

Abstract
The presence of melanocytes in the oral epithelium is a well-established fact, but their physiological functions are not well defined. Melanin provides protection from environmental stressors such as ultraviolet radiation and reactive oxygen species; and melanocytes function as stress-sensors having the capacity both to react to and to produce a variety of microenvironmental cytokines and growth factors, modulating immune, inflammatory and antibacterial responses. Melanocytes also act as neuroendocrine cells producing local neurotransmitters including acetylcholine, catecholamines and opioids, and hormones of the melanocortin system such as proopiomelanocortin, adrenocorticotropic hormone and α-melanocyte stimulating hormone, that participate in intracellular and in intercellular signalling pathways, thus contributing to tissue homeostasis.
There is a wide range of normal variation in melanin pigmentation of the oral mucosa. In general, darker skinned persons more frequently have oral melanin pigmentation than light-skinned persons. Variations in oral physiological pigmentation are genetically determined unless associated with some underlying disease.
In this article, we discuss some aspects of the biophysiology of oral melanocytes, of the functions of melanin, and of physiological oral pigmentation.
Keywords: Melanin, Oral melanocytes, Pigmentation, Melanocyte stem cell, Keratinocyte, Melanosome, Pigment transfer
Introduction
While under physiological conditions the number of melanocytes in the oral epithelium is the same regardless of racial/ethnic origin [1], the colour of oral mucosa varies between persons and is determined by several factors including the number and melanogenic activity of the melanocytes in the basal cell layer of the epithelium, differences in number, size, and distribution of melanosomes, differences in the type of melanins, and the masking effect of heavily keratinized epithelium [2-4]. The variations in melanin colourisation of the oral mucosa will be determined against a background of the degree of vascularization of the tissues and by the level of haemoglobin in the blood [2].
Melanocytes containing melanin are present in the basal cell layer of the epithelium even at those oral mucosal sites that show no visible signs of melanin pigmentation [5-7]. Oral melanocytes may or may not produce melanin, but as in the case of the skin, the amount of melanin they produce is genetically determined [2,6]. There are substantial variations in the degree of melanin pigmentation between persons of different racial/ethnic groups and between persons of the same racial/ethnic group, and these variations are normal [2,8,9]. Physiological/racial melanin pigmentation of the oral mucosa is common in black persons [7], and is more frequent in darker skinned whites (Caucasians) than in lighter skinned whites [10]. The oral mucosal melanin pigmentation may be patchy or uniform and most commonly affects the gingiva [9].
Melanocytes produce melanin in membrane-bound organelles termed melanosomes. Melanosomes have all the proteins and enzymes necessary for melanin biosynthesis, for maintaining the structure of the melanosome, and for the maturation of the immature pre-melanosome into a mature melanosome producing melanin [11,12]. As the melanosomes mature intracellularly, they are transported via microtubuli to the surfaces of the elongated dendrites of the melanocytes whence they are ultimately transferred to the keratinocytes in the ‘keratinocyte melanin unit’ [11,12]. Within the keratinocytes, the melanin is preferentially localised within the nuclei forming protective barriers known as supranuclear ‘caps’ oriented in such a way that they shield the nuclear DNA from ultraviolet (UV) radiation [13].
Non-physiological alterations in melanin pigmentation of the oral mucosa are related to genetic, metabolic, endocrine, chemical or physical factors, to infective agents and to inflammatory or neoplastic processes [6,9]. It has been reported that in about 30% of cases, oral mucosal melanoma develops at sites of hyperpigmentation [2,14]. However, the nature of the hyperpigmented pre-melanoma is obscure.
Most of our knowledge about the regulation and biology of oral melanocytes is derived from data obtained from research on epidermal melanocytes which are histologically and ultra-structurally similar [15,16]. However, when extrapolating data from epidermal melanocytes, one needs to bear in mind that as a rule, oral mucosal melanocytes under physiological circumstances are said to be less metabolically active [6]. Nevertheless, in some cases oral melanocytes may be inherently metabolically active, and in other cases an increase in metabolic activity may be in response to environmental triggers such as hormones, inflammation or injury [6].
The aim of this short review is to discuss some aspects of the biology and physiology of oral melanocytes, the function of melanin and critically to evaluate the concept of intraoral physiological pigmentation.
The origin of oral melanocytes
Melanocytes are melanin-producing cells originating from the neural crest [17]. During development melanocyte stem cells migrate from the neural crest to the skin and to mucous membranes. Active melanocytes are present in the stria vascularis of the cochlea, in the leptomeninges, in the substantia nigra and locus coerulus of the brain, and in the heart, where they play a number of as yet ill-defined roles [13,18].
Melanocyte stem cells have the capacity for self-renewal and for differentiation and thus can maintain the population of mature melanocytes. In the epidermis, the melanocyte stem cells reside in the bulge region of hair follicles [19,20], but the niche in which they reside in the oral mucosa is unknown.
Epidermal melanocyte stem cells give rise to transient-amplifying melanocyte precursors that settle without the stem cell niche where they differentiate into mature melanin producing melanocytes [19,21-24]. Stem cell factor (SCF) and its tyrosine kinase receptor C-kit signalling pathways are critical for epidermal melanocyte development during embryogenesis [17,20], notch signalling pathways are essential in the maintenance of adult melanocyte stem cells, and thus for melanocyte homeostasis [20], endothelin 1 plays a role in the differentiation of melanocyte precursors, and micropthalmia-associated transcription factor (MITF) with its cAMP response element play a critical role in melanogenesis [25].
In the skin, migration of precursor melanocytes from the dermis to their final destination in the basal cell layer of the epithelium is mediated by c-Kit/SCF, endothelin 1 and 3, hepatocyte growth factor (HGF) and basic fibroblast growth factor (bFGF). These precursor dermal melanocytes, as they pass through the basement membrane, express E-cadherin which later facilitates intercellular communication with neighbouring keratinocytes in the basal cell layer of the epithelium [1]. Sometimes, melanocyte precursors on their way to the epithelium may become arrested in the lamina propria/dermis, and as they have the capacity to produce melanin, if they are aggregated they will form nevi [26]. The functional activity of melanocytes, both those in the basal cell layer of the epithelium and those that may have become arrested in the lamina propria is influenced by signals from the neighbouring fibroblasts [13,27,28].
The keratinocyte-melanocyte unit
Mature melanocytes are elongated dendritic cells residing in the basal cell layer of the epithelium. They contain all the proteins required for melanin biosynthesis and for the structural maturation of melanosomes [12], including tryosinase (TYR), tyrosinase-related proteins-1 (TRP-1) and TRP-2, gp 100, and melanoma antigen recognizable by T lymphocytes (MART-1) [13,29-31].
In the basal cell layer of the epithelium the ratio of melanocytes to keratinocytes ranges from 1:10 [30] to 1:15 [6]. Melanocytes and keratinocytes form epidermal melanin units, each unit consists of one melanocyte and a group of about 36 neighbouring keratinocytes. The melanosomes produced by melanocytes are disseminated via a network of melanocytic dendritic processes to the keratinocytes of the epidermal melanin unit [25,32]. It appears that keratinocytes have some control over the process of dendritic melanosome transfer since the capacity of keratinocytes to phagocytose the melanosomes is influenced by the degree of activation of the protease-activated receptor 2 (PAR-2) on the surface of keratinocytes [33].
It has been suggested that through biological mediators present in the melanosomes transferred to keratinocytes, melanocytes can influence the functional activities of keratinocytes [13], and keratinocytes via an array of paracrine-like biological mediators that they secrete, have the capacity to regulate melanocyte melanogenesis [13,25,34]. These mediators include α-melanocyte stimulating hormone (α-MSH), adrenocorticotropin hormone (ACTH), β-endorphin, bFGF, endothelins, HGF and SCF [35].
However, such a mechanistic description is an oversimplification as the ratio of keratinocytes to melanocytes is different at different stages of growth and development, and in adulthood the ratio is variable being determined by biological mediators secreted in the local microenvironment [33]. Furthermore, the numerical density of melanocytes in the epithelium varies in different parts of the skin or oral mucosa, and between the same skin or mucosa sites in different persons, regardless of their racial/ethnic origin [36]. It appears that Langerhans cells in the epithelium, and fibroblasts in the subepithelial connective tissue play an important role in maintaining the functional activity of the epidermal melanin unit [30,31,33].
The adherence of melanocytes to keratinocytes is by means of tight junctions where they co-express E-cadherin cell adhesion molecules and by gap junctions [33]. E-cadherin supresses melanocyte proliferation, but a switch from E-cadherin to N-cadherin owing to metabolically or traumatically triggered events in the microenvironment obviates the melanocytes from growth suppression. Melanocytes expressing N-cadherin can freely proliferate, migrate and self-aggregate in nests, and exhibit longevity [32].
The function of oral melanocytes
The functions of melanocytes are not fully understood, but it is clear that the melanin that they produce determines the colour of skin, hair and eyes [37], and provides protection from stressors such as UV radiation, reactive oxygen species (ROS) and free radicals in the environment. Melanins also have the capacity to sequester metal ions and to bind certain drugs and organic molecules [32,38].
The colour of skin and probably of any pigmented part of the oral mucosa is genetically determined by the number and size of the melanosomes and the type of melanin (eumelanin, pheomelanin) that they produce. Environmental factors have only a modifying influence on skin colour, though on an evolutionary scale this influence can have a more profound effect. Melanosomes vary in size and contain both eumelanin and pheomelanin. More, larger eumelanin impart a dark colour to the skin [33]. Small, few pheomelanin-containing melanosomes are associated with light skin, and the spectrum of size and number of melanosomes and the balance of eumelanin to pheomelanin within the melanosomes, will determine all the other colour variations [32].
As melanin synthesis is an oxygen dependent process, paradoxically, it also generates ROS that may accumulate in the melanocytes and cause DNA damage, and in fact UV radiation exaggerates the production of ROS during the biosynthesis of melanin and more particularly pheomelanin [39]. Thus, melanin possesses both antioxidant and ROS-dependent cytotoxic properties [18].
Quinones and semiquinones which are intermediates of melanogenesis are toxic or mutagenic with the potential to cause cytogenetic instability [40]; and L-dopa, another intermediate of melanogenesis, has the capacity to inhibit the production of proinflammatory cytokines by T lymphocytes and monocytes, thus downregulating immune and inflammatory responses [38,40].
Melanins produced by melanocytes residing in the basal cell layer of the gingival epithelium have the capacity to neutralize ROS generated by dentogingival plaque-induced inflammation in the periodontal microenvironment [16]. Interestingly, a recent report shows that markers of gingival inflammation are reduced in subjects with pigmented gingiva compared to subjects with non-pigmented gingiva, despite comparable dentogingival plaque levels in both groups of subjects. However, one needs to bear in mind that the sulcular and junctional epithelium, in contrast to the keratinized epithelium usually does not harbour any melanocytes [16].
Melanosomes contain lysosomal enzymes including α-mannosidase, acid phosphatase, β-N acetylglycosaminidase, β-galactosidase, and acid lipase that can degrade bacteria [41]. Melanin itself can neutralise bacteria-derived enzymes and toxins, and since it has strong binding properties, it can also act as a physical barrier against microorganisms [41]. Furthermore, melanocytes can act as antigen presenting cells, can stimulate T-cell proliferation, and can phagocytose microorganisms [32,41]. Thus, melanocytes and their products have the capacity to inhibit proliferation of bacterial and fungal microorganisms [41].
As keratinocytes ascend through the cell layers of the epithelium and are shed, their melanosomal membranes undergo degradation with release of melanin ‘dust’ which becomes entangled in the keratin filaments of the desquamating surface cells. This melanin dust inactivates pathogenic chemicals, microbial toxins and other biologically active molecules [18]. Therefore, melanocytes and melanins may be viewed as an integral part of the innate immune system with a role in neutralising the products of superficial bacterial and fungal infective agents [41].
It is possible that from an evolutionary point of view, the primary role of melanocytes is not to produce melanin, for melanin does not confer a selective advantage to the organisms, but that melanin production is only a secondary specialization so melanocytes must have other more important functions to perform [13]. Melanocytes in skin, and perhaps in oral mucosa express genes encoding corticotropin releasing factor (CRF), proopiomelanocortin (POMC), ACTH, β endorphins, α-MSH and melanocortin-1 receptor (MC1R). These elements of the skin melanocortin system have the capacity to neutralize external noxious agents, to mediate local antimicrobial and immune responses, and to mediate local nociception [34,42]. UV radiation induces the generation of CRF, POMC and α-MSH by cutaneous keratinocytes and melanocytes, bringing about an increase in melanin biosynthesis [43], imparting protection against solar radiation [13].
Melanocyte-derived α-MSH, ACTH, and other peptides of POMC, stimulate the MC1R of neighbouring melanocytes, activating an intracellular signalling cascade involving the second messenger cAMP and the MITF [34]. The α-MSH/MC1R/cAMP/MITF pathway controls the transcription of tyrosine and is therefore essential for melanogenesis [25], and also determines the type and quantity of the melanin produced [1,13].
Regulation of melanogenesis
Melanogenesis comprises a process of oxidation of the amino acid tyrosine and derived aromatic compounds, forming two main types of polymeric phenolic compounds. Large, irregular granules of eumelanin which are black-brown in colour and smaller, more regular granules of pheomelanin which are yellow-red in colour [41]. Melanocytes are capable of producing both eumelanin and pheomelanin, and the proportion of the two melanins produced by a particular melanocyte is a function of availability of tyrosine, of reducing agents, and of the types of pigment enzymes expressed [1].
The melanocortin system through the cAMP/MITF pathway can stimulate proliferation of undifferentiated melanocytes, their maturation with fully formed dendritic processes, and upregulation of melanogenesis, thus increasing the formation of melanin [42,44]. α-MSH has the capacity to suppress inflammatory responses, because it can inhibit nuclear factor-κB (NF-κB) that regulates the expression of genes of proinflammatory cytokines. Thus, the same biological mediator of the melanocortin system that stimulates melanogenesis, also downregulates inflammatory responses [33].
It appears that the epidermal adrenergic signalling pathway plays a role in the regulation of skin pigmentation. Epidermal melanocytes express α1- and β2-adrenoceptors, the activation of which leads to an increase in melanin biosynthesis and a concurrent increase in the number and complexity of melanocytic dendrites. The adrenalin/β2-adrenoceptor/cAMP/MITF pathway like the α-MSH /MC1R/cAMP/MITF pathway has therefore the capacity to mediate melanin production and its dendritic distribution [44].
β-endorphin, an opioid peptide cleaved from POMC is positively associated with increased skin pigmentation. The β-endorphin/μ-opioid receptor/PKCβ isoform signalling pathway is expressed and functionally active, mediating differentiation and maturation of melanocytes with increased melanogenesis and dendricity [36,45]. To the best of our knowledge, the role of melanocortin, adrenergic and opioid systems in relation to melanogenesis in the oral epithelium is unknown.
Mediators of inflammation such as histamine and arachidonic acid metabolites trigger melanogenesis [41], and inflammatory cytokines such as TNF-α and IL-1α induce the secretion of melanogenic agents (SCF, HGF, bFGF, endothelins) by keratinocytes. Together these agents account for the melanin pigmentation sometimes observed in association with inflammatory conditions of skin or oral mucosa [41,46], such as oral lichen planus [6] and healing after periodontal surgery [47].
Physiological oral pigmentation
Variation in oral physiological melanin pigmentation is ascribable to variations in the activity of melanocytes in the basal cell layer of the oral epithelium. Such oral pigmentation is more common in darker-skinned persons regardless of their racial/ethnic origin [10,16]. This strongly suggests that physiological oral pigmentation is determined by genetic factors associated with melanogenesis [15]. Microscopical examination of physiologically pigmented oral mucosae shows increased melanin in the basal cell layer and sometimes in the upper portion of the lamina propria within melanophages, or simply as extracellular melanin particles [15]. These microscopical features are very similar to those found in melanotic maculae and smoker’s melanosis [3].
Physiological oral pigmentation and pathological oral pigmentations that may be similar in appearance should be differentiated. Diseases that may be confused with physiological oral pigmentation include Addison’s disease, neurofibromatosis, oral melanotic maculae, oral mucosal melanoma, drug-induced oral mucosal pigmentation, and to a much lesser extent Kaposi sarcoma, vascular malformations and haemangioma of the oral mucosa [48].
Physiological melanin pigmentation of the oral mucosa affects males and females equally, as asymptomatic, solitary or multiple brown maculae with well-defined borders (Figure 1) [49]. It may involve any part of the oral mucosa (Figures 2, 3, 4), but most frequently the gingiva [3,48,50]. In the gingivae, pigmentation when it occurs, is most commonly bilaterally symmetrical and does not transgress the mucogingival junction (Figure 2) [3,8,9,48,51], and in such cases sometimes the free gingiva is not pigmented (Figure 5) [3,7,48]. Occasionally, the alveolar mucosa is affected, and again the mucogingival junction is not transgressed (Figure 6). Oral pigmentation gradually appears during the first two decades of life [48] but affected subjects may be unaware of it [48,50].
Figure 1.

Multiple, light brown maculae with well-defined borders on the gingiva.
Figure 2.

Physiological oral pigmentation on the gingiva presenting as bilateral, symmetrical, dark brown discolouration of the labial gingiva, including the marginal and papillary gingiva but not transgressing the mucogingival junction.
Figure 3.
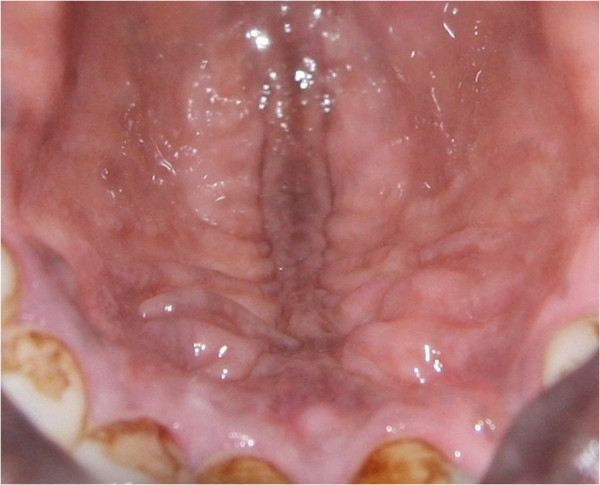
Diffuse, light brown physiological oral pigmentation on the hard palate.
Figure 4.

Uniform dark brown-black pigmentation of the lower lip and labial mucosa.
Figure 5.

Physiological oral pigmentation in a black male: a well demarcated brown band on the attached gingiva, not transgressing the mucogingival junction and not affecting the marginal/interdental papillary gingiva. (The foreign body at the upper left is a post-biopsy suture).
Figure 6.
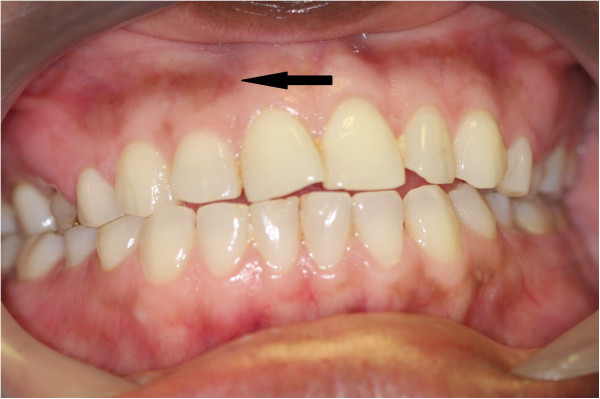
Light brown pigmentation of the alveolar mucosa not transgressing the mucogingival junction.
The extent and intensity of physiological oral pigmentation increases with age [52,53], concurrently with an increase in the number of oral melanocytes [15], perhaps because of the effects of potentially melanogenic stimuli such as recurrent minor functional injury, inflammatory conditions, medication, or tobacco smoke are cumulative [3].
The global frequency of physiological oral pigmentation in different racial/ethnic groups is unknown; but it has been reported that about 95% of healthy black persons in the United States are affected; that pigmentation is more extensive in the anterior than in the posterior portion of the mouth; and that when the gingiva is affected the buccal/labial surfaces are more intensely pigmented than the lingual/palatal surfaces [7]. It has been reported in one study from South Africa that oral pigmentation occurs in 98% of black persons [54].
Eighty to a hundred percent of Australian Aborigines have physiological oral pigmentation with equal gender distribution [52]. By contrast Fry and Almeyda [55] have reported that only about 5% of white people in England have physiological buccal mucosal pigmentation. In general, it appears that persons with more darkly pigmented skin are more likely to have physiological oral pigmentation [7].
Summary
Melanocytes can adjust and respond to biological, physical and chemical stimuli in their microenvironment, and can in turn generate physical and biochemical signals which may affect that microenvironment. Melanocytes have the capacity to mediate antimicrobial and immune responses and to act as neuroendocrine cells, and they produce melanin that provides protection from environmental stressors such as UV radiation, reactive oxygen species and free radicals. Why certain persons are more affected by physiological oral pigmentation is unknown, but certainly it occurs predominantly in darker skinned persons.
Consent
Written informed consent was obtained for the publication of this report and any accompanying images.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
Concept of paper was devised by LF. LF, AM, RAGK, MA, YJ, and JL wrote the manuscript. The manuscript was critically revised by LF, RAGK and JL. All authors read and approved the manuscript.
Contributor Information
Liviu Feller, Email: liviu.feller@ul.ac.za.
Aubrey Masilana, Email: graza@bucsmail.co.za.
Razia AG Khammissa, Email: razia.khammissa@ul.ac.za.
Mario Altini, Email: altini@iafrica.com.
Yusuf Jadwat, Email: yjadwat@gmail.com.
Johan Lemmer, Email: jbowman@iburst.co.za.
References
- Lin JY, Fisher DE. Melanocyte biology and skin pigmentation. Nature. 2007;445:843–850. doi: 10.1038/nature05660. [DOI] [PubMed] [Google Scholar]
- Pai A, Prasad S, Patil BA, Dyasanoor S, Hedge S. Oral pigmentation: case report and review of malignant melanoma with flow charts for diagnosis and treatment. Gen Dent. 2012;60:410–416. quiz 417-418. [PubMed] [Google Scholar]
- Meleti M, Vescovi P, Mooi WJ, van der Waal I. Pigmented lesions of the oral mucosa and perioral tissues: a flow-chart for the diagnosis and some recommendations for the management. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2008;105:606–616. doi: 10.1016/j.tripleo.2007.07.047. [DOI] [PubMed] [Google Scholar]
- Squier CA, Kremer MJ. Biology of oral mucosa and esophagus. J Natl Cancer Inst Monogr. 2001;29:7–15. doi: 10.1093/oxfordjournals.jncimonographs.a003443. [DOI] [PubMed] [Google Scholar]
- Barrett AW, Raja AM. The immunohistochemical identification of human oral mucosal melanocytes. Arch Oral Biol. 1997;42:77–81. doi: 10.1016/S0003-9969(96)00113-6. [DOI] [PubMed] [Google Scholar]
- Barrett AW, Scully C. Human oral mucosal melanocytes: a review. J Oral Pathol Med. 1994;23:97–103. doi: 10.1111/j.1600-0714.1994.tb01095.x. [DOI] [PubMed] [Google Scholar]
- Dummett CO. Physiologic pigmentation of the oral and cutaneous tissues in the Negro. J Dent Res. 1946;25:421–432. doi: 10.1177/00220345460250060201. [DOI] [PubMed] [Google Scholar]
- Chandra S, Keluskar V, Bagewadi A, Sah K. Extensive physiologic melanin pigmentation on the tongue: an unusual clinical presentation. Contemp Clin Dent. 2010;1:204–206. doi: 10.4103/0976-237X.73205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarthy PL, Shlkar G. In: Diseases of the Oral Mucosa. McCarthy PL, Shlkar G, editor. New York: McGraw-Hill; 1964. Distrubances in pigmentation; pp. 228–235. [Google Scholar]
- Amir E, Gorsky M, Buchner A, Sarnat H, Gat H. Physiologic pigmentation of the oral mucosa in Israeli children. Oral Surg Oral Med Oral Pathol. 1991;71:396–398. doi: 10.1016/0030-4220(91)90325-7. [DOI] [PubMed] [Google Scholar]
- Chi A, Valencia JC, Hu ZZ, Watabe H, Yamaguchi H, Mangini NJ, Huang H, Canfield VA, Cheng KC, Yang F, Abe R, Yamagishi S, Shabanowitz J, Hearing VJ, Wu C, Appella E, Hunt DF. Proteomic and bioinformatic characterization of the biogenesis and function of melanosomes. J Proteome Res. 2006;5:3135–3144. doi: 10.1021/pr060363j. [DOI] [PubMed] [Google Scholar]
- Marks MS, Seabra MC. The melanosome: membrane dynamics in black and white. Nat Rev Mol Cell Biol. 2001;2:738–748. doi: 10.1038/35096009. [DOI] [PubMed] [Google Scholar]
- Plonka PM, Passeron T, Brenner M, Tobin DJ, Shibahara S, Thomas A, Slominski A, Kadekaro AL, Hershkovitz D, Peters E, Nordlund JJ, Abdel-Malek Z, Takeda K, Paus R, Ortonne JP, Hearing VJ, Schallreuter KU. What are melanocytes really doing all day long…? Exp Dermatol. 2009;18:799–819. doi: 10.1111/j.1600-0625.2009.00912.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mihajlovic M, Vlajkovic S, Jovanovic P, Stefanovic V. Primary mucosal melanomas: a comprehensive review. Int J Clin Exp Pathol. 2012;5:739–753. [PMC free article] [PubMed] [Google Scholar]
- Eisen D. Disorders of pigmentation in the oral cavity. Clin Dermatol. 2000;18:579–587. doi: 10.1016/S0738-081X(00)00148-6. [DOI] [PubMed] [Google Scholar]
- Nilima S, Vandana KL. Melanin: a scavenger in gingival inflammation. Indian J Dent Res. 2011;22:38–43. doi: 10.4103/0970-9290.79973. [DOI] [PubMed] [Google Scholar]
- Thomas AJ, Erickson CA. The making of a melanocyte: the specification of melanoblasts from the neural crest. Pigment Cell Melanoma Res. 2008;21:598–610. doi: 10.1111/j.1755-148X.2008.00506.x. [DOI] [PubMed] [Google Scholar]
- Wood JM, Jimbow K, Boissy RE, Slominski A, Plonka PM, Slawinski J, Wortsman J, Tosk J. What’s the use of generating melanin? Exp Dermatol. 1999;8:153–164. doi: 10.1111/j.1600-0625.1999.tb00365.x. [DOI] [PubMed] [Google Scholar]
- Nishimura EK, Jordan SA, Oshima H, Yoshida H, Osawa M, Moriyama M, Jackson IJ, Barrandon Y, Miyachi Y, Nishikawa S. Dominant role of the niche in melanocyte stem-cell fate determination. Nature. 2002;416:854–860. doi: 10.1038/416854a. [DOI] [PubMed] [Google Scholar]
- Kumano K, Masuda S, Sata M, Saito T, Lee SY, Sakata-Yanagimoto M, Tomita T, Iwatsubo T, Natsugari H, Kurokawa M, Ogawa S, Chiba S. Both Notch1 and Notch2 contribute to the regulation of melanocyte homeostasis. Pigment Cell Melanoma Res. 2008;21:70–78. doi: 10.1111/j.1755-148X.2007.00423.x. [DOI] [PubMed] [Google Scholar]
- Cramer SF. Stem cells for epidermal melanocytes–a challenge for students of dermatopathology. Am J Dermatopathol. 2009;31:331–341. doi: 10.1097/DAD.0b013e31819cd0cb. [DOI] [PubMed] [Google Scholar]
- Aubin-Houzelstein G, Djian-Zaouche J, Bernex F, Gadin S, Delmas V, Larue L, Panthier JJ. Melanoblasts’ proper location and timed differentiation depend on Notch/RBP-J signaling in postnatal hair follicles. J Invest Dermatol. 2008;128:2686–2695. doi: 10.1038/jid.2008.120. [DOI] [PubMed] [Google Scholar]
- Shah KV, Chien AJ, Yee C, Moon RT. CTLA-4 is a direct target of Wnt/beta-catenin signaling and is expressed in human melanoma tumors. J Invest Dermatol. 2008;128:2870–2879. doi: 10.1038/jid.2008.170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlson JA, Murphy M, Slominski A, Vincent WL. In: Filed Cancerization: Basic Science and Clinical Applications. Dakubo GD E, editor. Ontario, Canada: Nova Science Publishers; 2011. Evidence of skin field cancerization; pp. 317–370. [Google Scholar]
- Schallreuter KU, Kothari S, Chavan B, Spencer JD. Regulation of melanogenesis–controversies and new concepts. Exp Dermatol. 2008;17:395–404. doi: 10.1111/j.1600-0625.2007.00675.x. [DOI] [PubMed] [Google Scholar]
- Neville BW, Damm DD, Allen CM, Bouquot JE. In: Oral and Maxillofacial Pathology. 3. Dolan J, editor. St Louis, Missouri: Saunders Elsevier; 2009. Acquired melanocytic nevus; p. 382. [Google Scholar]
- Yamaguchi Y, Passeron T, Watabe H, Yasumoto K, Rouzaud F, Hoashi T, Hearing VJ. The effects of dickkopf 1 on gene expression and Wnt signaling by melanocytes: mechanisms underlying its suppression of melanocyte function and proliferation. J Invest Dermatol. 2007;127:1217–1225. doi: 10.1038/sj.jid.5700629. [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y, Itami S, Watabe H, Yasumoto K, Abdel-Malek ZA, Kubo T, Rouzaud F, Tanemura A, Yoshikawa K, Hearing VJ. Mesenchymal-epithelial interactions in the skin: increased expression of dickkopf1 by palmoplantar fibroblasts inhibits melanocyte growth and differentiation. J Cell Biol. 2004;165:275–285. doi: 10.1083/jcb.200311122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paino F, Ricci G, De Rosa A, D’Aquino R, Laino L, Pirozzi G, Tirino V, Papaccio G. Ecto-mesenchymal stem cells from dental pulp are committed to differentiate into active melanocytes. Eur Cell Mater. 2010;20:295–305. doi: 10.22203/ecm.v020a24. [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y, Brenner M, Hearing VJ. The regulation of skin pigmentation. J Biol Chem. 2007;282:27557–27561. doi: 10.1074/jbc.R700026200. [DOI] [PubMed] [Google Scholar]
- Passeron T, Coelho SG, Miyamura Y, Takahashi K, Hearing VJ. Immunohistochemistry and in situ hybridization in the study of human skin melanocytes. Exp Dermatol. 2007;16:162–170. doi: 10.1111/j.1600-0625.2006.00538.x. [DOI] [PubMed] [Google Scholar]
- Tolleson WH. Human melanocyte biology, toxicology, and pathology. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2005;23:105–161. doi: 10.1080/10590500500234970. [DOI] [PubMed] [Google Scholar]
- Nordlund JJ. The melanocyte and the epidermal melanin unit: an expanded concept. Dermatol Clin. 2007;25:271–281. doi: 10.1016/j.det.2007.04.001. vii. [DOI] [PubMed] [Google Scholar]
- Slominski A, Wortsman J, Paus R, Elias PM, Tobin DJ, Feingold KR. Skin as an endocrine organ: implications for its function. Drug Discov Today Dis Mech. 2008;5:137–144. doi: 10.1016/j.ddmec.2008.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirobe T. Role of keratinocyte-derived factors involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. Pigment Cell Res. 2005;18:2–12. doi: 10.1111/j.1600-0749.2004.00198.x. [DOI] [PubMed] [Google Scholar]
- Slominski A, Tobin DJ, Shibahara S, Wortsman J. Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol Rev. 2004;84:1155–1228. doi: 10.1152/physrev.00044.2003. [DOI] [PubMed] [Google Scholar]
- Sturm RA. Molecular genetics of human pigmentation diversity. Hum Mol Genet. 2009;18:R9–R17. doi: 10.1093/hmg/ddp003. [DOI] [PubMed] [Google Scholar]
- Plonka PM, Grabacka M. Melanin synthesis in microorganisms–biotechnological and medical aspects. Acta Biochim Pol. 2006;53:429–443. [PubMed] [Google Scholar]
- Smit NP, van Nieuwpoort FA, Marrot L, Out C, Poorthuis B, van Pelt H, Meunier JR, Pavel S. Increased melanogenesis is a risk factor for oxidative DNA damage–study on cultured melanocytes and atypical nevus cells. Photochem Photobiol. 2008;84:550–555. doi: 10.1111/j.1751-1097.2007.00242.x. [DOI] [PubMed] [Google Scholar]
- Slominski A, Zbytek B, Slominski R. Inhibitors of melanogenesis increase toxicity of cyclophosphamide and lymphocytes against melanoma cells. Int J Canc Suppl J Int Canc Suppl. 2009;124:1470–1477. doi: 10.1002/ijc.24005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackintosh JA. The antimicrobial properties of melanocytes, melanosomes and melanin and the evolution of black skin. J Theor Biol. 2001;211:101–113. doi: 10.1006/jtbi.2001.2331. [DOI] [PubMed] [Google Scholar]
- Peters A. The self-similarity of the melanocortin system. Endocrinology. 2005;146:529–531. doi: 10.1210/en.2004-1475. [DOI] [PubMed] [Google Scholar]
- Millington GW. Proopiomelanocortin (POMC): the cutaneous roles of its melanocortin products and receptors. Clin Exp Dermatol. 2006;31:407–412. doi: 10.1111/j.1365-2230.2006.02128.x. [DOI] [PubMed] [Google Scholar]
- Grando SA, Pittelkow MR, Schallreuter KU. Adrenergic and cholinergic control in the biology of epidermis: physiological and clinical significance. J Invest Dermatol. 2006;123:1948–1965. doi: 10.1038/sj.jid.5700151. [DOI] [PubMed] [Google Scholar]
- Kauser S, Schallreuter KU, Thody AJ, Gummer C, Tobin DJ. Regulation of human epidermal melanocyte biology by beta-endorphin. J Invest Dermatol. 2003;120:1073–1080. doi: 10.1046/j.1523-1747.2003.12242.x. [DOI] [PubMed] [Google Scholar]
- Satomura K, Tokuyama R, Yamasaki Y, Yuasa T, Tatehara S, Ishimaru N, Hayashi Y, Nagayama M. Possible involvement of stem cell factor and endothelin-1 in the emergence of pigmented squamous cell carcinoma in oral mucosa. J Oral Pathol Med. 2007;36:621–624. doi: 10.1111/j.1600-0714.2007.00587.x. [DOI] [PubMed] [Google Scholar]
- Perlmutter S, Tal H. Repigmentation of the gingiva following surgical injury. J Periodontol. 1986;57:48–50. doi: 10.1902/jop.1986.57.1.48. [DOI] [PubMed] [Google Scholar]
- Kauzman A, Pavone M, Blanas N, Bradley G. Pigmented lesions of the oral cavity: review, differential diagnosis, and case presentations. J Can Dent Assoc. 2004;70:682–683. [PubMed] [Google Scholar]
- Caldeira PC, de Sousa SF, Gomez RS, Silva TA. Diffuse pigmentation of the oral mucosa. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2010;110:550–553. doi: 10.1016/j.tripleo.2010.06.005. [DOI] [PubMed] [Google Scholar]
- Lerman MA, Karimbux N, Guze KA, Woo SB. Pigmentation of the hard palate. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 2009;107:8–12. doi: 10.1016/j.tripleo.2008.07.022. [DOI] [PubMed] [Google Scholar]
- Gaeta GM, Satriano RA, Baroni A. Oral pigmented lesions. Clin Dermatol. 2002;20:286–288. doi: 10.1016/S0738-081X(02)00225-0. [DOI] [PubMed] [Google Scholar]
- Brown T. Oral pigmentation in the Aborigines of Kalumburu, North-West Australia. Arch Oral Biol. 1964;9:555–564. doi: 10.1016/0003-9969(64)90019-6. [DOI] [PubMed] [Google Scholar]
- Dummett CO, Barens G. Oromucosal pigmentation: an updated literary review. J Periodontol. 1971;42:726–736. doi: 10.1902/jop.1971.42.11.726. [DOI] [PubMed] [Google Scholar]
- van Wyk CW. Mouth pigmentation patterns in a group of healthy South African Bantu. S Afr Med J. 1970;44:177–180. [PubMed] [Google Scholar]
- Fry L, Almeyda JR. The incidence of buccal pigmentation in caucasoids and negroids in Britain. Br J Dermatol. 1968;80:244–247. doi: 10.1111/j.1365-2133.1968.tb11966.x. [DOI] [PubMed] [Google Scholar]