

Summary
Plasmodium knowlesi is a simian malaria parasite primarily infecting macaque species in Southeast Asia. Although its capacity to infect humans has been recognized since the early part of the last century, it has recently become evident that human infections are widespread and potentially life threatening. Historically, P. knowlesi has proven to be a powerful tool in early studies of malaria parasites, providing key breakthroughs in understanding many aspects of Plasmodium biology. However, the necessity to grow the parasite either in macaques or in vitro using macaque blood restricted research to laboratories with access to these resources. The recent adaptation of P. knowlesi to grow and proliferate in vitro in human red blood cells (RBCs) is therefore a substantial step towards revitalizing and expanding research on P. knowlesi. Furthermore, the development of a highly efficient transfection system to genetically modify the parasite makes P. knowlesi an ideal model to study parasite biology. In this review we elaborate on the importance of P. knowlesi in earlier phases of malaria research and highlight the future potential of the newly available human adapted P. knowlesi parasite lines.
Introduction
Malaria remains one of the most important infectious diseases of humans, causing almost 1 million deaths per year and extensive morbidity (Murray et al., 2012). Caused by six species of single cell protozoan pathogens from the genus Plasmodium, severe disease and death in Africa is associated with Plasmodium falciparum infections, whereas Plasmodium vivax also causes a large number of cases and significant morbidity worldwide. Plasmodium knowlesi is a simian malaria parasite phylogenetically closely related to P. vivax (Pain et al., 2008) and has long provided many advantages as an experimental model in the laboratory. It was first identified by Knowles and Das Gupta in 1932, in a cynomolgus macaque (Macaca fascicularis). In this first description, they demonstrated that the parasite could also infect humans by blood passage, but the first natural P. knowlesi infection was not identified until 1965 (Chin et al., 1965). Today, it is recognized as a major cause of clinical malaria in South East Asia (Singh et al., 2013).
Plasmodium parasites have a complicated lifecycle involving both mosquito and specific vertebrate hosts. For most of its development in the vertebrate it is an obligate intracellular parasite, first invading and undergoing asexual multiplication in hepatocytes and then multiple cycles of replication in red blood cells (RBCs). During growth within an infected RBC the parasite replicates to form a schizont, which undergoes segmentation producing invasive forms known as merozoites. These are highly polarized cells with a complex set of organelles and machinery for invasion of RBCs, but are also amongst the smallest of all eukaryotic organisms. Unlike P. falciparum, which produces 20 to 30 merozoites each about one micron in diameter, P. knowlesi produces fewer (average 10, but up to 16; Coatney et al., 1971) but larger merozoites (Fig. 1A).
Figure 1. Adaptation procedure and switch in host cell preference of P. knowlesi.
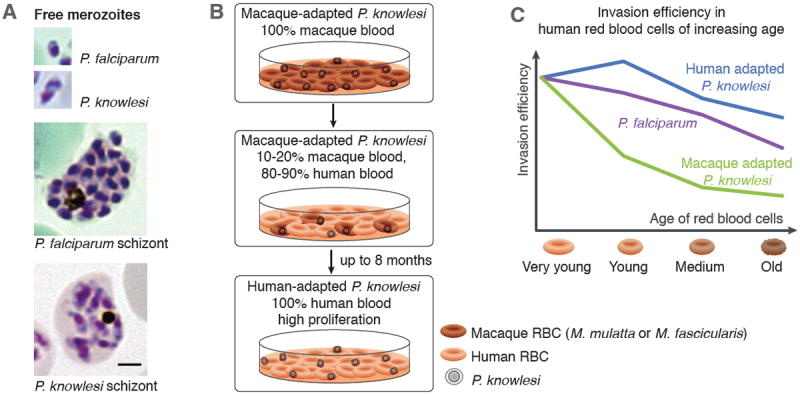
A. Comparison of P. falciparum and P. knowlesi schizonts and merozoites. P. knowlesi has fewer but larger merozoites in a mature schizont than P. falciparum.
B. Schematic representation of the process used to adapt macaque restricted P. knowlesi parasites to in vitro growth in human RBCs. Macaque RBCs are depicted in dark-red and human RBCs in red. Scale bar represents 2μm.
C. Switch in host cell preference to adapt to growth in RBCs. Graph shows the restriction of P. knowlesi grown in macaque blood to very young human RBCs and the subsequent expansion during adaptation to human blood towards a wider range of host cells. The subtle preference of P. falciparum for younger cells is also shown as reference.
The laboratory study of malaria has been aided by many critical developments, including the establishment of in vivo animal models, laboratory colonisation of mosquito vectors and the in vitro culture adaptation of erythrocytic stages. Here, we review the advances in the development of P. knowlesi as a model for understanding these various aspects of Plasmodium biology. We will highlight the recent technical developments of the P. knowlesi in vitro system and provide an outlook for the exploitation of the unique and exciting possibilities this model system offers for future research.
P. knowlesi is a zoonotic infection of humans
After the initial description of a P. knowlesi human infection (Chin et al., 1965), a subsequent survey suggested that the parasite was a rare zoonosis of humans (Warren et al., 1970). However, in 2004, Singh and colleagues were studying a large focus of apparent P. malariae clinical cases in Kapit Division, Malaysian Borneo. Genetic analysis revealed that these parasites had been misdiagnosed due to similar blood smear morphology, and that 120 out of 208 cases were P. knowlesi (Singh et al., 2004). Screening of archival blood smears then showed that P. knowlesi had been present in the region since at least 1996 (Lee et al., 2009). Most clinical P. knowlesi cases are uncomplicated with nonspecific febrile illness and thrombocytopenia. However, the rates of severe malaria are similar to that of P. falciparum, reaching 6.5% in a district hospital in Sarawak, Malaysia (Daneshvar et al., 2009). The most common complications are respiratory distress and renal failure, and these are often associated with hyperparasitaemia (Barber et al., 2013).
P. knowlesi is now known to be the most common cause of malaria in many areas of Malaysian Borneo, and a significant contributor in Peninsular Malaysia. Human cases of P. knowlesi have since been discovered throughout South East Asia, including the Philippines, Singapore, Indonesia, Vietnam, Cambodia, Thailand and Myanmar (Singh et al., 2013). While malaria cases caused by other Plasmodium species are on the decline in Sabah, Malaysian Borneo, P. knowlesi cases increased over 10-fold between 2004 and 2011, indicating that this parasite is an emerging threat in the region (William et al., 2013). However, there is as of yet no direct evidence for human-to-human transmission by mosquitoes in the field.
P. knowlesi: pioneering malaria research
P. knowlesi offers many important advantages as an experimental model for malaria and this was first demonstrated in several animal studies. While the parasite produces a mild chronic infection in its natural macaque host, Macaca fascicularis, it was commonly maintained experimentally in the rhesus macaque, Macaca mulatta, in which it produces fulminating infections (Coatney et al., 1971). Before in vitro culture of P. falciparum was achieved and widely adopted, P. knowlesi rhesus infections provided a source of highly synchronous parasites for studies in the laboratory.
The P. knowlesi animal models provided groundbreaking knowledge on several aspects of Plasmodium infection, including the basis of immunity. A landmark finding was the protection provided by passive immunization of newly infected monkeys with serum from chronically infected ones (Coggeshall et al., 1937). These transfer experiments and the discovery of antibodies targeting the parasite, demonstrated the importance of the immune response in controlling disease progression (Coggeshall, 1940). Shortly after, the first immunization studies were conducted using killed blood stage P. knowlesi or avian P. lophurae parasites (Freund et al., 1945a, Freund et al., 1945b). Both studies showed promising although not entirely protective effects, providing early evidence that a whole parasite vaccine may provide a valuable tool to prevent malaria parasite infection.
Antigenic variation mediated by the successive expression of variant proteins is an important immune evasion mechanism in Plasmodium parasites and was first described in P. knowlesi (Brown et al., 1965). This study paved the way to identify and characterize the corresponding gene family called SICAvar in P. knowlesi (Howard et al., 1983) and later the var genes in P. falciparum.
The invasion of a RBC by a merozoite is a crucial stage in the course of infection. This highly complex process, involving various receptor-ligand interactions, is thus of great research interest. The first of these receptor-ligand pairs was discovered in P. knowlesi, and comprises the Duffy binding protein-α (PkDBP-α) and the Duffy antigen receptor for chemokines (DARC) on human RBCs (Adams et al., 1990; Miller et al., 1975), and is also shared with P. vivax (Horuk et al., 1993). In total, the P. knowlesi genome encodes three DBPs (Pain et al., 2008). While being dispensable for invading rhesus macaque RBCs, PkDBP-α is pivotal for the invasion of human RBCs (Singh et al., 2005). The functions of the other two DBPs (DBP-β and -γ) are unknown but since they bind to rhesus but not human RBCs, it is likely that they contribute to rhesus RBC invasion (Chitnis et al., 1994). These studies highlight some of the seminal contributions made using P. knowlesi in the early days of malaria research.
From in vivo to in vitro studies
Although research on P. knowlesi in non-human primates was of great importance for the understanding of parasite biology, the lack of a robust continuous in vitro cultivation system restricted research to laboratories with access to suitable animal facilities. The first successful in vitro culture was achieved in 1945 when Ball and colleagues were able to maintain P. knowlesi in culture in rhesus RBCs for periods of up to 6 erythrocytic cycles (Ball et al., 1945). Follow-up studies provided insights into the importance of different components of the culture medium (Anfinsen et al., 1946; Geiman et al., 1966; Trigg, 1968); these detailed efforts were essential for optimizing experimental procedures such as invasion assays.
Although no long-term culture was possible, there were other advantages to studying this parasite. Groundbreaking studies in the 1970s included the first detailed live-microscopy investigation of P. knowlesi invasion (Dvorak et al., 1975). Dennis and colleagues developed a procedure to isolate live merozoites from infected rhesus RBCs using a polycarbonate sieve (Dennis et al., 1975), facilitating detailed electron microscopic studies of the invasion process and the identification of the moving junction between parasite and host cell (Aikawa et al., 1978; Bannister et al., 1975). While recent advances with P. falciparum have now allowed both the real-time imaging of invasion (Gilson et al., 2009) and purification of viable merozoites (Boyle et al., 2010), P. knowlesi still has the significant advantage of size and better viability. The application of new technologies such as super-resolution microscopy (Riglar et al., 2011) or long-term live microscopy (Grüring et al., 2011) to the P. knowlesi system will be particularly fruitful.
Although the achievement of Trager and Jensen to establish culture conditions for P. falciparum (Trager et al., 1976) diminished the use of P. knowlesi, the knowledge gained could be extrapolated for P. knowlesi culture. Further developments allowed the extended culture of P. knowlesi for several months (Butcher, 1979; Wickham et al., 1980), but required frequent media changes or intricate systems and were not widely used. A breakthrough was achieved in 2002 when Kocken and colleagues adapted the P. knowlesi H strain to continuous culture, developing an in vitro system that produced enough parasites to allow for transfection experiments (Kocken et al., 2002). Importantly, the in vitro adapted line was readily transferred back into rhesus monkeys, allowing genetic manipulation of parasites in vitro and the study of their phenotype in vivo. However, the requirement for rhesus macaque RBCs still restricted research to a few laboratories with access to monkey blood; the need for P. knowlesi parasite lines able to grow in human blood in vitro was clear (Kocken et al., 2009).
Adaptation of P. knowlesi to human red blood cells
Although human infections, either artificially induced or naturally acquired, have been known for decades, attempts to adapt P. knowlesi to stably proliferate in human RBCs failed, probably due to low replication rates (Kocken et al., 2009). This hurdle has now been overcome as our laboratories have reported the successful adaptation of P. knowlesi deriving from the H strain to grow in human RBCs (Lim et al., 2013; Moon et al., 2013). In both studies, parasites initially rapidly died out when grown exclusively in human RBCs but could be slowly adapted by initially growing them in a mixture of macaque (either M. mulatta or M. fascicularis) and human RBCs (Fig. 1B). Interestingly, after initial adaptation in human RBCs and proliferation rates of approximately two fold per day, the parasites eventually adapted further with rates increasing to up to five fold per day (Moon et al., 2013). This suggests two important adaptation steps, first at the level of invasion and second during the intra-erythrocytic growth phase. Whether the two lines underwent the same genetic or epigenetic changes remains an open question and is currently under investigation. A major step towards adaptation in one of the lines was a change in host RBC preference for invasion. While not restricted to cells of a certain age in macaques, P. knowlesi was found to invade mainly young human RBCs. In contrast, the human-adapted line had an increased invasion efficiency into older RBCs, providing access to a greater repertoire of suitable host cells (Fig. 1C). Mathematical modeling suggests that this expansion likely drives the high parasitaemias observed in some clinical infections (Lim et al., 2013). However, invasion of both adapted P. knowlesi lines was still DARC dependent since no invasion was observed in Duffy-negative human RBCs (Moon et al., 2013) or in cells treated with chymotrypsin depleting DARC from the RBC surface (Lim et al., unpublished). It will be of great interest to dissect the exact receptor requirements for the human lines as the DARC level on the surface of mature RBCs is diminished compared to reticulocytes (Liu et al., 2009). Furthermore, the human RBC-adapted P. knowlesi parasites retain their competence to efficiently invade macaque RBCs, showing that the invasion pathways used between the original and human RBC-adapted parasites are not mutually exclusive (Lim et al., 2013; Moon et al., 2013). The analysis of the adaptation mechanism in these lines could thus be leveraged to understand the basis of the successful zoonosis causing human P. knowlesi infections.
These human RBC-adapted lines are now available to the research community and their widespread use will greatly boost research on P. knowlesi, which is urgently needed in view of its potential as an emergent human infection.
Transfection of P. knowlesi
Genetic modification of the malaria parasite is a cornerstone of modern malaria research and has revealed countless details of parasite biology. However, P. falciparum is poorly amenable to genetic manipulation. Until very recently, transfection efficiencies in P. falciparum were only around 10-6 (O’Donnell et al., 2002), rendering the generation of transgenic parasites laborious and inefficient. Certain rodent malaria models, notably P. berghei, have higher transfection efficiencies of between 10-2 and 10-3 (Janse et al., 2006), but these parasites cannot be cultured in vitro, creating a substantial bottleneck for the high-throughput generation and analysis of transgenic parasites.
The first transfections in P. knowlesi were carried out in 1997 using episomal constructs, with P. berghei or P. falciparum regulatory regions driving the T. gondii dihydrofolate reductase-thymidylate synthase selectable marker (van der Wel et al., 1997). Schizonts obtained from infected macaques were electroporated and reinoculated into recipient macaques, enabling isolation of transgenic parasites in just over a week of drug selection. Similar methods were used later to integrate constructs by double-crossover homologous recombination to create a circumsporozoite protein (CSP) gene knockout line. Critically, for the first time these authors were able to demonstrate that the whole process could be carried out entirely in vitro using a line adapted to grow in culture with macaque RBCs (Kocken et al., 2002). Additional selectable markers previously used in other Plasmodium species were also demonstrated to work in P. knowlesi providing positive selection resistance to neomycin, Blasticidin and WR99210 as well as negative selection by susceptibility to Ganciclovir (van der Wel et al., 2004).
More recently, transfection has been achieved using the human-adapted line. Synchronous late stage schizonts from the A1H.1 line were transfected using the Amaxa electroporation system (Moon et al., 2013), which had in the past significantly improved transfection efficiency in P. berghei (Janse et al., 2006). Expression of transgenes was also improved by the use of P. knowlesi specific promoter regions. Transfection with an episomal construct containing a GFP expression cassette resulted in ~30% GFP positive parasites the day following transfection (Moon et al., 2013). This indicates the highest recorded transfection efficiency for any human malaria parasite and for the first time, parasites could be analyzed in the first growth cycle after transfection. Using linearized DNA constructs targeted to a genomic locus via single crossover recombination, integrated parasites lines could be obtained in just over a week. Integration efficiency was sufficiently high to allow immediate dilution cloning, meaning that extensive drug cycling is not required. Importantly, the whole procedure could be carried out solely using human RBCs and with 10-fold fewer schizonts per transfection (108) than used in previous methods. Subsequent work has demonstrated that this efficiency also holds true for constructs targeting other loci (Moon et al., unpublished). The transfection efficiency in the other human adapted line has yet to be tested.
Towards establishing transmission of P. knowlesi in the lab
The sexual stages of malaria parasites known as gametocytes form when a subset of replicating asexual blood stages switch to the sexual development pathway. Once induced, P. falciparum gametocytes take around 10 days to fully mature, while P. knowlesi gametocyte formation requires about 48 hours, which may prove a significant experimental advantage (Coatney et al., 1971). Early work demonstrated that P. knowlesi could be readily transmitted via mosquitoes in the laboratory from macaque to macaque and even human to human (Chin et al., 1968). Unfortunately, none of the P. knowlesi lines adapted to in vitro growth produce gametocytes (Moon et al., 2013; Zeeman et al., 2013, Lim et al., unpublished) and attempts to induce gametocytogenesis have been so far unsuccessful. It is well established that parasite lines maintained by extended blood passage or in culture may lose their ability to form gametocytes (Janse et al., 1992). Therefore, it might be necessary to culture-adapt lines that have recently been transmitted by mosquitoes and retain the ability to produce gametocytes.
There is also a need to identify suitable vectors for use in the laboratory. The most commonly used experimental malaria parasite vector, Anopheles stephensi, cannot effectively transmit P. knowlesi. While the parasite is able to form midgut oocysts in An. stephensi, the sporozoites released from these oocysts are unable to invade the mosquito salivary glands (Coatney et al., 1971). Early fieldwork identified Anopheles hackeri as a vector for P. knowlesi in Malaysia (Wharton et al., 1961), but more recent work has implicated other vectors, including Anopheles cracens in peninsular Malaysia (Vythilingam et al., 2008) and Anopheles latens in Malaysian Borneo (Vythilingam et al., 2006). Colonies of An. cracens have been established in the laboratory and although their maintenance remains challenging, this species may represent the vector of choice for P. knowlesi transmission in the laboratory (Amir et al., 2013). While significant challenges remain, P. knowlesi may provide a useful alternative to study sexual stages.
The use of P. knowlesi as a model for P. vivax
Alongside P. falciparum, P. vivax causes a very high burden of malaria globally, but currently lacks an in vitro model. Historically, common aspects of P. knowlesi and P. vivax invasion, especially their use of DARC as a receptor to invade human RBCs, enabled the understanding P. vivax invasion using P. knowlesi as a surrogate (Horuk et al., 1993; Singh et al., 2002). Beside the potential to scrutinize the use of DBP as a vaccine candidate, the P. knowlesi system provides the unique opportunity to investigate the role of the reticulocyte binding-like proteins (RBLs), another major ligand family that mediates invasion. The binding properties of the two members present in the P. knowlesi genome (PkNBPXa and PkNBPXb) have recently been investigated (Meyer et al., 2009; Semenya et al., 2012). While both members bind to rhesus macaque RBCs, PkNBPXa also binds human RBCs (Semenya et al., 2012). An ortholog of NBPXb has recently been identified in a de novo assembly of the P. vivax genome (Hester et al., 2013). Studies on these invasion molecules could shed light on the adaptation to human RBCs. The sequencing of the P. vivax and P. knowlesi genomes revealed a near perfect synteny interspersed with expansions of species-specific genes (Carlton et al., 2008; Pain et al., 2008). The two genomes also share more than 90 genes with no orthologs in P. falciparum (Frech et al., 2011), indicating that the accessibility of the P. knowlesi model may prove beneficial for the study of other aspects of P. vivax biology.
However, notable differences between the two parasites limit the use of P. knowlesi as a bona fide P. vivax model. Unlike P. knowlesi, P. vivax forms hypnozoites, dormant liver stage parasites that can lead to disease relapses years after the initial infection. Developments in a more closely related simian malaria model, Plasmodium cynomolgi, have provided an alternative means to study this phenomenon (Voorberg-van der Weel et al., 2013). Nevertheless, as long as P. cynomolgi and P. vivax lack a robust in vitro system, P. knowlesi remains a highly accessible and valuable tool to further explore aspects of P. vivax biology.
Future studies
The high transfection efficiency obtained in P. knowlesi, combined with the ability to culture the parasites in human RBCs means that the system is ideal for the development of both novel reverse genetics and high-throughput genetic techniques (Fig. 2). Conditional knockout systems have proved crucial in determining the functions of essential blood stage proteins. Several conditional knockout/knockdown systems have been developed for use in P. falciparum, including the use of ligand-regulatable FKBP protein destabilization domains (ddFKBP) (Armstrong et al., 2007), as well as the DiCre conditional recombinase system (Collins et al., 2013). These systems could be quickly adapted for use in P. knowlesi. Similarly, genome-editing tools such as the zinc-finger nucleases have revolutionized many fields, and recently have been demonstrated to target genomic integration with unprecedented efficiency in P. falciparum (Straimer et al., 2012). This approach, and analogous systems such the TALEN (transcription activator-like effector nucleases) (Christian et al., 2010) and the CRISPR (clustered regularly interspaced short palindromic repeat)-Cas system (Ran et al., 2013), could be readily adapted for use in P. knowlesi to facilitate genomic integration of DNA constructs in a matter of days.
Figure 2. Outline of new tools to advance malaria research using human red blood cell adapted P. knowlesi.
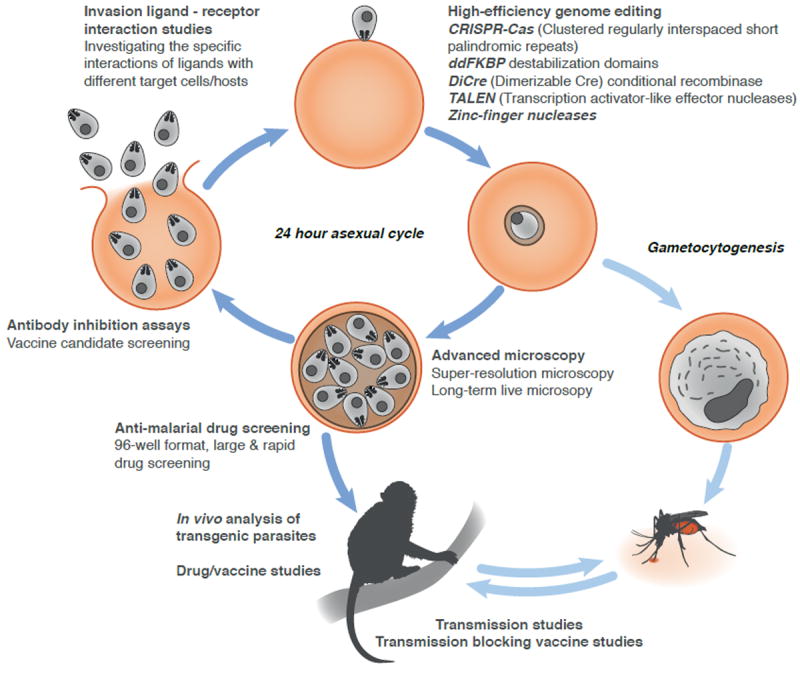
The 24h P. knowlesi life cycle is depicted, starting with the invasion of a merozoite and subsequent development from ring to schizont stage. A selection of tools that can be applied to the human adapted P. knowlesi line is shown.
The generation of a gametocyte-producing-line and transmission competent mosquito colonies would allow the research to expand to other life stages.
Human RBC-adapted P. knowlesi also provides exciting opportunities for high-throughput genome-scale transgenesis. The ability to culture P. knowlesi in a microplate format, combined with the high transfection efficiency, allows for the generation of pools of recombinant parasites. The development of recombineering techniques for high-throughput generation of DNA constructs for use in P. berghei has greatly benefited the field (Pfander et al., 2011). Application of such techniques to P. knowlesi, which can be cloned and grown in 96-well plates, will provide an important resource for the malaria research community. Taken together, human RBCs-adapted P. knowlesi could emerge as an ideal system for forward genetic studies in malaria.
Acknowledgments
We thank Kathryn Shaw Saliba for critical reading of the manuscript. Christof Grüring is supported by a fellowship from the Swiss National Science Foundation. Robert W. Moon is supported by a Medical Research Council Career Development Fellowship. Work in the Holder and Blackman labs is supported by the MRC (File reference number U117532067 and U117532063, respectively). Manoj T. Duraisingh is supported by an NIH Award (R01AI091787). We apologize to colleagues whose work we could not cite due to space restrictions.
References
- Adams JH, Hudson DE, Torii M, Ward GE, Wellems TE, Aikawa M, et al. The Duffy receptor family of Plasmodium knowlesi is located within the micronemes of invasive malaria merozoites. Cell. 1990;63:141–153. doi: 10.1016/0092-8674(90)90295-p. [DOI] [PubMed] [Google Scholar]
- Aikawa M, Miller LH, Johnson J, Rabbege J. Erythrocyte entry by malarial parasites. A moving junction between erythrocyte and parasite. J Cell Biol. 1978;77:72–82. doi: 10.1083/jcb.77.1.72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amir A, Sum JS, Lau YL, Vythilingam I, Fong MY. Colonization of Anopheles cracens: a malaria vector of emerging importance. Parasit Vectors. 2013;6:81. doi: 10.1186/1756-3305-6-81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anfinsen CB, Geiman QM, McKee RW, Ormsbee RA, Ball EG. Studies on malarial strasites: VIII. Factors affecting the growth of Plasmodium knowlesi in vitro. J Exp Med. 1946;84:607–621. doi: 10.1084/jem.84.6.607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Armstrong CM, Goldberg DE. An FKBP destabilization domain modulates protein levels in Plasmodium falciparum. Nat Meth. 2007;4:1007–1009. doi: 10.1038/nmeth1132. [DOI] [PubMed] [Google Scholar]
- Ball EG, Anfinsen CB, Geiman QM, McKee RW, Ormsbee RA. In vitro growth and multiplication of the malaria parasite, Plasmodium knowlesi. Science. 1945;101:542–544. doi: 10.1126/science.101.2630.542. [DOI] [PubMed] [Google Scholar]
- Bannister LH, Butcher GA, Dennis ED, Mitchell GH. Structure and invasive behaviour of Plasmodium knowlesi merozoites in vitro. Parasitol. 1975;71:483–491. doi: 10.1017/s0031182000047247. [DOI] [PubMed] [Google Scholar]
- Barber BE, William T, Grigg MJ, Menon J, Auburn S, Marfurt J, et al. A prospective comparative study of knowlesi, falciparum, and vivax malaria in Sabah, Malaysia: high proportion with severe disease from Plasmodium knowlesi and Plasmodium vivax but no mortality with early referral and Artesunate therapy. Clin Infect Dis. 2013;56:383–397. doi: 10.1093/cid/cis902. [DOI] [PubMed] [Google Scholar]
- Boyle MJ, Wilson DW, Richards JS, Riglar DT, Tetteh KK, Conway DJ, et al. Isolation of viable Plasmodium falciparum merozoites to define erythrocyte invasion events and advance vaccine and drug development. Proc Natl Acad Sci USA. 2010;107:14378–14383. doi: 10.1073/pnas.1009198107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown KN, Brown IN. Immunity to malaria: antigenic variation in chronic infections of Plasmodium knowlesi. Nature. 1965;208:1286–1288. doi: 10.1038/2081286a0. [DOI] [PubMed] [Google Scholar]
- Butcher GA. Factors affecting the in vitro culture of Plasmodium falciparum and Plasmodium knowlesi. Bull World Health Organ. 1979;57:17–26. [PMC free article] [PubMed] [Google Scholar]
- Carlton JM, Adams JH, Silva JC, Bidwell SL, Lorenzi H, Caler E, et al. Comparative genomics of the neglected human malaria parasite Plasmodium vivax. Nature. 2008;455:757–763. doi: 10.1038/nature07327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chin W, Contacos PG, Coatney GR, Kimball HR. A naturally acquired quotidian-type malaria in man transferable to monkeys. Science. 1965;149:865. doi: 10.1126/science.149.3686.865. [DOI] [PubMed] [Google Scholar]
- Chin W, Contacos PG, Collins WE, Jeter MH, Alpert E. Experimental mosquito-transmission of Plasmodium knowlesi to man and monkey. Am J Trop Med Hyg. 1968;17:355–358. doi: 10.4269/ajtmh.1968.17.355. [DOI] [PubMed] [Google Scholar]
- Chitnis CE, Miller LH. Identification of the erythrocyte binding domains of Plasmodium vivax and Plasmodium knowlesi proteins involved in erythrocyte invasion. J Exp Med. 1994;180:497–506. doi: 10.1084/jem.180.2.497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christian M, Cermak T, Doyle EL, Schmidt C, Zhang F, Hummel A, et al. Targeting DNA double-strand breaks with TAL Effector Nucleases. Genetics. 2010;186:757–761. doi: 10.1534/genetics.110.120717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coatney GR, Collins WE, McWilson W, Contacos PG. The Primate Malarias. Bethesda: U.S. Department of Health, Education and Welfare; 1971. [Google Scholar]
- Coggeshall LT, Kumm HW. Demonstration of passive immunity in experimental monkey malaria. J Exp Med. 1937;66:177–190. doi: 10.1084/jem.66.2.177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coggeshall LT. The occurrence of malaria antibodies in human serum following induced infection with Plasmodium knowlesi. J Exp Med. 1940;72:21–31. doi: 10.1084/jem.72.1.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins CR, Das S, Wong EH, Andenmatten N, Stallmach R, Hackett F, et al. Robust inducible Cre recombinase activity in the human malaria parasite Plasmodium falciparum enables efficient gene deletion within a single asexual erythrocytic growth cycle. Mol Microbiol. 2013;88:687–701. doi: 10.1111/mmi.12206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daneshvar C, Davis TME, Cox-Singh J, Rafa’ee MZ, Zakaria SK, Divis PCS, Singh B. Clinical and laboratory features of human Plasmodium knowlesi infection. Clin Infect Dis. 2009;49:852–860. doi: 10.1086/605439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dennis ED, Mitchell GH, Butcher GA, Cohen S. In vitro isolation of Plasmodium knowlesi merozoites using polycarbonate sieves. Parasitol. 1975;71:475–481. doi: 10.1017/s0031182000047235. [DOI] [PubMed] [Google Scholar]
- Dvorak JA, Miller LH, Whitehouse WC, Shiroishi T. Invasion of erythrocytes by malaria merozoites. Science. 1975;187:748–750. doi: 10.1126/science.803712. [DOI] [PubMed] [Google Scholar]
- Frech C, Chen N. Genome comparison of human and non-human malaria parasites reveals species subset-specific genes potentially linked to human disease. PLoS Comput Biol. 2011;7:e1002320. doi: 10.1371/journal.pcbi.1002320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freund J, Thomson KJ, Sommer HE, Walter AW, Schenkein EL. Immunization of rhesus monkeys against malarial infection (P. knowlesi) with killed parasites and adjuvants. Science. 1945a;102:202–204. doi: 10.1126/science.102.2643.202. [DOI] [PubMed] [Google Scholar]
- Freund J, Sommer HE, Walter AW. Immunization against malaria: vaccination of ducks with killed parasites incorporated with adjuvants. Science. 1945b;102:200–202. doi: 10.1126/science.102.2643.200. [DOI] [PubMed] [Google Scholar]
- Geiman QM, Siddiqui WA, Schnell JV. Plasma replacement for in vitro culture of Plasmodium knowlesi. Science. 1966;153:1129–1130. doi: 10.1126/science.153.3740.1129. [DOI] [PubMed] [Google Scholar]
- Gilson PR, Crabb BS. Morphology and kinetics of the three distinct phases of red blood cell invasion by Plasmodium falciparum merozoites. Int J Parasitol. 2009;39:91–96. doi: 10.1016/j.ijpara.2008.09.007. [DOI] [PubMed] [Google Scholar]
- Grüring C, Heiber A, Kruse F, Ungefehr J, Gilberger TW, Spielmann T. Development and host cell modifications of Plasmodium falciparum blood stages in four dimensions. Nat Commun. 2011;2:165. doi: 10.1038/ncomms1169. [DOI] [PubMed] [Google Scholar]
- Hester J, Chan ER, Menard D, Mercereau-Puijalon O, Barnwell J, Zimmerman PA, et al. De novo assembly of a field isolate genome reveals novel Plasmodium vivax erythrocyte invasion genes. PLoS Negl Trop Dis. 2013;7:e2569. doi: 10.1371/journal.pntd.0002569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howard RJ, Barnwell JW, Kao V. Antigenic variation of Plasmodium knowlesi malaria : identification of the variant antigen on infected erythrocytes. Proc Natl Acad Sci U S A. 1983;80:4129–4133. doi: 10.1073/pnas.80.13.4129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horuk R, Chitnis CE, Darbonne WC, Colby TJ, Rybicki A, Hadley TJ, et al. A receptor for the malarial parasite Plasmodium vivax: the erythrocyte chemokine receptor. Science. 1993;261:1182–1184. doi: 10.1126/science.7689250. [DOI] [PubMed] [Google Scholar]
- Janse CJ, Franke-Fayard B, Mair GR, Ramesar J, Thiel C, Engelmann S, et al. High efficiency transfection of Plasmodium berghei facilitates novel selection procedures. Mol Biochem Parasitol. 2006;145:60–70. doi: 10.1016/j.molbiopara.2005.09.007. [DOI] [PubMed] [Google Scholar]
- Janse CJ, Ramesar J, van den Berg FM, Mons B. Plasmodium berghei in vivo generation and selection of karyotype mutants and non-gametocyte producer mutants. Exp Parasitol. 1992;74:1–10. doi: 10.1016/0014-4894(92)90133-u. [DOI] [PubMed] [Google Scholar]
- Kocken CHM, Ozwara H, van der Wel A, Beetsma AL, Mwenda JM, Thomas AW. Plasmodium knowlesi provides a rapid in vitro and in vivo transfection system that enables double-crossover gene knockout studies. Infect Immun. 2002;70:655–660. doi: 10.1128/IAI.70.2.655-660.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kocken CHM, Zeeman A-M, Voorberg-van der Wel A, Thomas AW. Transgenic Plasmodium knowlesi: relieving a bottleneck in malaria research? 2009 doi: 10.1016/j.pt.2009.05.009. [DOI] [PubMed] [Google Scholar]
- Lee KS, Cox-Singh J, Brooke G, Matusop A, Singh B. Plasmodium knowlesi from archival blood films: Further evidence that human infections are widely distributed and not newly emergent in Malaysian Borneo. Int J Parasitol. 2009;39:1125–1128. doi: 10.1016/j.ijpara.2009.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim C, Hansen E, DeSimone TM, Moreno Y, Junker K, Bei A, et al. Expansion of host cellular niche can drive adaptation of a zoonotic malaria parasite to humans. Nat Commun. 2013;4:1638. doi: 10.1038/ncomms2612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu J, Guo X, Mohandas N, Chasis JA, An X. Membrane remodeling during reticulocyte maturation. Blood. 2009;115:2021–2027. doi: 10.1182/blood-2009-08-241182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer EVS, Semenya AA, Okenu DMN, Dluzewski AR, Bannister LH, Barnwell JW, et al. The reticulocyte binding-like proteins of P. knowlesi locate to the micronemes of merozoites and define two new members of this invasion ligand family. Mol Biochem Parasitol. 2009;165:111–121. doi: 10.1016/j.molbiopara.2009.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller LH, Mason SJ, Dvorak JA, McGinniss MH, Rothman IK. Erythrocyte receptors for (Plasmodium knowlesi) malaria: Duffy blood group determinants. Science. 1975;189:561–563. doi: 10.1126/science.1145213. [DOI] [PubMed] [Google Scholar]
- Moon RW, Hall J, Rangkuti F, Ho YS, Almond N, Mitchell GH, et al. Adaptation of the genetically tractable malaria pathogen Plasmodium knowlesi to continuous culture in human erythrocytes. Proc Natl Acad Sci USA. 2013;110:531–536. doi: 10.1073/pnas.1216457110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murray CJ, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ, Haring D, et al. Global malaria mortality between 1980 and 2010: a systematic analysis. Lancet. 2012;379:413–431. doi: 10.1016/S0140-6736(12)60034-8. [DOI] [PubMed] [Google Scholar]
- O’Donnell RA, Freitas-Junior LH, Preiser PR, Williamson DH, Duraisingh MT, McElwain TF, et al. A genetic screen for improved plasmid segregation reveals a role for Rep20 in the interaction of Plasmodium falciparum chromosomes. EMBO J. 2002;21:1231–1239. doi: 10.1093/emboj/21.5.1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pain A, Böhme U, Berry AE, Mungall K, Finn RD, Jackson AP, et al. The genome of the simian and human malaria parasite Plasmodium knowlesi. Nature. 2008;455:799–803. doi: 10.1038/nature07306. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfander C, Anar B, Schwach F, Otto TD, Brochet M, Volkmann K, et al. A scalable pipeline for highly effective genetic modification of a malaria parasite. Nat Meth. 2011;8:1078–1082. doi: 10.1038/nmeth.1742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ran FA, Hsu PD, Lin C-Y, Gootenberg JS, Konermann S, Trevino AE, et al. Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell. 2013;154:1380–1389. doi: 10.1016/j.cell.2013.08.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riglar DT, Richard D, Wilson DW, Boyle MJ, Dekiwadia C, Turnbull L, et al. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host & Microbe. 2011;9:9–20. doi: 10.1016/j.chom.2010.12.003. [DOI] [PubMed] [Google Scholar]
- Semenya AA, Tran TM, Meyer EV, Barnwell JW, Galinski MR. Two functional reticulocyte binding-like (RBL) invasion ligands of zoonotic Plasmodium knowlesi exhibit differential adhesion to monkey and human erythrocytes. Malar J. 2012;11:228. doi: 10.1186/1475-2875-11-228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh AP, Puri SK, Chitnis CE. Antibodies raised against receptor-binding domain of Plasmodium knowlesi Duffy binding protein inhibit erythrocyte invasion. Mol Biochem Parasitol. 2002;121:21–31. doi: 10.1016/s0166-6851(02)00017-8. [DOI] [PubMed] [Google Scholar]
- Singh AP, Ozwara H, Kocken CHM, Puri SK, Thomas AW, Chitnis CE. Targeted deletion of Plasmodium knowlesi Duffy binding protein confirms its role in junction formation during invasion. Mol Microbiol. 2005;55:1925–1934. doi: 10.1111/j.1365-2958.2005.04523.x. [DOI] [PubMed] [Google Scholar]
- Singh B, Kim Sung L, Matusop A, Radhakrishnan A, Shamsul SS, Cox-Singh J, et al. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet. 2004;363:1017–1024. doi: 10.1016/S0140-6736(04)15836-4. [DOI] [PubMed] [Google Scholar]
- Singh B, Daneshvar C. Human infection and detection of Plasmodium knowlesi. Clin Microbiol Rev. 2013;26:165–184. doi: 10.1128/CMR.00079-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Straimer J, Lee MCS, Lee AH, Zeitler B, Williams AE, Pearl JR, et al. Site-specific genome editing in Plasmodium falciparum using engineered zinc-finger nucleases. Nat Meth. 2012;9:993–998. doi: 10.1038/nmeth.2143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trager W, Jensen J. Human malaria parasites in continuous culture. Science. 1976;193:673–675. doi: 10.1126/science.781840. [DOI] [PubMed] [Google Scholar]
- Trigg PI. A new continuous perfusion technique for thecultivation of malaria parasites in vitro. Trans R Soc Trop Med Hyg. 1968;62:371–378. doi: 10.1016/0035-9203(68)90088-6. [DOI] [PubMed] [Google Scholar]
- van der Wel AM, Tomás AM, Kocken CHM, Malhotra P, Janse CJ, Waters AP, et al. Transfection of the primate malaria parasite Plasmodium knowlesi using entirely heterologous constructs. J Exp Med. 1997;185:1499–1503. doi: 10.1084/jem.185.8.1499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van der Wel AM, Kocken CHM, Pronk TC, Franke-Fayard B, Thomas AW. New selectable markers and single crossover integration for the highly versatile Plasmodium knowlesi transfection system. Mol Biochem Parasitol. 2004;134:97–104. doi: 10.1016/j.molbiopara.2003.10.019. [DOI] [PubMed] [Google Scholar]
- Voorberg-van der Wel A, Zeeman AM, van Amsterdam SM, van den Berg A, Klooster EJ, Iwanaga S, et al. Transgenic fluorescent Plasmodium cynomolgi liver stages enable live imaging and purification of Malaria hypnozoite-forms. PLoS ONE. 2013;8:e54888. doi: 10.1371/journal.pone.0054888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vythilingam I, Noorazian YM, Huat TC, Jiram AI, Yusri YM, Azahari AH, et al. Plasmodium knowlesi in humans, macaques and mosquitoes in peninsular Malaysia. Parasit Vectors. 2008;1:26. doi: 10.1186/1756-3305-1-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vythilingam I, Tan CH, Asmad M, Chan ST, Lee KS, Sing B. Natural transmission of Plasmodium knowlesi to humans by Anopheles latens in Sarawak, Malaysia. Trans R Soc Trop Med Hyg. 2006;100:1087–1088. doi: 10.1016/j.trstmh.2006.02.006. [DOI] [PubMed] [Google Scholar]
- Warren M, Cheong WH, Fredericks HK, Coatney GR. Cycles of jungle malaria in West Malaysia. Am J Trop Med Hyg. 1970;19:383–393. doi: 10.4269/ajtmh.1970.19.383. [DOI] [PubMed] [Google Scholar]
- Wharton RH, Eyles DE. Anopheles hackeri, a vector of Plasmodium knowlesi in Malaya. Science. 1961;134:279–280. doi: 10.1126/science.134.3474.279. [DOI] [PubMed] [Google Scholar]
- Wickham JM, Dennis ED, Mitchell GH. Long term cultivation of a simian malaria parasite (Plasmodium knowlesi) in a semi-automated apparatus. Trans R Soc Trop Med Hyg. 1980;74:789–792. doi: 10.1016/0035-9203(80)90202-3. [DOI] [PubMed] [Google Scholar]
- William T, Rahman HA, Jelip J, Ibrahim MY, Menon J, Grigg MJ, et al. Increasing incidence of Plasmodium knowlesi malaria following control of P. falciparum and P. vivax Malaria in Sabah, Malaysia. PLoS Negl Trop Dis. 2013;7:e2026. doi: 10.1371/journal.pntd.0002026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeeman A-M, Voorberg-van der Wel A, Kocken CHM. Ex vivo culture of Plasmodium vivax and Plasmodium cynomolgi and in vitro culture of Plasmodium knowlesi blood stages. Methods Mol Biol. 2013;923:35–49. doi: 10.1007/978-1-62703-026-7_4. [DOI] [PubMed] [Google Scholar]