

ABSTRACT
Influenza viruses of the H6 subtype have been isolated from wild and domestic aquatic and terrestrial avian species throughout the world since their first detection in a turkey in Massachusetts in 1965. Since 1997, H6 viruses with different neuraminidase (NA) subtypes have been detected frequently in the live poultry markets of southern China. Although sequence information has been gathered over the last few years, the H6 viruses have not been fully biologically characterized. To investigate the potential risk posed by H6 viruses to humans, here we assessed the receptor-binding preference, replication, and transmissibility in mammals of a series of H6 viruses isolated from live poultry markets in southern China from 2008 to 2011. Among the 257 H6 strains tested, 87 viruses recognized the human type receptor. Genome sequence analysis of 38 representative H6 viruses revealed 30 different genotypes, indicating that these viruses are actively circulating and reassorting in nature. Thirty-seven of 38 viruses tested in mice replicated efficiently in the lungs and some caused mild disease; none, however, were lethal. We also tested the direct contact transmission of 10 H6 viruses in guinea pigs and found that 5 viruses did not transmit to the contact animals, 3 viruses transmitted to one of the three contact animals, and 2 viruses transmitted to all three contact animals. Our study demonstrates that the H6 avian influenza viruses pose a clear threat to human health and emphasizes the need for continued surveillance and evaluation of the H6 influenza viruses circulating in nature.
IMPORTANCE Avian influenza viruses continue to present a challenge to human health. Research and pandemic preparedness have largely focused on the H5 and H7 subtype influenza viruses in recent years. Influenza viruses of the H6 subtype have been isolated from wild and domestic aquatic and terrestrial avian species throughout the world since their first detection in the United States in 1965. Since 1997, H6 viruses have been detected frequently in the live poultry markets of southern China; however, the biological characterization of these viruses is very limited. Here, we assessed the receptor-binding preference, replication, and transmissibility in mammals of a series of H6 viruses isolated from live poultry markets in southern China and found that 34% of the viruses are able to bind human type receptors and that some of them are able to transmit efficiently to contact animals. Our study demonstrates that the H6 viruses pose a clear threat to human health.
INTRODUCTION
Avian influenza viruses continue to present a challenge to human health. In the last century, human influenza pandemics occurred and resulted in significant mortality and morbidity; genetic evidence indicates that these pandemic strains were partially or entirely derived from avian influenza viruses (1–3). The H5N1 highly pathogenic influenza viruses have spread to over 60 countries and caused devastating avian influenza outbreaks. They have also transmitted to humans, resulting in 380 fatal cases among the 641 documented infections (4). The H7N7 avian influenza virus that caused highly pathogenic avian influenza in Holland in 2003 was associated with conjunctivitis in 347 humans (5). Infection with H7N7 influenza virus was confirmed in 87 of these cases; one person, a veterinarian, died as a result of the infection. H9N2 subtype avian influenza viruses have widely circulated in the world since their first detection in turkeys in Wisconsin in 1966 (6); they have also been detected in humans in China (7). In 2013, the newly emerged H7N9 viruses caused the deaths of 45 of 136 infected humans in mainland China as of 25 October (8), although these viruses did not cause any disease in chickens or ducks (9). Thus, avian influenza viruses currently circulating in poultry represent a major public threat.
H6 subtype influenza viruses are also widely distributed worldwide. An H6 influenza virus was first isolated from a turkey in Massachusetts in 1965, and since then H6 viruses have been isolated with increasing frequency from wild and domestic aquatic and terrestrial avian species (10–24). A surveillance study from Europe and the Americas found that H6 was the most abundantly detected influenza virus subtype and that it had a broader host range than other subtypes (25). Although the H6 viruses isolated to date have largely caused asymptomatic infections in waterfowl, infection of chickens with H6 viruses has been associated with decreased egg production, upper respiratory tract infection, morbidity, and increased mortality (10–13). Several studies have shown that certain H6 viruses can infect and cause illness in mice and ferrets (26, 27). An H6N6 virus was also detected in pigs in Guangdong province in China in 2010 (28), and an H6N1 virus was isolated from a human with flu-like symptoms in Taiwan in 2013 (29).
Southern China has been considered an epicenter of influenza virus due to its poultry breeding and trading style. Domestic poultry farming in southern China occurs in high-density settings but in a free-range manner. There are no biosafety measures in place on these farms, especially in those that breed yellow meat chickens and ducks. This creates an environment where migratory birds and domestic ducks are in close contact, sharing water, food, and habitat. The poultry in southern China is mainly traded through live poultry markets, where birds from different sources can be hosted together for several days. As a result, avian influenza viruses from different sources coexist in the open duck farms and live poultry markets, facilitating interspecies transmission and viral gene reassortment. H5N1 influenza viruses that have been detected in ducks in this region are complicated reassortants that have progressively acquired the ability to infect and kill mammals (30, 31). Surveillance studies have revealed the existence of different subtypes of H6 viruses in the live poultry markets in southern China (32–36), which is where the H5N1 influenza viruses and H7N9 viruses jumped to humans. Although a vast amount of sequence analysis of the H6 viruses has been performed (25, 32–39), there has been relatively little biologic characterization of the H6 viruses from China. Here, we fully characterized the genetic variation, receptor-binding specificity, replication capability, and transmission in mammals of a series of H6 influenza viruses that were detected in the live poultry markets in southern China between 2008 and 2011. Our results reveal the potential threat to public health posed by H6 influenza viruses.
MATERIALS AND METHODS
Ethics statements and facility.
The present study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the Ministry of Science and Technology of the People's Republic of China. The protocol was approved by the Committee on the Ethics of Animal Experiments of the Harbin Veterinary Research Institute (HVRI) of the Chinese Academy of Agricultural Sciences (CAAS) (approval numbers BRDW-XBS–09S for mice and BRDW-TS–09S for guinea pigs).
Virus isolation and identification.
The H6 viruses used in this study were isolated from live poultry markets between 2008 and 2011 in China during routine surveillance. Viruses were isolated in embryonated chicken eggs. Hemagglutinin (HA) subtypes of influenza viruses were identified by using the hemagglutination inhibition (HI) test. Neuraminidase (NA) subtypes were determined by means of direct sequencing. The viruses selected for sequence analysis and animal study were biologically cloned by three rounds of limiting dilution in embryonated specific-pathogen-free eggs.
Receptor-binding analysis by means of hemagglutination assays.
Hemagglutination (HA) assays using resialylated chicken red blood cells (cRBCs) were performed as described previously (40) with minor modifications. cRBCs were enzymatically desialylated with Vibrio cholerae neuraminidase (Sigma-Aldrich, St. Louis, MO, USA) and then resialylated by using α-2-6-N-sialyltransferase (Sigma-Aldrich, St. Louis, MO, USA) and CMP-sialic acid (Sigma-Aldrich, St. Louis, MO, USA). The minimum cutoff value for the HA assay was 2.
Receptor-binding analysis using a solid-phase direct-binding assay.
Receptor specificity was analyzed by use of a solid-phase direct-binding assay as described previously (41) using two different glycopolymers: α-2,3-sialylglycopolymer [Neu5Acα2-3Galβ1-4GlcNAcβ1-pAP (para-aminophenyl)-alpha-polyglutamic acid (α-PGA)] and the α-2,6-sialylglycopolymer [Neu5Acα2-6Galβ1-4GlcNAcβ1-pAP (para-aminophenyl)-alpha-polyglutamic acid (α-PGA)]. Chicken antiserum against Chicken/Guangdong/S1312/2010 (H6N2) [CK/GD/S1312/10 (H6N2)] virus was used as the primary antibody and a horseradish peroxidase (HRP)-conjugated goat-anti-chicken antibody (Sigma-Aldrich, St. Louis, MO, USA) was used as the secondary antibody. Absorbance was determined at a wavelength of 490 nm. The cutoff value for the glycan binding assays was the background value of the well with 100 ng of glycopolymer in the absence of added virus.
Genetic and phylogenetic analyses.
Viral RNA was extracted with an RNeasy minikit (Qiagen, Valencia, CA) and was reverse transcribed. PCR amplification was performed by using segment-specific primers (primer sequences are available upon request). The PCR products were purified with a QIAquick PCR purification kit (Qiagen) and sequenced on an Applied Biosystems DNA analyzer. The nucleotide sequences were edited using the Seqman module of the DNAStar package, and phylogenetic analyses were carried out with the PHYLIP program of the ClustalX software package (version 1.81) using a neighbor-joining algorithm. Bootstrap values of 1,000 were used.
Antigenic analyses.
Antigenic analyses were performed by using HI tests with guinea pig antisera generated against the indicated viruses. To generate the antisera, we intranasally (i.n.) inoculated Hartley strain female guinea pigs weighing 300 to 350 g with 106 50% egg infective doses (EID50) of test viruses indicated in Table 4 and collected sera 3 weeks after the injection. Sera were treated with Vibrio cholerae receptor-destroying enzyme (Denka-Seiken) before being tested for the presence of HI antibody with 0.5% (vol/vol) chicken erythrocytes. The minimum cutoff value for the HI assay was 10.
TABLE 4.
Antigenic analysis of H6 avian influenza viruses isolated in China
| Virus (HA group) | HI antibody titer of antiserum against virus (HA group)a: |
||||
|---|---|---|---|---|---|
| CK/GD/S1312/10 (H6N2) (1) | DK/HuB/S1114/09 (H6N2) (2) | DK/ZJ/S4204/10 (H6N6) (3) | DK/GD/S4192/08 (H6N2) (4) | DK/GD/S1419/11 (H6N6) (5) | |
| CK/GD/S1312/10 (H6N2) (1) | 640 | 80 | 40 | <10 | <10 |
| DK/GD/S1328/10 (H6N2) (1) | 640 | 80 | 80 | <10 | <10 |
| GS/GD/S1384/10 (H6N2) (1) | 640 | 80 | 80 | <10 | <10 |
| DK/GD/S1289/10 (H6N2) (1) | 320 | 40 | 80 | <10 | <10 |
| DK/HuN/S3047/09 (H6N2) (1) | 320 | 40 | 80 | <10 | <10 |
| CK/GD/S1453/10 (H6N2) (1) | 640 | 80 | 80 | <10 | <10 |
| DK/HuB/S4135/10 (H6N2) (1) | 320 | 40 | 80 | <10 | <10 |
| CK/GD/S1414/10 (H6N6) (1) | 640 | 80 | 80 | <10 | <10 |
| DK/ZJ/S1023/10 (H6N6) (1) | 320 | 80 | 160 | <10 | <10 |
| DK/GD/S3180/10 (H6N6) (1) | 640 | 80 | 80 | <10 | <10 |
| DK/HuN/S4273/10 (H6N6) (1) | 320 | 80 | 80 | <10 | <10 |
| CK/GD/S1311/10 (H6N6) (1) | 640 | 80 | 40 | <10 | <10 |
| CK/HuN/S4495/10 (H6N6) (1) | 320 | 40 | 80 | <10 | <10 |
| CK/HuN/S3003/09 (H6N6) (1) | 320 | 40 | 80 | <10 | <10 |
| DK/HuB/S1114/09 (H6N2) (2) | 320 | 160 | 40 | <10 | <10 |
| DK/HuB/S4170/08 (H6N2) (2) | 160 | 160 | 40 | <10 | <10 |
| DK/HuN/S1284/09 (H6N2) (2) | 320 | 160 | 160 | <10 | <10 |
| DK/GD/S4018/10 (H6N6) (2) | 160 | 80 | 160 | <10 | <10 |
| DK/GD/S1663/09 (H6N6) (2) | 80 | 80 | 160 | <10 | <10 |
| CK/GX/S4029/10 (H6N6) (2) | 160 | 80 | 80 | <10 | <10 |
| DK/ZJ/S4204/10 (H6N6) (3) | 80 | <10 | 320 | <10 | <10 |
| CK/HuN/S4191/09 (H6N2) (3) | 80 | <10 | 320 | <10 | <10 |
| DK/HuN/S4386/09 (H6N2) (3) | 80 | <10 | 320 | <10 | <10 |
| DK/HuB/S1366/09 (H6N2) (3) | 80 | <10 | 320 | <10 | <10 |
| CK/GX/S4381/10 (H6N6) (3) | 40 | <10 | 320 | <10 | <10 |
| DK/GX/S4111/10 (H6N6) (3) | 80 | <10 | 320 | <10 | <10 |
| DK/GD/S4192/08 (H6N2) (4) | 80 | <10 | 160 | 160 | <10 |
| DK/GD/S1566/09 (H6N2) (4) | 40 | <10 | 160 | 80 | <10 |
| DK/GD/S1419/11 (H6N6) (5) | 40 | 20 | 20 | <10 | 1280 |
| CK/GD/S2346/09 (H6N2) (5) | 40 | 40 | 80 | <10 | 160 |
| DK/FJ/S4081/08 (H6N2) (5) | 40 | 40 | 40 | <10 | 160 |
| DK/GD/S4251/10 (H6N6) (5) | 20 | 20 | 20 | <10 | 640 |
| GS/GD/1/96 (H5N1)b | <10 | <10 | <10 | <10 | <10 |
| Newcastle disease virus (LaSota strain)b | <10 | <10 | <10 | <10 | <10 |
Antiserum was generated by intranasally inoculating guinea pigs with 106 EID50 of the indicated virus. Homologous titers are underlined.
An H5N1 virus and a Newcastle disease virus served as negative antigen controls.
Studies with mice.
Groups of eight 6-week-old female BALB/c mice (Beijing Experimental Animal Center, Beijing, China) were lightly anesthetized with CO2 and inoculated i.n. with 106 EID50 of H6 influenza virus in a volume of 50 μl of medium. Three of the eight mice were euthanized on day 3 postinoculation (p.i.) for titration of virus from the lungs, nasal turbinate, kidneys, spleen, and brain. The remaining five mice were monitored daily for weight loss and mortality for a total of 2 weeks.
Studies with guinea pigs.
Hartley strain female guinea pigs weighing 300 to 350 g (Vital River Laboratories, Beijing, China) serologically negative for influenza viruses were used in these studies. Ketamine (20 mg/kg) and xylazine (1 mg/kg) were used to anesthetize the animals via intramuscular injection.
To investigate the replication of H6 influenza viruses, we anesthetized groups of two guinea pigs and inoculated them i.n. with 106 EID50 of test virus in a 0.3-ml volume of medium (0.15 ml per nostril). The animals were euthanized on day 3 p.i., and nasal washes and lung tissues were collected for virus titration in eggs.
For the transmission studies, groups of three animals were inoculated i.n. with 106 EID50 of test virus and housed in a cage placed inside an isolator. Three naive animals were introduced into the same cage 24 h later. Nasal washes were collected at 2-day intervals, beginning on day 2 p.i. (1 day postcontact) and titrated in eggs. Sera were collected on day 21 p.i. and treated with Vibrio cholerae receptor-destroying enzyme (Denka-Seiken) before being tested for the presence of HI antibody with 0.5% (vol/vol) chicken erythrocytes. To prevent inadvertent physical transmission of virus by the investigators, the contact guinea pigs were always handled first, and gloves, implements, and napkins on the work surface were changed between animals. The ambient conditions for these studies were set as 20 to 22°C and 30% to 40% relative humidity. The airflow in the isolator was horizontal with a speed of 0.1 m/s.
Viral replication in human lung cells (A549).
Virus was inoculated into A549 monolayers at a multiplicity of infection (MOI) of 0.01. The cells were incubated at 37°C in Opti-MEM (Gibco, Grand Island, NY, USA) containing 0.2 μg/ml l-(tosylamido-2-phenyl) ethyl chloromethyl ketone (TPCK)-treated trypsin (Sigma-Aldrich, St. Louis, MO, USA). Virus-containing culture supernatant was collected at various time points (hours postinfection) and titrated in embryonated chicken eggs. The growth data shown in Fig. 4 are averages of the results of three independent experiments.
FIG 4.
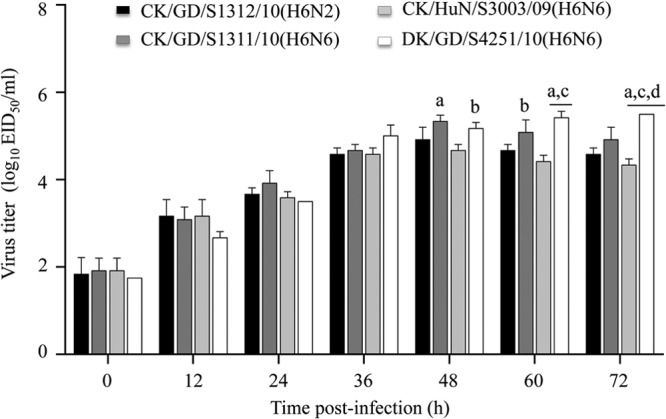
Multicycle replication of H6 avian influenza viruses in A549 cells. A549 monolayers were inoculated at an MOI of 0.01 with virus, and the culture supernatants were collected at the indicated time points and then titrated in eggs. a, P < 0.01 compared with titers in CK/HuN/S3003/09 (H6N6)-infected cells; b, P < 0.05 compared with titers in CK/HuN/S3003/09 (H6N6)-infected cells; c, P < 0.01 compared with titers in CK/GD/S1312/10 (H6N2)-infected cells; d, P < 0.05 compared with titers in CK/GD/S1311/10 (H6N6)-infected cells.
Statistical analysis.
The affinities of the viruses to different glycans were compared by use of two-way analysis of variance. The replication of different viruses in A549 cells was compared by use of the Student-Newman-Keuls test. Transmission efficiencies of different viruses were compared by use of Fisher's exact test.
Nucleotide sequence accession numbers.
The nucleotide sequences of the 38 H6 viruses determined in this study have been deposited in the Global Initiative on Sharing All Influenza Data (GISAID) database under numbers EPI500847 to EPI501150.
RESULTS
Isolation of influenza viruses from live poultry markets in southern China.
We collected 29,305 samples from poultry markets and poultry slaughterhouses in Shanghai, Anhui, Zhejiang, Jiangsu, Fujian, Hubei, Hunan, Henan, Jiangxi, Shandong, Guangxi, and Guangdong provinces from the winter of 2008 to the spring of 2011. All samples were inoculated individually into 10-day-old embryonated chicken eggs for virus isolation. The HA subtype was confirmed by use of an HI test with antisera against the 16 HA subtypes of avian influenza viruses. Nine different HA subtypes of avian influenza viruses were detected, including H1 (9 strains), H3 (134 strains), H4 (302 strains), H5 (285 strains), H6 (1,145 strains), H7 (15 strains), H9 (1,881 strains), H10 (44 strains), and H11 (31 strains). The 1,145 H6 subtype viruses were isolated from the samples that were collected from Guangdong, Guangxi, Hunan, Hubei, Fujian, and Zhejiang provinces.
Receptor-binding specificity of H6 avian influenza viruses.
Binding to α-2,6-linked sialic acids (Sias) (human type receptors) is a prerequisite for an avian influenza virus to transmit from human to human (42–45). From the 1,145 H6 isolates, we selected 1 to 3 viruses that were isolated from the birds of each owner in the live poultry markets (a total of 257 H6 subtype viruses were thus selected) and tested their affinities (approximately 22% of the total viruses isolated) for α-2,6-linked Sias by using hemagglutination (HA) assays with cRBCs resialylated with α-2,6-glycans as described previously (40). We found that 39 of 187 (20.9%) H6N2 viruses and 48 of 70 (68.6%) H6N6 viruses agglutinated the cRBCs resialylated with α-2,6-glycans (Table 1).
TABLE 1.
H6 influenza viruses that bind to human-like receptorsa
| Location isolated | H6N2 |
H6N6 |
||
|---|---|---|---|---|
| No. of strains tested | No. of strains binding to α-2,6-linked sialic acid | No. of strains tested | No. of strains binding to α-2,6-linked sialic acid | |
| Guangdong | 110 | 25 | 27 | 23 |
| Guangxi | 14 | 1 | 11 | 10 |
| Fujian | 14 | 1 | 12 | 0 |
| Hunan | 33 | 6 | 9 | 6 |
| Hubei | 12 | 6 | 7 | 6 |
| Zhejiang | 4 | 0 | 4 | 3 |
| Total | 187 | 39 | 70 | 48 |
Viruses were identified by using a hemagglutination assay with chicken red blood cells resialylated with α-2,6-glycans.
We further confirmed the binding of H6 subtype viruses to α-2,6-glycans by using a solid-phase binding assay as described previously by Imai et al. (41). Of the 87 H6 viruses that agglutinated the cRBCs resialylated with α-2,6-glycans, 9 were selected for this assay (Fig. 1A). Another virus, A/chicken/Hunan/S3003/2009 (H6N6) [CK/HuN/S3003/09 (H6N6)], that did not agglutinate the cRBCs resialylated with α-2,6-glycans was included as a control. Eight of the 10 viruses bound to both α-2,3-glycans and α-2,6-glycans, although their affinity for the α-2,3-glycans was higher than that for the α-2,6-glycans. One virus, A/duck/Guangdong/S4251/2010 (H6N6) [DK/GD/S4251/10 (H6N6)], bound to the α-2,6-glycans with high affinity and to the α-2,3-glycans with very low affinity, whereas another virus, CK/HuN/S3003/09 (H6N6), bound to the α-2,3-glycans with high affinity but to the α-2,6-glycans with very low affinity (Fig. 1B). The results of the two assays were in agreement, confirming that about 34% of the H6 influenza viruses circulating in the live poultry markets of southern China have acquired the ability to recognize the human type receptors.
FIG 1.

Characterization of the receptor-binding properties of H6 influenza viruses. (A) Hemagglutination titers of H6 influenza viruses with 0.5% cRBCs treated as follows: cBRCs, untreated; desialylated (Desial) cRBCs, treated with Vibrio cholerae neuraminidase; α-2,6 cRBCs, treated with VCNA and resialylated with α-2,6-glycans. The dashed line indicates the lower limit of detection. (B) Glycan-binding specificity of H6 viruses. The binding of the viruses to two different glycans (α-2,3-glycans, blue; α-2,6-glycans, red) was tested. The data shown are the means of three repeats; the error bars indicate standard deviations. Significant differences were detected between the affinities for the two glycans.
Molecular and phylogenetic analysis.
We demonstrated that of the 257 H6 isolates tested, 87 viruses were able to bind to the α-2,6-glycans in the hemagglutination assay with resialylated α-2,6 cRBCs, including 39 H6N2 viruses and 48 H6N6 viruses. To understand their genetic relationship, we sequenced 37 representative viruses (17 H6N2 viruses and 20 H6N6 viruses) that bound to the α-2,6-glycans and came from the birds of different owners. One virus [CK/HuN/S3003/09 (H6N6)], which bound only to the α-2,3-glycans and was used as a control in some experiments, was also sequenced. The HA gene of all of these viruses had only one basic amino acid, arginine, in the cleavage site of HA1 and HA2. The amino acid changes of A138S (H3 numbering, which is used throughout the manuscript), Q226L, and G228S, as well as the absence of glycosylation at positions 158 to 160 in HA, have been reported to favor the affinity of influenza viruses for human type receptors (40, 41, 46, 47). A138S was present in five H6 influenza viruses tested in this study and was also detected in the H6N6 virus previously isolated from a pig (Table 2). Q226L and G228S were not detected in any of the 38 avian viruses we sequenced here; however, G228S was detected in both swine and human H6 isolates (Table 2). There are five potential glycosylation sites in HA1, at positions 21 to 23, 33 to 35, 169 to 171, 291 to 293, and 296 to 298. Four of these sites were highly conserved among the 38 viruses, whereas the potential glycosylation site at position 169 to 171 was absent from 10 of the 38 avian viruses and from the swine H6N6 influenza virus (Table 2).
TABLE 2.
Molecular characteristics of H6 avian influenza viruses
| Virusa | Amino acid(s) at indicated key position(s) in HAb |
Amino acid deletion in NA (position) | Amino acid at position 31 of M2 | Amino acid deletion at position 218–230 in NS1 protein | ||
|---|---|---|---|---|---|---|
| 138 | 228 | 169–171 | ||||
| TW/2/13 (H6N1) | A | S | NNT | NA | N | No |
| SW/GD/K6/10 (H6N6) | S | S | RNT | Yes (59–69) | S | Yes |
| CK/GD/S1312/10 (H6N2) | A | G | NNT | Yes (63–65) | N | Yes |
| DK/HuB/S4135/10 (H6N2) | A | G | NNT | Yes (63–65) | S | Yes |
| DK/HuB/S4170/08 (H6N2) | A | G | NNT | No | S | No |
| DK/HuN/S1284/09 (H6N2) | A | G | NNT | Yes (67–76) | S | Yes |
| CK/HuN/S4191/09 (H6N2) | A | G | TNT | No | S | Yes |
| DK/HuN/S4386/09 (H6N2) | A | G | RNT | No | S | Yes |
| DK/HuB/S1366/09 (H6N2) | S | G | TNT | No | S | Yes |
| CK/GD/S2346/09 (H6N2) | S | G | RNT | No | S | Yes |
| DK/FJ/S4081/08 (H6N2) | S | G | RNT | No | S | No |
| CK/GD/S1311/10 (H6N6) | A | G | NNT | No | N | Yes |
| CK/HuN/S4495/10 (H6N6) | A | G | NNT | No | S | No |
| CK/GX/S4381/10 (H6N6) | A | G | RNT | No | S | Yes |
| DK/GX/S4111/10 (H6N6) | A | G | RNT | No | S | Yes |
| DK/ZJ/S4204/10 (H6N6) | A | G | RNT | No | S | Yes |
| DK/GD/S4251/10 (H6N6) | S | G | RNT | Yes (59–69) | S | Yes |
| DK/GD/S1419/11 (H6N6) | S | G | RNT | Yes (59–69) | S | Yes |
| Other 22 virusesc | A | G | NNT | No | S | Yes |
The HA genes of H6 viruses showed great diversity and formed phylogenetically different groups. The 38 isolates we sequenced in this study could be divided into five groups (Fig. 2A). The intragroup virus homology was over 98%, while the intergroup homology was less than 95%. There were 20, 6, 6, 2, and 4 viruses in groups 1, 2, 3, 4, and 5, respectively. Group 1 contained strains from Hunan, Hubei, Guangdong, and Zhejiang provinces, and one H6N1 virus isolated from Guangxi by others (48) also clustered in this group. The group 2 viruses came from Guangdong, Guangxi, Hunan, and Hubei provinces. The group 3 viruses came from Hunan, Hubei, Zhejiang, and Guangxi provinces, and the H6N6 virus in this group had also been detected previously in Hubei province (49). The two viruses in group 4 were isolated from Guangdong province, while the four viruses in group 5 were from Guangdong and Fujian provinces. Of note, the H6N6 virus in group 5 had also been isolated from pigs in Guangdong province in 2010 (Fig. 2A).
FIG 2.

Phylogenetic analyses of the H6 viruses isolated from 2008 to 2011 in China. The phylogenetic trees were generated with the PHYLIP program of the ClustalX software package (version 1.81). The nine trees were generated based on the following sequences: HA nucleotides (nt) 18 to 1718, N2 nt 20 to 1429, N6 nt 20 to 1432, PB2 nt 28 to 2307, PB1 nt 25 to 2298, PA nt 25 to 2175, NP nt 46 to 1542, M nt 26 to 1007, and NS nt 27 to 864. The phylogenetic trees of HA (A), N2 NA (B), and N6 NA (C) were rooted to A/Turkey/Canada/1963 (H6N2), A/Turkey/England/1969 (H3N2), and A/Duck/England/1/1956 (H11N6), respectively. The phylogenetic trees of PB2 (D), PB1 (E), PA (F), NP (G), M (H), and NS (I) were rooted to A/Equine/Prague/1/56 (H7N7). Sequences of viruses with names in black were downloaded from available databases; viruses with names in other colors were sequenced in this study. The colors of the virus names in the NA, PB2, PB1, PA, NP, M, and NS trees match with those used in the HA tree. Abbreviations: AN, avian; BWT, blue-winged teal; CK, chicken; DK, duck; EC, Eastern China; EW, Eurasian wigeon; FJ, Fujian; GD, Guangdong; GS, goose; GX, Guangxi; HK, Hongkong; HuB, Hubei; HuN, Hunan; MDK, Muscovy duck; ML, mallard; SW, swine; TL, teal; WDK, wild duck; ZJ, Zhejiang.
The 17 N2 neuraminidase (NA) genes also formed 5 groups, and there were 8, 5, 1, 1, and 2 viruses in groups 1, 2, 3, 4, and 5, respectively (Fig. 2B). The NA gene of A/duck/Hunan/S1248/2009 (H6N2) [DK/HuN/S1284/09 (H6N2)] has a 30-nucleotide deletion at positions 197 to 226 that results in the loss of 10 amino acids at position 67 to 76. The NA gene of two viruses in group 5 has a nine-nucleotide deletion at position 187 to 195 that results in the loss of three amino acids at position 63 to 65. The 21 N6 NA genes formed 3 groups, with 16, 2, and 3 viruses in groups 1, 2, and 3, respectively (Fig. 2C). The NA of the two viruses in group 2 has a 33-nucleotide deletion at position 175 to 207 that results in the loss of 11 amino acids at position 59 to 69 (Table 2).
Several amino acid changes in PB2, including T271A, E627K, and D701N, contribute to the increased virulence and transmission of influenza viruses in mammals (40, 45, 52, 53). The H6 viruses we sequenced in this study had none of these mutations. The PB2 gene showed distinct diversity and formed six groups in the phylogenetic tree (Fig. 2D; see also Fig. S1A in the supplemental material). The PB2 in groups 1, 2, and 5 was shared by both the H6N2 and H6N6 viruses, whereas the PB2 in groups 4 and 6 was detected only in H6N2 viruses and that in group 3 was detected only in H6N6 viruses (Table 3).
TABLE 3.
Genotype and manifestation in mice of H6 influenza viruses
| Virus | Group of each gene segment in the phylogenetic tree |
Genotype | Virus titers in organs in mice (log10 EID50)a |
Maximum body wt change (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HA | NA | PB2 | PB1 | PA | NP | M | NS | Nasal turbinate | Lung | |||
| DK/GD/S1328/10 (H6N2) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | A1 | 0.8 ± 0.4 | 5.8 ± 0.4 | +8.4 |
| GS/GD/S1384/10 (H6N2) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | A1 | 1.8 ± 0.5 | 5.3 ± 0.3 | +10.8 |
| DK/GD/S1289/10 (H6N2) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | A2 | 0.8 ± 0.6 | 5.5 ± 0.3 | +10.0 |
| DK/HuN/S3047/09 (H6N2) | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | A3 | ND | 1.8 ± 0.4 | +11.1 |
| CK/GD/S1453/10 (H6N2) | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 2 | A4 | ND | 5.3 ± 0.3 | +9.6 |
| CK/GD/S1312/10 (H6N2) | 1 | 5 | 1 | 5 | 1 | 1 | 5 | 5 | A5 | 2.3 ± 0.1 | 4.4 ± 0.1 | −6.1 |
| DK/HuB/S4135/10 (H6N2) | 1 | 5 | 6 | 5 | 4 | 5 | 5 | 5 | A6 | 0.7 ± 0.3 | 2.2 ± 0.4 | +8.2 |
| DK/HuB/S4170/08 (H6N2) | 2 | 2 | 5 | 4 | 5 | 2 | 4 | 4 | A7 | ND | 2.9 ± 1 | +13.7 |
| DK/HuB/S1114/09 (H6N2) | 2 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | A8 | 1.4 ± 0.3 | 5.5 ± 0.3 | +8.2 |
| DK/HuN/S1284/09 (H6N2) | 2 | 4 | 1 | 2 | 2 | 1 | 1 | 3 | A9 | 0.7 ± 0.3 | 5.4 ± 0.1 | −1.4 |
| CK/HuN/S4191/09 (H6N2) | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | A10 | 0.7 ± 0.3 | 1.5 | +11.1 |
| DK/HuN/S4386/09 (H6N2) | 3 | 1 | 4 | 1 | 1 | 1 | 1 | 2 | A11 | 0.7 ± 0.3 | 5.4 ± 0.1 | +12.8 |
| DK/HuB/S1366/09 (H6N2) | 3 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | A12 | ND | 5.3 ± 0.4 | +9.7 |
| DK/GD/S4192/08 (H6N2) | 4 | 1 | 1 | 1 | 3 | 1 | 1 | 2 | A13 | 1.3 ± 0.3 | 5.3 ± 0.4 | +7.9 |
| DK/GD/S1566/09 (H6N2) | 4 | 2 | 1 | 1 | 3 | 1 | 1 | 2 | A14 | ND | ND | +10.2 |
| CK/GD/S2346/09 (H6N2) | 5 | 2 | 5 | 4 | 5 | 2 | 4 | 4 | A15 | 1.5 | 5.7 ± 0.1 | +9.2 |
| DK/FJ/S4081/08 (H6N2) | 5 | 2 | 5 | 4 | 5 | 2 | 4 | 4 | A15 | 1.8 ± 0.1 | 5.8 ± 0.6 | +9.3 |
| CK/GD/S1414/10 (H6N6) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | B1 | 1.5 ± 0.3 | 4.8 ± 0.4 | +11.8 |
| DK/ZJ/S1023/10 (H6N6) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | B1 | ND | 5.8 ± 0.6 | +8.9 |
| DK/GD/S3073/10 (H6N6) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | B1 | 2 ± 0.7 | 4.1 ± 0.3 | −3.7 |
| GS/GD/S4362/09 (H6N6) | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | B1 | 1.2 ± 0.3 | 5.8 ± 0.5 | +9.3 |
| DK/GD/S1155/11 (H6N6) | 1 | 1 | 1 | 1 | 2 | 2 | 1 | 2 | B2 | 0.8 ± 0.4 | 4.9 ± 0.5 | +6.6 |
| DK/GD/S3180/10 (H6N6) | 1 | 1 | 1 | 1 | 3 | 1 | 1 | 1 | B3 | 0.7 ± 0.3 | 1.6 ± 0.1 | +13.4 |
| DK/HuN/S4273/10 (H6N6) | 1 | 1 | 1 | 3 | 2 | 1 | 1 | 2 | B4 | 2.3 ± 0.1 | 5.5 ± 0.4 | +8.2 |
| CK/GD/S1311/10 (H6N6) | 1 | 1 | 1 | 5 | 1 | 1 | 5 | 5 | B5 | 1.8 ± 0.4 | 5.3 ± 0.1 | −5.7 |
| DK/ZJ/S1134/11 (H6N6) | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | B6 | ND | 5.4 ± 0.4 | +6.1 |
| DK/GD/S3225/10 (H6N6) | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | B6 | 1.2 ± 0.3 | 3.4 ± 0.1 | −5.8 |
| DK/GD/S3468/10 (H6N6) | 1 | 1 | 2 | 2 | 1 | 1 | 1 | 1 | B7 | 0.9 ± 0.4 | 5.6 ± 0.1 | +13.8 |
| CK/HuN/S4495/10 (H6N6) | 1 | 1 | 5 | 3 | 2 | 3 | 1 | 4 | B8 | 2.4 ± 0.9 | 4.4 ± 0.1 | +11.6 |
| CK/HuN/S3003/09 (H6N6) | 1 | 3 | 1 | 1 | 2 | 1 | 3 | 1 | B9 | 0.7 ± 0.3 | 4.0 | +8.2 |
| DK/GD/S4018/10 (H6N6) | 2 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | B10 | 0.6 ± 0.1 | 4.8 ± 0.6 | +10.5 |
| DK/GD/S1663/09 (H6N6) | 2 | 3 | 1 | 1 | 1 | 1 | 1 | 1 | B11 | 1.2 ± 0.6 | 4.3 ± 0.7 | +9.5 |
| CK/GX/S4029/10 (H6N6) | 2 | 3 | 2 | 2 | 1 | 1 | 1 | 1 | B12 | 1.2 ± 0.3 | 2 ± 0.5 | +11.0 |
| CK/GX/S4381/10 (H6N6) | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | B13 | 0.6 ± 0.1 | 1.6 ± 0.1 | +7.4 |
| DK/GX/S4111/10 (H6N6) | 3 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | B13 | 0.7 ± 0.3 | 3.8 ± 0.4 | +9.2 |
| DK/ZJ/S4204/10 (H6N6) | 3 | 1 | 1 | 2 | 2 | 1 | 1 | 1 | B14 | ND | 5.3 ± 0.4 | +9.1 |
| DK/GD/S4251/10 (H6N6) | 5 | 2 | 3 | 2 | 1 | 4 | 2 | 3 | B15 | 3.8 ± 0.4 | 5.3 ± 0.1 | −2.9 |
| DK/GD/S1419/11 (H6N6) | 5 | 2 | 3 | 2 | 1 | 4 | 2 | 3 | B15 | 2.4 ± 0.1 | 5.9 ± 0.5 | −2.5 |
Six-week-old BALB/c mice were inoculated intranasally with 106 EID50 of each virus in a 50-μl volume of medium. Three mice from each group were killed on day 3 postinoculation, and virus titers were determined in samples of nasal turbinate, lung, spleen, kidney, and brain in eggs. Virus was not detected from the spleen, kidney, or brain of any mouse; therefore, these data are not shown in the table. Data are presented as means ± standard deviations. ND, virus not detected in sample.
The PB1, PA, NP, M, and NS genes of the H6 viruses also showed distinct diversity, and each formed five groups in the phylogenetic trees (Fig. 2E to I; see also Fig. S1B to E in the supplemental material). The PB1 in groups 1, 2, and 5 was shared by both the H6N2 and H6N6 viruses, the PB1 in group 4 was detected only in H6N2 viruses, and the PB1 in group 3 was detected only in H6N6 viruses. The PA in groups 1, 2, and 3 was detected in both H6N2 and H6N6 viruses, whereas the PA in groups 4 and 5 was detected only in H6N2 viruses. The NP in groups 1 and 2 was shared by both H6N2 and H6N6 viruses, whereas the NP in groups 3 and 4 was detected only in H6N6 viruses and the NP in group 5 was detected only in an H6N2 virus. The M in groups 1 and 5 was shared by both H6N2 and H6N6 viruses, whereas the M in groups 2 and 3 was detected only in H6N6 viruses and the M in group 4 was detected only in H6N2 viruses. The S31N mutation was detected in the M2 of the A/chicken/Guangdong/S1312/2010 (H6N2) [CK/GD/S1312/10 (H6N2)] and A/chicken/Guangdong/S1311/2010 (H6N6) [CK/GD/S1311/10 (H6N6)] viruses (Table 3), implying that these two viruses may be resistant to amantadine and rimantadine. The NS genes in all 5 groups were shared by both H6N2 and H6N6 viruses (Table 3). The NS1 of three viruses, A/duck/Hubei/S4170/2008 (H6N2) [DK/HuB/S4170/08 (H6N2)], A/duck/Fujian/S4081/2008 (H6N2) [DK/FJ/S4081/08 (H6N2)], and A/chicken/Hunan/S4495/2010 (H6N6) [CK/HuN/S4495/10 (H6N6)], encoded 230 amino acids, whereas the NS1 of the other 35 viruses encoded only 217 amino acids (Table 2), with a deletion of amino acids 218 to 230, which was previously reported to be associated with the attenuated phenotype of influenza viruses in mice (54, 55).
On the basis of this genomic diversity, we divided the viruses examined in this study into 30 genotypes, including 15 H6N2 virus genotypes, A1 to A15, and 15 H6N6 virus genotypes, B1 to B15 (Table 3). It is interesting to note that the CK/GD/S1311/10 (H6N6) and CK/GD/S1312/10 (H6N2) viruses differ only in their NA gene; they have identical PB2, PB1, PA, HA, NP, M, and NS genes.
Antigenic analysis.
Antisera, generated in guinea pigs, to selected H6 viruses from each HA gene group were used for antigenic analyses by hemagglutination inhibition (HI) assays with 0.5% cRBCs. As indicated in Table 4, the antisera against CK/GD/S1312/10 (H6N2) (HA group 1) and DK/ZJ/S4204/10 (H6N6) (HA group 3) cross-reacted well with all of the viruses tested, although the HI titers of these viruses were 2- to 32-fold lower than the homologous titers. The antisera against DK/HuB/S1114/09 (H6N2) (HA group 2) cross-reacted well with viruses bearing the HA gene from groups 1, 2, and 5 but did not react with viruses bearing the HA gene from groups 3 and 4 (Table 4). The antisera against A/duck/Guangdong/S4192/2008 (H6N2) [DK/GD/S4192/08 (H6N2)] (HA group 4) and A/duck/Guangdong/S1410/2011 (H6N6) [DK/GD/S1419/11 (H6N6)] (HA group 5) cross-reacted only with viruses bearing the HA gene from group 4 and group 5, respectively (Table 4). These results indicate that the H6 viruses circulating in nature exhibit severe antigenic drift.
Replication of H6 viruses in mice.
To investigate the replication and virulence of H6 influenza viruses in mammals, we inoculated groups of eight 6-week-old BALB/c mice with 106 EID50 of each virus. Three mice in each group were killed on day 3 p.i. for virus detection in organs, including nasal turbinate, lung, spleen, kidney, and brain; the other five mice in each group were observed for a total of 14 days for body weight changes and death.
One virus, A/duck/Guangdong/S1566/2009 (H6N2) [DK/GD/S1566/09 (H6N2)], was not detected in any organs tested (Table 3). The other 37 viruses were all detected in the lungs of mice, with mean titers ranging from 1.5 to 5.9 log10 EID50; however, only 30 of these 37 viruses were also detected in the nasal turbinate of mice, with titers ranging from 0.6 to 3.8 log10 EID50 (Table 3). None of the viruses were detected in the spleen, kidneys, or brain of mice. Seven of the viruses caused mice to experience body weight loss (range, 1.4% to 6.1%), whereas mice infected with the other 31 viruses gained body weight (Table 3). These results indicate that most H6 influenza viruses can replicate in the respiratory system of mice without preadaptation but that only a few of them cause mild disease in mice.
Transmission of H6 influenza viruses in guinea pigs.
The above results indicated that the H6 avian influenza viruses can efficiently infect and replicate in mice and that some of the viruses have acquired the ability to bind to the human-like receptors. We, therefore, investigated whether these viruses could also transmit among mammals by testing the 9 viruses that bound to the α-2,6-glycan (shown in Fig. 1) and replicated efficiently in both the nasal turbinate and lung of mice in guinea pigs, and we also included one virus, CK/HuN/S3003/09 (H6N6), that bound only to the α-2,3-glycan in this test. To evaluate virus replication and tissue tropism in inoculated guinea pigs, we intranasally (i.n.) inoculated two guinea pigs with 106 EID50 of each virus. The nasal washes and lung tissues from each guinea pig were collected on day 3 p.i. for viral titration. All viruses were detected in the nasal washes and lungs of these guinea pigs except for the CK/HuN/S3003/09 (H6N6) virus, which was detected only in the nasal washes. There was considerable variation in the titers among the strains (Table 5).
TABLE 5.
Replication and transmission of H6 avian influenza viruses in guinea pigs
| Virus | Viral titersa (log10 EID50) in: |
No. positive for seroconversion/total no. (HI titers) |
Direct contact transmission | ||
|---|---|---|---|---|---|
| Nasal turbinate | Lung | Inoculated | Contact | ||
| CK/GD/S1312/10 (H6N2) | 3.8, 4.5 | 1.0, 1.0 | 3/3 (160–320) | 3/3 (80–320) | Highly efficient |
| DK/GD/S4192/08 (H6N2) | 3.5, 5.0 | 1.5, 3.5 | 3/3 (160) | 0/3 | None |
| DK/HuB/S1114/09 (H6N2) | 5.0, 5.0 | 1.3, 1.0 | 3/3 (10–80) | 1/3 (10) | Inefficient |
| GS/GD/S1384/10 (H6N2) | 2.5, 3.0 | 2.3, 2.3 | 3/3 (10–40) | 0/3 | None |
| CK/GD/S1311/10 (H6N6) | 4.3, 4.5 | 1.0, 1.0 | 3/3 (40–160) | 3/3 (20–80) | Highly efficient |
| CK/HuN/S4495/10 (H6N6) | 3.0, 4.5 | 1.8, 1.0 | 3/3 (20–40) | 1/3 (20) | Inefficient |
| DK/GD/S3073/10 (H6N6) | 2.3, 2.5 | 1.0, 1.0 | 3/3 (80, 160) | 0/3 | None |
| DK/GD/S4251/10 (H6N6) | 5.0, 4.8 | 5.5, 5.5 | 3/3 (160–320) | 1/3 (80) | Inefficient |
| DK/GD/S1155/11 (H6N6) | 3.5, 5.0 | 3.5, 2.5 | 3/3 (80–160) | 0/3 | None |
| CK/HuN/S3003/09 (H6N6) | 3.0, 2.5 | ND, NDb | 3/3 (10) | 0/3 | None |
Viral titers are shown as individual titers for both guinea pigs.
ND, virus not detected in sample.
To test virus transmissibility, we inoculated i.n. three guinea pigs with 106 EID50 of test virus and housed them in a cage within an isolator. Twenty-four hours later, three naive guinea pigs were placed in the same cage. Evidence of transmission was based on the detection of virus in the nasal wash and on seroconversion at the end of the 3-week observation period. As shown in Fig. 3, in the DK/GD/S4192/08 (H6N2)-, A/goose/Guangdong/S1384/2010 (H6N2) [GS/GD/S1384/10 (H6N2)-, A/duck/Guangdong/S3073/2010 (H6N6) [DK/GD/S3073/10 (H6N6)]-, A/duck/Guangdong/S1155/2011 [DK/GD/S1155/11 (H6N6)]-, and CK/HuN/S3003/09 (H6N6)-inoculated groups, virus was detected in the nasal washes of all three inoculated guinea pigs, but not in any of the contact guinea pigs (Fig. 3B, D, G, I, and J). In the A/duck/Hubei/S1114/2009 (H6N2) [DK/HuB/S1114/09 (H6N2)]-, A/duck/Hubei/S4495/2010 (H6N6) [HuB/S4495/10 (H6N6)]-, and DK/GD/S4251/10 (H6N6)-inoculated groups, virus was detected in the nasal washes of all three inoculated guinea pigs and was also detected in the nasal washes of one of the three contact animals (Fig. 3C, F, and H). In the CK/GD/S1312/10 (H6N2)- and CK/GD/S1311/10 (H6N6)-inoculated groups, virus was detected in the nasal washes of all three inoculated guinea pigs and in all three contact guinea pigs (Fig. 3A and E). Seroconversion occurred in all inoculated animals and in the contact animals that were virus positive (Table 5). These results indicate that the transmissibility of H6 avian influenza viruses in guinea pigs varies among strains, with some strains transmitting efficiently to the contact animals.
FIG 3.
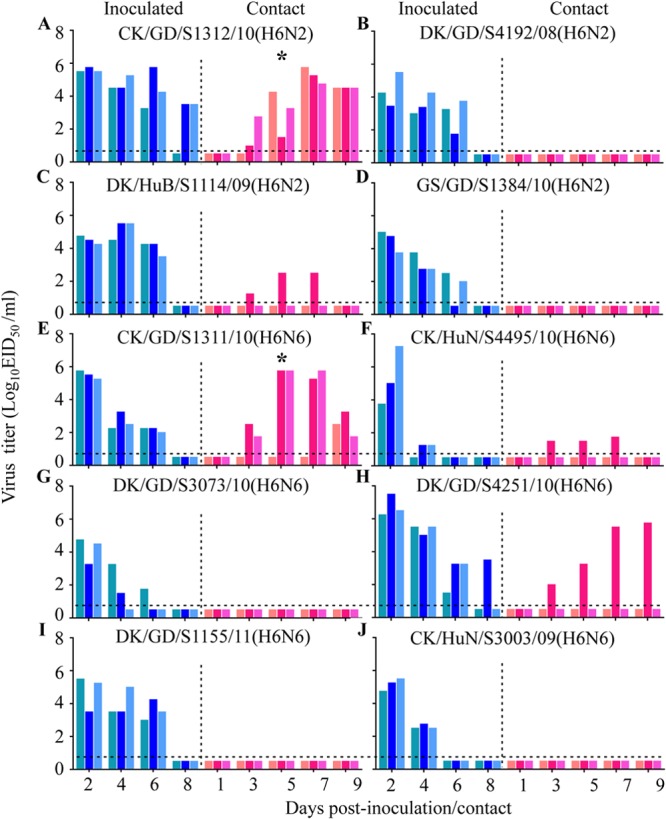
Transmission of H6 avian influenza viruses in guinea pigs. Groups of three guinea pigs were inoculated i.n. with 106 EID50 of test virus, and 24 h later, three contact guinea pigs were placed in each cage. Nasal washes were collected every 2 days from all animals beginning 2 days p.i. for detection of virus shedding. (A) CK/GD/S1312/10 (H6N2); (B) DK/GD/S4192/08 (H6N2); (C) DK/HuB/S1114/09 (H6N2); (D) GS/GD/S1384/10 (H6N2); (E) CK/GD/S1311/10 (H6N6); (F) CK/HuN/S4495/10 (H6N6); (G) DK/GD/S3073/10 (H6N6); (H) DK/GD/S4251/10 (H6N6); (I) DK/GD/S1155/11 (H6N6); and (J) CK/HuN/S3003/09 (H6N6). Each color bar represents the virus titer from an individual animal. The horizontal dashed lines in these panels indicate the lower limit of detection. Asterisks indicate that transmission efficiency was significantly higher than that of the DK/GD/S4192/08 (H6N2), GS/GD/S1384/10 (H6N2), DK/GD/S3073/10 (H6N6), DK/GD/S1155/11 (H6N6), and CK/HuN/S3003/09 (H6N6) viruses.
Replication of H6 viruses in A549 cells.
To better assess the potential risk to humans of the H6 influenza viruses, we compared the multicycle growth of four of these viruses in A549: CK/GD/S1312/10 (H6N2) and CK/GD/S1311/10 (H6N6), which bound to both α-2,3-glycans and α-2,6-glycans; and DK/GD/S4251/10 (H6N6) and CK/HuN/S3003/09 (H6N6), which bound to α-2,6-glycans and α-2,3-glycans, respectively. All of the four viruses grew efficiently in A549 cells; however, the viral titers in the CK/GD/S1311/10 (H6N6)- and DK/GD/S4251/10 (H6N6)-infected cells were significantly higher than those in the CK/HuN/S3003/09 (H6N6)-infected cells at 48 and 60 h postinfection. The viral titers in the DK/GD/S4251/10 (H6N6)-infected cells were also significantly higher than those in the CK/GD/S1312/10 (H6N2)-infected cells at 60 h postinfection and than those in cells infected with the other three viruses at 72 h postinfection (Fig. 4). The preferential binding of DK/GD/S4251/10 (H6N6) to α-2,6-glycans may have contributed to its high level of replication in A549 cells.
DISCUSSION
H6 subtype influenza viruses are widely distributed, having been detected in different species of wild birds and domestic poultry all over the world (10–24, 32–34, 36, 37, 57, 58). Here, we found that both H6N2 and H6N6 viruses have coexisted in the live poultry markets in several provinces of southern China. Our genetic analysis indicated that these viruses are complicated reassortants. We found that they can replicate in mice without preadaptation, that about 34% of them have acquired the ability to bind to the human-like receptor, and that some of them transmit efficiently between guinea pigs by direct contact.
The receptor-binding preference of HA has important implications for influenza virus replication and transmission (59–62). The HA of human influenza virus subtypes preferentially recognizes α-2,6-linked Sias, whereas the HA of avian influenza virus subtypes preferentially recognizes α-2,3-linked Sias (avian-type receptors) (61, 63). In the present study, we found that about 34% of the H6 influenza viruses derived from live poultry markets between 2008 and 2011 could bind to the α-2,6-linked Sias, although they retained high affinity for α-2,3-linked Sias. While it remains unclear what factors push an avian influenza virus to acquire the mutations that allow it to recognize the human-like receptor, these mutations do occur in different subtypes of avian influenza viruses. The recently emerged H7N9 viruses bind to both α-2,3- and α-2,6-linked Sias, although their affinities for different glycans vary among strains (9, 64–67). H5N1 avian influenza viruses have been detected in domestic poultry and wild birds in many countries and have been classified into different clades based on the diversity of their HA genes. The clade 2.2 viruses and some strains in other clades have acquired the ability to bind to both α-2,3- and α-2,6-linked Sias (68), and the mutation of 160A, which results in the absence of glycosylation at positions 158 to 160 in HA, permits these H5N1 viruses to bind to α-2,6-linked Sias (40). Mutagenesis studies have confirmed that two more amino acid mutations, 226L/228S or 224N/226L, combined with the absence of glycosylation at positions 158 to 160 in HA, enable the H5N1 influenza viruses to preferentially recognize α-2,6-linked Sias (40, 41). A G228S change was reported in the HA of the swine H6 virus and in that of human H6 virus isolates (Table 2), but the receptor-binding preferences of these strains was not reported. None of the avian H6 viruses that bound to α-2,6-linked Sias have the 226L/228S changes in their HA; therefore, it remains to be seen what mutations contributed to the affinity of H6 viruses to α-2,6-linked Sias.
We established that the H6 viruses circulating in nature have undergone severe antigenic drift; this was particularly evident from the antiserum generated from viruses bearing the HA gene from groups 4 and 5, which did not cross-react with strains bearing the HA genes from any other group. A vaccine against H6 influenza virus has never been used in China; therefore, this observed antigenic drift of H6 viruses is the result of the natural evolution of these viruses in different host species. Similar changes have been observed in H5N1 avian influenza viruses; for example, an H5N1 reassortant (clade 7.2 virus) bearing new HA, NA, and PB1 genes that was detected in northern China in 2006 has undergone severe antigenic drift from previously detected viruses of other clades (30). The naturally occurring antigenic drift of different H6 influenza viruses thus poses a challenge for vaccine strain selection for pandemic preparedness.
The mouse model has been widely used to evaluate the virulence of avian influenza viruses to mammals. Studies using this model found that the early detected H9N2 and H5N1 avian influenza viruses were unable to infect mammals but that they progressively acquired the ability to infect and cause disease and even death in mammalian species during their circulation in nature (31, 69). A previous study indicated that the H7N9 human isolates replicated much more efficiently and were more lethal in mice than were the viruses isolated from birds and that the PB2 627K mutation likely contributed to the increased virulence of the human isolates to mice (9). Gillim-Ross et al. tested 14 representative H6 viruses isolated from different avian species on two continents over a span of 36 years and found that all of the viruses could replicate in the lung of mice and that some of them were lethal (26). In the present study, we found that 37 of 38 tested viruses could replicate efficiently in the lungs of mice; although some of the viruses caused some degree of weight loss in the mice, none of them were lethal. Since these viruses can bind to the receptor in human respiratory tract cells, they may be able to infect humans easily, and during their replication in humans, they could acquire further mutations that would make them more lethal to humans, as occurred with the H7N9 viruses (9, 70).
We demonstrated that the transmissibility of H6 viruses in guinea pigs varied among strains. Some strains were efficiently transmitted to contact animals, whereas others were only inefficiently transmitted or were unable to be transmitted to contact animals despite acquiring the ability to bind to α-2,6-linked sialic acid. We speculate that the inefficiency or inability of these viruses to transmit may stem from suboptimal compatibility of their genomic segments, resulting in the failure of the viruses to be released from the infected animals. Previous studies with guinea pigs also found that some H5N1 viruses were able to transmit to contact animals, and further studies demonstrated that the amino acid change of T160A in HA, conferring binding to α-2,6-linked Sias, and the amino acid change of D701N or E627K in PB2 are important for the transmission of the H5N1 viruses in guinea pigs (40, 53). Of interest, the two H6 viruses that were efficiently transmitted to the contact guinea pigs, CK/GD/S1312/10 (H6N2) and CK/GD/S1311/10 (H6N6), shared the same gene constellation with the exception that their NA genes were of either the N2 or N6 subtype. Neither virus has any of the known amino acid changes associated with virus transmissibility, suggesting that some other amino acid changes may have contributed to the transmission of these viruses.
Genome analysis of 38 H6 viruses revealed 30 genotypes, reflecting the active circulation of these viruses in this region and their frequent reassortment with other viruses. Historically, reassortment has led to the emergence and spread of pandemic viruses in immunologically naive human populations (1, 71–73). A recent study demonstrated that H5N1 reassortants bearing certain genes from the transmissible 2009/H1N1 virus could become transmissible by respiratory droplet in mammals (74). Although human influenza virus genes have not yet been detected in H6 viruses, one cannot rule out the possibility that such reassortment will occur in the future. The 2009/H1N1 pandemic virus is still circulating in humans and pigs around the world (75–78), and H6 virus infection of pigs and humans has been recorded in different geographic areas (28, 29, 79, 80). Clearly, the possibility exists for H6 reassortants that are transmissible by respiratory droplet in mammals to be generated by deriving genes from transmissible human influenza viruses.
Supplementary Material
ACKNOWLEDGMENTS
We thank S. Watson for editing the manuscript and Yamasa Corporation Co. Ltd. for synthesizing the sialylglycopolymer.
This work was supported by the Ministry of Agriculture (CARS-42-G08) and by the Ministry of Science and Technology (KJYJ-2013-01-01 and 2012ZX10004214).
Footnotes
Published ahead of print 5 February 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JVI.03292-13.
REFERENCES
- 1.Kawaoka Y, Krauss S, Webster RG. 1989. Avian-to-human transmission of the PB1 gene of influenza A viruses in the 1957 and 1968 pandemics. J. Virol. 63:4603–4608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Taubenberger JK, Reid AH, Krafft AE, Bijwaard KE, Fanning TG. 1997. Initial genetic characterization of the 1918 “Spanish” influenza virus. Science 275:1793–1796. 10.1126/science.275.5307.1793 [DOI] [PubMed] [Google Scholar]
- 3.Webster RG, Bean WJ, Gorman OT, Chambers TM, Kawaoka Y. 1992. Evolution and ecology of influenza A viruses. Microbiol. Rev. 56:152–179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.World Health Organization. 8 October 2013. Cumulative number of confirmed human cases for avian influenza A(H5N1) reported to WHO, 2003–2013. World Health Organization, Geneva, Switzerland: http://www.who.int/influenza/human_animal_interface/EN_GIP_20131008CumulativeNumberH5N1cases.pdf [Google Scholar]
- 5.Abbott A. 2003. Human fatality adds fresh impetus to fight against bird flu. Nature 423:5. 10.1038/423005b [DOI] [PubMed] [Google Scholar]
- 6.Homme PJ, Easterday BC, Anderson DP. 1970. Avian influenza virus infections. II. Experimental epizootiology of influenza A-turkey-Wisconsin-1966 virus in turkeys. Avian Dis. 14:240–247 [PubMed] [Google Scholar]
- 7.Peiris M, Yuen KY, Leung CW, Chan KH, Ip PL, Lai RW, Orr WK, Shortridge KF. 1999. Human infection with influenza H9N2. Lancet 354:916–917. 10.1016/S0140-6736(99)03311-5 [DOI] [PubMed] [Google Scholar]
- 8.World Health Organization. 25 October 2013. Number of confirmed human cases of avian influenza A(H7N9) reported to WHO. World Health Organization, Geneva, Switzerland: http://www.who.int/influenza/human_animal_interface/influenza_h7n9/10u_ReportWebH7N9Number.pdf [Google Scholar]
- 9.Zhang Q, Shi J, Deng G, Guo J, Zeng X, He X, Kong H, Gu C, Li X, Liu J, Wang G, Chen Y, Liu L, Liang L, Li Y, Fan J, Wang J, Li W, Guan L, Li Q, Yang H, Chen P, Jiang L, Guan Y, Xin X, Jiang Y, Tian G, Wang X, Qiao C, Li C, Bu Z, Chen H. 2013. H7N9 influenza viruses are transmissible in ferrets by respiratory droplet. Science 341:410–414. 10.1126/science.1240532 [DOI] [PubMed] [Google Scholar]
- 10.Abolnik C, Bisschop S, Gerdes T, Olivier A, Horner R. 2007. Outbreaks of avian influenza H6N2 viruses in chickens arose by a reassortment of H6N8 and H9N2 ostrich viruses. Virus Genes 34:37–45. 10.1007/s11262-006-0007-6 [DOI] [PubMed] [Google Scholar]
- 11.Choi YK, Seo SH, Kim JA, Webby RJ, Webster RG. 2005. Avian influenza viruses in Korean live poultry markets and their pathogenic potential. Virology 332:529–537. 10.1016/j.virol.2004.12.002 [DOI] [PubMed] [Google Scholar]
- 12.Wang CW, Wang CH. 2003. Experimental selection of virus derivatives with variations in virulence from a single low-pathogenicity H6N1 avian influenza virus field isolate. Avian Dis. 47:1416–1422. 10.1637/6093 [DOI] [PubMed] [Google Scholar]
- 13.Woolcock PR, Suarez DL, Kuney D. 2003. Low-pathogenicity avian influenza virus (H6N2) in chickens in California, 2000-02. Avian Dis. 47:872–881. 10.1637/0005-2086-47.s3.872 [DOI] [PubMed] [Google Scholar]
- 14.Alexander DJ. 2003. Report on avian influenza in the Eastern Hemisphere during 1997-2002. Avian Dis. 47:792–797. 10.1637/0005-2086-47.s3.792 [DOI] [PubMed] [Google Scholar]
- 15.Chin PS, Hoffmann E, Webby R, Webster RG, Guan Y, Peiris M, Shortridge KF. 2002. Molecular evolution of H6 influenza viruses from poultry in Southeastern China: prevalence of H6N1 influenza viruses possessing seven A/Hong Kong/156/97 (H5N1)-like genes in poultry. J. Virol. 76:507–516. 10.1128/JVI.76.2.507-516.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Downie JC, Laver WG. 1973. Isolation of a type A influenza virus from an Australian pelagic bird. Virology 51:259–269. 10.1016/0042-6822(73)90426-1 [DOI] [PubMed] [Google Scholar]
- 17.Hanson BA, Stallknecht DE, Swayne DE, Lewis LA, Senne DA. 2003. Avian influenza viruses in Minnesota ducks during 1998-2000. Avian Dis. 47:867–871. 10.1637/0005-2086-47.s3.867 [DOI] [PubMed] [Google Scholar]
- 18.Kinde H, Read DH, Daft BM, Hammarlund M, Moore J, Uzal F, Mukai J, Woolcock P. 2003. The occurrence of avian influenza A subtype H6N2 in commercial layer flocks in Southern California (2000-02): clinicopathologic findings. Avian Dis. 47:1214–1218. 10.1637/0005-2086-47.s3.1214 [DOI] [PubMed] [Google Scholar]
- 19.Panigrahy B, Senne DA, Pedersen JC. 2002. Avian influenza virus subtypes inside and outside the live bird markets, 1993-2000: a spatial and temporal relationship. Avian Dis. 46:298–307. 10.1637/0005-2086(2002)046[0298:AIVSIA]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 20.Senne DA. 2003. Avian influenza in the Western Hemisphere including the Pacific Islands and Australia. Avian Dis. 47:798–805. 10.1637/0005-2086-47.s3.798 [DOI] [PubMed] [Google Scholar]
- 21.Slemons RD, Hansen WR, Converse KA, Senne DA. 2003. Type A influenza virus surveillance in free-flying, nonmigratory ducks residing on the eastern shore of Maryland. Avian Dis. 47:1107–1110. 10.1637/0005-2086-47.s3.1107 [DOI] [PubMed] [Google Scholar]
- 22.Spackman E, Stallknecht DE, Slemons RD, Winker K, Suarez DL, Scott M, Swayne DE. 2005. Phylogenetic analyses of type A influenza genes in natural reservoir species in North America reveals genetic variation. Virus Res. 114:89–100. 10.1016/j.virusres.2005.05.013 [DOI] [PubMed] [Google Scholar]
- 23.Webby RJ, Woolcock PR, Krauss SL, Walker DB, Chin PS, Shortridge KF, Webster RG. 2003. Multiple genotypes of nonpathogenic H6N2 influenza viruses isolated from chickens in California. Avian Dis. 47:905–910. 10.1637/0005-2086-47.s3.905 [DOI] [PubMed] [Google Scholar]
- 24.Shortridge KF, Butterfield WK, Webster RG, Campbell CH. 1977. Isolation and characterization of influenza A viruses from avian species in Hong Kong. Bull. World Health Organ. 55:15–20 [PMC free article] [PubMed] [Google Scholar]
- 25.Munster VJ, Baas C, Lexmond P, Waldenstrom J, Wallensten A, Fransson T, Rimmelzwaan GF, Beyer WE, Schutten M, Olsen B, Osterhaus AD, Fouchier RA. 2007. Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathog. 3:e61. 10.1371/journal.ppat.0030061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gillim-Ross L, Santos C, Chen Z, Aspelund A, Yang CF, Ye D, Jin H, Kemble G, Subbarao K. 2008. Avian influenza H6 viruses productively infect and cause illness in mice and ferrets. J. Virol. 82:10854–10863. 10.1128/JVI.01206-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nam JH, Kim EH, Song D, Choi YK, Kim JK, Poo H. 2011. Emergence of mammalian species-infectious and -pathogenic avian influenza H6N5 virus with no evidence of adaptation. J. Virol. 85:13271–13277. 10.1128/JVI.05038-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang G, Kong W, Qi W, Long LP, Cao Z, Huang L, Qi H, Cao N, Wang W, Zhao F, Ning Z, Liao M, Wan XF. 2011. Identification of an H6N6 swine influenza virus in southern China. Infect. Genet. Evol. 11:1174–1177. 10.1016/j.meegid.2011.02.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Yuan J, Zhang L, Kan X, Jiang L, Yang J, Guo Z, Ren Q. 2013. Origin and molecular characteristics of a novel 2013 avian influenza A(H6N1) virus causing human infection in Taiwan. Clin. Infect. Dis. 57:1367–1368. 10.1093/cid/cit479 [DOI] [PubMed] [Google Scholar]
- 30.Li Y, Shi J, Zhong G, Deng G, Tian G, Ge J, Zeng X, Song J, Zhao D, Liu L, Jiang Y, Guan Y, Bu Z, Chen H. 2010. Continued evolution of H5N1 influenza viruses in wild birds, domestic poultry, and humans in China from 2004 to 2009. J. Virol. 84:8389–8397. 10.1128/JVI.00413-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chen H, Deng G, Li Z, Tian G, Li Y, Jiao P, Zhang L, Liu Z, Webster RG, Yu K. 2004. The evolution of H5N1 influenza viruses in ducks in southern China. Proc. Natl. Acad. Sci. U. S. A. 101:10452–10457. 10.1073/pnas.0403212101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Cheung CL, Vijaykrishna D, Smith GJ, Fan XH, Zhang JX, Bahl J, Duan L, Huang K, Tai H, Wang J, Poon LL, Peiris JS, Chen H, Guan Y. 2007. Establishment of influenza A virus (H6N1) in minor poultry species in southern China. J. Virol. 81:10402–10412. 10.1128/JVI.01157-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Huang K, Bahl J, Fan XH, Vijaykrishna D, Cheung CL, Webby RJ, Webster RG, Chen H, Smith GJ, Peiris JS, Guan Y. 2010. Establishment of an H6N2 influenza virus lineage in domestic ducks in southern China. J. Virol. 84:6978–6986. 10.1128/JVI.00256-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Huang K, Zhu H, Fan X, Wang J, Cheung CL, Duan L, Hong W, Liu Y, Li L, Smith DK, Chen H, Webster RG, Webby RJ, Peiris M, Guan Y. 2012. Establishment and lineage replacement of H6 influenza viruses in domestic ducks in southern China. J. Virol. 86:6075–6083. 10.1128/JVI.06389-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Deng G, Tan D, Shi J, Cui P, Jiang Y, Liu L, Tian G, Kawaoka Y, Li C, Chen H. 2013. Complex reassortment of multiple subtypes of avian influenza viruses in domestic ducks at the Dongting Lake region of China. J. Virol. 87:9452–9462. 10.1128/JVI.00776-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zhao G, Lu X, Gu X, Zhao K, Song Q, Pan J, Xu Q, Duan Z, Peng D, Hu S, Wang X, Liu X. 2011. Molecular evolution of the H6 subtype influenza A viruses from poultry in eastern China from 2002 to 2010. Virol. J. 8:470. 10.1186/1743-422X-8-470 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee MS, Chang PC, Shien JH, Cheng MC, Chen CL, Shieh HK. 2006. Genetic and pathogenic characterization of H6N1 avian influenza viruses isolated in Taiwan between 1972 and 2005. Avian Dis. 50:561–571. 10.1637/7640-050106R.1 [DOI] [PubMed] [Google Scholar]
- 38.Rimondi A, Xu K, Craig MI, Shao H, Ferreyra H, Rago MV, Romano M, Uhart M, Sutton T, Ferrero A, Perez DR, Pereda A. 2011. Phylogenetic analysis of H6 influenza viruses isolated from rosy-billed pochards (Netta peposaca) in Argentina reveals the presence of different HA gene clusters. J. Virol. 85:13354–13362. 10.1128/JVI.05946-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hoffmann E, Stech J, Leneva I, Krauss S, Scholtissek C, Chin PS, Peiris M, Shortridge KF, Webster RG. 2000. Characterization of the influenza A virus gene pool in avian species in southern China: was H6N1 a derivative or a precursor of H5N1? J. Virol. 74:6309–6315. 10.1128/JVI.74.14.6309-6315.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gao Y, Zhang Y, Shinya K, Deng G, Jiang Y, Li Z, Guan Y, Tian G, Li Y, Shi J, Liu L, Zeng X, Bu Z, Xia X, Kawaoka Y, Chen H. 2009. Identification of amino acids in HA and PB2 critical for the transmission of H5N1 avian influenza viruses in a mammalian host. PLoS Pathog. 5:e1000709. 10.1371/journal.ppat.1000709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Imai M, Watanabe T, Hatta M, Das SC, Ozawa M, Shinya K, Zhong G, Hanson A, Katsura H, Watanabe S, Li C, Kawakami E, Yamada S, Kiso M, Suzuki Y, Maher EA, Neumann G, Kawaoka Y. 2012. Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets. Nature 486:420–428. 10.1038/nature10831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Munster VJ, de Wit E, van den Brand JM, Herfst S, Schrauwen EJ, Bestebroer TM, van de Vijver D, Boucher CA, Koopmans M, Rimmelzwaan GF, Kuiken T, Osterhaus AD, Fouchier RA. 2009. Pathogenesis and transmission of swine-origin 2009 A(H1N1) influenza virus in ferrets. Science 325:481–483. 10.1126/science.1177127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Maines TR, Jayaraman A, Belser JA, Wadford DA, Pappas C, Zeng H, Gustin KM, Pearce MB, Viswanathan K, Shriver ZH, Raman R, Cox NJ, Sasisekharan R, Katz JM, Tumpey TM. 2009. Transmission and pathogenesis of swine-origin 2009 A(H1N1) influenza viruses in ferrets and mice. Science 325:484–487. 10.1126/science.1177238 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tumpey TM, Maines TR, Van Hoeven N, Glaser L, Solorzano A, Pappas C, Cox NJ, Swayne DE, Palese P, Katz JM, Garcia-Sastre A. 2007. A two-amino acid change in the hemagglutinin of the 1918 influenza virus abolishes transmission. Science 315:655–659. 10.1126/science.1136212 [DOI] [PubMed] [Google Scholar]
- 45.Zhang Y, Zhang Q, Gao Y, He X, Kong H, Jiang Y, Guan Y, Xia X, Shu Y, Kawaoka Y, Bu Z, Chen H. 2012. Key molecular factors in hemagglutinin and PB2 contribute to efficient transmission of the 2009 H1N1 pandemic influenza virus. J. Virol. 86:9666–9674. 10.1128/JVI.00958-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Matrosovich MN, Gambaryan AS, Teneberg S, Piskarev VE, Yamnikova SS, Lvov DK, Robertson JS, Karlsson KA. 1997. Avian influenza A viruses differ from human viruses by recognition of sialyloligosaccharides and gangliosides and by a higher conservation of the HA receptor-binding site. Virology 233:224–234. 10.1006/viro.1997.8580 [DOI] [PubMed] [Google Scholar]
- 47.Nidom CA, Takano R, Yamada S, Sakai-Tagawa Y, Daulay S, Aswadi D, Suzuki T, Suzuki Y, Shinya K, Iwatsuki-Horimoto K, Muramoto Y, Kawaoka Y. 2010. Influenza A (H5N1) viruses from pigs, Indonesia. Emerg. Infect. Dis. 16:1515–1523. 10.3201/eid1610.100508 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xie Z, Xie L, Zhou C, Liu J, Pang Y, Deng X, Xie Z, Fan Q. 2012. Complete genome sequence analysis of an H6N1 avian influenza virus isolated from Guangxi pockmark ducks. J. Virol. 86:13868–13869. 10.1128/JVI.02700-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Yao Y, Wang H, Chen Q, Zhang H, Zhang T, Chen J, Xu B, Wang H, Sun B, Chen Z. 2013. Characterization of low-pathogenic H6N6 avian influenza viruses in central China. Arch. Virol. 158:367–377. 10.1007/s00705-012-1496-3 [DOI] [PubMed] [Google Scholar]
- 50. Reference deleted.
- 51.Hatta M, Gao P, Halfmann P, Kawaoka Y. 2001. Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science 293:1840–1842. 10.1126/science.1062882 [DOI] [PubMed] [Google Scholar]
- 52.Li Z, Chen H, Jiao P, Deng G, Tian G, Li Y, Hoffmann E, Webster RG, Matsuoka Y, Yu K. 2005. Molecular basis of replication of duck H5N1 influenza viruses in a mammalian mouse model. J. Virol. 79:12058–12064. 10.1128/JVI.79.18.12058-12064.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Steel J, Lowen AC, Mubareka S, Palese P. 2009. Transmission of influenza virus in a mammalian host is increased by PB2 amino acids 627K or 627E/701N. PLoS Pathog. 5:e1000252. 10.1371/journal.ppat.1000252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Jackson D, Hossain MJ, Hickman D, Perez DR, Lamb RA. 2008. A new influenza virus virulence determinant: the NS1 protein four C-terminal residues modulate pathogenicity. Proc. Natl. Acad. Sci. U. S. A. 105:4381–4386. 10.1073/pnas.0800482105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Obenauer JC, Denson J, Mehta PK, Su X, Mukatira S, Finkelstein DB, Xu X, Wang J, Ma J, Fan Y, Rakestraw KM, Webster RG, Hoffmann E, Krauss S, Zheng J, Zhang Z, Naeve CW. 2006. Large-scale sequence analysis of avian influenza isolates. Science 311:1576–1580. 10.1126/science.1121586 [DOI] [PubMed] [Google Scholar]
- 56. Reference deleted.
- 57.Reperant LA, Fuckar NS, Osterhaus AD, Dobson AP, Kuiken T. 2010. Spatial and temporal association of outbreaks of H5N1 influenza virus infection in wild birds with the 0°C isotherm. PLoS Pathog. 6:e1000854. 10.1371/journal.ppat.1000854 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kim HR, Lee YJ, Lee KK, Oem JK, Kim SH, Lee MH, Lee OS, Park CK. 2010. Genetic relatedness of H6 subtype avian influenza viruses isolated from wild birds and domestic ducks in Korea and their pathogenicity in animals. J. Gen. Virol. 91:208–219. 10.1099/vir.0.015800-0 [DOI] [PubMed] [Google Scholar]
- 59.Glaser L, Stevens J, Zamarin D, Wilson IA, Garcia-Sastre A, Tumpey TM, Basler CF, Taubenberger JK, Palese P. 2005. A single amino acid substitution in 1918 influenza virus hemagglutinin changes receptor binding specificity. J. Virol. 79:11533–11536. 10.1128/JVI.79.17.11533-11536.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Matrosovich M, Tuzikov A, Bovin N, Gambaryan A, Klimov A, Castrucci MR, Donatelli I, Kawaoka Y. 2000. Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals. J. Virol. 74:8502–8512. 10.1128/JVI.74.18.8502-8512.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Herfst S, Schrauwen EJ, Linster M, Chutinimitkul S, de Wit E, Munster VJ, Sorrell EM, Bestebroer TM, Burke DF, Smith DJ, Rimmelzwaan GF, Osterhaus AD, Fouchier RA. 2012. Airborne transmission of influenza A/H5N1 virus between ferrets. Science 336:1534–1541. 10.1126/science.1213362 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Vines A, Wells K, Matrosovich M, Castrucci MR, Ito T, Kawaoka Y. 1998. The role of influenza A virus hemagglutinin residues 226 and 228 in receptor specificity and host range restriction. J. Virol. 72:7626–7631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rogers GN, Paulson JC. 1983. Receptor determinants of human and animal influenza virus isolates: differences in receptor specificity of the H3 hemagglutinin based on species of origin. Virology 127:361–373. 10.1016/0042-6822(83)90150-2 [DOI] [PubMed] [Google Scholar]
- 64.Zhou J, Wang D, Gao R, Zhao B, Song J, Qi X, Zhang Y, Shi Y, Yang L, Zhu W, Bai T, Qin K, Lan Y, Zou S, Guo J, Dong J, Dong L, Wei H, Li X, Lu J, Liu L, Zhao X, Huang W, Wen L, Bo H, Xin L, Chen Y, Xu C, Pei Y, Yang Y, Zhang X, Wang S, Feng Z, Han J, Yang W, Gao GF, Wu G, Li D, Wang Y, Shu Y. 2013. Biological features of novel avian influenza A (H7N9) virus. Nature 499:500–503. 10.1038/nature12379 [DOI] [PubMed] [Google Scholar]
- 65.Watanabe T, Kiso M, Fukuyama S, Nakajima N, Imai M, Yamada S, Murakami S, Yamayoshi S, Iwatsuki-Horimoto K, Sakoda Y, Takashita E, McBride R, Noda T, Hatta M, Imai H, Zhao D, Kishida N, Shirakura M, de Vries RP, Shichinohe S, Okamatsu M, Tamura T, Tomita Y, Fujimoto N, Goto K, Katsura H, Kawakami E, Ishikawa I, Watanabe S, Ito M, Sakai-Tagawa Y, Sugita Y, Uraki R, Yamaji R, Eisfeld AJ, Zhong G, Fan S, Ping J, Maher EA, Hanson A, Uchida Y, Saito T, Ozawa M, Neumann G, Kida H, Odagiri T, Paulson JC, Hasegawa H, Tashiro M, Kawaoka Y. 2013. Characterization of H7N9 influenza A viruses isolated from humans. Nature 501:551–555. 10.1038/nature12392 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Tharakaraman K, Jayaraman A, Raman R, Viswanathan K, Stebbins NW, Johnson D, Shriver Z, Sasisekharan V, Sasisekharan R. 2013. Glycan receptor binding of the influenza A virus H7N9 hemagglutinin. Cell 153:1486–1493. 10.1016/j.cell.2013.05.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Richard M, Schrauwen EJ, de Graaf M, Bestebroer TM, Spronken MI, van Boheemen S, de Meulder D, Lexmond P, Linster M, Herfst S, Smith DJ, van den Brand JM, Burke DF, Kuiken T, Rimmelzwaan GF, Osterhaus AD, Fouchier RA. 2013. Limited airborne transmission of H7N9 influenza A virus between ferrets. Nature 501:560–563. 10.1038/nature12476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Stevens J, Blixt O, Chen LM, Donis RO, Paulson JC, Wilson IA. 2008. Recent avian H5N1 viruses exhibit increased propensity for acquiring human receptor specificity. J. Mol. Biol. 381:1382–1394. 10.1016/j.jmb.2008.04.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Li C, Yu K, Tian G, Yu D, Liu L, Jing B, Ping J, Chen H. 2005. Evolution of H9N2 influenza viruses from domestic poultry in mainland China. Virology 340:70–83. 10.1016/j.virol.2005.06.025 [DOI] [PubMed] [Google Scholar]
- 70.Shi J, Deng G, Liu P, Zhou J, Guan L, Li W, Li X, Guo J, Wang G, Fan J, Wang J, Li Y, Jiang Y, Liu L, Tian G, Li C, Chen H. 2013. Isolation and characterization of H7N9 viruses from live poultry markets—implication of the source of current H7N9 infection in humans. Chin. Sci. Bull. 58:1857–1863. 10.1007/s11434-013-5873-4 [DOI] [Google Scholar]
- 71.Scholtissek C, Rohde W, Von Hoyningen V, Rott R. 1978. On the origin of the human influenza virus subtypes H2N2 and H3N2. Virology 87:13–20. 10.1016/0042-6822(78)90153-8 [DOI] [PubMed] [Google Scholar]
- 72.Lindstrom SE, Cox NJ, Klimov A. 2004. Genetic analysis of human H2N2 and early H3N2 influenza viruses, 1957-1972: evidence for genetic divergence and multiple reassortment events. Virology 328:101–119. 10.1016/j.virol.2004.06.009 [DOI] [PubMed] [Google Scholar]
- 73.Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, Sessions WM, Xu X, Skepner E, Deyde V, Okomo-Adhiambo M, Gubareva L, Barnes J, Smith CB, Emery SL, Hillman MJ, Rivailler P, Smagala J, de Graaf M, Burke DF, Fouchier RA, Pappas C, Alpuche-Aranda CM, Lopez-Gatell H, Olivera H, Lopez I, Myers CA, Faix D, Blair PJ, Yu C, Keene KM, Dotson PD, Jr, Boxrud D, Sambol AR, Abid SH, St George K, Bannerman T, Moore AL, Stringer DJ, Blevins P, Demmler-Harrison GJ, Ginsberg M, Kriner P, Waterman S, Smole S, Guevara HF, Belongia EA, Clark PA, Beatrice ST, Donis R, Katz J, Finelli L, Bridges CB, Shaw M, Jernigan DB, Uyeki TM, Smith DJ, Klimov AI, Cox NJ. 2009. Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science 325:197–201. 10.1126/science.1176225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhang Y, Zhang Q, Kong H, Jiang Y, Gao Y, Deng G, Shi J, Tian G, Liu L, Liu J, Guan Y, Bu Z, Chen H. 2013. H5N1 hybrid viruses bearing 2009/H1N1 virus genes transmit in guinea pigs by respiratory droplet. Science 340:1459–1463. 10.1126/science.1229455 [DOI] [PubMed] [Google Scholar]
- 75.Yan JH, Xiong Y, Yi CH, Sun XX, He QS, Fu W, Xu XK, Jiang JX, Ma L, Liu Q. 2012. Pandemic (H1N1) 2009 virus circulating in pigs, Guangxi, China. Emerg. Infect. Dis. 18:357–359. 10.3201/eid1802.111346 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Pasma T, Joseph T. 2010. Pandemic (H1N1) 2009 infection in swine herds, Manitoba, Canada. Emerg. Infect. Dis. 16:706–708. 10.3201/eid1604.091636 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Sreta D, Tantawet S, Na Ayudhya SN, Thontiravong A, Wongphatcharachai M, Lapkuntod J, Bunpapong N, Tuanudom R, Suradhat S, Vimolket L, Poovorawan Y, Thanawongnuwech R, Amonsin A, Kitikoon P. 2010. Pandemic (H1N1) 2009 virus on commercial swine farm, Thailand. Emerg. Infect. Dis. 16:1587–1590. 10.3201/eid1610.100665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Chowell G, Echevarria-Zuno S, Viboud C, Simonsen L, Grajales Muniz C, Rascon Pacheco RA, Gonzalez Leon M, Borja Aburto VH. 2012. Recrudescent wave of pandemic A/H1N1 influenza in Mexico, winter 2011-2012: age shift and severity. PLoS Curr. 4:RRN1306. 10.1371/currents.RRN1306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Kayali G, Ortiz EJ, Chorazy ML, Gray GC. 2010. Evidence of previous avian influenza infection among US turkey workers. Zoonoses Public Health 57:265–272. 10.1111/j.1863-2378.2009.01231.x [DOI] [PubMed] [Google Scholar]
- 80.Myers KP, Setterquist SF, Capuano AW, Gray GC. 2007. Infection due to 3 avian influenza subtypes in United States veterinarians. Clin. Infect. Dis. 45:4–9. 10.1086/518579 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.