

Abstract
Objective
We aimed to investigate how losartan exerts protective effects on human umbilical vein endothelial cell injury induced by small, dense, LDL (sLDL) cholesterol particles.
Methods
sLDL cholesterol was isolated by a 2-steps method and the nuclear translocation and activation of nuclear factor-κB (NF-κB) in endothelial cells was observed by confocal microscopy and electrophoretic mobility shift assays.
Results
Losartan greatly inhibited the nuclear translocation of NF-κB induced by sLDL cholesterol in a dose-dependent manner.
Conclusions
sLDL cholesterol may be involved in endothelial dysfunction possibly through NF-κB activation; losartan protects against sLDL cholesterol-inducing endothelial cell injury by inhibiting NF-κB activation, suggesting that losartan may play a role in the prevention and treatment of cardiovascular disease.
Key Words: losartan; nuclear factor-κB; small, dense, LDL
Introduction
During the past 2 decades it has become evident that the vascular endothelium is an active paracrine, endocrine, and autocrine organ that is not only indispensable for the regulation of vascular tone and blood flow, but also for the maintenance of vascular homeostasis.1 When endothelial cells get damaged, they can trigger an inflammation response through secretion of inflammatory factors and regulation of the leukocyte adhesion surface, which then leads to the initiation and expansion of atherosclerosis (As).1 So, endothelial dysfunction is thought to be the key event and the most important risk factor in the process of As.
Endothelial dysfunction has many causes, such as hyperlipidemia, hypertension, diabetes, and smoking. Studies have shown that LDL cholesterol becomes oxidized in response to oxidative stress and results in the formation of oxidized LDL (ox-LDL) cholesterol. Ox-LDL cholesterol is a vital factor that can activate nuclear factor-κB (NF-κB) and increase expression of inflammatory factors, such as adhesion molecule and tissue factors, and then cause endothelial dysfunction.2,3 Small, dense LDL (sLDL) cholesterol is a subclass of LDL. In recent years, a large number of epidemiologic and clinical studies have shown the role elevated sLDL levels play in the initiation of As and that it is a risk factor in coronary heart disease.4,5
Angiotensin II (AngII), the major effector of the renin-angiotensin system, also contributes to endothelial dysfunction.6,7 Angiotensin receptor antagonists (ARBs) can antagonize the effect of AngII at the receptor level, and therefore have been widely used in clinical practice in the form of antihypertensive drugs. Large-scale clinical trials have indicated that losartan, a kind of ARB, can prevent As through down-regulating NF-κB, thus leading to the inhibition of oxygen free radicals, adhesion molecules, monocyte-macrophage chemokines, and stromal metalloproteinase.8–11 However, little research is done in China to investigate the mechanisms of how losartan inhibits As. Here, we report that losartan can inhibit nuclear translocation of NF-κB induced by sLDL cholesterol in human umbilical vein endothelial cells, adding to our understanding about the possible molecular mechanism for lipid disorders and the independent antihypertensive ways by which ARBs are involved in anti-As treatment.
Materials and Methods
Antibodies and reagents
Losartan used in our study was a gift from Xiuzheng Pharmaceutical Group Company Limited. Edathamil disodium (EDTA) was acquired from Sigma-Aldrich (Shanghai, China), whereas Dulbecco’s modification of Eagle's medium, Serum, total RNA extraction reagent, and Trypsin were procured from GIBCO Life Technology Company (Shanghai, China). Rabbit polyclonal antibodies against p65 were obtained from Santa Cruz Biotechnology (Dallas, Texas). Yahui Biomedical Engineering Company (Beijing, China) was the source for α-32P-deoxycytidine triphosphate.
Isolation and measurement of sLDL cholesterol
The 2-step ultracentrifugation method was used to extract and separate sLDL cholesterol as described previously.12 This method consists of 2 steps. The first step is to separate sLDL; the second step is to identify sLDL. Zhong Ke Mai Chen Technology Co Ltd (Beijing, China) was used to measure the protein concentration of sLDL cholesterol.
Human umbilical vein endothelial cell culture, identification, and treatment
Human umbilical vein endothelial cells were isolated from freshly obtained healthy maternal newborn umbilical cords and grown in tissue culture. Cultured endothelial cells were observed by phase contrast microscopy and identified for growth factor-related antigen. Cells, subcultured for 2–5 passages, were used for experiments. To examine the effect of losartan on sLDL cholesterol-induced NF-κB activation, cells were treated with either 100 μg/mL sLDL cholesterol or with different concentrations of losartan (10–7, 10–6, or 10–5 mol/L) plus 100 g/mL sLDL cholesterol. In the losartan interference experiment, cells were first incubated with losartan for 2 hours, and then sLDL cholesterol (100 g/mL) was added. Tumor necrosis factor-α (TNF-α) (200 U/mL) treatment was used as a positive control.
Preparation of nuclear extracts
Nuclear extracts were isolated as described previously.13 Briefly, cells were washed with ice-cold phosphate-buffered saline (PBS) and pelleted, and then the cell pellet was resuspended in a hypotonic buffer (10 mM N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid [HEPES], pH 7.9, 10 mM potassium chloride, 0.1 mM EDTA, 0.1 mM ethylene glycol tetraacetic acid [EGTA], 1 mM dithiothreitol [DTT], 0.5 mM phenylmethylsulfonyl fluoride [PMSF], 1 mM sodium fluoride, and 1 mM sodium orthovandate), incubated for 15 minutes on ice, and then lysed by the addition of 0.5% octylphenoxypolyethoxyethanol followed by vigorous vortexing for 10 seconds. The nuclei were pelleted and resuspended in extraction buffer (20 mM HEPES, pH 7.9, 400 mM sodium chloride, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 1 mM PMSF, 1 mM sodium fluoride, and 1 mM sodium orthovandate), and the tube was vigorously shaken at 4°C for 15 minutes on a shaking platform. The nuclear extracts were then centrifuged, and the supernatants were aliquoted and stored at –80°C. Protein concentrations can be determined according to the method of Bradford (BioRad, Veenendaal, the Netherlands).
Fluorescence confocal microscopy
Endothelial cells were treated with appropriate stimulus. After treatment for 24 hours, cells were fixed in 4% paraformaldehyde for 10 minutes followed by incubation in 0.5% polyethylene glycol octylphenol in PBS for 10 minutes for permeabilization, and were blocked with 10% goat serum in PBS for 1 hour. Cells were then incubated for 1 hour in 1:100 diluted solution of anti-p65-antibody in 10% goat serum followed by several washes and incubation in 1:100 solution of the secondary antibody fluorescein-goat-anti-rabbit immunoglobulin G for the detection of p65 protein. Washed cells were mounted in Vecta Shield (Vector Laboratories, Burlingame, California) containing 0.1 μg/mL 4′, 6-diamidino-2-phenylindole to stain nuclei. FV-REVIEW STATI10N software was used to determine the relative amount of p65 translocation.
Electrophoretic Mobility Shift Assay (EMSA)
EMSAs were performed by incubating 14 μg nuclear extract with 20 fmol of the 32P-end-labeled NF-κB consensus oligonucleotide 5′-CGCTTGATGAGTCAGCCGGAA-3′ (Promega, Madison, Wisconsin) for 15 minutes at 37°C. The incubation mixture included 2–3 μg poly-deoxy-inosinic-deoxy-cytidylic acid in a binding buffer (25 mM HEPES [pH 7.9], 0.5 mM EDTA, 0.5 mM DTT, 1% octylphenoxypolyethoxyethanol 5% glycerol, and 50 mM sodium chloride). The DNA-protein complex thus formed was separated from free oligonucleotide on 7.5% native polyacrylamide gel, using buffer containing 50 mM tris(hydroxymethyl)aminomethane, 200 mM glycine (pH 8.5), and 1 mM EDTA. The gel was dried, and the radioactive bands were visualized and quantified by a PhosphorImager (Molecular Dynamics, Sunnyvale, California), using ImageQuant software. The specificity of the NF-κB band was confirmed by using antibodies against p65 protein.
Statistics
SPSS version 10.0 software (1999, IBM-SPSS, Inc, Armonk, New York) was used to establish a database. Pairwise comparisons is done by using independent samples t test, and all experimental data are expressed as mean (SEM).
Results
Isolation and identification of sLDL cholesterol
The LDL cholesterol, which was obtained firstly by centrifugation, was then separated by ultracentrifugation from plasma. sLDL cholesterol was isolated (density = 1.044–1.066 g/mL), negatively stained, and observed. A transmission electron microscope was used to measure particle size and the result showed that all particles were <25.5 nm (Figure 1). Thus, our method is reliable and the sLDL cholesterol that was isolated with this method can be used for the following study.
Figure 1.

The small, dense, LDL (sLDL) cholesterol was isolated and observed by transmission electron microscope.
Losartan inhibits the nuclear translocation of NF-κB induced by sLDL cholesterol
To test the effect of losartan on the p65 translocation induced by sLDL cholesterol in endothelial cells, immunostaining of p65 was performed in the presence or absence of sLDL cholesterol. As show in Figure 2A, without sLDL cholesterol, p65 was mainly distributed in the cytoplasm. TNF-α treatment was used as a positive control. Under sLDL stimulation, p65 translocated to the nucleus. Compared with control cells, losartan can inhibit the translocation of p65 in dose-dependent fashion. Semiquantitative analysis was also performed (Figure 2B). Moreover, EMSA analysis confirmed our results (Figure 2C). These data support a notion that losartan inhibits the nuclear translocation of NF-κB induced by sLDL cholesterol in a dose-dependent manner.
Figure 2.
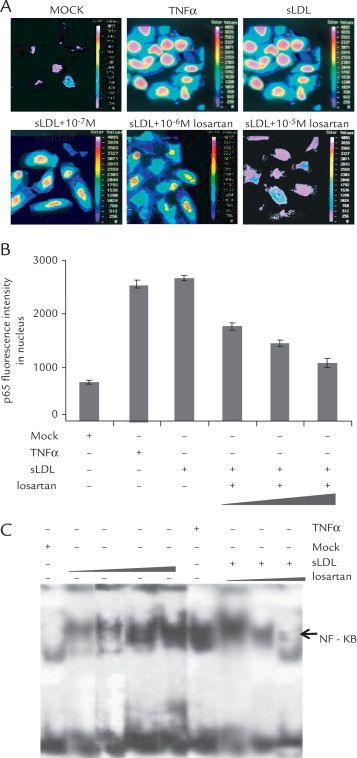
Losartan inhibits the nuclear translocation of nuclear factor-κB (NF-κB) induced by small, dense LDL (sLDL) cholesterol particles. (A) Subcellular localization of p65 under different stimulus. Endothelial cells were treated with 200 U/mL tumor necrosis factor-α (TNF-α), 100 μg/mL sLDL cholesterol, or 100 μg/mL sLDL cholesterol plus different concentrations of losartan (10–7 mol/L, 10–6 mol/L, or 10–5 mol/L). Twenty-four hours after incubation, rhodamine staining of p65 was visualized by fluorescence microscopy. (B) Semiquantitative analysis of p65 translocation. Endothelial cells were treated with 200 U/mL TNF-α, 100 μg/mL sLDL, or 100 μg/mL sLDL plus different concentrations of losartan (10–7 mol/L, 10–6 mol/L, or 10–5 mol/L). Twenty-four hours after incubation, subcellular distribution of p65 was scored in at least 10 cells. The results are means (SD) of 3 independent determinations. (C) Endothelial cells were exposed to different stimuli; nuclear extracts were made and analyzed with electrophoretic mobility shift assay as described in the text. A representative blot presented here shows that sLDL cholesterol (50 μg/mL, 100 μg/mL, and 150 μg/mL) activates NF-κB in a dose-dependent manner (lanes 2–5). Losartan (10–7 mol/L, 10–6 mol/L, and 10–5 mol/L) can inhibit the sLDL cholesterol-induced NF-κB activation in a dose-dependent manner (lanes 7–9). Treatment with 200 U/mL TNF-α was used a positive control (lane 6).
Discussion
The LDL cholesterol in plasma is made up of particles of unequal size and density. By density gradient ultracentrifugation or nondenaturing gradient gel method, the particles can be divided into many subclasses, mainly 3 types: low-density, larger size particles; intermediate-density, median size particles (LDL); and high-density, smaller size particles (sLDL). In recent years, a large number of epidemiologic and clinical studies have indicated that an increased proportion of sLDL cholesterol in total LDL cholesterol levels plays an important role in the occurrence of As, which is a risk factor for coronary heart disease.14–16 The generally acknowledged mechanisms that lead to As are the fact that sLDL cholesterol particles more readily permeate the inner arterial wall and deposit in the arterial intima, and that reducing acylneuraminate in the surface of sLDL cholesterol leads to the decrease of polyanion and more combination with proteoglycan in the arterial wall. Thus, sLDL cholesterol adheres to arterial walls and gives rise to the deposition of cholesterol; the spatial conformation of apolipoprotein B is not easily combined with the plasma LDL cholesterol receptor; therefore, the plasma LDL cholesterol receptor removal pathway is not efficient. Due to the reduction of free cholesterol in the lipid layer of sLDL cholesterol surface and increase of polyunsaturated acid, sLDL cholesterol can be easily oxidized and taken advantage by macrophages to form foam cells. Therefore, sLDL cholesterol is a pathogenic factor for As, and it will become 1 of the indicators that predicts As disease.
Because it is characterized by chronic inflammation and the hyperplasia process, As may be a NF-κB-mediated chronic inflammatory disease.17 It is reported that NF-κB is involved in the regulation of many intercellular gene expressions that are related to As (including the expression of vascular cell adhesion molecule-1 and intercellular adhesion molecule-1), promotes the local adhesion of monocytes and their migration to atherosclerotic injury predilection sites,18 and thus participates in the occurrence and development of As. Moreover, studies showed that ox-LDL increases expression of adhesion molecules, decreases NF-κB activity, inhibits the upregulation of adhesion molecules, prevents the formation of foam cells, and slows the progress of As.19,20 Therefore, all these studies suggest that NF-κB can regulate the expression of adhesion molecules and play an important role in the pathogenesis of As.
In our study, in the absence of losartan, NF-κB was significantly accumulated in the nucleus with sLDL cholesterol stimulation. In the presence of losartan, however, the sLDL-stimulated accumulation of NF-κB in the nucleus was greatly inhibited. This result was confirmed by EMSA. These experiments indicate that sLDL can be involved in the initiation and progression of atherosclerosis, possibly through releasing of inflammatory cytokines that were caused by activation of NF-κB, whereas losartan can prevent the progress of As by inhibiting the activation of NF-κB. sLDL has also been reported to affect human umbilical vein endothelial cells and enhance the activity of activated protein-1.16 In addition, AngII may also participate in the regulation of As-related genes, possibly through regulation of some important transcription factors, such as specificity protein-1 and activated protein-1, in human umbilical vein endothelial cells.8 Therefore, future investigations will need to address whether or not sLDL cholesterol influences the damage effect through other transcription factors and if losartan protects endothelial cells by other mechanisms.
Conflicts of Interests
The authors have indicated that they have no conflicts of interest regarding the content of this article.
Acknowledgments
Dr. Guo was in charge of the study performance. Dr. Cheng was in charge of the study design. Dr. Fu was in charge of the literature search and data collection.
Footnotes
This is an open-access article distributed under the terms of the Creative Commons Attribution-NonCommercial-No Derivative Works License, which permits non-commercial use, distribution, and reproduction in any medium, provided the original author and source are credited.
References
- 1.Tousoulis D., Kampoli A.M., Papageorgiou N. Pathophysiology of atherosclerosis: the role of inflammation. Curr Pharm Des. 2011;17:4089–4110. doi: 10.2174/138161211798764843. [DOI] [PubMed] [Google Scholar]
- 2.Leonarduzzi G., Gamba P., Gargiulo S. Inflammation-related gene expression by lipid oxidation-derived products in the progression of atherosclerosis. Free Radic Biol Med. 2012;52:19–34. doi: 10.1016/j.freeradbiomed.2011.09.031. [DOI] [PubMed] [Google Scholar]
- 3.Mitra S., Goyal T., Mehta J.L. Oxidized LDL, LOX-1 and atherosclerosis. Cardiovasc Drugs Ther. 2011;25:419–429. doi: 10.1007/s10557-011-6341-5. [DOI] [PubMed] [Google Scholar]
- 4.Alabakovska S.B., Tosheska K.N., Spiroski M.Z. Low density lipoprotein size in relation to carotid intima-media thickness in coronary artery disease. Bratisl Lek Listy. 2012;113:87–91. doi: 10.4149/bll_2012_020. [DOI] [PubMed] [Google Scholar]
- 5.Maeda S., Nakanishi S., Yoneda M. Associations between small dense LDL, HDL subfractions (HDL2, HDL3) and risk of atherosclerosis in Japanese-Americans. J Atheroscler Thromb. 2012;19:444–452. doi: 10.5551/jat.11445. [DOI] [PubMed] [Google Scholar]
- 6.Hirata Y., Fukuda D., Sata M. Critical role of renin-angiotensin system in the pathogenesis of atherosclerosis [in Japanese] Nihon Rinsho. 2011;69:55–59. [PubMed] [Google Scholar]
- 7.Sata M., Fukuda D. Crucial role of renin-angiotensin system in the pathogenesis of atherosclerosis. J Med Invest. 2010;57:12–25. doi: 10.2152/jmi.57.12. [DOI] [PubMed] [Google Scholar]
- 8.Aoyama T., Minatoguchi S. The effect of ARB on prevention of atherosclerosis [in Japanese] Nihon Rinsho. 2011;69:92–99. [PubMed] [Google Scholar]
- 9.Hirohata A., Yamamoto K., Miyoshi T. Impact of olmesartan on progression of coronary atherosclerosis a serial volumetric intravascular ultrasound analysis from the OLIVUS (impact of OLmesarten on progression of coronary atherosclerosis: evaluation by intravascular ultrasound) trial. J Am Coll Cardiol. 2010;55:976–982. doi: 10.1016/j.jacc.2009.09.062. [DOI] [PubMed] [Google Scholar]
- 10.Fujino M., Miura S., Kiya Y. A small difference in the molecular structure of angiotensin II receptor blockers induces AT(1) receptor-dependent and -independent beneficial effects. Hypertens Res. 2010;33:1044–1052. doi: 10.1038/hr.2010.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Negro R. Endothelial effects of antihypertensive treatment: focus on irbesartan. Vasc Health Risk Manag. 2008;4:89–101. doi: 10.2147/vhrm.2008.04.01.89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sun G., Wang Y., Sun L. Clinical significance of Hiwi gene expression in gliomas. Brain Res. 2011;1373:183–188. doi: 10.1016/j.brainres.2010.11.097. [DOI] [PubMed] [Google Scholar]
- 13.Timchenko N.A., Wilde M., Nakanishi M. CCAAT/enhancer-binding protein alpha (C/EBP alpha) inhibits cell proliferation through the p21 (WAF-1/CIP-1/SDI-1) protein. Genes Dev. 1996;10:804–815. doi: 10.1101/gad.10.7.804. [DOI] [PubMed] [Google Scholar]
- 14.Koba S., Yokota Y., Hirano T. Small LDL-cholesterol is superior to LDL-cholesterol for determining severe coronary atherosclerosis. J Atheroscler Thromb. 2008;15:250–260. doi: 10.5551/jat.e572. [DOI] [PubMed] [Google Scholar]
- 15.Norata G.D., Raselli S., Grigore L. Small dense LDL and VLDL predict common carotid artery IMT and elicit an inflammatory response in peripheral blood mononuclear and endothelial cells. Atherosclerosis. 2009;206:556–562. doi: 10.1016/j.atherosclerosis.2009.03.017. [DOI] [PubMed] [Google Scholar]
- 16.Toft-Petersen A.P., Tilsted H.H., Aaroe J. Small dense LDL particles—a predictor of coronary artery disease evaluated by invasive and CT-based techniques: a case-control study. Lipids Health Dis. 2011;10:21. doi: 10.1186/1476-511X-10-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Demer L., Tintut Y. The roles of lipid oxidation products and receptor activator of nuclear factor-kappaB signaling in atherosclerotic calcification. Circ Res. 2011;108:1482–1493. doi: 10.1161/CIRCRESAHA.110.234245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pamukcu B., Lip G.Y., Shantsila E. The nuclear factor-kappa B pathway in atherosclerosis: a potential therapeutic target for atherothrombotic vascular disease. Thromb Res. 2011;128:117–123. doi: 10.1016/j.thromres.2011.03.025. [DOI] [PubMed] [Google Scholar]
- 19.Lei L., Li H., Yan F. Porphyromonas gingivalis lipopolysaccharide alters atherosclerotic-related gene expression in oxidized low-density-lipoprotein-induced macrophages and foam cells. J Periodontal Res. 2011;46:427–437. doi: 10.1111/j.1600-0765.2011.01356.x. [DOI] [PubMed] [Google Scholar]
- 20.Wiesner P., Choi S.H., Almazan F. Low doses of lipopolysaccharide and minimally oxidized low-density lipoprotein cooperatively activate macrophages via nuclear factor kappa B and activator protein-1: possible mechanism for acceleration of atherosclerosis by subclinical endotoxemia. Circ Res. 2010;107:56–65. doi: 10.1161/CIRCRESAHA.110.218420. [DOI] [PMC free article] [PubMed] [Google Scholar]