

Abstract
Liver X receptors (LXRα and LXRβ) are key transcription factors in cholesterol metabolism that regulate cholesterol biosynthesis/efflux and bile acid metabolism/excretion in the liver and numerous organs. In macrophages, LXR signaling modulates cholesterol handling and the inflammatory response, pathways involved in atherosclerosis. Since regulatory pathways of LXR transcription control are well understood, in the present study we aimed at identifying post-transcriptional regulators of LXR activity. MicroRNAs (miRs) are such post-transcriptional regulators of genes that in the canonical pathway mediate mRNA inactivation. In silico analysis identified miR-206 as a putative regulator of LXRα but not LXRβ. Indeed, as recently shown, we found that miR-206 represses LXRα activity and expression of LXRα and its target genes in hepatic cells. Interestingly, miR-206 regulates LXRα differently in macrophages. Stably overexpressing miR-206 in THP-1 human macrophages revealed an up-regulation and miR-206 knockdown led to a down-regulation of LXRα and its target genes. In support of these results, bone marrow-derived macrophages (BMDMs) from miR-206 KO mice also exhibited lower expression of LXRα target genes. The physiological relevance of these findings was proven by gain- and loss-of-function of miR-206; overexpression of miR-206 enhanced cholesterol efflux in human macrophages and knocking out miR-206 decreased cholesterol efflux from MPMs. Moreover, we show that miR-206 expression in macrophages is repressed by LXRα activation, while oxidized LDL and inflammatory stimuli profoundly induced miR-206 expression. We therefore propose a feed-back loop between miR-206 and LXRα that might be part of an LXR auto-regulatory mechanism to fine tune LXR activity.
Abbreviations: miR, Micro-RNA; LXRs, liver X receptors; ApoE, apolipoprotein E; ABCs, ATP-binding cassette transporters; KO, knockout; SREBP, sterol regulatory element-binding protein
Keywords: Micro-RNA, ox-LDL, LXR target gene, ABC, ApoA-I, HDL
Graphical abstract
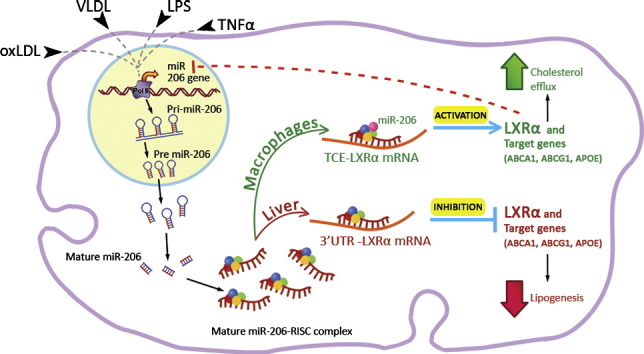
Highlights
-
•
Functional differences of miR-206 in the liver and macrophages
-
•
In the liver, miR-206 suppresses LXRα expression and signaling.
-
•
In macrophages, miR-206 increases LXRα abundance and promotes cholesterol efflux.
-
•
In macrophages, LXRα activation represses miR-206 expression.
-
•
In macrophages, pro-inflammatory stimuli increase miR-206 expression.
1. Introduction
Whole body metabolism and the cellular homeostasis of cholesterol are complex and governed by de novo biosynthesis, cholesterol absorption and cholesterol elimination [1]. In dyslipidemia, which predisposes to atherosclerosis, the balance between cholesterol uptake, synthesis and cellular export is disrupted [2,3]. Cholesterol homeostasis is intricately regulated by a battery of transcription factors among which liver X receptors (LXRs) are nuclear receptors that play a crucial role in transcriptional regulation of lipid metabolism and inflammation [1]. LXRβ is ubiquitously expressed whereas LXRα is expressed in all metabolically active tissues such as the liver, adipose tissue, kidney, intestine, and in cells such as the macrophages, lung and adrenal glands. LXRs are “cholesterol sensors” activated by elevated intracellular oxysterols and intermediates of the cholesterol biosynthetic pathway, which in turn transcriptionally regulate genes that are involved in reverse cholesterol transport, cholesterol conversion to bile acids and intestinal absorption [2].
In macrophages, LXRs regulate several genes involved in cholesterol transport and lipid metabolism such as the ATP-binding cassette transporters (ABCs) ABCA1 and ABCG1, apolipoprotein E (ApoE), lipoprotein lipase (LPL) and phospholipid transfer protein (PLTP). Activation of LXR in macrophages promotes cholesterol efflux and protects against development of atherosclerosis. An E3 ubiquitin ligase (IDOL) which is a direct LXRα target gene has been shown to reduce cholesterol accumulation in cells by post-translationally degrading low-density lipoprotein receptor (LDLR), very low-density lipoprotein receptor (VLDLR) and ApoE receptor 2 (ApoER2) [4–6]. In addition to its role in cholesterol homeostasis LXRα has been implicated as regulator of inflammation and innate immunity. Recently, LXRα has also been reported to be atheroprotective by other mechanisms such as in the control of iron homeostasis, in which it enhances iron recycling capabilities by increasing iron release from macrophages [7].
Though athero-protective, the major drawback of LXR agonists as potential drugs to treat cardiovascular diseases is their stimulating effect on hepatic lipogenesis and increase of plasma triglyceride levels thereby promoting hepato-steatosis [8]. Hence a better understanding of the mechanisms controlling tissue specific functions of LXR is of immense interest [9,10].
MicroRNAs (miRs) are important post-transcriptional regulators of gene expression that play crucial roles in numerous biological pathways. miRs are 20–25 nt small non-coding RNA molecules that control gene expression through base pairing between the seed sequence of the miR and its complementary match sequence on the target mRNA [11]. miRs play important roles in the physiology and pathophysiology of cell function, differentiation and growth [12,13]. There are more than 70 human diseases that find direct correlation to the differential expression of a single tissue specific miR or a specific family of miRs [12,13].
In this paper we report a novel miR-206 mediated regulation of macrophage LXRα gene expression in both human and mouse macrophages. Interestingly, we observe that this regulation occurs differently in the liver. We also observe a reciprocal feed-back repression of miR-206 expression in response to LXRα activation in macrophages.
2. Methods
2.1. Animals
All animal experiments were carried out in accordance with the guidelines of the Division of Genetic Engineering and Animal Experiments, Austrian Federal Ministry of Science and Research (Vienna, Austria). miR-206 knockout (KO) mice were obtained from Dr. Eric N. Olson's laboratory (Department of Molecular Biology, University of Texas Southwestern Medical Center, TX). WT mice and miR-206 KO mice were on a mixed background (29SvEv–C57BL/6). All animals were fed ad libitum with normal chow diet (caloric intake = 11.9% from fat; Ssniff®, Soest, Germany) and were maintained in a 12 h light/dark cycle in a temperature controlled environment.
2.2. Cell lines and cell experiments
HepG2 and COS-7 cells used in this study were obtained from American Type Culture Collection (ATCC) and were grown in DMEM supplemented with 10% fetal bovine serum and 1% pen–strep (penicillin–streptomycin). THP-1 cells were maintained in RPMI media (Gibco; Invitrogen, CA, USA) supplemented with 10% fetal bovine serum and 1% pen–strep. For differentiating THP-1 cells, the cells were treated with 100 nM phorbol 12-myristate 13-acetate (PMA; Sigma# P 8139, St. Louis, USA) for 3 days.
Mouse bone marrow-derived macrophages (BMDMs) and peritoneal macrophages (MPMs) were prepared as described earlier [14]. Human peripheral blood mononuclear cells were isolated from normolipidemic individuals and were differentiated into monocyte-derived macrophages (MDMs) as previously described [7].
THP-1 cells were starved for 12 h and treated with LXR agonists TO-901317 (TO9, 1–10 μM) and GW3965 (GW, 1 μM) for 24 h. Alternatively, cells were incubated for 24 h with the given amounts of acLDL, oxLDL or VLDL. Lipoproteins were prepared as described [15,16].
2.2.1. LPS treatment and TNFα treatment
PMA-treated THP-1 cells were serum starved for 12 h and treated with 10 ng/ml TNFα (PeproTech, NJ). Differentiated MDMs were treated with 100 ng/ml LPS for 24 h. For details see Supplementary material.
2.3. Computational prediction
The microRNA that post-transcriptionally regulated LXR was identified using publicly available prediction software suite, TargetScan (http://www.targetscan.org/). The software suite, RNAhybrid (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/) was used further to identify the microRNA that had the most favorable binding energy to the LXR mRNA. For predicting microRNA promoter interaction MicroPIR prediction software was used.
2.4. miR-206 overexpression or knockout
miR-206 was expressed in HepG2 cells by transfection with 100 nM of synthetic precursor miR-206 (Ambion®, TX, USA) using Metafectene PRO. The transfection was carried out as per the manufacturer's instruction.
For miR-206 overexpression or knockout in THP-1 cells, pCDH-CMV-MCS-EF1-copGFP cDNA cloning and expression vector (SBI# CD511B-1, PA, USA) or miRzip lentiviral vector (SBI# MZIPpa/aa-1) was co-transfected with packaging vectors VSVG and PCMV (Clontech, CA, USA) into HEK293T cells using the standard calcium phosphate method. Viruses were harvested by centrifuging the supernatant at 2000 g for 20 min at 4 °C. The supernatant was filtered and purified further and the viral stocks were aliquoted and maintained at − 80 °C until use. THP-1 cells were transfected using the viral particles and GFP positive cells were sorted out using FACSCalibur (BD Biosciences, New Jersey, USA).
2.5. Protein extraction, Western blot, RNA extraction and quantitative real time PCR analyses
Protein extraction and Western blot analysis were carried out as described [17]. Details of the antibody dilutions used for Western blot are as follows: LXRα (1:500 dilution, #ab41902, Abcam, Cambridge, UK), ABCA1 (1:1000, #NB400-105, Novus Biologicals, Cambridge, UK), GAPDH (1:1000, #sc-365062, Santa Cruz Biotechnology, TX, USA), and Calnexin (1:1000, #sc-11397, Santa Cruz Biotechnology, TX, USA).
RNA extraction and quantitative real time PCR are described in detail in the Supplementary material. In short, total RNA was extracted with Trifast (Peqlab, Erlangen, Germany) and reverse transcribed (High Capacity cDNA Reverse Transcription Kit, Applied Biosystems, Carlsbad, CA). The quantity of RNA used in quantitative real time PCR was optimized prior to obtain ct values within the range of 18–30.
For mature miR-206 quantification the miRCURY LNA™ Universal RT microRNA PCR system (Exiqon, Vedbaek, Denmark) was used together with miR-206 or U6 (reference gene) specific predesigned LNA primers. Quantitative real time PCR was carried out using gene specific primers and QuantiFast SYBR Green PCR Kit (Qiagen, Germany) on LC480 (Roche Diagnostics, Basel, Switzerland). All samples were normalized to cyclophilin A mRNA expression. Experiments were carried out in triplicates.
2.6. Luciferase reporter assays
Details of these experiments are given in the Supplementary material. In short, a 52 base pair putative miR-206 target sequence from the 3′UTR region of LXR, or a scrambled negative control was cloned using PremiR-Reporter constructs and synthetic Pre-miR206 from Ambion and the expression plasmid for miR206 from SBI (System Biosciences, PA, USA).
For the luciferase assay, 100 ng/ml of the PremiR-Reporter construct was co-transfected with 25 ng/ml β-Gal control plasmid and also with the 100 nM synthetic precursor-miR206 strand or the 1 μg/ml of miR-206 expression vector into COS-7 cells using X-tremeGENE siRNA Transfection Kit (Roche). After 36 h, cells were lysed and 50 μl of the lysate was used for β-Gal assay and 10 μl (2 ×) was used for luciferase assay carried out using the manufacturer's protocol.
2.7. Cholesterol efflux assay
Cholesterol efflux assay was performed in MPMs after loading with [3H]cholesterol labeled ac-LDL as described earlier [15]. Cholesterol efflux assay in THP-1 cells was performed as described in the Supplementary methods section.
2.8. Statistics
All statistical analyses were performed using GraphPad Prism 5.0. In cholesterol efflux assays the differences between groups were analyzed using two-tailed, unpaired Student's t-test. All qRT-PCR experiments were calculated using REST2009.
3. Results
3.1. miR-206 targets the conserved 3′UTR of LXRα in humans and several mammals
To identify post-transcriptional regulators of LXRα, we performed in silico analysis of LXRα mRNA sequence using the TargetScan microRNA prediction tool. TargetScan analysis revealed miR-206 to be a putative miR that potentially targets the 3′UTR of LXRα mRNA. Fig. 1a shows the stem miR-206 loop structure, the mature miR-206 sequence and the conserved binding site of miR-206 on the LXRα 3′UTR of human and several mammals.
Fig. 1.

a) Conserved binding site for miR-206 on LXRα mRNA. Top: Stem loop structure of miR-206 from which the mature miR-206 is formed. The seed sequence of miR-206 that can putatively bind to the LXRα 3′UTR region is represented in pink and the conserved miR-206 binding site on LXRα mRNA is highlighted in blue.
b) Luciferase reporter assay: Relative luciferase activity of the LXR 3′UTR construct and the 3′UTR scrambled construct. Mean values ± SEM from three separate experiments are as indicated;***p < 0.001.
We further examined the feasibility of the predicted interaction using the RNAhybrid program that predicts the minimum free binding energy (mfe) of miR206–LXRα interaction. This analysis revealed that the interaction between miR206–LXRα has a favorable mfe of (−) 23.5 kcal/mol. LXRβ lacked miR-206 binding sites and therefore was not among the predicted miR-206 targets.
To determine whether the predicted miR-206 binding site on LXRα 3′UTR was functional, we cloned the 3′UTR region of LXRα (LXR 3′UTR clone) or a scrambled seed sequence clone of the 3′UTR LXRα (scrambled control) into a miR-luciferase reporter construct. As shown in Fig. 1b, luciferase activity of the LXRα 3′UTR clone was markedly reduced in COS7 cells transfected with a miR-206 expression plasmid whereas the cells transfected with the scrambled control plus the miR-206 expression plasmid showed no reduction in the luciferase activity. As indicated in Fig. 1b, the basal luciferase activities of both the LXR 3′UTR clone and the scrambled control were comparable.
3.2. miR-206 represses mRNA expression of LXRα and its target genes in HepG2 cells
While this study was in progress, Zhong et al. reported that miR-206 reduces LXRα signaling in mouse primary hepatocytes and in human HepG2 cells [18]. In order to confirm and extend these observations, we studied the expression of LXRα and its target genes in HepG2 cells transfected with pre-miR-206. Supplementary Fig. S2.1a shows that in miR-206 overexpressing HepG2 cells treated with the LXRα agonist TO901317 (TO9, 10 μM), LXRα expression was reduced by 50%, and the expression of the target genes ABCA1, ABCG1, SREBP-1c and ABCG5 was reduced by 75–85%. ABCG8 expression was not significantly affected. LXRα protein expression in TO9-activated miR-206 expressing cells was also reduced by up to 50% (Supplementary Fig. S2.1b).
3.3. The expression of LXRα and its target genes is increased in miR206 KO liver
In order to substantiate the physiological relevance of these findings, LXRα mediated pathways were investigated in miR-206 KO mice. In Fig. 2a we show, that LXRα protein expression in miR-206 KO liver is markedly increased by up to 75%. In addition, we noticed a significant 1.5–5 fold increase in the LXRα target gene mRNA expression (ABCA1, ABCG1, ApoE and SREBP-1c) in the liver of miR206 KO mice (Fig. 2b).
Fig. 2.

Effect of miR-206 knockout on mRNA and protein expression of LXRα and its target genes in the liver. a) Western blot of LXRα from the liver of miR-206 KO and WT mice fed normal diet (n = 6). b) mRNA abundance of LXRα and its target genes in WT and in miR 206 KO liver by qRT-PCR (n = 4). Mean values ± SEM from three separate experiments are indicated;*p < 0.05; **p < 0.01; ***p < 0.001.
In summary, we show that miR-206 directly interacts with the 3′UTR region of LXRα and represses its translation in HepG2 cells and in mouse liver and thereby interferes with LXRα mediated signaling pathways.
3.4. miR-206 regulates LXRα expression differently in the liver and in human macrophages
As LXRα plays a crucial role in regulating cholesterol efflux and homeostasis in macrophages [9], we were further interested to study the effect of miR-206 in macrophage cholesterol homeostasis. For this purpose, we stably overexpressed (THP-1miR206 OE) or knocked down (THP-1miR206 KO) miR-206 in the human macrophage cell line THP-1 using lentiviral transduction. The relative expressions of miR-206 and of ABCA-1 in these cells are shown in Supplementary Fig. S2.2. Surprisingly, Western blot analysis from THP-1miR206 OE cells revealed that LXRα protein expression was markedly induced in untreated as well as in LXR agonist-treated cells compared to control THP-1 cells. In contrast, though not significant both basal as well as the TO9 or GW-induced LXRα protein expressions were moderately reduced in THP-1miR206 KO when compared to control THP-1 cells (Fig. 3a). In THP-1miR206 KO, TO9 or GW-induced LXRα protein expression is remarkably lower when compared to LXRα protein expression in THP-1miR206 OE cells.
Fig. 3.

Regulation of LXRα expression and its target genes by miR-206 in THP-1 cells. After differentiation, protein and mRNA expressions of LXRα and its target genes were determined in control THP-1, THP-1miR206 OE and THP-1miR206 KO cells. a) Western blot of basal or TO9 or GW-induced LXRα protein from control THP-1, THP-1miR206 KO and THP-1miR206 OE cells. Western blots are from one representative experiment out of four experiments (t test: THP-1miR206 OE vs. control and TO9 or GW-induced expression of LXRα in THP-1miR206 OE vs. control THP-1 cells; $p < 0.05; THP-1miR206 OE vs. THP-1miR206 KO and TO9 or GW-induced expression of LXRα in THP-1miR206 OE vs. THP-1miR206 KO; #p < 0.05; untreated control vs. TO9 or GW-induced expression of LXRα in THP-1, THP-1miR2-6-KO and THP-1miR206-OE cells; *p < 0.05; ***p < 0.001). b) Basal mRNA abundance of LXRα target genes in THP-1 cells by qRT-PCR. c) TO9-induced mRNA abundance of LXRα target genes by qRT-PCR. Mean values ± SEM from three separate experiments are indicated; *p < 0.05; ***p < 0.001.
qRT-PCR gene expression profiling of several LXRα target genes revealed that basal mRNA levels of ABCA1, ABCG1, ApoE and SREBP-1c were increased by 3–18 fold in THP-1miR206 OE cells (Fig. 3b), whereas LXRα target genes were less expressed in miR-206 KO THP-1 cells (Fig. 3b), compared to control THP-1 cells. Comparable upregulation in THP-1miR206 OE cells and downregulation in THP-1miR206 KO cells were observed after treatment with TO9 or GW (Fig. 3c and Supplementary Fig. S2.3).
3.5. LXRα target gene expression is reduced in miR-206 KO murine macrophages
To examine whether this miR206–LXRα regulatory pathway is also operative in murine systems, we studied MPMs and BMDMs obtained from miR-206 KO and WT mice. Gene expression profiling of miR206 KO MPMs revealed that under basal conditions, the expression of LXRα target genes ABCA1, ABCG1, ApoE and SREBP-1c was about 40–80% lower compared to WT MPMs (Fig. 4a). Comparable results were also observed after treatment of MPMs with GW (Fig. 4b), yet SREBP-1c expression was affected to a much greater extent as compared to untreated WT MPMs (90% vs. 55%, Fig. 4a and b). TO9 treatment had lower effect on LXRα target gene expression as compared to GW treatment (Fig. 4b, c).
Fig. 4.

Effect of miR-206 knockout on mRNA LXRα target genes in untreated and LXRα ligand treated MPMs. a) Basal mRNA abundance of LXRα target genes in miR206 KO and WT MPMs by qRT-PCR. b) TO9-induced mRNA abundance of LXRα and its target genes in miR206 KO and WT MPMs by qRT-PCR. c) GW-induced mRNA abundance of LXRα and its target genes in miR206 KO and WT MPMs by qRT-PCR. Mean values ± SEM from three separate experiments are indicated; *p < 0.05; **p < 0.01; ***p < 0.001.
Similar experiments were performed in BMDMs (Fig. 5). Here, ABCA1 expression was not significantly different between WT BMDMs and miR206 KO BMDMs neither under basal conditions nor in the presence of the LXRα ligands TO9 or GW. ABCG1 and SREBP-1c expressions were lower in miR206 KO BMDMs under basal conditions and also after TO9 treatment and GW treatment, and ApoE expression was lower under basal conditions and after TO9 treatment (Fig. 5a–c). Notably, the expression of ABCG1, ApoE and SREBP-1c was affected in miR-206 KO MPMs to a greater extent than in miR206 KO BMDMs.
Fig. 5.

Effect of miR-206 knockout on mRNA LXRα target genes in untreated and LXRα treated BMDMs. a) Basal mRNA abundance of LXRα target genes in miR206 KO and WT BMDMs by qRT-PCR. b) TO9-induced mRNA abundance of LXRα and its target genes in miR206 KO and WT BMDMs by qRT-PCR. c) GW-induced mRNA abundance of LXRα and its target genes in miR206 KO and WT BMDMs by qRT-PCR. Mean values ± SEM from three separate experiments are indicated; *p < 0.05; **p < 0.01; ***p < 0.001.
3.6. miR-206 overexpression in THP-1 cells increases cholesterol efflux to HDL and human serum
To investigate whether the increase in LXRα and its target genes has functional relevance on human macrophages, we performed cholesterol efflux assays in THP-1miR206 OE, THP-1miR206 KO and control THP-1 cells. Briefly, cells were incubated with [3H]cholesterol for 24 h, and the efflux to ApoA-I, HDL (ApoB depleted serum) and human serum was measured after stimulation with TO9. Fig. 6a demonstrates that all three cell lines effluxed cholesterol efficiently to ApoA-I, human serum and HDL. However, THP-1miR206 overexpression showed significantly enhanced cholesterol efflux capacity to human serum when compared to control THP-1 and THP-1miR206 KO cells. Though not significant, THP-1miR206 OE showed an increasing tendency to efflux cholesterol to HDL when compared to control THP-1 cells. In contrast to THP-1miR206 OE, THP-1miR206 KO showed a significantly lower cholesterol efflux to HDL when compared to control THP-1miR206 OE cells. Cholesterol efflux to ApoA-I and to BSA (control) was not significantly affected by miR-206 overexpression or knockout under these experimental conditions (Fig. 6a).
Fig. 6.

Cholesterol efflux assay in THP-1, THP-1miR206 OE and THP-1miR206 KO cells. Control THP-1 cells, THP-1miR206 OE and THP-1miR206 KO were loaded with [3H]cholesterol for 24 h and cholesterol efflux to human serum, ApoB depleted serum (HDL), and ApoA-I was determined. a) Percentage of cholesterol efflux to ApoA-I, HDL and human serum was calculated relative to the unspecific cholesterol efflux to BSA. Statistical significance is indicated (t test: THP-1miR206 OE vs. control and THP-1miR206 OE vs. THP-1miR206 KO efflux to human serum; #p < 0.05; THP-1miR206 OE vs. THP-1miR206 KO efflux to HDL; $p < 0.05; efflux to ApoA-I, HDL and human serum compared to control-BSA; *p < 0.05; **p < 0.01; ***p < 0.001). b) Cholesterol efflux assay from WT and miR206KO MPMs to ApoA-I, HDL and whole human serum: MPMs were isolated and loaded with 3H-acLDL for 24 h and the cholesterol efflux assay was performed. Results are mean ± SEM of triplicate experiments. Statistical significance is indicated (t test; *p < 0.05; ***p < 0.001).
Collectively, these data indicate that the miR206–LXRα pathway is functional and of potential physiological relevance in facilitating cholesterol efflux from human macrophages.
3.7. miR206 KO mouse macrophages display reduced cholesterol efflux to HDL and ApoA-I
In order to substantiate these findings and to examine the physiological relevance of the miR206–LXRα pathway in the murine system, cholesterol efflux was also investigated in MPMs from miR206 KO and WT mice. Fig. 6b shows that the efflux of cholesterol to BSA, ApoA-I, HDL and human serum in the presence of TO9 was all significantly lower in miR206 KO MPMs as compared to WT MPMs underlining the physiological significance of miR-206 on LXRα target genes.
3.8. LXR activation represses miR-206 expression in THP-1 macrophages whereas ox-LDL, VLDL and other inflammatory stimuli induce miR-206 expression
To determine whether a feedback mechanism between LXRα and miR-206 may exist, we activated LXR in THP-1 cells with TO9 (100 nM, 500 nM and 1 μM) and measured the expression levels of miR-206. Notably, LXR activation by TO9 drastically repressed miR-206 expression (Fig. 7a). We were also interested in identifying factors that may mediate miR-206 expression in macrophages. Thus, THP-1 cells were incubated in the presence of native LDL, oxLDL or VLDL. 50–100 μg/ml of oxLDL treatment or 50 μg/ml of VLDL treatment increased the expression of miR-206 in THP-1 cells by 1.5–2 fold compared to THP-1 cells incubated with native LDL (Fig. 7b).
Fig. 7.

Influence of LXR activation, modified lipoproteins, TNFα and LPS on miR-206 expression in THP-1 cells and human MDMs.
THP-1 cells were differentiated and treated with a) 100 nM, 500 nM or 1 μM of TO9; b) 50 μg/ml of native LDL (control), 50 μg/ml VLDL or 50 or 100 μg/ml oxLDL. c) Incubation with 10 μg/ml of TNFα. d) Incubation with 100 ng/ml of LPS. The relative abundance of miR-206 was measured by qRT-PCR. Values are mean ± SEM of three separate experiments; *p < 0.05; **p < 0.01; ***p < 0.001.
As oxLDL robustly induced the expression of miR-206, we were further interested to know whether oxLDL-induced inflammatory cytokines such as TNFα or inflammatory mediators such as LPS might influence the expression of miR-206. As shown in Fig. 7c a strong induction of miR-206 expression (20-fold) in THP-1 cells was treated with 10 ng/ml of TNFα. Consistently, LPS treatment of primary human monocyte-derived macrophages also resulted in a significantly increased expression of miR-206 (Fig. 7d).
4. Discussion
The major findings of this report are: 1) miR-206 targets LXRα and affects signaling pathways differently in the liver and macrophages of both mouse and human, 2) the identification of novel feedback regulation between LXR and miR206, miR-206 activates LXRα mediated pathway in cholesterol efflux and LXRα autoregulates its own expression by repressing miR-206 and 3) and the expression of miR-206 is induced by patho-physiological stimuli such as VLDL, oxLDL, LPS and TNFα.
miRs play important roles in a diverse spectrum of events in development, cell growth and differentiation as well as in metabolism. The varying expression patterns and the tissue specific functionality further emphasize the systemic importance of microRNAs in health and diseases [13]. There is an increasing body of evidence illustrating the role of miRs in cholesterol homeostasis and atherosclerosis: Sun et al. [19] recently published that miR-26 directly targets ABCA1 and ARL-7 RNA and controls LXR-dependent cholesterol efflux from RAW264.7 cells. Ramirez et al. [20] found a similar effect of miR-758 on ABCA1 expression and concomitantly reduced cholesterol efflux from human and mouse macrophages. Rayner et al. [21,22] and Horie et al. [23] reported that miR-33, that is co-transcribed with the SREBP-2 gene targets several LXR regulated genes and that miR-33 overexpression ameliorates cholesterol efflux from macrophages. Antagonizing miR-33 in mice promoted reverse cholesterol transport and regression of atherosclerosis [21]. Recently two independent groups published an LXRα and farnesoid X receptor (FXR) induced expression of miR-144 that targeted ABCA1 mRNA and reduced ABCA1 mediated cholesterol homeostasis in the liver [24,25]. Importantly, miR-1/206 and miR-613 have been identified as negative regulators of LXRα expression and lipogenesis in the liver [18,26]. Notably, most of these metabolic miRs that have been identified are reported to repress their target genes.
In the present study we identified a cell-specific activity for miR-206, which not only acts as a repressor of LXRα signaling in the liver cells but also intriguingly acts as an activator of LXRα in macrophages. This differential change in function is operative in both human and mouse macrophage systems and affects cholesterol homeostasis.
In line with earlier reports, we also observe a decrease in LXRα expression in HepG2 cells transfected with miR-206 and a concomitant down-regulated expression of LXRα target genes, such as ABCA1, ABCG1 ABCG5 and SREBP-1c. In contrast, THP-1miR206 OE cells showed induction of LXRα and its target genes such as ABCA1, ABCG1, ApoE and SREBP-1c accompanied with an increased cholesterol efflux capacity. Exactly opposite effects were achieved with lenti-virus mediated knockdown of miR-206 in THP-1 cells, as well as in miR206KO BMDMs and miR-206KO MPMs, which further substantiates the activation of LXRα pathway by miR-206 in macrophages. Interestingly, the basal unspecific cholesterol efflux to BSA was also reduced in miR-206KO MPMs. Though purified albumin added to cell culture is a relatively poor promoter of efflux, numerous studies report that aqueous diffusion contributes significantly to the total cell cholesterol flux [27]. Therefore, the reduction in efflux to BSA that we observe in miR-206KO MPMs might have been contributed by the aqueous transfer mediated by albumin.
As miRs were traditionally described as negative regulators of gene expression our results are quite surprising. Many recent reports suggest that miRs can not only repress but also activate translation by multiple mechanisms. Vasudevan [28] recently reported that during cell cycle, miRs oscillate between activation and repression; in proliferating cells miRs repress the translation whereas in G1/G0 arrest miRs facilitate translational activation. One reported mechanism for activation is the preferential interaction of AGO2–FXR1 complex (part of the miR ribonucleoprotein complex) with the AU rich regions on the target mRNA during cell cycle arrest [29]. They also report that under serum starved conditions let-7 interaction with the 3′UTR region of HMGA2 leads to activation; this mechanism also needed the interaction of AGO2–FXR1 complex with the HMGA2 mRNA [30]. A recent report from the same group suggests that nuclear localization of AGO2–FXR1 complex along with the target mRNA and miR is a requirement for translational activation of the mRNA [31].
Interestingly, mature miR-206 has been reported to have both nuclear localization and cytosolic localization but the functional relevance of compartmentalized localization is unknown. miR-206 is also one out of the few miRs that were reported to have translational activation capabilities [32,33]. Lin et al. [34] reported a feedback regulatory loop between KLF4 and miR-206 and the context specific activation of KLF4 by miR-206 in terminally differentiated cells. In our experimental conditions, THP-1 monocytes were differentiated into macrophages with 100 nM PMA for three days. PMA treatment initially induces cell cycle arrest and then terminal differentiation in macrophages. Thus after three days of differentiation, THP-1 macrophages are terminally differentiated cells which are thought to be incapable of further proliferation. As suggested earlier, it is conceivable that FXR1–AGO2–miR-206 complex trans-activates LXRα in THP-1miR206-OE cells by interacting with the 3’′UTR of LXR.
Similarly, miR-206-KO BMDMs were differentiated for 10 days which also yields terminally differentiated macrophages. miR-206 KO MPMs are already differentiated macrophages which are activated in response to the injected thioglycollate stimuli. In miR-206-KO macrophages the modest reduction of LXRα expression and less pronounced induction of TO9 or GW mediated LXRα signaling further emphasize the native requirement of miR-206 in activating LXRα signaling. Interestingly, in Fig. 3b and c, we observe a negative correlation in ABCG1 expression in THP-1miR206 OE and THP-1miR206 KO. Consistently, we also observed a reduction in ABCG1 expression in miR-206 KO BMDMs and MPMs (Figs. 4 & 5). This may explain the pronounced differences in cholesterol efflux to HDL and complete serum in comparison to ApoA-I. It is intriguing to speculate that the observed up- and downregulation of ABCG1 by miR-206 overexpression or knockdown might be at least in part independent of LXRα, i.e. the consequence of the direct regulation of ABCG1 expression by miR-206. However, further work is needed to experimentally address this possibility.
miRs have also been reported to activate gene expression transcriptionally by binding to the promoter sequence of the target gene [35]. To examine the possibility of such an interaction we performed an in silico RNAhybrid analysis which also showed a favorable minimum free binding energy (mfe) of (−) 20.3 kcal/mol. MicroPIR (microRNA-promoter interaction resource) is a recently developed online resource to predict putative miR-promoter interactions. In fact, miR-206 is one among the predicted ones which can bind at position − 4809th on the ABCG1 promoter with a mfe of (−) 25.50. MicroPIR also predicted miR-206 interaction with ApoE (at − 3937) and ABCA1 (at − 907). The fact that miR-206 has been shown to localize also in the nucleus and can interact with the promoters of the abovementioned genes suggests that such a mechanism might also be operative in macrophages.
Lipoproteins have been recently shown to carry miRs either through ABCA1 mediated miR loading to HDL or through binding to circulatory miRs [36]. miRs carried by HDL get activated inside the cell and can regulate gene expression. Interestingly, miR-206 is also carried in the circulation by HDL [36]. Physiologically, HDL molecules carrying miR-206 can deliver it to macrophages by SRB-1 mediated pathway and once inside the cells miR-206 can activate LXRα/ABCG1 mediated signaling which can in turn initiate cholesterol efflux to HDL [36]. This miR206–LXRα activation in macrophages might be part of the complex cholesterol homeostasis machinery. Another important finding is the negative feedback regulation of miR-206 by LXRα, which might be part of the LXR auto-regulatory mechanism to fine tune its own transcription and expression.
Our findings that VLDL and oxLDL as well as inflammatory cytokines and mediators such as TNFα and LPS induce the expression of miR-206 highlight the importance of this miR in diseases such as atherosclerosis and metabolic syndrome.
To summarize, we report a previously unknown pathway of miR-206 mediated LXRα function in macrophages that influence macrophage cholesterol homeostasis. As LXRα is involved in various mechanisms of macrophage biology and functionality, it will be important to investigate in detail the influence of the miR-206 mediated LXRα signaling in a patho-physiological context such as atherosclerosis.
Acknowledgements
The authors highly appreciate the technical assistance of Anton Ibovnik, Etrika Bulqiza and Silvia Povoden. This work was supported by the PhD Program in Molecular Medicine of the Medical University of Graz, the Austrian Science Fund (DK-MCD W1226, SFB-LIPOTOX F30, P22832) and the Marietta Blau Scholarship (OeAD). We thank Drs Rhonda Bassel and Eric Olson, Department of Molecular Biology, University of Texas SW-Dallas Medical Center for providing us with the miR-206 KO mice. BS is a member of the Institut Universitaire de France.
Appendix A. Supplementary data
Supplementary material.
References
- 1.Ostlund R.E., Jr. A minimal model for human whole body cholesterol metabolism. Am. J. Physiol. 1993;265:E513–E520. doi: 10.1152/ajpendo.1993.265.3.E513. [DOI] [PubMed] [Google Scholar]
- 2.Repa J.J., Mangelsdorf D.J. The role of orphan nuclear receptors in the regulation of cholesterol homeostasis. Annu. Rev. Cell Dev. Biol. 2000;16:459–481. doi: 10.1146/annurev.cellbio.16.1.459. [DOI] [PubMed] [Google Scholar]
- 3.Lusis A.J. Atherosclerosis. Nature. 2000;407:233–241. doi: 10.1038/35025203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Im S.-S., Osborne T.F. Liver X receptors in atherosclerosis and inflammation. Circ. Res. 2011;108:996–1001. doi: 10.1161/CIRCRESAHA.110.226878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sorrentino V., Nelson J.K., Maspero E., Marques A.R., Scheer L., Polo S., Zelcer N. The LXR–IDOL axis defines a clathrin-, caveolae-, and dynamin-independent endocytic route for LDLR internalization and lysosomal degradation. J. Lipid Res. 2013;54:2174–2184. doi: 10.1194/jlr.M037713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Tontonoz P. Transcriptional and posttranscriptional control of cholesterol homeostasis by liver X receptors. Cold Spring Harb. Symp. Quant. Biol. 2011;76:129–137. doi: 10.1101/sqb.2011.76.010702. [DOI] [PubMed] [Google Scholar]
- 7.Bories G., Colin S., Vanhoutte J., Derudas B., Copin C., Fanchon M., Daoudi M., Belloy L., Haulon S., Zawadzki C., Jude B., Staels B., Chinetti-Gbaguidi G. Liver X receptor activation stimulates iron export in human alternative macrophages. Circ. Res. 2013;113:1196–1205. doi: 10.1161/CIRCRESAHA.113.301656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Schultz J.R., Tu H., Luk A., Repa J.J., Medina J.C., Li L., Schwendner S., Wang S., Thoolen M., Mangelsdorf D.J., Lustig K.D., Shan B. Role of LXRs in control of lipogenesis. Genes Dev. 2000;14:2831–2838. doi: 10.1101/gad.850400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zelcer N., Tontonoz P. Liver X receptors as integrators of metabolic and inflammatory signaling. J. Clin. Invest. 2006;116:607–614. doi: 10.1172/JCI27883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Calkin A.C., Tontonoz P. Liver X receptor signaling pathways and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2010;30:1513–1518. doi: 10.1161/ATVBAHA.109.191197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bartel D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/s0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 12.Sacco J., Adeli K. MicroRNAs: emerging roles in lipid and lipoprotein metabolism. Curr. Opin. Lipidol. 2012;23:220–225. doi: 10.1097/MOL.0b013e3283534c9f. [DOI] [PubMed] [Google Scholar]
- 13.Soifer H.S., Rossi J.J., Saetrom P. MicroRNAs in disease and potential therapeutic applications. Mol. Ther. 2007;15:2070–2079. doi: 10.1038/sj.mt.6300311. [DOI] [PubMed] [Google Scholar]
- 14.Weischenfeldt J., Porse B. 2008. Bone Marrow-derived Macrophages (BMM): Isolation and Applications, CSH Protocols. ((2008) pdb prot5080) [DOI] [PubMed] [Google Scholar]
- 15.Chandak P.G., Obrowsky S., Radovic B., Doddapattar P., Aflaki E., Kratzer A., Doshi L.S., Povoden S., Ahammer H., Hoefler G., Levak-Frank S., Kratky D. Lack of acyl-CoA:diacylglycerol acyltransferase 1 reduces intestinal cholesterol absorption and attenuates atherosclerosis in apolipoprotein E knockout mice. Biochim. Biophys. Acta. 2011;1811:1011–1020. doi: 10.1016/j.bbalip.2011.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Malle E., Hazell L., Stocker R., Sattler W., Esterbauer H., Waeg G. Immunologic detection and measurement of hypochlorite-modified LDL with specific monoclonal antibodies. Arterioscler. Thromb. Vasc. Biol. 1995;15:982–989. doi: 10.1161/01.atv.15.7.982. [DOI] [PubMed] [Google Scholar]
- 17.Chennamsetty I., Kostner K.M., Claudel T., Vinod M., Frank S., Weiss T.S., Trauner M., Kostner G.M. Nicotinic acid inhibits hepatic APOA gene expression: studies in humans and in transgenic mice. J. Lipid Res. 2012;53:2405–2412. doi: 10.1194/jlr.M029769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zhong D., Huang G., Zhang Y., Zeng Y., Xu Z., Zhao Y., He X., He F. MicroRNA-1 and microRNA-206 suppress LXRalpha-induced lipogenesis in hepatocytes. Cell. Signal. 2013;25:1429–1437. doi: 10.1016/j.cellsig.2013.03.003. [DOI] [PubMed] [Google Scholar]
- 19.Sun D., Zhang J., Xie J., Wei W., Chen M., Zhao X. MiR-26 controls LXR-dependent cholesterol efflux by targeting ABCA1 and ARL7. FEBS Lett. 2012;586:1472–1479. doi: 10.1016/j.febslet.2012.03.068. [DOI] [PubMed] [Google Scholar]
- 20.Ramirez C.M., Davalos A., Goedeke L., Salerno A.G., Warrier N., Cirera-Salinas D., Suarez Y., Fernandez-Hernando C. MicroRNA-758 regulates cholesterol efflux through posttranscriptional repression of ATP-binding cassette transporter A1. Arterioscler. Thromb. Vasc. Biol. 2011;31:2707–2714. doi: 10.1161/ATVBAHA.111.232066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Rayner K.J., Sheedy F.J., Esau C.C., Hussain F.N., Temel R.E., Parathath S., van Gils J.M., Rayner A.J., Chang A.N., Suarez Y., Fernandez-Hernando C., Fisher E.A., Moore K.J. Antagonism of miR-33 in mice promotes reverse cholesterol transport and regression of atherosclerosis. J. Clin. Invest. 2011;121:2921–2931. doi: 10.1172/JCI57275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rayner K.J., Suarez Y., Davalos A., Parathath S., Fitzgerald M.L., Tamehiro N., Fisher E.A., Moore K.J., Fernandez-Hernando C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science. 2010;328:1570–1573. doi: 10.1126/science.1189862. (New York, N.Y.) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Horie T., Ono K., Horiguchi M., Nishi H., Nakamura T., Nagao K., Kinoshita M., Kuwabara Y., Marusawa H., Iwanaga Y., Hasegawa K., Yokode M., Kimura T., Kita T. MicroRNA-33 encoded by an intron of sterol regulatory element-binding protein 2 (Srebp2) regulates HDL in vivo. Proc. Natl. Acad. Sci. U. S. A. 2010;107:17321–17326. doi: 10.1073/pnas.1008499107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ramírez C.M., Rotllan N., Vlassov A.V., Dávalos A., Li M., Goedeke L., Aranda J.F., Cirera-Salinas D., Araldi E., Salerno A., Wanschel A., Zavadil J., Castrillo A., Kim J., Suárez Y., Fernández-Hernando C. Control of cholesterol metabolism and plasma high-density lipoprotein levels by microRNA-144. Circ. Res. 2013;112:1592–1601. doi: 10.1161/CIRCRESAHA.112.300626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.de Aguiar Vallim T.Q., Tarling E.J., Kim T., Civelek M., Baldan A., Esau C., Edwards P.A. MicroRNA-144 regulates hepatic ATP binding cassette transporter A1 and plasma high-density lipoprotein after activation of the nuclear receptor farnesoid X receptor. Circ. Res. 2013;112:1602–1612. doi: 10.1161/CIRCRESAHA.112.300648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ou Z., Wada T., Gramignoli R., Li S., Strom S.C., Huang M., Xie W. MicroRNA hsa-miR-613 targets the human LXRalpha gene and mediates a feedback loop of LXRalpha autoregulation. Mol. Endocrinol. 2011;25:584–596. doi: 10.1210/me.2010-0360. (Baltimore, MD) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sankaranarayanan S., de la Llera-Moya M., Drazul-Schrader D., Phillips M.C., Kellner-Weibel G., Rothblat G.H. Serum albumin acts as a shuttle to enhance cholesterol efflux from cells. J. Lipid Res. 2013;54:671–676. doi: 10.1194/jlr.M031336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Vasudevan S. Posttranscriptional upregulation by microRNAs. Wiley Interdiscip. Rev. RNA. 2012;3:311–330. doi: 10.1002/wrna.121. [DOI] [PubMed] [Google Scholar]
- 29.Vasudevan S., Steitz J.A. AU-rich-element-mediated upregulation of translation by FXR1 and Argonaute 2. Cell. 2007;128:1105–1118. doi: 10.1016/j.cell.2007.01.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vasudevan S., Tong Y., Steitz J.A. Switching from repression to activation: microRNAs can up-regulate translation. Science. 2007;318:1931–1934. doi: 10.1126/science.1149460. (New York, N.Y.) [DOI] [PubMed] [Google Scholar]
- 31.Truesdell S.S., Mortensen R.D., Seo M., Schroeder J.C., Lee J.H., LeTonqueze O., Vasudevan S. MicroRNA-mediated mRNA translation activation in quiescent cells and oocytes involves recruitment of a nuclear microRNP. Sci. Rep. 2012;2:842. doi: 10.1038/srep00842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ritland Politz J.C., Zhang F., Pederson T. MicroRNA-206 colocalizes with ribosome-rich regions in both the nucleolus and cytoplasm of rat myogenic cells. Proc. Natl. Acad. Sci. 2006;103:18957–18962. doi: 10.1073/pnas.0609466103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Foldes-Papp Z., Konig K., Studier H., Buckle R., Breunig H.G., Uchugonova A., Kostner G.M. Trafficking of mature miRNA-122 into the nucleus of live liver cells. Curr. Pharm. Biotechnol. 2009;10:569–578. doi: 10.2174/138920109789069332. [DOI] [PubMed] [Google Scholar]
- 34.Lin C.C., Liu L.Z., Addison J.B., Wonderlin W.F., Ivanov A.V., Ruppert J.M. A KLF4–miRNA-206 autoregulatory feedback loop can promote or inhibit protein translation depending upon cell context. Mol. Cell. Biol. 2011;31:2513–2527. doi: 10.1128/MCB.01189-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Majid S., Dar A.A., Saini S., Yamamura S., Hirata H., Tanaka Y., Deng G., Dahiya R. MicroRNA-205-directed transcriptional activation of tumor suppressor genes in prostate cancer. Cancer. 2010;116:5637–5649. doi: 10.1002/cncr.25488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Vickers K.C., Palmisano B.T., Shoucri B.M., Shamburek R.D., Remaley A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011;13:423–433. doi: 10.1038/ncb2210. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material.