

Abstract
Collagen VI is significant in the progression of numerous types of cancer. Type VI collagen consists of three α-chains and collagen VI α3 (COL6A3) encodes the α3 chain. The overexpression of COL6A3 has been demonstrated to correlate with high-grade ovarian cancer and contributes to cisplatin resistance; however, its role in human gastric cancer (GC) remains unclear. Using microarray meta-analysis, COL6A3 was observed to be frequently overexpressed in the GC tissues, furthermore, this overexpression was identified in five GC cell lines. A microarray-based co-expression network analysis was conducted and identified a total of 62 genes that were co-expressed with COL6A3, with the majority of the genes being involved in cancer-related processes, such as cell differentiation, migration and adhesion. Network analysis of these 62 genes demonstrated that fibronectin 1, a well-characterized oncogene, was located at the center of the COL6A3 co-expression network. Therefore, COL6A3 may act as an oncogene in human GC and the antagonism of COL6A3 may be an effective therapeutic treatment for GC.
Keywords: collagen VI α3, microarray, meta-analysis, gastric cancer
Introduction
Gastric cancer (GC) is the fourth most common type of malignancy worldwide, which results in 989,600 novel cases and 738,000 fatalities annually, specifically in Asian countries (1). Recent advancements in diagnosis and treatment modalities have been made, however, the prognosis of GC patients remains poor. As current therapeutic strategies are insufficient and do not achieve complete tumor ablation, it is important to analyze the molecular mechanisms of GC and identify novel biomarkers, as well as targets for therapeutic approaches, which may improve the clinical outcome for GC patients.
Collagen VI was initially identified as an extracellular matrix protein. It forms a microfilament network and binds to extracellular matrix proteins via its functional subdomains, which is important for the organization of fibrillar collagens and adhesion to the basement membrane (2). Collagen VI has recently attracted interest due to its involvement in breast and ovarian cancers (3–5). It is composed of three distinct α-chains (α1, -2 and -3) and collagen VI α3 (COL6A3) encodes the α3 chain, which is markedly longer than the other two chains (6). In a previous study, COL6A3 was shown to be upregulated in ovarian cancer (7), and Sherman-Baust et al (5) identified that the expression of COL6A3 was correlated with cisplatin resistance in ovarian cancer cell lines. Furthermore, highly or moderately differentiated ovarian tumors expressed lower levels of COL6A3 than poorly differentiated tumors, which indicated that the expression of COL6A3 was associated with the grade of the ovarian tumor (5). A recent exon array analysis study demonstrated that an alternative long isoform of COL6A3 was expressed, almost exclusively, in cancer samples, and may potentially serve as a novel cancer biomarker (8). Currently, the majority of studies relating to the oncogenic role of this gene focus on ovarian and breast cancer, however, the expression pattern and the biological functions of COL6A3 in human GC remain unknown.
In the present study, the authors investigated whether the expression level of COL6A3 was altered in GC, and a microarray meta-analysis was performed in order to assess the functional characteristics and molecular mechanisms of COL6A3 in GC.
Materials and methods
Gene expression patterns in GC
The Oncomine database (http://www.oncomine.org) was used to examine the differences in the transcriptional profiles between GC tissues and the adjacent normal tissues (9). Only the datasets that contained cancer versus normal analysis at the mRNA expression level were selected for analysis in the present study. In total, four GeneChip datasets, consisting of 318 paired GC and non-cancerous tissues, were selected according to the criteria shown in Table I.
Table I.
Oncomine datasets obtained for use in the present study.
| Dataset (Ref no.) | Samples | Data link |
|---|---|---|
| Chen Gastric (11) | 103 gastric adenocarcinomas and 29 normal gastric mucosa samples | http://genome-www.stanford.edu/gastric_cancer2/index.shtml |
| Cho Gastric (12) | 65 gastric adenocarcinoma, 19 paired surrounding normal tissue and six gastrointestinal stromal tumor samples | http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE13861 |
| D’Errico Gastric (13) | 31 paired gastric carcinoma and adjacent normal gastric mucosa and seven unmatched gastric carcinoma samples | http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE13911 |
| Wang Gastric (14) | 12 paired gastric carcinoma and normal gastric mucosa samples and three normal gastric tissue samples | http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE19826 |
Cell culture
Five human GC cell lines (AGS, HGC-27, BGC-823, SGC-7901 and MGC80-3) and one immortalized gastric cell line (GES-1) were purchased from Shanghai Institute of Cell Biology (Shanghai, China). All cell lines were incubated in Dulbecco’s modified Eagle’s medium (Gibco-BRL, Carlsbad, CA, USA) with 10% fetal bovine serum (SAFC Biosciences Inc., Lenexa, KS, USA), 100 U/ml penicillin and 100 mg/ml streptomycin (Sigma-Aldrich, St. Louis, MO, USA).
Quantitative polymerase chain reaction (qPCR) analysis
TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) was used to extract the total RNA from whole cells, and reverse-transcription was conducted using a TaqMan® Reverse Transcription kit (Applied Biosystems, Foster City, CA, USA). The DNA was amplified using an ABI® 7500 Real-Time PCR system (Applied Biosystems) and SYBR Premix Ex Taq (Takara, Kusatsu, Japan). The ΔΔCt method was used to calculate the relative RNA expression, which was normalized to GAPDH expression. PCR was performed using the following primers: forward, 5′-GAGACGCAGTGAGTGGGAAA-3′ and reverse, 5′-AGAGTCTTGTGCTGCTTGCT-3′ for COL6A3; and forward, 5′-CTCTCTGCTCCTCCTGTTCGAC-3′ and reverse, 5′-TGAGCGATGTGGCTCGGCT-3′ for GAPDH.
Co-expression analysis
The Oncomine database co-expression analysis tool was used to conduct the co-expression analysis of the microarray datasets. Using the co-expression score, the top 150 genes of each dataset were selected. The genes that appeared in at least two of the three datasets were defined as COL6A3 co-expressed genes.
Gene ontology (GO) and pathway enrichment analysis
GO and pathway enrichment analysis were conducted to examine COL6A3 co-expressed genes using the Database for Annotation, Visualization and Integrated Discovery (DAVID; http://david.abcc.ncifcrf.gov/). The categories, GOTERM_BP_3, GOTERM_CC_2 and GOTERM_MF_3 were selected, and the other options were set as defaults.
Construction of the gene interaction network
The gene interaction network was constructed using a gene expression pattern scanner (GePS: http://www.genomatix.de/) as described previously (10).
Statistical analysis
The independent Student’s t test was used to analyze the differences between two groups. Statistical analysis was performed using SPSS software version 16.0 (SPSS, Chicago, IL, USA). Data are presented as the means ± SD. P<0.05 was considered to indicate a statistically significant difference.
Results
COL6A3 is commonly overexpressed in GC
To determine the changes in the transcriptional pattern of GC cells, microarray datasets from the studies by Chen et al (11), Cho et al (12), D’Errico et al (13) and Wang et al (14) were analyzed using the Oncomine database. COL6A3 demonstrated a significant overexpression in the GC cells (P=3.98×10−15; Fig. 1A). To confirm this finding, the expression of COL6A3 in one immortalized gastric cell line (GES-1) and five GC cell lines (AGS, HGC-27, BGC-823, SGC-7901, MGC80-3) was analyzed using qPCR. The five GC cell lines exhibited ≥2.5-fold overexpression of COL6A3 compared with that of GES-1 cells (Fig. 1B).
Figure 1.
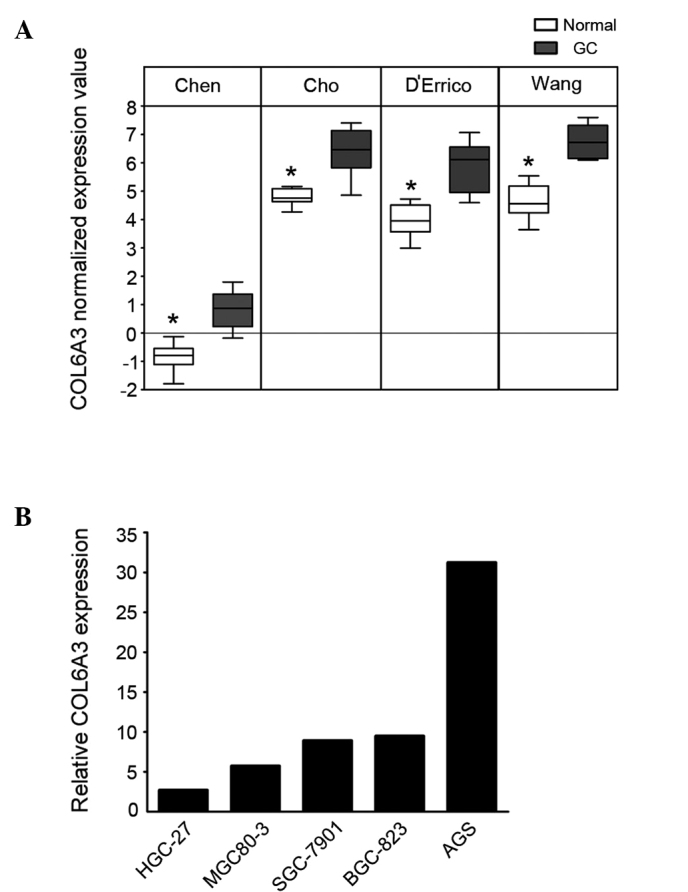
COL6A3 was overexpressed in gastric carcinoma tissue. (A) The expression pattern of COL6A3 in four GC datasets that were obtained using the Oncomine database; whiskers, 10th and 90th percentile; box boundaries, 75th and 25th percentile; line within the box, median. *P<0.001. (B) Relative COL6A3 expression of five GC cell lines (HGC-27, MGC80-3, SGC-7901, BGC-823 and AGS) compared with the mean value of a normal GC cell line (GES-1). COL6A3; collagen VI α3; GC, gastric cancer.
Genes co-expressed with COL6A3
A previous study indicated that genes which are co-expressed in different conditions may be functionally related or co-regulated (15). Therefore, a microarray co-expression analysis was conducted to identify the genes that were co-expressed with COL6A3. The dataset from the study by D’Errico et al (13) did not contain any co-expression data, therefore, the other three datasets consisting of 249 paired tissues were selected for inclusion in the co-expression analysis. Using a cut-off of the top 150 genes, which were identified by the co-expression score from each dataset, and with at least two appearances on the co-expressed list, 62 genes were identified as genes that were co-expressed with COL6A3 (Table II).
Table II.
Collagen VI α3 co-expressed genes with the cut-off for selection defined as an appearance in two datasets.
| Gene | Gene name | No. of appearances |
|---|---|---|
| COL6A3 | Collagen type VI α3 | 3 |
| COL1A2 | Collagen type I α2 | 3 |
| COL1A1 | Collagen type I α1 | 3 |
| COL12A1 | Collagen type XII α1 | 3 |
| THY1 | Thy-1 cell surface antigen | 3 |
| THBS2 | Thrombospondin 2 | 3 |
| BGN | Biglycan | 3 |
| CTHRC1 | Collagen triple helix repeat containing 1 | 3 |
| SULF1 | Sulfatase 1 | 3 |
| FAP | Fibroblast activation protein-α | 3 |
| SFRP4 | Secreted frizzled-related protein 4 | 3 |
| TIMP1 | Tissue inhibitor of metallopeptidase 1 | 3 |
| WNT2 | Wingless-type mouse mammary tumor virus integration site family member 2 | 3 |
| COL11A1 | Collagen type XI α1 | 3 |
| BMP1 | Bone morphogenetic protein 1 | 3 |
| SPOCK1 | Sparc/osteonectin cwcv and kazal-like domains proteoglycan (testican) 1 | 3 |
| SERPINH1 | Serpin peptidase inhibitor clade H (heat shock protein 47) member 1 (collagen binding protein 1) | 2 |
| CPXM1 | Carboxypeptidase X (M14 family) member 1 | 2 |
| INHBA | Inhibin β A | 2 |
| CDH11 | Cadherin 11, type 2, OB-cadherin (osteoblast) | 2 |
| RAB31 | Member of the RAS oncogene family | 2 |
| ANTXR1 | Anthrax toxin receptor 1 | 2 |
| NID2 | Nidogen 2 (osteonidogen) | 2 |
| PDGFRB | Platelet-derived growth factor receptor β polypeptide | 2 |
| COL4A2 | Collagen type IV α2 | 2 |
| COL4A1 | Collagen type IV α1 | 2 |
| TGFBI | Transforming growth factor β-induced (68kDa) | 2 |
| PLAU | Plasminogen activator urokinase | 2 |
| PRRX1 | Paired related homeobox 1 | 2 |
| LOX | Lysyl oxidase | 2 |
| PLXDC2 | Plexin domain containing 2 | 2 |
| LAMC1 | Laminin γ1 (formerly LAMB2) | 2 |
| OLFML2B | Olfactomedin-like 2B | 2 |
| CLDN4 | Claudin 4 | 2 |
| FAM83D | Family with sequence similarity 83, member D | 2 |
| ITGB5 | Integrin β5 | 2 |
| TNC | Tenascin C | 2 |
| SNAI2 | Snail family zinc finger 2 | 2 |
| FRMD6 | FERM domain containing 6 | 2 |
| COL6A1 | Collagen type VI α1 | 2 |
| NUAK1 | NUAK family, SNF1-like kinase 1 | 2 |
| HSPG2 | Heparan sulfate proteoglycan 2 | 2 |
| NOTCH3 | Notch 3 | 2 |
| CD276 | Cluster of differentiation 276 molecule | 2 |
| WNT5A | Wingless-type mouse mammary tumor virus integration site family member 5A | 2 |
| ECM1 | Extracellular matrix protein 1 | 2 |
| PDPN | Podoplanin | 2 |
| TNFAIP6 | Tumor necrosis factor α-induced protein 6 | 2 |
| ADAM12 | A disintegrin and metallo-peptidase domain 12 | 2 |
| GAS1 | Growth arrest-specific 1 | 2 |
| THBS1 | Thrombospondin 1 | 2 |
| COL10A1 | Collagen type X α1 | 2 |
| FNDC1 | Fibronectin type III domain containing 1 | 2 |
| SPHK1 | Sphingosine kinase 1 | 2 |
| MMP11 | Matrix metallopeptidase 11 (stromelysin 3) | 2 |
| CST1 | Cystatin SN | 2 |
| KRT80 | Keratin 80 | 2 |
| PMEPA1 | Prostate transmembrane protein, androgen induced 1 | 2 |
| SPP1 | Secreted phosphoprotein 1 | 2 |
| TNFRSF11B | Tumor necrosis factor receptor superfamily, member 11b | 2 |
| IGF2BP3 | Insulin-like growth factor 2 mRNA binding protein 3 | 2 |
| MFAP2 | Microfibrillar-associated protein 2 | 2 |
| EHD2 | EH-domain containing 2 | 2 |
GO and pathway enrichment analysis of COL6A3 co-expressed genes
GO and pathway enrichment analysis were conducted using the DAVID functional annotation chart tool (16) to further analyze the underlying mechanisms of COL6A3 and its co-expressed genes. In total, 36 biological process, seven cellular constituents, seven molecular function terms and six Kyoto encyclopedia of genes and genomes pathways were indicated to be significantly enriched (P<0.01; Table III). The extracellular matrix organization indicated the most marked enrichment among the GO biological process terms. The predominant function of COL6A3 has been identified to be the organization of matrix components, which supported the reliability of the present analysis. Furthermore, cell processes, such as cell differentiation, cell-substrate adhesion, regulation of cell proliferation, regulation of cell migration, cell motion and cell migration, which are considered to be cancer-related biological processes, were enriched (Fig. 2). This result indicated that COL6A3 may have been involved in the biological processes that promote the progression of GC.
Table III.
GO and pathway enrichment analysis of COL6A3 co-expressed genes.
| Category | Term | Function | Count | P-value | Fold enrichment | FDR |
|---|---|---|---|---|---|---|
| GOTERM _BP_3 | GO:0030198 | ECM organization | 11 | 5.88×10−12 | 26.86738026 | 8.02×10−9 |
| GO:0048731 | System development | 29 | 2.30×10−9 | 3.161608227 | 3.13×10−6 | |
| GO:0048513 | Organ development | 24 | 2.54×10−8 | 3.507740409 | 3.47×10−5 | |
| GO:0009653 | Anatomical structure morphogenesis | 19 | 2.42×10−7 | 4.032045523 | 3.30×10−4 | |
| GO:0009888 | Tissue development | 14 | 9.88×10−7 | 5.347765641 | 1.35×10−3 | |
| GO:0022603 | Regulation of anatomical structure morphogenesis | 7 | 1.89×10−4 | 8.119324546 | 2.57×10−1 | |
| GO:0030154 | Cell differentiation | 17 | 3.01×10−4 | 2.637947926 | 4.10×10−1 | |
| GO:0051093 | Negative regulation of developmental process | 7 | 4.64×10−4 | 6.865374809 | 6.31×10−1 | |
| GO:0031589 | Cell-substrate adhesion | 5 | 5.47×10−4 | 12.96014632 | 7.43×10−1 | |
| GO:0051239 | Regulation of multicellular organismal process | 12 | 7.52×−4 | 3.253176537 | 1.02 | |
| GO:0048519 | Negative regulation of biological process | 17 | 9.44×10−4 | 2.383179224 | 1.28 | |
| GO:0050793 | Regulation of developmental process | 10 | 9.95×10−4 | 3.768825934 | 1.35 | |
| GO:0060348 | Bone development | 5 | 1.28×10−3 | 10.32597024 | 1.73 | |
| GO:0009887 | Organ morphogenesis | 9 | 1.33×10−3 | 4.053492573 | 1.80 | |
| GO:0006928 | Cell motion | 8 | 2.20×10−3 | 4.278212512 | 2.96 | |
| GO:0042127 | Regulation of cell proliferation | 10 | 2.90×10−3 | 3.227685742 | 3.88 | |
| GO:0032101 | Regulation of response to external stimulus | 5 | 3.27×10−3 | 7.988014715 | 4.36 | |
| GO:0002683 | Negative regulation of immune system process | 4 | 4.02×10−3 | 12.24187315 | 5.34 | |
| GO:0009611 | Response to wounding | 8 | 4.05×10−3 | 3.834247063 | 5.39 | |
| GO:0030334 | Regulation of cell migration | 5 | 4.06×10−3 | 7.515351122 | 5.40 | |
| GO:0016477 | Cell migration | 6 | 4.13×10−3 | 5.522149303 | 5.49 | |
| GO:0016337 | Cell-cell adhesion | 6 | 4.13×10−3 | 5.522149303 | 5.49 | |
| GO:0050865 | Regulation of cell activation | 5 | 4.60×10−3 | 7.257681941 | 6.08 | |
| GO:0008284 | Positive regulation of cell proliferation | 7 | 5.02×10−3 | 4.295005013 | 6.64 | |
| GO:0009790 | Embryonic development | 8 | 5.90×10−3 | 3.577730534 | 7.75 | |
| GO:0007566 | Embryo implantation | 3 | 5.94×10−3 | 25.40188679 | 7.80 | |
| GO:0044259 | Multicellular organismal macromolecule metabolic process | 3 | 6.33×10−3 | 24.58247109 | 8.30 | |
| GO:0040012 | Regulation of locomotion | 5 | 6.37×10−3 | 6.615074686 | 8.34 | |
| GO:0051272 | Positive regulation of cell motion | 4 | 6.39×10−3 | 10.36811706 | 8.36 | |
| GO:0040017 | Positive regulation of locomotion | 4 | 6.39×10−3 | 10.36811706 | 8.36 | |
| GO:0048870 | Cell motility | 6 | 6.46×10−3 | 4.964538135 | 8.46 | |
| GO:0051270 | Regulation of cell motion | 5 | 6.48×10−3 | 6.580799687 | 8.49 | |
| GO:0048523 | Negative regulation of cellular process | 14 | 8.98×10−3 | 2.142327802 | 11.57 | |
| GO:0050867 | Positive regulation of cell activation | 4 | 9.00×10−3 | 9.153833078 | 11.59 | |
| GO:0009792 | Embryonic development ending in birth or egg hatching | 6 | 9.13×10−3 | 4.563213196 | 11.76 | |
| GO:0032844 | Regulation of homeostatic process | 4 | 9.67×10−3 | 8.912942734 | 12.41 | |
| GOTERM_CC_3 | GO:0031012 | ECM | 26 | 2.51×10−26 | 19.52139523 | 2.54×10−23 |
| GO:0005578 | Proteinaceous ECM | 25 | 1.40×10−25 | 20.23702331 | 1.41×10−22 | |
| GO:0044420 | ECM part | 15 | 8.00×10−18 | 33.20947414 | 8.10×10−15 | |
| GO:0005581 | Collagen | 10 | 5.53×10−15 | 74.00968523 | 5.62×10−12 | |
| GO:0005604 | Basement membrane | 6 | 1.13×10−5 | 19.92568449 | 1.14×10−2 | |
| GO:0005615 | Extracellular space | 12 | 4.49×10−5 | 4.537820116 | 4.55×10−2 | |
| GO:0005886 | Plasma membrane | 25 | 3.82×10−3 | 1.71454791 | 3.80 | |
| GO:0031252 | Cell leading edge | 3 | 9.65×10−2 | 5.631171702 | 64.22 | |
| GOTERM_MF_3 | GO:0019838 | Growth factor binding | 6 | 3.98×10−5 | 15.21982507 | 3.80×10−2 |
| GO:0005518 | Collagen binding | 4 | 3.06×10−4 | 29.59410431 | 2.92×10−2 | |
| GO:0005102 | Receptor binding | 11 | 1.21×10−3 | 3.306790436 | 1.15 | |
| KEGG_PATHWAY | hsa04512 | ECM-receptor interaction | 14 | 1.41×10−16 | 28.25 | 9.99×10−14 |
| hsa04510 | Focal adhesion | 14 | 1.52×10−11 | 11.80597015 | 1.35×10−08 |
GO, gene ontolgy; COL6A3, collagen VI α3; FDR, false discovery rate; BP; biological process; CC, cellular constituent; ECM, extracellular matrix; MF, molecular function; KEGG, Kyoto encyclopedia of genes and genomes. P<0.01 indicated a statistically significant difference.
Figure 2.
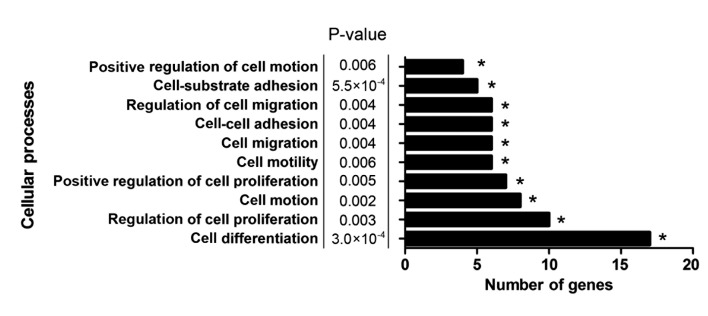
Gene ontology analysis of collagen VI α3 co-expressed genes was conducted using the Database for Annotation, Visualization and Integrated Discovery functional annotation chart tool. *P<0.01 for the pathway enrichment of COL6A3 co-expressed genes compared with Homo sapiens transcriptome background.
Network analysis of COL6A3
A network analysis was conducted using Genomatix GePS to construct the functional connections of COL6A3 co-expressed genes. FN1 was highlighted in this network, as it functionally associated with 50 (81.9%) COL6A3 co-expressed genes, which indicated that FN1 may act as a significant regulator in the COL6A33 regulatory network (Fig. 3).
Figure 3.

Network construction of COL6A3 co-expressed genes. The biological interactions of COL6A3 co-expressed genes were analyzed and visualized using a gene expression pattern scanner. The category of each gene is distinguished by its shape for factors, such as kinases and transporters. The direction of the arrow demonstrates whether a gene is upstream or downstream of another gene. Dashed line, co-cited genes; solid line, genes with an expertly curated connection. Genes with no interactions are not shown.
Discussion
COL6A3 is located on chromosome 2q37 and codes for the α-3 chain, one of the three α-chains of type VI collagen. It is hypothesized that COL6A3 accelerates cell anchoring and signaling through its interaction with integrin (17) and disruption of this gene results in muscular dystrophy (2). In addition to integrin, COL6A3 interacts with other matrix components, such as decorin, hyaluronan, heparan sulfate and NG2 proteoglycans (18). Furthermore, COL6A3 may promote neural crest cell migration and attachment, which is significant in the later stages of neural crest development (19).
Recently, COL6A3 has received increasing attention, due to its abnormal expression and the occurrence of alternative splicing in numerous types of cancer. Previous genome exon array studies have identified cancer-specific alternative splicing of exons 3, 4 and 6 of COL6A3 in colon, pancreatic, bladder and prostate cancer (8,20). Furthermore, COL6A3 was identified to be overexpressed in pancreatic (21) and ovarian cancer (7), which was associated with the poor differentiation of tumor cells (5). Although COL6A3 has been investigated in numerous other types of cancer, its biological mechanisms and expression pattern in GC remain unclear.
In the era of post-genomic medicine, microarray meta-analysis has been demonstrated to be an effective strategy for identifying gene expression changes in various types of cancer (22,23). In the present study, a microarray meta-analysis was performed to identify that COL6A3 was frequently overexpressed in hepatocellular carcinoma tissues, indicating that an increased expression of COL6A3 was associated with the carcinogenesis of GC. The underlying mechanisms that result in the increased expression of COL6A3 may relate to the transcriptional regulation of transforming growth factor (TGF)-β (24), however, this requires further investigation. To further define the biological mechanisms of COL6A3, a co-expression analysis was conducted to investigate the genes that are functionally related to, or co-regulated by, COL6A3. This identified 62 co-expression genes for COL6A3, the majority of which are involved in the processes of extracellular matrix organization such as lysyl oxidase, collagen type IV α2, TGF-β-induced and laminin γ1 (Table II). The functional network analysis of these co-expression genes was dominated by FN1, which demonstrated its predominant functional connections with other genes. FN1 is an adhesive protein of the extracellular matrix and it contains two apparently identical subunits with a range of binding sites for cell surface and extracellular ligands. It has been indicated that FN1 is involved in various aspects of cancer-related biological processes, such as cellular adhesion and migration. FN1 was identified to be overexpressed in hepatocellular, gastrointestinal, head and neck cancers (25,26), which indicated its involvement in tumorigenesis. Furthermore, Waalkes demonstrated that advanced-stage renal cancer patients exhibited increased FN1 expression when compared with patients exhibiting organ-confined diseases (27). Thus, the present study provided a mechanistic insight into the role of COL6A3 in GC.
In conclusion, the present study indicated that COL6A3 was regularly overexpressed in GC cells. A list of potential partner genes of COL6A3 was generated, the majority of which are involved in cancer-related processes, and a functional network of COL6A3 was constructed, which provided promising results to enable future studies to identify the precise role of COL6A3.
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Bönnemann CG. The collagen VI-related myopathies: muscle meets its matrix. Nat Rev Neurol. 2011;7:379–390. doi: 10.1038/nrneurol.2011.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Iyengar P, Espina V, Williams TW, et al. Adipocyte-derived collagen VI affects early mammary tumor progression in vivo, demonstrating a critical interaction in the tumor/stroma microenvironment. J Clin Invest. 2005;115:1163–1176. doi: 10.1172/JCI23424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Schäffler A, Schölmerich J, Buechler C. Mechanisms of disease: adipokines and breast cancer - endocrine and paracrine mechanisms that connect adiposity and breast cancer. Nat Clin Pract Endocrinol Metab. 2007;3:345–354. doi: 10.1038/ncpendmet0456. [DOI] [PubMed] [Google Scholar]
- 5.Sherman-Baust CA, Weeraratna AT, Rangel LB, et al. Remodeling of the extracellular matrix through overexpression of collagen VI contributes to cisplatin resistance in ovarian cancer cells. Cancer Cell. 2003;3:377–386. doi: 10.1016/s1535-6108(03)00058-8. [DOI] [PubMed] [Google Scholar]
- 6.Aumailley M, Mann K, von der Mark H, Timpl R. Cell attachment properties of collagen type VI and Arg-Gly-Asp dependent binding to its alpha 2(VI) and alpha 3(VI) chains. Exp Cell Res. 1989;181:463–474. doi: 10.1016/0014-4827(89)90103-1. [DOI] [PubMed] [Google Scholar]
- 7.Ismail RS, Baldwin RL, Fang J, Browning D, Karlan BY, Gasson JC, Chang DD. Differential gene expression between normal and tumor-derived ovarian epithelial cells. Cancer Res. 2000;60:6744–6749. [PubMed] [Google Scholar]
- 8.Thorsen K, Sørensen KD, Brems-Eskildsen AS, et al. Alternative splicing in colon, bladder, and prostate cancer identified by exon array analysis. Mol Cell Proteomics. 2008;7:1214–1224. doi: 10.1074/mcp.M700590-MCP200. [DOI] [PubMed] [Google Scholar]
- 9.Rhodes DR, Yu J, Shanker K, et al. Large-scale meta-analysis of cancer microarray data identifies common transcriptional profiles of neoplastic transformation and progression. Proc Natl Acad Sci USA. 2004;101:9309–9314. doi: 10.1073/pnas.0401994101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xing C, Zhou W, Ding S, et al. Reversing effect of ring finger protein 43 inhibition on malignant phenotypes of human hepatocellular carcinoma. Mol Cancer Ther. 2013;12:94–103. doi: 10.1158/1535-7163.MCT-12-0672. [DOI] [PubMed] [Google Scholar]
- 11.Chen X, Leung SY, Yuen ST, et al. Variation in gene expression patterns in human gastric cancers. Mol Biol Cell. 2003;14:3208–3215. doi: 10.1091/mbc.E02-12-0833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cho JY, Lim JY, Cheong JH, et al. Gene expression signature-based prognostic risk score in gastric cancer. Clin Cancer Res. 2011;17:1850–1857. doi: 10.1158/1078-0432.CCR-10-2180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.D’Errico M, de Rinaldis E, Blasi MF, et al. Genome-wide expression profile of sporadic gastric cancers with microsatellite instability. Eur J Cancer. 2009;45:461–469. doi: 10.1016/j.ejca.2008.10.032. [DOI] [PubMed] [Google Scholar]
- 14.Wang Q, Wen YG, Li DP, et al. Upregulated INHBA expression is associated with poor survival in gastric cancer. Med Oncol. 2012;29:77–83. doi: 10.1007/s12032-010-9766-y. [DOI] [PubMed] [Google Scholar]
- 15.Fierro AC, Vandenbussche F, Engelen K, Van de Peer Y, Marchal K. Meta analysis of gene expression data within and across species. Curr Genomics. 2008;9:525–534. doi: 10.2174/138920208786847935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Huang da W, Sherman BT, Lempicki RA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc. 2009;4:44–57. doi: 10.1038/nprot.2008.211. [DOI] [PubMed] [Google Scholar]
- 17.Pfaff M, Aumailley M, Specks U, Knolle J, Zerwes HG, Timpl R. Integrin and Arg-Gly-Asp dependence of cell adhesion to the native and unfolded triple helix of collagen type VI. Exp Cell Res. 1993;206:167–176. doi: 10.1006/excr.1993.1134. [DOI] [PubMed] [Google Scholar]
- 18.Burg MA, Tillet E, Timpl R, Stallcup WB. Binding of the NG2 proteoglycan to type VI collagen and other extracellular matrix molecules. J Biol Chem. 1996;271:26110–26116. doi: 10.1074/jbc.271.42.26110. [DOI] [PubMed] [Google Scholar]
- 19.Perris R, Kuo HJ, Glanville RW, Bronner-Fraser M. Collagen type VI in neural crest development: distribution in situ and interaction with cells in vitro. Dev Dyn. 1993;198:135–149. doi: 10.1002/aja.1001980207. [DOI] [PubMed] [Google Scholar]
- 20.Gardina PJ, Clark TA, Shimada B, et al. Alternative splicing and differential gene expression in colon cancer detected by a whole genome exon array. BMC Genomics. 2006;7:325. doi: 10.1186/1471-2164-7-325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Arafat H, Lazar M, Salem K, et al. Tumor-specific expression and alternative splicing of the COL6A3 gene in pancreatic cancer. Surgery. 2011;150:306–315. doi: 10.1016/j.surg.2011.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Smith DD, Saetrom P, Snøve O, Jr, Lundberg C, Rivas GE, Glackin C, Larson GP. Meta-analysis of breast cancer microarray studies in conjunction with conserved cis-elements suggest patterns for coordinate regulation. BMC Bioinformatics. 2008;9:63. doi: 10.1186/1471-2105-9-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rybaczyk LA, Bashaw MJ, Pathak DR, Huang K. An indicator of cancer: downregulation of monoamine oxidase-A in multiple organs and species. BMC Genomics. 2008;9:134. doi: 10.1186/1471-2164-9-134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Verrecchia F, Chu ML, Mauviel A. Identification of novel TGF-beta/Smad gene targets in dermal fibroblasts using a combined cDNA microarray/promoter transactivation approach. J Biol Chem. 2001;276:17058–17062. doi: 10.1074/jbc.M100754200. [DOI] [PubMed] [Google Scholar]
- 25.Torbenson M, Wang J, Choti M, Ashfaq R, Maitra A, Wilentz RE, Boitnott J. Hepatocellular carcinomas show abnormal expression of fibronectin protein. Mod Pathol. 2002;15:826–830. doi: 10.1097/01.MP.0000024257.83046.7C. [DOI] [PubMed] [Google Scholar]
- 26.Warawdekar UM, Zingde SM, Iyer KS, Jagannath P, Mehta AR, Mehta NG. Elevated levels and fragmented nature of cellular fibronectin in the plasma of gastrointestinal and head and neck cancer patients. Clin Chim Acta. 2006;372:83–93. doi: 10.1016/j.cca.2006.03.023. [DOI] [PubMed] [Google Scholar]
- 27.Waalkes S, Atschekzei F, Kramer MW, et al. Fibronectin 1 mRNA expression correlates with advanced disease in renal cancer. BMC Cancer. 2010;10:503. doi: 10.1186/1471-2407-10-503. [DOI] [PMC free article] [PubMed] [Google Scholar]