

Abstract
The yeast Wickerhamomyces anomalus has been investigated for several years for its wide biotechnological potential, especially for applications in the food industry. Specifically, the antimicrobial activity of this yeast, associated with the production of Killer Toxins (KTs), has attracted a great deal of attention. The strains of W. anomalus able to produce KTs, called “killer” yeasts, have been shown to be highly competitive in the environment. Different W. anomalus strains have been isolated from diverse habitats and recently even from insects. In the malaria mosquito vector Anopheles stephensi these yeasts have been detected in the midgut and gonads. Here we show that the strain of W. anomalus isolated from An. stephensi, namely WaF17.12, is a killer yeast able to produce a KT in a cell-free medium (in vitro) as well as in the mosquito body (in vivo). We showed a constant production of WaF17.12-KT over time, after stimulation of toxin secretion in yeast cultures and reintroduction of the activated cells into the mosquito through the diet. Furthermore, the antimicrobial activity of WaF17.12-KT has been demonstrated in vitro against sensitive microbes, showing that strain WaF17.12 releases a functional toxin. The mosquito-associated yeast WaF17.12 thus possesses an antimicrobial activity, which makes this yeast worthy of further investigations, in view of its potential as an agent for the symbiotic control of malaria.
Introduction
Wickerhamomyces anomalus (formerly Hansenula anomala and Pichia anomala) is a Saccharomycetes yeast with wide biotechnological potential [1], traditionally used in the agro-food sector, as a biopreservation agent, suitable to improve feed and food safety. The suitability of W. anomalus as a biopreservation agent is certified by the European Food Safety Authority (EFSA), with the attribution of a qualified presumption of safety status at level-1 (QPS-1) [2]. Fields of application for this yeast include biocontrol, food fermentation, biofuel production, and the production of therapeutic molecules used in human medicine.
W. anomalus is highly tolerant to environmental stresses, and is adapted to a wide range of growth conditions, in terms of temperature (3–37°C), pH value (2–12) and osmolarity [3]. This robustness makes this yeast highly competitive in many different habitats. It has been isolated from very diverse sources, including flowering plants, fruit skins, dairy and baked food products, contaminated oil, salted foods, wastewater, marine environments, human tissues and even the gut of insects (flies, beetles and mosquitoes) [1].
An additional character displayed by W. anomalus is a wide spectrum antimicrobial activity (something very unusual in yeasts), being active against a variety of microorganisms including other yeasts, filamentous fungi and bacteria [1]. Several inhibition mechanisms have been proposed such as competition for nutrients, production of ethyl acetate and direct killing by Killer Toxins (KTs). The yeast “killer” phenomenon derives from the lethal activity of KTs secreted by self-immune killer strains on sensitive yeasts, expressing specific cell-wall KT receptors (KTRs) [4].
KTs are a group of glycoproteins with a variable molecular weight, whose activity is characterized by a large range of optimal pH and temperatures [5]. These antimicrobial properties make W. anomalus a valuable bio-control agent on different substrates; for example the inhibition of the fungus Botrytis cinerea on fruits [6], [7], and of Penicillium spp., Aspergillus spp. and Enterobacteriaceae on cereal grain [8]. The mechanism of bacterial inhibition is yet unknown, but it is likely that W. anomalus KTs (WaKTs) play a main role, as demonstrated on different Gram-negative bacteria and fungi [9], [10]. Killer activity mediated by WaKTs has been well characterized against sensible strains of fungi. For example, WaKT-mediated inhibition was demonstrated in Candida albicans both in vitro and in vivo [11]–[13]. Other studies highlighted that a KT-derived ‘killer’ peptide (KP), characterized by a candidacidal activity, aggregates on the surface of C. albicans cells, particularly on budding scars, where β1,3-glucans (the major component of the KTRs) are exposed [14]. KP has also been shown to be effective against Saccharomyces cerevisiae, including some strains resistant to conventional antifungal drugs [15], and against a wide spectrum of microbes beyond bacteria and fungi, including the protozoan parasites Leishmania infantum, L. major and Acanthamoeba castellanii [16], [17].
The mechanism of action of WaKTs is mainly based on the interaction with cell-wall carbohydrates, where β-glucanase-induced hydrolysis of glucan into glucose is demonstrated [18]. In this context, several authors have investigated the β-glucanase activity of WaKTs from different W. anomalus strains. In the strain K (successfully applied as a bio-control agent against B. cinerea on apple) and in killer strains isolated from olive brine, exoglucanase-encoding genes are responsible for the toxic exo-β-1,3-glucanase activity [19], [20]. The same WaKT-mediated enzymatic activity in the marine W. anomalus strain YF07b is directed against pathogenic fungi of a crab [21]. Thus, arthropod-associated W. anomalus could exert a protective function against pathogens, in addition to the already demonstrated nutritional role (such as the case of the mutualistic yeast-beetle association with a nutritional dependence in the coleopteran Doubledaya bucculenta [22]). Among insects, W. anomalus has also been detected in the inner body of Drosophila sp.[23] and in mosquitoes of public health importance [24]. Particularly, it has been shown to be associated with the malaria vector Anopheles stephensi, where the yeast is present in the midgut as well as in the gonads [25].The association between Anopheles mosquitoes and W. anomalus deserves to be further investigated, in order to explore the possibility that this yeast could be developed as an agent for the symbiotic control (SC) of malaria [26], in addition to other candidates for SC, such as the bacteria Asaia spp. and Wolbachia, and the fungus Metarhizium robertsii [27]–[29].The objective of this work is to investigate whether the W. anomalus strain isolated from An. stephensi displays killer activity against model microorganisms, and whether it produces a KT antimicrobial molecule within the mosquito body.
Materials and Methods
Yeast Strains
Five strains belonging to the yeast species W. anomalus, C. albicans and S. cerevisiae have been used: (i) WaF17.12 isolated from An. stephensi mosquitoes [25], (ii) WaATCC 96603, (iii) WaUM3, (iv) CaATCC 24433 and (v) ScATCC 2601. WaATCC 96603 and WaUM3 were used as reference strains, since they are respectively WaKT-producing (WaKT-resistant) and WaKT-non-producing (WaKT-susceptible) strains [30], [31]. The strain WaUM3 was used both as negative control in the experiments aimed at studying the expression of WaKT and susceptible target in antimicrobial assays. CaATCC 24433 and ScATCC 2601 were selected as additional WaKT-susceptible targets [32]–[34].
Mosquitoes
The experiments were performed using a colony of An. stephensi (Liston) reared at University of Camerino (Italy). Mosquitoes were maintained at standard aseptic conditions of 30°C and 95% humidity. Larvae were grown in tanks filled with water containing sterile minced commercial mouse food. The adults were fed with 5% sterile sugar solution. Specific diet for IFA assays is described in a paragraph below.
Production of Crude WaKT Extracts
The strains WaF17.12, WaATCC 96603 and WaUM3 were grown in culture conditions suitable to stimulate the production of soluble toxins in liquid medium, as reported by İzgü et al. (2006) [33]: YPD broth (20 g/l peptone, 20 g/l glucose, 10 g/l yeast extract), buffered at pH 4.5 or pH 8 with 0.1 M citric acid and 0.2 M K2HPO4, and incubated at 26°C for 36 h at 70 rpm [32]. The yeast cells were removed by centrifugation at 2000 g 4°C for 10 min. The supernatants were filtered and concentrated 100-fold using Pierce Concentrators (Thermo Fisher Scientific Inc, Waltham, Massachusetts, USA) with a cut off of 9 kDa, by centrifugation at 4°C for 5 h at 2000 g.
Western Blot Analysis of WaKT Crude Extracts
The crude WaKT extracts from WaF17.12, WaATCC 96603 and WaUM3 were analyzed by electrophoresis in 7% polyacrylamide gel, at 100 Volts for 4 h in a minigel system (Bio-Rad Laboratories, Richmond, Calif., USA). Proteins were electrically transferred to a polyvinylidene difluoride (PVDF) membrane at 30 Volts overnight. PVDF membrane was incubated for 1 h at room temperature with 3% non-fat powdered milk in phosphate-buffered saline (PBS) at pH 7.2 and 2% tween-20 (PBS-T). Subsequently, the membrane was incubated at 37°C for 1 h with a 1∶50 dilution of monoclonal antibody (mAb) KT4 produced against a WaKT of W. anomalus ATCC 96603 [14], [30] in PBS-T, washed three times in PBS-T and incubated for 1 h at room temperature with a secondary, peroxidase-conjugated, anti-mouse Ab. After three washings with PBS-T, the membrane was incubated 1 min with the proper substrate (Ablotlite, Euroclone, Italy) and detected by ChemiDoc 2000R (Kodak, USA).
WaKT Activity Assays
The tests to evaluate the toxic activity of WaF17.12 on susceptible yeast strains were preliminary performed using supernatants from W. anomalus cultures against S. cerevisiae. WaF17.12, WaATCC 96603 (positive control) and WaUM3 (negative control) were grown at conditions that stimulate WaKT production, as previously described (see paragraph on the Production of crude WaKT extracts), at pH 4.5. Then the three supernatants were filtered and used as growing-media for the yeast strain ScATCC 2601. S. cerevisiae was inoculated in fresh YPD medium pH 4.5 for normal growth control. After an overnight incubation at 26°C and 70 rpm, optical densities (OD) were measured by a NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific Inc, Waltham, Massachusetts, USA).
An antimicrobial assay was carried out on solid medium plate against Asaia sp. (α-proteobacterium, Gram-negative), isolated from An. stephensi mosquito [27]. An overnight Asaia culture, grown in liquid GLY medium (25 g/l glycerol, 10 g/l yeast extract, pH 5) at 30°C at 100 rpm, was diluted to 0.1 OD (107 cells/ml) in sterile water and seeded on GLY agar plate (25 g/l glycerol, 10 g/l yeast extract, 20 g/l agar pH 5). In accordance with Lopes et al. (2010) [35], five microliters of three dilutions (109, 108 and 107 cell/ml) of a WaF17.12 yeast culture, grown in stimulating conditions, were spotted in duplicate and the plates were incubated for 72 h at 20°C. Five microliters of a WaUM3 culture (109 cell/ml) were spotted as negative control. As positive control, the same experiment was performed seeding ScATCC 2601 (susceptible strain) on a YPD plate (10 g/l yeast extract, 20 g/l bacterial peptone, 20 g/l glucose, 20 g/l agar, 0.03 g/l methylene blue). After 72 h, plates were observed for the presence of inhibition halo around the spots.
Further killer activity tests of strain WaF17.12 were carried out against the susceptible strains CaATCC 24433 and WaUM3, using WaKT crude extracts obtained from WaF17.12, WaATCC 96603 (positive control) and WaUM3 (negative control) at different pH conditions (4.5 and 8); these WaKT crude extracts were prepared as described above. The sensitive strains were grown overnight in YPD medium buffered at pH 4.5 (CaATCC 24433 or WaUM3) or pH 8 (WaUM3) with 0.1 M citric acid and 0.2 M K2HPO4. Cells (105 per well) of each susceptible strain were spotted in a 96-well microtiter plate and incubated with the three KT crude extracts at a final concentration of 50X or 19X in YPD medium. To control the normal growth of the susceptible strains, cells were incubated with fresh YPD and free medium was spotted in additional wells to rule out a possible environmental contamination. After 12 h of incubation at 26°C, the toxic activity of crude extracts against the susceptible strains was evaluated by checking cell morphology, cell density (established by OD measure) and by analysing the cellular vitality through a trypan blue assay on a Neubauer counting chamber using a 40X optical microscope objective (Carl Zeiss Axio Observer.Z1, Milan, Italy).
Statistical Analysis
All the above assays with crude extracts (50X or 19X) on the susceptible strains CaATCC 24433 and WaUM3 were performed in triplicates and vital cells/ml were counted twice. The analysed data are thus those that derived from three cell counts for each assay (using the means of the two counts for each assay). The data from the cell counts were analyzed using the software GraphPad (available at http://www.graphpad.com/) and statistical analysis was performed using One Way ANOVA followed by Bonferroni’s Multiple Comparison Tests. Statistical significance is expressed as a p-value, p<0.05.
Analysis of WaKT Expression by In vitro and In vivo Immunofluorescence Assays (IFA)
WaKT expression was tested by IFA using mAbKT4 both in vitro on cultivated yeasts and in vivo within the mosquito body. Strains WaF17.12, WaATCC 96603 (positive control) and WaUM3 (negative control) were grown at the conditions that stimulate toxin production (see paragraph on the Production of crude WaKT extracts), at pH 4.5. Cells were then centrifuged at 2000 g for 10 min at 4°C and washed twice in PBS 1X, for the in vitro assay, or three times in 9 g/l NaCl and resuspended in 50 g/l sucrose solution at a concentration of 108 cells/ml to be added to the diet of newly emerged adult mosquitoes, for in vivo assay. In the latter experiment four cages were prepared containing mosquitoes fed with: i) sterile sugar solution, ii) sugar solution added with stimulated WaATCC 96603 cells, iii) sugar solution added with stimulated WaF17.12 cells or iv) sugar solution added with stimulated WaUM3 cells. After two days of feeding, the mosquitoes were moved in new cages and fed with sterile sucrose solution only. Hence, 10, 20, and 27 days post yeasts introduction mosquitoes were analyzed. Around 60 organs (midguts and gonads) were dissected and analyzed per each time point (total 171).
For IFA, cultivated yeast cells and dissected mosquito organs were fixed on slides with 4% paraformaldehyde for 10 min at 4°C and washed twice with PBS 1X. Organs were permeabilized with 0.5% Triton X-100 in PBS 1X for 10 min at room temperature and washed three times with PBS 1X. The slides were incubated in 1% BSA in PBS 1X for 30 min at room temperature and, successively, for 1 h at 37°C in mAbKT4 diluted 1∶1000 in PBS 1X. After three washings in PBS 1X, they were incubated for 30 min at 37°C with anti-mouse IgG Alexa Fluor 594 conjugate (Invitrogen, Life Technologies) diluted 1∶100 in PBS 1X and washed three times for 10 min with PBS 1X. Slides were mounted with glycerol and visualized by an epifluorescent microscope (Carl Zeiss Axio Observer.Z1, Milan, Italy).
Immunohistochemistry
Five mosquitoes from cages i, ii, iii and iv were analyzed for the immunolocalization of WaKT. The abdominal segments were excised and immediately fixed in 4% buffered formalin solution for 2 h at room temperature. After dehydration in a graded ethanol series, the samples were cleared in xylene and embedded in paraffin at 56–58°C. Sections (5 micron thick) were mounted on gelatin-coated slides, rehydrated and processed for immunohistochemical staining using the Vectastain ABC kit reagents (Vector Laboratories, Burlingame, CA). After inactivation of the endogenous peroxidase (0.3% H2O2 in methanol for 30 min) and blocking of endogenous avidin-binding activity (avidin-biotin blocking kit; Vector Laboratories), sections were incubated for 30 min in normal goat serum diluted 1∶5 with 1% bovine serum albumin (BSA; Sigma, St. Louis, MO) in 0.05 M phosphate-buffered saline (PBS), pH 7.6. Incubation of sections with the monoclonal antibody mAbKT4 was performed at dilutions from 1∶100 to 1∶2,000 in PBS containing 1% BSA overnight at room temperature, in a humid chamber. After washing, sections were incubated with goat biotinylated anti-mouse IgG (1∶200 in PBS) for 45 min, followed by washing in PBS and treatment with the avidin-biotin-peroxidase complex (1∶100) for 45 min. After washing in PBS, the immunoreactive polypeptides were visualized in all sections by incubation for 4 min with VIP substrate (Vector Laboratories) and the reaction was stopped by rinsing with tap water. Finally, sections were dehydrated and mounted in Eukitt. Immunohistochemical controls were carried out by incubation with preimmune serum or with PBS plus 1% BSA, in place of the primary antibody. Analysis has been performed using a photomicroscope Leica DM2500.
Cloning and Sequencing of WaF17.12exoglucanase-encoding Genes (EXG1 and EXG2)
Genomic DNA was extracted from a culture of the strain WaF17.12 using a JetFlex kit (Genomed, Germany). Amplifications of EXG1 and EXG2 genes were obtained using specific oligonucleotides (described in Table S1) designed on the W. anomalus BS91 EXG1 and EXG2 sequences available in the data bases (accessions: JQ734563 and JQ734566). Reaction mixtures were prepared in 25 µl using: 0.6 U of Dream Taq Polymerase (Fermentas, Thermo Fisher Scientific Inc, Waltham, Massachusetts, USA), 0.25 mMdNTPs, 1X Taq Polymerase Buffer, 0.2 µM each primer and 50 ng DNA. Reactions were run for 2 min at 95°C and cycled 30 times through 30 sec at 95°C, 30 sec at 55°C and 40 sec at 72°C. Finally, reactions were kept for 8 min at 72°C. PCR products were then resolved in 1% agarose gel stained with ethidium bromide. The fragments were cloned in T-Vector System following manufacture’s instructions (Promega, USA). The cloned fragments were sequenced after colony PCR using the plasmid primers SP6 (5′-atttaggtgacactatagaat-3′) and T7 (5′-aatacgactcactataggg-3′). The obtained sequences were firstly analysed by BLASTN. (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Sequence alignment were then generated by ClustalW (http://www.genome.jp/tools/clustalw/). Sequences of WaF17.12 genes EXG1 and EXG2 were deposited through the EMBL-Bank.
Results
Characterization of WaF17.12-KT Using Monoclonal Antibody KT4 (mAbKT4)
Western blot analysis was carried out on WaKT crude extracts from the strains WaF17.12 (from An. stephensi), WaATCC 96603 (WaKT-producing strain) and WaUM3 (WaKT-non-producing strain) cultivated at a growth condition that stimulates toxin production [32], [33]. The monoclonal antibody (mAbKT4) used for labelling was directed against a specific WaKT, produced by the reference strain WaATCC 96603 and well characterised for its wide antimicrobial activity; mAbKT4 has already been shown to cross-react with the toxin produced by other strains of killer yeasts [14], [30]. Western blot analysis revealed a single band of the same size in both WaF17.12 (mosquito strain) and WaATCC 96603 (positive control) KT crude extracts; no signal was present in the extract from WaUM3 (negative control) (see Figure S1). These results show that yeast strain WaF17.12 from An. stephensi [25] releases a protein molecule in the cultivation medium that is labelled by the monoclonal antibody directed against WaKT; this indicates that WaF17.12 likely secretes a killer toxin.
Further evidence for the capacity of WaF17.12 to produce a killer toxin was obtained by immunofluorescence assay (IFA) using mAbKT4 in vitro, on free yeast cells, and in vivo, in the mosquito. Staining with mAbKT4 produced a clear fluorescent signal onto the cell surface of free yeast cells from strains WaF17.12 and WaATCC 96603, while no signal was revealed in WaUM3 ( Figure 1A, B and C respectively). Taken together, the results of Western blot analysis and in vitro IFA on free cells are coherent, and provide evidence that strain WaF17.12, once isolated from mosquito and cultivated in stimulating conditions, produces WaKT.
Figure 1. In vitro detection of WaKT by IFA assay using mAbKT4.
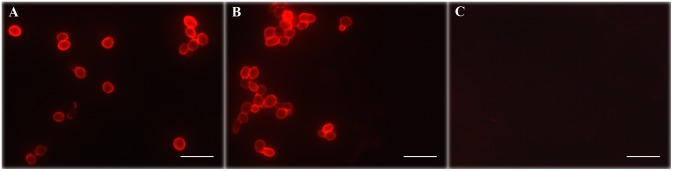
Free yeast cells of W. anomalus strains WaF17.12 (A), WaATCC 96603 (B) and WaUM3 (C) are shown. Red signal shows WaKT concentrated in the yeast cell wall layers of WaF17.12 (A) and WaATCC 96603 (B) (bars = 20 µm). Phace contrast of panels A, B and C are shown in Figure S6.
Afterwards, IFA assay was carried out in vivo in An. stephensi, checking the toxin presence in mosquito organs that are known to harbour W. anomalus [25]. Staining with mAbKT4 revealed very occasional and faint WaKT signals within the analysed guts and gonads. To investigate if the observed limited staining could derive from a scarce production of the killer toxin in the mosquito body, we carried out an in vivo IFA assay using mAbKT4 after feeding mosquitoes with a sugar meal containing yeast cells stimulated in vitro. Mosquitoes diet was supplemented for two days with WaF17.12, WaATCC 96603 (WaKT-producing strain) or WaUM3 (WaKT-non-producing strain), except a control group fed on sterile sugar solution. After each yeast treatments mosquitoes were fed on sterile sugar solution and were then dissected for the recovery of the organs (guts and gonads) at specific time points, for IFA observations. In table 1 are reported the percentages of positive samples detected on the 10th, 20th or 27th day after each yeast treatments (detailed data on male and female organs are given in table S2).
Table 1. In vivo detection of WaKTs in An. stephensi by IFA with mAbKT4.
| Days after yeast introduction | Mosquito feeding | WaKT detection %( )° |
| 10 | (i) Sterile sugar solution | 29% (5*/17) |
| (ii) Sugar solution plus WaATCC | 50% (8/16) | |
| (iii) Sugar solution plus WaF17.12 | 56% (9/16) | |
| (iv) Sugar solution plus WaUM3 | 19% (3*/16) | |
| 20 | (i) Sterile sugar solution | 60% (9*/15) |
| (ii) Sugar solution plus WaATCC | 93% (13/14) | |
| (iii) Sugar solution plus WaF17.12 | 92% (11/12) | |
| (iv) Sugar solution plus WaUM3 | 50% (8*/16) | |
| 27 | (i) Sterile sugar solution | 54% (7*/13) |
| (ii) Sugar solution plus WaATCC | 83% (10/12) | |
| (iii) Sugar solution plus WaF17.12 | 92% (11/12) | |
| (iv) Sugar solution plus WaUM3 | 42% (5*/12) | |
| Total samples analysed: 171 | ||
WaKT positive organs/total organs analyzed.
*Occasional WaKT signals were detected in these samples.
IFA analysis at different time points highlighted that both WaF17.12 and WaATCC 96603, once stimulated in culture and reintroduced in the mosquitoes, were characterized by an intense WaKT signal. In the majority of the samples analyzed, WaKT signal was detected up to four weeks after the introduction of the yeast-enriched meal ( Table 1 ). After WaF17.12 introduction, fluorescence analysis revealed the presence of an abundant WaKT signal in both female and male midguts ( Figure 2A and B ; see also immunolocalization in Figure S2) and gonads ( Figure 2C and D ). Similar staining was observed in the samples fed with the reference strain WaATCC 96603 (not shown). In contrast, in mosquitoes supplemented with WaUM3 and in the control group, the percentages of positive samples are lower than in the other mosquitoes and we observed only a few faintly stained yeast cells, that might perhaps represent resident (not introduced) yeasts. Figure 3 shows a comparison of the typical WaKT signal detected in mosquitoes fed with WaF17.12 or with sterile sugar solution only.
Figure 2. In vivo detection of WaF17.12-KT by IFA assay using mAbKT4.

Red stained yeasts are visible in the mosquito organs after feeding sugar solution supplemented with WaF17.12 cells cultivated in WaKT stimulating condition: Female (A) and male (B) guts analyzed on the 10th day after yeast introduction (bar = 200 µm), female (C) and male gonads (D) analyzed respectively on the 20th and 27th day after yeast introduction (bar = 50 µm). Negative control (gut treated with secondary antibody only) is shown in Figure S7.
Figure 3. WaF17.12-KT fluorescence signal comparison in female mosquito midguts with or without yeast introduction.

Female mosquito midguts on the 10th day after feeding with sugar solution enriched with stimulated WaF17.12 (A and B) and sterile sugar solution (C and D). Red stained yeasts are abundantly visible in (A), while only few cells are detected in the control sample (C). Images B and D (bar = 20 µm) are magnification of samples shown in A and C respectively (bar = 200 µm) White arrow in A and C indicates the section of sample enlarged in B and D.
IFA analysis also revealed some differences between guts and gonads: we observed that the WaKT signal on the 10th day was detected almost exclusively in the guts, whereas in the following two time points the number of positive gonads increased and became comparable to that of the guts (Table S2).
Significant WaKT signal was detected even in part of the offspring of WaF17.12 fed mosquitoes, suggesting a vertical transmission of the yeasts and a long-term effect of the in vitro induction of KT secretion (Figure S3).
Crude WaF17.12-KT Activity Assays
Experiments performed using yeast culture supernatants showed antimicrobial activity of WaF17.12 against the susceptible strain ScATCC 2601 [34], belonging to the species S. cerevisiae. Specifically the growth of the target yeast ScATCC 2601 was inhibited when inoculated in the liquid culture medium obtained from the supernatants of WaATCC 96603 (WaKT-producing strain) and WaF17.12 grown at condition that stimulate WaKT production, but not using supernatant from WaUM3 (WaKT-non-producing strain) (see Figure S4).
In-depth antimicrobial activity assays were carried out using crude WaKT extracts from WaF17.12, WaATCC 96603 and WaUM3 against two additional WaKT susceptible yeast strains, CaATCC 24433 [32], belonging to the species C. albicans, and WaUM3 itself [30], [31]. The results obtained using different concentrations of the crude extracts (50X and 19X) were comparable and data about the lower concentration assay are shown in figure 4 (those from the 50X concentration are not shown). Since it is reported that WaKT activity shows its maximum around acidic pH values, with a peak at pH 4.5 [21], [33], this was the pH value used in assays for both sensitive target strains ( Figure 4A and B ). To test possible killer activity at not ideal pH conditions, we performed an additional assay at pH 8 ( Figure 4C ) on WaUM3, considering this target as the most suitable one [4]. In the three assays the results of toxin activity revealed a growth inhibition of both susceptible strains when treated with WaKT crude extracts from WaF17.12 and WaATCC 96603, while no effect was detected with WaUM3 extract. More in detail, cell counts of CaATCC 24433 ( Figure 4A ) and WaUM3 ( Figure 4B and C ) showed a growth inhibition after treatment with both of the crude extracts from the two WaKT-producing strains (p<0.01, p<0.001 and p<0.01 respectively). These data confirmed that the susceptibility of the two target strains to toxic activity of WaF17.12 was comparable to that exerted by the reference strain WaATCC 96603. Importantly, significant antimicrobial activity against WaUM3 was observed also at pH 8 ( Figure 4C ) although the toxic effect appeared to be lower than those at acidic pH, in agreement with published results [33]. Bonferroni’s tests have been performed comparing the number of cells/ml of the target strains after each treatment with crude extracts from WaF17.12, or WaATCC 96603 (positive control), versus treatment with crude extract from WaUM3 (negative control).
Figure 4. Crude Wa F17.12-KT extract activity assays.

The susceptible strains CaATCC 24433 (A) and WaUM3 (B and C) were incubated for 12 h at 26°C at pH 4.5 (A and B) and 8 (C) with WaKT crude extracts (19X) from WaF17.12, WaATCC 96603 (WaKT-producing strain) or WaUM3 (WaKT non-producing strain). Crude WaKT activity was evaluated using the trypan blue test reporting the number of vital cells/ml of the two target strains and the values are reported as mean±SEM (n = 6). Asterisks refers to the statistical significance of the differences, according to the One Way ANOVA and Bonferroni’s Multiple Comparation Tests.
Since a WaKT-mediated activity on Gram-negative bacteria has been described [9], we investigated the effect of a culture WaF17.12 stimulated for KT production on the An. stephensi bacterial symbiont Asaia sp., that is known to massively inhabit the mosquito midgut and gonads [27]. The assays showed that the growth of Asaia sp. colonies in vitro was not inhibited by the WaF17.12, suggesting that this yeast does not exert its antimicrobial activity against Asaia symbionts (see Figure S5).
Sequence Analysis of WaF17.12 Exoglucanase-encoding Genes EXG1 and EXG2
The EXG1 and EXG2 complete coding genes were amplified, cloned and sequenced. We identified two Open Reading Frame (ORF) sequences of 1497 bps and 1284 bps, respectively. Corresponding amino acid sequences are of 499 residues in the case of EXG1 and 428 for EXG2.
The EXG1 and EXG2 sequences showed a 100% and 99% similarity respectively with the GenBank W. anomalus sequences JQ734563 and JQ734566 [20]. Sequences of WaF17.12 EXG1 and EXG2 genes were deposited through the EMBL-Bank under the accession numbers HG316785 (EXG1) and HG316786 (EXG2).
Discussion
The Yeast Strain WaF17.12 Isolated from Mosquito Secretes WaKT Both In vitro and In vivo
Western blot analysis of cultural crude extracts showed that strain WaF17.12, isolated from the malaria vector An. stephensi, releases a molecule in the medium that is stained by mAbKT4, a monoclonal antibody targeted on killer yeast toxins. By in vitro IFA assay on free cells, we verified that cultivated WaF17.12 yeasts also produce a molecule recognized by mAbKT4, concentrated at the surface of the yeast. This is in accordance with published results indicating toxin secretion through the cell wall in W. anomalus WaKT-producing strains [36], [37]. Overall, the above results strongly suggest that WaF17.12 yeast strain from mosquitoes produces a killer toxin.
IFA analyses also pinpointed that when cultures of WaF17.12 stimulated for toxin production were reintroduced within the mosquito by the diet, the WaKT signal was still present several weeks after the yeast reintroduction, i.e. for the entire mosquito life span (a relevant mAbKT4-staining was found even in the offspring of these mosquitoes). Particularly, the toxin signal was detected in the midgut and gonads, in both male and female mosquitoes. The WaKT signal in mosquitoes fed with WaF17.12 (or the reference strain WaATCC 96603) was quite abundant especially within the female midgut. Instead, the mosquitoes fed with sterile sugar solution or with WaUM3 showed few stained yeast cells. Reasonably, this occasional WaKT signal could derive from the presence of residing (not introduced) yeasts capable of producing the toxin (and possibly related with WaF17.12).
Crude WaKT Extract from WaF17.12 shows In vitro Antimicrobial Activity
Bio-assays demonstrated that WaF17.12 exhibits antimicrobial effects against different susceptible strains, inhibiting their growth in cultivation media. Similarly to other strains of W. anomalus that have been described to be active against C. albicans, we demonstrated that WaF17.12 exerted an antimicrobial effect in vitro on a strain from this species, CaATCC 24433. Since ascomycetous yeasts are frequently described to infect mosquitoes [24], [38], [39], future studies could investigate the possible detrimental effects of killer yeast strains against other yeasts sharing the same habitat within the insect body.
WaKT activity assays at pH 4.5 and pH 8 demonstrated that the killer activity of WaF17.12 against WaUM3 spans a wide range of pH values. This finding fits well with possible antimicrobial competition of W. anomalus in the mosquito body, since different physiological conditions can occur in vivo, for example in the different organs as well as at the intestinal level depending on the diet. Particularly, it is described that pH in the female midgut can vary around basic values after a blood meal [40], [41]. Concerning temperature, our experiments were carried out at 26°C, the suggested condition for toxic assays [32]. Nevertheless, antimicrobial tests demonstrated that the maximum WaKTs activity ranges from 18°C to 30°C and its decrement starts at around 40°C [33]. Adaptability of KT activity to different pH values and temperatures is an important requisite for potential applications of killer yeast strains in the biotechnological control of mosquito transmitted diseases. In this context, the inefficacy of WaF17.12-KT against Asaia sp. supports a feasible cohabitation of the bacteria and the yeast in the mosquito midgut and gonads, suggesting the possible synergistic use of W. anomalus in combination with other microbial candidates for SC.
Exoglucanase-encoding Genes as Possible Mediators of W. anomalus Killer Activity
Antimicrobial activity of toxins produced by killer yeasts is well documented in literature [21], [42], [43], while little is known about the genetics of this antimicrobial phenotype. Some indications suggested that WaKTs are chromosomally inherited [44], but the chromosomal localization of the genes is still missing [5]. Of particular interest was the identification of genes responsible for the synthesis of exo- β-1,3-glucanase [19] and the characterization of its enzymatic activity, adapted to a wide range of temperature and acidic pH [45]–[47]. This activity is responsible for the cleavage of the β-glycosidic bond of β-glucan, a major cell wall component of yeasts, [33]. Here, we reported the identification of two genes in the mosquito yeast strain WaF17.12, EXG1 and EXG2, that encode for exo-1,3-β-glucanases. Nevertheless hypothesis of a possible association between antimicrobial phenotype of WaF17.12with exo-β-1,3-glucanase activity needs further characterization of these genes. Particularly, the amplification of both EXG-genes also in the non-killer strain WaUM3 lays the basis for later studies encompassing possible polymorphisms and differences in the transcriptional activity of these genes in KT-producing and non KT-producing strains. A clarification of this aspect may contribute to explain the molecular basis of the killer phenomenon.
Conclusions and Prospects for Future Applications
The results here reported show that mosquito yeast strain WaF17.12: (i) secretes a molecule recognized by a monoclonal antibody specific for killer yeast toxins; (ii) possesses antimicrobial activity against sensitive yeast strains/species. This allows us to conclude that the mosquito yeast WaF17.12 can be regarded as a killer yeast, capable of producing a killer toxin. We would thus indicate this toxin as WaF17.12-KT.
W. anomalus yeasts had been detected in mosquitoes a few years ago, but their biological role in these insects is still unknown [24], [25]. Arthropod-associated W. anomalus strains could exert a nutritional role [22] and/or a protective function mediated by antimicrobial activity against pathogens [48], and this led us to hypothesize that W. anomalus might play similar roles also in mosquitoes [24], [25]. The novel finding that the strain isolated from An. stephensi displays an antimicrobial phenotype supports the hypothesis of a protective function. The protective role of symbiotic microorganisms against pathogens in different arthropod species is emerging as a promising research field, with a high potential for future applications toward the control of vector-borne diseases [29], [49]. However, most of the papers so far published in this field deal with bacterial symbionts, and the work here presented provides the first evidence for the presence of a yeast expressing a killer phenotype in mosquitoes.
The presence of a W. anomalus killer strain in the female mosquito midgut is quite interesting, considering that the gut represents a numeric bottleneck during the life cycle of Plasmodium, and this could make the parasites more vulnerable to the action of antagonistic microorganisms [50]. Future studies should thus investigate whether KT-producing strains of W. anomalus possess anti-Plasmodium activity, and whether enough toxin is produced by the yeasts naturally present in the mosquito gut. On the other hand, our results show that cultivated yeasts can easily colonize mosquitoes, and then continue with toxin production for at least four weeks. Killer yeasts could also be selected for an increased secretion of KT at conditions that simulates the mosquito gut, and for more effective anti-malaria activity. A possible association between antimicrobial phenotype of KT-producing strains with exo-β-1,3-glucanase seems very interesting, particularly considering that the enzyme target carbohydrate (β-1,3-glucan) is already known to be present in some parasites oocyst walls as Toxoplasma and Eimeria [51] and implicated in the immunoprotection against Plasmodium berghei infection in mice [52].
Yeasts naturally effective against malaria (or even after selection for increased activity) would likely be more easy to be accepted by the public opinion for field applications, compared to the genetically modified microorganisms foreseen by the so-called paratransgenesis approach [53].
Data Accessibility
DNA sequences are accessible at http://www.ebi.ac.uk/ena/data/view/HG316785-HG316786.
Supporting Information
Western blot analysis of Wa KT crude extracts. WaF17.12, WaATCC 96603 (WaKT-producer strain) and WaUM3 (WaKT-not-producer strain) crude extracts, obtained after concentration of supernatants from yeast cultures stimulated for WaKT secretion, were analyzed using mAbKT4. (1) Standard molecular weight marker expressed in kDa and WaKT crude extracts from (2) WaUM3, (3) WaATCC 96603 and (4) WaF17.12. A single band at around 250 kDa was revealed in extracts from WaATCC 96603 and WaF17.12.
(DOC)
Immunolocalization of Wa F17.12-KT in An. Stephensi abdominal sections. Abdominal histological sections of adult female mosquitoes treated with sterile sugar solution (A) or with sugar solution enriched with stimulated yeast cultures of WaF17.12 (B). White asterisk and black arrow in (B) indicate positive staining surrounding the intestinal epithelium and the presence of WaF17.12 cells in the midgut lumen (ML), respectively. WaF17.12-KT signal in panel (B) is shown in the mosquito gut on the 10th day post yeast treatment. mAbKT4 is able to recognize both the WaF17.12-KT on the surface of the yeast cell and in soluble form, as demonstrated by IFA and Western blot analysis, respectively. Interestingly, the staining obtained by immunohistochemistry appears to localize on both yeast cells surface and intestinal epithelium, supporting the hypothesis that WaF17.12-KT is secreted in vivo and present in the gut lumen.
(DOC)
In vivo detection of Wa F17.12-KT in mosquito offspring by IFA assay using mAbKT4. Red stained yeasts are visible (white arrow) in the female mosquito midgut (F1 generation from parental mosquitoes fed for two days with sugar solution plus stimulated WaF17.12 cells) (bar = 50 µm).
(DOC)
Antimicrobial activity of Wa F17.12 against S. cerevisiae . The susceptible strain ScATCC 2601 was inoculated in fresh YPD medium (A) or in filtered broth culture surnatants of the strains WaUM3 (B), WaATCC 96603 (C) and WaF17.12 (D) grown at conditions that stimulate WaKT production. After an overnight incubation at 26°C and 70 rpm, optical densities (OD) of each ScATCC 2601 culture were measured. ScATCC 2601 growth rates compared to growth control (A) showed to be slightly affected in (B), whereas there is a strong decrease in (C) and (D). (A): 5.97 OD (2×108 cells/ml*); (B): 3.83 OD (1.3×108 cells/ml*); (C): 1.04 OD (3.5×107 cells/ml*); (D): 1.04 OD (3.5×107 cells/ml*). *Concentration (cells/ml) of ScATCC 2601 has been evaluated by the assessment of a standard growth curve.
(DOC)
Growth inhibition evaluation of Wa F17.12-KT against S. cerevisiae and Asaia sp. Cultures of ScATCC 2601 (A) and Asaia sp. (B) were seeded on YPD agar and GLY agar, respectively. The plates were incubated for 72 h at 20°C with three dilution of the activated WaF17.12 culture: 109 cells/ml (1 and 4), 108 cells/ml (2 and 5) and 107 cells/ml (3 and 6). WaUM3 culture (109 cell/ml) was spotted as negative control (N). A growth inhibition halo is distinguishable surrounding the yeast colonies only in the plate A (black arrow). The presence of a pink ring of Asaia sp. around the yeast colonies (black arrow) can be observed in plate B.
(DOC)
In vitro detection of Wa KT by IFA assay using mAbKT4 (phase contrast of Fig. 1 in the main test). Free yeast cells of W. anomalus strains WaF17.12 (A), WaATCC 96603 (B) and WaUM3 (C) phase contrast images corresponding to images of Fig. 1 (A), (B) and (C) respectively.
(DOC)
In vivo detection of Wa F17.12-KT by IFA assay using mAbKT4 (negative control for IFA experiments). Female gut from mosquito fed with cultivated WaF17.12 after treatment with secondary antibody only.
(DOC)
Oligonucleotides used in EXG1 and EXG2 genes molecular analysis. List of primers used in EXG genes sequence analysis.
(DOC)
Detection of Wa KT signal in mosquito organs by IFA with mAbKT4. Four cages containing mosquitoes fed with: (i) Sterile sugar solution, (ii) sugar solution enriched with WaATCCC 96603 (WaKT-producing strain), (iii) sugar solution enriched with WaF17.12 and (iv) sugar solution enriched with WaUM3 (WaKT non-producing strain). Organs were dissected 10, 20 and 27 days after yeast introduction and analyzed using a fluorescence microscope.
(DOC)
Acknowledgments
We would like to thank Tiziana Zucconi Galli Fonseca for editing the English usage in the manuscript.
Funding Statement
The research leading to these results has received funding from the European Union Seventh Framework Programme ([FP7/2007-2013] [FP7/2007-2011]) under grant agreement n° 281222 to IR and from Prin 2009 (grant 2009L27YC8_003), from the Italian Ministry of Education, University and Research (MIUR) and by the EU-FP7 Capacities-Infrastructure 2008 (grant 228421) to GF. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Walker GM (2011) Pichia anomala: cell physiology and biotechnology relative to other yeasts. Antonie Van Leeuwenhoek 99: 25–34. [DOI] [PubMed] [Google Scholar]
- 2. Sundh I, Melin P (2010) Safety and regulation of yeasts intentionally added to the food or feed chains. Antonie van Leeuwenhoek 99: 113–119. [DOI] [PubMed] [Google Scholar]
- 3. Fredlund E, Druvefors U, Boysen ME, Lingsten KJ, Schnurer J (2002) Physiological characteristics of the biocontrol yeast Pichia anomala J121. FEMS Yeast Res 2: 395–402. [DOI] [PubMed] [Google Scholar]
- 4.Bevan EA, Makover M (1963) The physiological basis of the killer character in yeasts. In: Geert SJ editor. Genetics Today XI International Congress on Genetics. Oxford Pergamon Press, Oxford. Vol 1, 53–58.
- 5. Passoth V, Fredlund E, Druvefors UA, Schnurer J (2006) Biotechnology, physiology and genetics of the yeast Pichia anomala . FEMS Yeast Res 6: 3–13. [DOI] [PubMed] [Google Scholar]
- 6. Friel D, Pessoa NM, Vandenbol M, Jijakli MH (2007) Separate and combined disruptions of two exo-beta-1,3-glucanase genes decrease the efficiency of Pichia anomala (strain K) biocontrol against Botrytis cinerea on apple. Mol Plant Microbe Interact 20: 371–379. [DOI] [PubMed] [Google Scholar]
- 7. Jijakli MH (2011) Pichia anomala in biocontrol for apples: 20 years of fundamental research and practical applications. Antonie Van Leeuwenhoek 99: 93–105. [DOI] [PubMed] [Google Scholar]
- 8. Olstorpe M, Passoth V (2011) Pichia anomala in grain biopreservation. Antonie Van Leeuwenhoek 99: 57–62. [DOI] [PubMed] [Google Scholar]
- 9. Polonelli L, Morace G (1986) Reevaluation of the yeast killer phenomenon. J ClinMicrobiol 24: 866–869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Passoth V, Schnürer J (2003) Non-conventional yeasts in antifungal application. In: de Winde JH editor. Functional genetics of industrial yeasts. Springer Verlag, Berlin.pp. 297–329.
- 11. Sawant AD, Abdelal AT, Ahearn DG (1988) Anti-Candida albicans activity of Pichia anomala as determined by a growth rate reduction assay. Appl Environ Microbiol 54: 1099–1103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Buzzini P, Martini A (2001) Discrimination between Candida albicans and other pathogenic species of the genus Candida by their differential sensitivities to toxins of a panel of killer yeasts. J ClinMicrobiol 39: 3362–3364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Polonelli L, Conti S, Gerloni M, Campani L, Mantovani MP, et al. (1990) Production of yeast killer toxin in experimentally infected animals. Mycopathologia 110: 169–175. [DOI] [PubMed] [Google Scholar]
- 14. Polonelli L, Magliani W, Ciociola T, Giovati L, Conti S (2011) From Pichia anomala killer toxin through killer antibodies to killer peptides for a comprehensive anti-infective strategy. Antonie Van Leeuwenhoek 99: 35–41. [DOI] [PubMed] [Google Scholar]
- 15. Conti S, Magliani W, Giovati L, Libri I, Maffei DL, et al. (2008) Screening of a Saccharomyces cerevisiae nonessential gene deletion collection for altered susceptibility to a killer peptide. New Microbiol 31: 143–145. [PubMed] [Google Scholar]
- 16. Savoia D, Scutera S, Raimondo S, Conti S, Magliani W, et al. (2006) Activity of an engineered synthetic killer peptide on Leishmania major and Leishmania infantum promastigotes. ExpParasitol 113: 186–192. [DOI] [PubMed] [Google Scholar]
- 17. Fiori PL, Mattana A, Dessi D, Conti S, Magliani W, et al. (2006) In vitro acanthamoebicidal activity of a killer monoclonal antibody and a synthetic peptide. J AntimicrobChemother 57: 891–898. [DOI] [PubMed] [Google Scholar]
- 18. Magliani W, Conti S, Giovati L, Maffei DL, Polonelli L (2008) Anti-beta-glucan-likeimmunoprotectivecandidacidalantiidiotypicantibodies. Front Biosci 13: 6920–6937. [DOI] [PubMed] [Google Scholar]
- 19. Grevesse C, Lepoivre P, Jijakli MH (2003) Characterization of the Exoglucanase-Encoding Gene PaEXG2 and Study of Its Role in the Biocontrol Activity of Pichia anomala Strain K. Phytopathology. 93: 1145–1152. [DOI] [PubMed] [Google Scholar]
- 20. Muccilli S, Wemhoff S, Restuccia C, Meinhardt F (2013) Exoglucanase-encoding genes from three Wickerhamomyces anomalus killer strains isolated from olive brine. Yeast 30: 33–43. [DOI] [PubMed] [Google Scholar]
- 21. Wang X, Chi Z, Yue L, Li J (2007) Purification and characterization of killer toxin from a marine yeast Pichia anomala YF07b against the pathogenic yeast in crab. CurrMicrobiol 55: 396–401. [DOI] [PubMed] [Google Scholar]
- 22. Toki W, Tanahashi M, Togashi K, Fukatsu T (2012) Fungal farming in a non-social beetle. PLoS One 7: e41893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Zacchi L, Vaughan-Martini A (2002) Yeasts associated with insects in agricultural areas of Perugia, Italy. Ann. Microbiol52: 237–244. [Google Scholar]
- 24. Ricci I, Mosca M, Valzano M, Damiani C, Scuppa P, et al. (2011a) Different mosquito species host Wickerhamomyces anomalus (Pichia anomala): perspectives on vector-borne diseases symbiotic control. Antonie Van Leeuwenhoek 99: 43–50. [DOI] [PubMed] [Google Scholar]
- 25. Ricci I, Damiani C, Scuppa P, Mosca M, Crotti E, et al. (2011b) The yeast Wickerhamomyces anomalus (Pichia anomala) inhabits the midgut and reproductive system of the Asian malaria vector Anopheles stephensi . Environ Microbiol 13: 911–921. [DOI] [PubMed] [Google Scholar]
- 26. Wang S, Jacobs-Lorena M (2013) Genetic approaches to interfere with malaria transmission by vector mosquitoes. Trends Biotechnol 31: 185–193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Favia G, Ricci I, Damiani C, Raddadi N, Crotti E, et al. (2007) Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc Natl Acad Sci U S A 104: 9047–9051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Fang W, Vega-Rodriguez J, Ghosh AK, Jacobs-Lorena M, Kang A, et al. (2011) Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 331: 1074–1077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Bian G, Joshi D, Dong Y, Lu P, Zhou G, et al. (2013) Wolbachia invades Anopheles stephensi populations and induces refractoriness to Plasmodium infection. Science 340: 748–751. [DOI] [PubMed] [Google Scholar]
- 30. Polonelli L, Morace G (1987) Production and characterization of yeast killer toxin monoclonal antibodies. J ClinMicrobiol 25: 460–462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Polonelli L, Seguy N, Conti S, Gerloni M, Bertolotti D, et al. (1997) Monoclonal yeast killer toxin-like candidacidal anti-idiotypic antibodies. ClinDiagn Lab Immunol 4: 142–146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Guyard C, Seguy N, Cailliez JC, Drobecq H, Polonelli L, et al. (2002) Characterization of a Williopsissaturnus var. mrakii high molecular weight secreted killer toxin with broad-spectrum antimicrobial activity. J Antimicrob Chemother 49: 961–971. [DOI] [PubMed] [Google Scholar]
- 33. İzgü F, Altınbay D, Acun T (2006) Killer toxin of Pichia anomala NCYC 432; purification, characterization and its exo-β-1,3-glucanase activity. Enzyme MicrobTechnol39: 669–676. [Google Scholar]
- 34. Magliani W, Conti S, Maffei DL, Ravanetti L, Polonelli L (2007) Antiidiotype-derived killer peptides as new potential tools to combat HIV-1 and AIDS-related opportunistic pathogens. Antiinfect Agents Med Chem6: 263–272. [Google Scholar]
- 35. Lopes CA, Sangorrín MP (2010) Optimization of killer assays for yeast selection protocols. Rev Argent Microbiol. 42: 298–306. [DOI] [PubMed] [Google Scholar]
- 36. Cailliez JC, Gerloni M, Morace G, Conti S, Cantelli C, et al. (1992) Ultrastructural immunodetection of a Pichia anomala killer toxin: a preliminary study. Biol Cell 75: 19–23. [DOI] [PubMed] [Google Scholar]
- 37. Cailliez JC, Cantelli C, Seguy N, Conti S, Gerloni M, et al. (1994) Killer toxin secretion through the cell wall of the yeast Pichia anomala . Mycopathologia 126: 173–177. [DOI] [PubMed] [Google Scholar]
- 38. Gusmão DS, Santos AV, Marini DC, Russo Ede S, Peixoto AM, et al. (2007) First isolation of microorganisms from the gut diverticulum of Aedes aegypti (Diptera: Culicidae): new perspectives for an insect-bacteria association. MemInst Oswaldo Cruz 102: 919–924. [DOI] [PubMed] [Google Scholar]
- 39. Gusmão DS, Santos AV, Marini DC, Bacci M Jr, Berbert-Molina MA, et al. (2010) Culture-dependent and culture-independent characterization of microorganisms associated with Aedes aegypti (Diptera: Culicidae) (L.) and dynamics of bacterial colonization in the midgut. Acta Trop 115: 275–281. [DOI] [PubMed] [Google Scholar]
- 40. Billker O, Miller AJ, Sinden RE (2000) Determination of mosquito bloodmeal pH in situ by ion-selective microelectrode measurement: implications for the regulation of malarial gametogenesis. Parasitology 120: 547–551. [DOI] [PubMed] [Google Scholar]
- 41. del Pilar Corena M, VanEkeris L, Salazar MI, Bowers D, Fiedler MM, et al. (2005) Carbonic anhydrase in the adult mosquito midgut. J ExpBiol 208: 3263–3273. [DOI] [PubMed] [Google Scholar]
- 42. Peng Y, Chi ZM, Wang XH, Li J (2009) Purification and molecular characterization of exo-beta-1,3-glucanases from the marine yeast Williopsis saturnus WC91–2. Appl Microbiol Biotechnol 85: 85–94. [DOI] [PubMed] [Google Scholar]
- 43. Wang XX, Chi Z, Peng Y, Wang XH, Ru SG, et al. (2012) Purification, characterization and gene cloning of the killer toxin produced by the marine-derived yeast Williopsis saturnus WC91–2. Microbiol Res 167: 558–563. [DOI] [PubMed] [Google Scholar]
- 44. Kagiyama S, Aiba T, Kadowaki K, Mogi K (1988) New killer toxins of halophilic Hansenula anomala . Agric Biol Chem 52: 1–7. [Google Scholar]
- 45. Lachance MA, Villa TG, Phaff HJ (1977) Purification and partial characterization of an exo-beta-glucanase from the yeast Kluyveromyces aestaurii . Can J Biochem 55: 1001–1006. [DOI] [PubMed] [Google Scholar]
- 46. Molina M, Cenamor R, Sanchez M, Nombela C (1989) Purification and some properties of Candida albicans exo-1,3-beta-glucanase. J Gen Microbiol 135: 309–314. [DOI] [PubMed] [Google Scholar]
- 47. Xu Z, Shih MC, Poulton JE (2006) An extracellular exo-beta-(1,3)-glucanase from Pichia pastoris: purification, characterization, molecular cloning, and functional expression. Protein Expr Purif 47: 118–127. [DOI] [PubMed] [Google Scholar]
- 48. Wang L, Yue L, Chi Z, Wang X (2008) Marine killer yeasts active against a yeast strain pathogenic to crab Portunus trituberculatus . Dis Aquat Organ 80: 211–218. [DOI] [PubMed] [Google Scholar]
- 49. Koehler S, Kaltenpoth M (2013) Maternal and Environmental Effects on Symbiont-Mediated Antimicrobial Defense. J Chem Ecol. 39: 978–988. [DOI] [PubMed] [Google Scholar]
- 50. Abraham EG, Jacobs-Lorena M (2004) Mosquito midgut barriers to malaria parasite development. Insect Biochem Mol Biol 34: 667–671. [DOI] [PubMed] [Google Scholar]
- 51. Bushkin GG, Motari E, Magnelli P, Gubbels MJ, Dubey JP, et al. (2012) beta-1,3-glucan, which can be targeted by drugs, forms a trabecular scaffold in the oocyst walls of Toxoplasma and Eimeria . MBio 3: e00258–12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Maheshwari R, Siddiqui MU (1989) Immunoprotection by beta-1,3 glucan antigen combination in Plasmodium berghei infection in mice. Indian J Med Res 89: 396–403. [PubMed] [Google Scholar]
- 53. Cereghino GP, Cregg JM (1999) Applications of yeast in biotechnology: protein production and genetic analysis. Curr Opin Biotechnol 10: 422–427. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Western blot analysis of Wa KT crude extracts. WaF17.12, WaATCC 96603 (WaKT-producer strain) and WaUM3 (WaKT-not-producer strain) crude extracts, obtained after concentration of supernatants from yeast cultures stimulated for WaKT secretion, were analyzed using mAbKT4. (1) Standard molecular weight marker expressed in kDa and WaKT crude extracts from (2) WaUM3, (3) WaATCC 96603 and (4) WaF17.12. A single band at around 250 kDa was revealed in extracts from WaATCC 96603 and WaF17.12.
(DOC)
Immunolocalization of Wa F17.12-KT in An. Stephensi abdominal sections. Abdominal histological sections of adult female mosquitoes treated with sterile sugar solution (A) or with sugar solution enriched with stimulated yeast cultures of WaF17.12 (B). White asterisk and black arrow in (B) indicate positive staining surrounding the intestinal epithelium and the presence of WaF17.12 cells in the midgut lumen (ML), respectively. WaF17.12-KT signal in panel (B) is shown in the mosquito gut on the 10th day post yeast treatment. mAbKT4 is able to recognize both the WaF17.12-KT on the surface of the yeast cell and in soluble form, as demonstrated by IFA and Western blot analysis, respectively. Interestingly, the staining obtained by immunohistochemistry appears to localize on both yeast cells surface and intestinal epithelium, supporting the hypothesis that WaF17.12-KT is secreted in vivo and present in the gut lumen.
(DOC)
In vivo detection of Wa F17.12-KT in mosquito offspring by IFA assay using mAbKT4. Red stained yeasts are visible (white arrow) in the female mosquito midgut (F1 generation from parental mosquitoes fed for two days with sugar solution plus stimulated WaF17.12 cells) (bar = 50 µm).
(DOC)
Antimicrobial activity of Wa F17.12 against S. cerevisiae . The susceptible strain ScATCC 2601 was inoculated in fresh YPD medium (A) or in filtered broth culture surnatants of the strains WaUM3 (B), WaATCC 96603 (C) and WaF17.12 (D) grown at conditions that stimulate WaKT production. After an overnight incubation at 26°C and 70 rpm, optical densities (OD) of each ScATCC 2601 culture were measured. ScATCC 2601 growth rates compared to growth control (A) showed to be slightly affected in (B), whereas there is a strong decrease in (C) and (D). (A): 5.97 OD (2×108 cells/ml*); (B): 3.83 OD (1.3×108 cells/ml*); (C): 1.04 OD (3.5×107 cells/ml*); (D): 1.04 OD (3.5×107 cells/ml*). *Concentration (cells/ml) of ScATCC 2601 has been evaluated by the assessment of a standard growth curve.
(DOC)
Growth inhibition evaluation of Wa F17.12-KT against S. cerevisiae and Asaia sp. Cultures of ScATCC 2601 (A) and Asaia sp. (B) were seeded on YPD agar and GLY agar, respectively. The plates were incubated for 72 h at 20°C with three dilution of the activated WaF17.12 culture: 109 cells/ml (1 and 4), 108 cells/ml (2 and 5) and 107 cells/ml (3 and 6). WaUM3 culture (109 cell/ml) was spotted as negative control (N). A growth inhibition halo is distinguishable surrounding the yeast colonies only in the plate A (black arrow). The presence of a pink ring of Asaia sp. around the yeast colonies (black arrow) can be observed in plate B.
(DOC)
In vitro detection of Wa KT by IFA assay using mAbKT4 (phase contrast of Fig. 1 in the main test). Free yeast cells of W. anomalus strains WaF17.12 (A), WaATCC 96603 (B) and WaUM3 (C) phase contrast images corresponding to images of Fig. 1 (A), (B) and (C) respectively.
(DOC)
In vivo detection of Wa F17.12-KT by IFA assay using mAbKT4 (negative control for IFA experiments). Female gut from mosquito fed with cultivated WaF17.12 after treatment with secondary antibody only.
(DOC)
Oligonucleotides used in EXG1 and EXG2 genes molecular analysis. List of primers used in EXG genes sequence analysis.
(DOC)
Detection of Wa KT signal in mosquito organs by IFA with mAbKT4. Four cages containing mosquitoes fed with: (i) Sterile sugar solution, (ii) sugar solution enriched with WaATCCC 96603 (WaKT-producing strain), (iii) sugar solution enriched with WaF17.12 and (iv) sugar solution enriched with WaUM3 (WaKT non-producing strain). Organs were dissected 10, 20 and 27 days after yeast introduction and analyzed using a fluorescence microscope.
(DOC)
Data Availability Statement
DNA sequences are accessible at http://www.ebi.ac.uk/ena/data/view/HG316785-HG316786.