

Background: Glucose activates CaMKII in β-cells, how this influences glucose homeostasis has not been determined.
Results: Inhibiting CaMKII in mouse β-cells causes glucose intolerance by reducing Ca2+ entry and insulin secretion.
Conclusion: CaMKII is a β-cell Ca2+ sensor that amplifies secretagogue-induced Ca2+ entry and insulin secretion to maintain glucose homeostasis.
Significance: This provides the first evidence that β-cell CaMKII modulates glucose homeostasis under physiological and insulin resistant states.
Keywords: Calcium Channels, Calcium Imaging, CaMKII, Diabetes, Insulin, Insulin Secretion, Islet, Pancreatic Islets
Abstract
Glucose-stimulated insulin secretion (GSIS) from pancreatic β-cells is caused by Ca2+ entry via voltage-dependent Ca2+ channels. CaMKII is a key mediator and feedback regulator of Ca2+ signaling in many tissues, but its role in β-cells is poorly understood, especially in vivo. Here, we report that mice with conditional inhibition of CaMKII in β-cells show significantly impaired glucose tolerance due to decreased GSIS. Moreover, β-cell CaMKII inhibition dramatically exacerbates glucose intolerance following exposure to a high fat diet. The impairment of islet GSIS by β-cell CaMKII inhibition is not accompanied by changes in either glucose metabolism or the activities of KATP and voltage-gated potassium channels. However, glucose-stimulated Ca2+ entry via voltage-dependent Ca2+ channels is reduced in islet β-cells with CaMKII inhibition, as well as in primary wild-type β-cells treated with a peptide inhibitor of CaMKII. The levels of basal β-cell cytoplasmic Ca2+ and of endoplasmic reticulum Ca2+ stores are also decreased by CaMKII inhibition. In addition, CaMKII inhibition suppresses glucose-stimulated action potential firing frequency. These results reveal that CaMKII is a Ca2+ sensor with a key role as a feed-forward stimulator of β-cell Ca2+ signals that enhance GSIS under physiological and pathological conditions.
Introduction
Insulin secretion from pancreatic β-cells is caused by glucose-stimulated Ca2+ entry. Although the primary insulin secretory machinery is directly stimulated by Ca2+, protein phosphorylation modulates the coupling of Ca2+ entry to insulin secretion (1). Studies in insulin-secreting cell lines and in isolated β-cells suggest that Ca2+-dependent kinases such as Ca2+/calmodulin-dependent protein kinase II (CaMKII)2 are key regulators of insulin secretion (2–5). However, the role of CaMKII in modulating GSIS in vivo under physiological and pathological conditions is poorly understood.
Glucose-stimulated activation of β-cell CaMKII temporally correlates with GSIS from isolated pancreatic islets (5). Secretagogue stimulation of Ca2+ entry activates β-cell CaMKII-enhancing GSIS, whereas inhibition of CaMKII activity blunts islet GSIS (6–8). However, these acute in vitro studies do not address the role of CaMKII in modulating islet β-cell function and glucose homeostasis in vivo (2).
Here, we investigated the physiological role of CaMKII in pancreatic β-cells using a transgenic mouse model that allows tetracycline-inducible inhibition of β-cell CaMKII activity. We find that inhibition of β-cell CaMKII significantly impairs glucose tolerance by reducing l-type Ca2+ channel (LTCC) facilitation, Ca2+ entry, and insulin secretion. These studies reveal that β-cell CaMKII is an integral Ca2+ sensor in a positive feedback pathway that regulates cytosolic and endoplasmic reticulum (ER) Ca2+ levels to enhance insulin secretion.
EXPERIMENTAL PROCEDURES
Mouse β-Cell CaMKII Inhibition
For β-cell-specific CaMKII inhibition, transgenic animals were utilized; these are a cross between C57BL/6 mice with a tetracycline transactivator expressed in β-cells via an insulin promoter (RIP-rtTA, The Jackson Laboratory, 008250) (9) and C57BL/6 mice with a CaMKII pseudosubstrate inhibitor peptide fused to GFP under the expression of a tetracycline operator-controlled promoter (EAC3I-GFP; see Fig. 1) (10). To induce expression of EAC3I-GFP these mice were treated with doxycycline (2 mg·ml−1; RPI, Mount Prospect, IL) in the drinking water for either 11 or 14 days as indicated. Controls were RIP-rtTA animals or RIP-rtTA + tetO-GFP mice (Jackson ImmunoResearch Laboratories, 018913); as indicated, both were treated with doxycline identically to the EAC3I-GFP mice. For acute inhibition of islet-cell CAMKII, C57BL/6 islets were dispersed into 20-cell clusters and then treated with the cell-permeable CAMKII inhibitor peptide, autocamtide-2 related inhibitory peptide II (AIP2), which is an Antennapedia transport peptide fused to AIP2 (EMD Millipore). Islet cells were incubated with 20 μm AIP2 in 2 mm glucose for 20 min immediately before Ca2+ imaging.
FIGURE 1.

Doxycycline induces inhibition of β-cell CaMKII activity in a transgenic mouse. A, schematic of the transgenic design of a mouse model with doxycycline-inducible β-cell CaMKII inhibition. B, epifluorescent detection of β-cell specific GFP in a pancreas of an EAC3I-GFP + RIPrtTA mouse on doxycycline for 1.5 weeks; white bar is equivalent to 20 μm. C, differential interference contrast (DIC) image of an islet cell cluster from an EAC3I-GFP + RIPrtTA mouse on doxycycline for 1.5 weeks stained for GFP (red) and insulin (green), white bar is equivalent to 10 μm. D, islet phosphosynapsin 1 (top panels) and GAPDH (lower panels) from islets expressing rtTA or rtTA + EAC3I-GFP in β-cells. E, total pancreatic insulin from RIPrtTA or EAC3I-GFP + RIPrtTA mice treated with doxycycline for 1.5 weeks (n = 4 pancreata/group; NS, nonsignificant). F, total islet insulin from RIPrtTA or EAC3I-GFP + RIPrtTA mice treated with doxycycline for 1.5 weeks (n = 4 independent islet preparations/group; NS, nonsignificant). G and H, doxycycline-treated RIPrtTA EAC3I-GFP (G) and doxycycline-treated RIPrtTA (H) pancreatic sections stained for insulin (red), glucagon (green), and nuclei (blue); white bars are equivalent to 20 μm.
Mouse Diets and Glucose Tolerance Testing
Mice were placed either on a normal chow diet or a high fat diet (HFD, 60 kcal% fat; Research Diets, Inc.) and monitored for glucose tolerance. The glucose tolerance test (GTT) was performed as described previously by injecting 2 mg/kg dextrose (animals on a normal chow diet) or 1 mg/kg (animals on a HFD) and monitoring blood glucose at the indicated time points post glucose injection (11). The mice fed a normal chow diet were treated with doxycycline at 6 weeks of age, and GTT was performed at 7.5 weeks of age. A cohort of mice was also placed on a HFD at 3 weeks of age for 2 months when GTT was performed; at this time, doxycycline was added to the drinking water for 14 days in the presence of the HFD following which another GTT was performed.
Mouse Islet and β-Cell Isolation
Islets were isolated from pancreata of mice, using collagenase digestion and Ficoll gradients as described previously (12). Islets were plated or dissociated in 0.005% trypsin, placed on glass coverslips, and cultured for 16 h in RPMI 1640 medium supplemented with 10% fetal calf serum, concentrations of glucose-specified, 100 international units ml−1 penicillin, and 100 mg ml−1 streptomycin. Dissociated β-cells were specifically used in all voltage clamp experiments recording Ca2+ currents. β-Cells on the periphery of intact islets were recorded in current clamp mode in all of the membrane potential recordings. Cells and islets were maintained in a humidified incubator at 37 °C under an atmosphere of 95% air and 5% CO2.
Western Blot Analysis
Mouse islets in groups of 50 were treated with 1 μm ionomycin for 2 min. Protein extracts were prepared from islets by extraction with SDS loading buffer (1% SDS, 30 mmol/liter Tris-HCl (pH 6.8), 5% β-mercaptoethanol, 5% glycerol, and 0.1% bromphenol blue) with protease and phosphatase inhibitors at 80 °C for 10 min. After electrophoresis through a 4–12% denaturing polyacrylamide gel, proteins were prepared as a Western blot on a nitrocellulose membrane (Bio-Rad). Anti-phosphosynapsin antibody (Santa Cruz Biotechnology) or anti-GAPDH (Rockland Immunochemicals) was used to probe the membrane at 1:250 or 1:700 dilution, respectively, in PBS, 0.1% Tween 20, and 3% powdered dried milk followed by an HRP-coupled secondary antibody (Jackson ImmunoResearch Laboratories) at 1:5000 in the same solution. The membranes were washed in PBS containing 0.1% Tween between and after antibody incubations; HRP was illuminated using Pico Signal (Pierce) and exposed on Kodak X-Omat Blue film.
Perforated Patch Electrophysiology
Patch electrodes (2–4 micro-ohms) loaded with solution containing (in mmol·liter−1) 140 KCl, 1 MgCl2[H2O]6, 10 EGTA, 10 HEPES (pH 7.25 with KOH) and the pore-forming antibiotic amphotericin B (Sigma) were used to record islet-attached β-cells (13). Islets were perfused with Krebs-Ringer-HEPES buffer containing 119 mmol·liter−1 NaCl, 2 mmol·liter−1 CaCl2, 4.7 mmol·liter−1 KCl, 10 mmol·liter−1 HEPES, 1.2 mmol·liter−1 MgSO4, 1.2 mmol·liter−1 KH2PO4, adjusted to pH 7.35 with NaOH, with the indicated concentrations of glucose and compounds. Cells on the periphery of islets were sealed in voltage clamp at −80 mV, and access was obtained over several minutes through perforations by amphotericin (13). After being switched to current clamp, cells that had a resting membrane voltage near −65 mV in 2 mm glucose and fired voltage-dependent Ca2+ channel (VDCC)-dependent action potentials post glucose-stimulation were assumed to be β-cells.
Whole Cell Voltage Clamp Electrophysiological Recordings
Voltage-activated currents were recorded using whole-cell ruptured patch clamp with an Axopatch 200B amplifier and pCLAMP10 software (Molecular Devices, Sunnyvale, CA). For calcium channel recordings, patch electrodes (2–4 micro-ohms) were loaded with intracellular solution containing the following: 102 mmol·liter−1 CsCl, 10 mmol·liter−1 tetraethylamonium chloride, 0.1 mmol·liter−1 tolbutamide, 10 mmol·liter−1 EGTA, 1 mmol·liter−1 MgCl2, 3 mmol·liter−1 Na2ATP, 5 mmol·liter−1 HEPES, pH 7.4, adjusted with CsOH. Cells were perifused with a Krebs-Ringer buffer (KRB) containing the following: 82 mmol·liter−1 NaCl, 20 mmol·liter−1 tetraethylamonium chloride, 0.1 mmol·liter−1 tolbutamide, 30 mmol·liter−1 CaCl2, 5 mmol·liter−1 CsCl, 1 mmol·liter−1 MgCl2, 0.1 mmol·liter−1 EGTA, 10 mmol·liter−1 glucose, 5 mmol·liter−1 HEPES, pH 7.4, adjusted with NaOH. For KATP channel recordings, patch electrodes were loaded with the following: 0.1 mmol·liter−1 MgATP, 140 mmol·liter−1 KCl, 1 mmol·liter−1 MgCl2, 10 mmol·liter−1 HEPES. Cells were perifused with Krebs-Ringer-HEPES buffer with 14 mm glucose and 20 mm tetraethylamonium chloride. For KV channel recordings, patch electrodes were loaded with the following: MgATP, 140 mmol·liter−1 KCl, 1 mmol·liter−1 MgCl2, 10 mmol·liter−1 HEPES. Cells were perifused with Krebs-Ringer-HEPES buffer with 14 mm glucose and 100 μm tolbutamide.
Measurement of Cytoplasmic Calcium
Islets were incubated (20 min at 37 °C) in KRB supplemented with 2 μm Fura-2 acetoxymethyl ester (Molecular Probes, Eugene, OR). Fluorescence imaging was performed using a Nikon Eclipse TE2000-U microscope equipped with an epifluorescence illuminator (SUTTER, Inc.) a CCD camera (HQ2, Photometrics, Inc.) and Nikon Elements software (NIKON, Inc.). Cells were perifused at 37 °C at a flow of 2 ml/min with appropriate KRB-based solutions that contained glucose concentrations and compounds specified in the figures. Relative Ca2+ concentrations were quantified every 5 s by determining the ratio of emitted fluorescence intensities at excitation wavelengths of 340 and 380 nm (F340/F380). The relative Ca2+ is plotted as the average FURA-2 ratio (F340/F380) of each experimental group ± the S.E. The averaged area under the curve (AUC) measurement for FURA-2 ratios during the indicated time period (in minutes) was plotted as a bar graph. Data were analyzed using Excel and GraphPad Prism software and compared by Student's t test.
Insulin Secretion Measurements
Mouse islets were allowed to recover following isolation overnight. For insulin measurements, islets were incubated with DMEM with 5.6 mm glucose overnight followed by treatment with the indicated glucose concentrations and analyzed for insulin content using a radioimmunoassay or ELISA-based detection kit (ALPCO Diagnostics, Salem, NH), and data are presented as means ± S.E. For static insulin secretion, islets were incubated in 2 mm glucose containing KRB for 1 h followed by the indicated treatments, and insulin was assayed by ELISA-based detection.
Islet Immunofluorescence Staining
Pancreas cubes from RIP-rtTA+EAC3I-GFP and RIP-rtTA animals were fixed in 2% paraformaldehyde, paraffin embedded and cut into 5-μm sections on a microtome. Islet β-cells were stained using insulin (Millipore) and glucagon (Sigma) antibodies at 1:300 in combination with fluorescein isothiocyanate-conjugated secondary antibodies (Jackson ImmunoResearch Laboratories) together with DAPI nuclear stain (Invitrogen).
Islet NAD(P)H fluorescence Imaging
NAD(P)H fluorescence imaging was performed on a LSM780 confocal microscope (Zeiss, Inc.), using a tunable Chameleon Ti:Sapphire laser (Coherent, Inc.). Multiphoton excitation was obtained using a 710-nm wavelength. The emitted light was spectrally resolved using the λ-mode configuration of the LSM780, with a band-pass of 412–631 nm and a spectral resolution of 8.6 nm. The pixel dwell time was set at 50.4 μs, and the pixel size was 0.277 μm. We used a Fluor 40× oil objective, with a numerical aperture of 1.30 and dishes with a 0.16–0.19-mm-thick glass bottom (Mattek). The NAD(P)H emission and GFP emission were extracted by linear unmixing, using ZEN software (Zeiss, Inc.). The reference spectrum for NAD(P)H was obtained measuring the emission from a freshly made solution of β-NADH (Sigma, N8160) in 0.01 m Tris, pH 8.5. The reference spectrum for GFP was measured from a solution of purified GFP in PBS, pH 7.4. The unmixed images were processed using Fiji software (14) to measure the average NAD(P)H intensity across the entire islet and only in the GFP-expressing cells. During the imaging, the islets where maintained in KRBH (Krebs-Ringer bicarbonate/HEPES buffer), pH 7.40. The islets were kept at 37 °C, in a humidified atmosphere with 5% CO2, using the microscope stage incubator. The images were collected 8 min post glucose stimulus and 30 μm deep in the islet.
RESULTS
CaMKII Activity Modulates Glucose Tolerance by Enhancing Glucose-stimulated Insulin Secretion
To investigate the physiological role of β-cell CaMKII during GSIS, we created a mouse model which allows tetracycline-inducible inhibition of CaMKII. These animals are a cross between mice with a reverse tetracycline transactivator expressed in β-cells via an insulin promoter (RIP-rtTA) and mice with a CaMKII pseudosubstrate inhibitor peptide fused to GFP under the control of a tetracycline operator (tetO)-controlled promoter (EAC3I-GFP; Fig. 1A) (10). Doxycycline treatment of these animals, for 1.5 weeks, induces expression of EAC3I-GFP specifically in pancreatic β-cells, which can be visualized with GFP fluorescence (Fig. 1B). EAC3I-GFP expression occurs in a majority of β-cells, resembling previously published reports on RIP-rtTA mice showing doxycycline induced expression of tetO-controlled transgenes in 68 to 80% of β-cells (Fig. 1, B and C) (15, 16). β-Cell expression of EAC3I-GFP causes a significant reduction in Ca2+-induced serine 553 phosphorylation of islet synapsin 1 compared with controls (RIP-rtTA islets), consistent with the effective inhibition of CaMKII activity by EAC3I-GFP expression (Fig. 1D) (17). However, total pancreatic insulin levels were equivalent between mice with β-cell EAC3I-GFP expression and controls (RIP-rtTA pancreata; Fig. 1E). Furthermore, β-cell EAC3I-GFP expression does not cause any changes in islet morphology, as β-cell size and insulin immunofluorescence were equivalent to controls (Fig. 1, F and G). Therefore, conditional β-cell expression of EAC3I-GFP is a good model for assessing the role of CaMKII during physiological insulin secretion.
The effect of β-cell CaMKII inhibition on glucose homeostasis was assessed using intraperitoneal (intraperitoneal) glucose tolerance tests. EAC3I-GFP expression in β-cells resulted in a significant impairment of glucose tolerance compared with controls (RIP-rtTA mice; Fig. 2A, n = 13 per group). Insulin secretion assays comparing pancreatic islets isolated from control and EAC3I-GFP mice showed that inhibition of β-cell CaMKII suppresses insulin secretion (Fig. 2, B–D). Although there was no effect of CaMKII inhibition on basal insulin secretion with 5.6 mm glucose, which resembles blood glucose concentrations during fasting (Fig. 2, B and C), both first and second phase insulin secretion in response to 16.7 mm glucose were reduced in islets with β-cell CaMKII inhibition compared with controls (RIP-rtTA islets). Inhibiting β-cell CaMKII only significantly reduced GSIS at glucose concentrations >7 mm, which resemble blood glucose levels (10 mm) reached postprandially (Fig. 2D). In combination, these data show that CaMKII plays a key role in modulating glucose homeostasis and GSIS in vivo.
FIGURE 2.
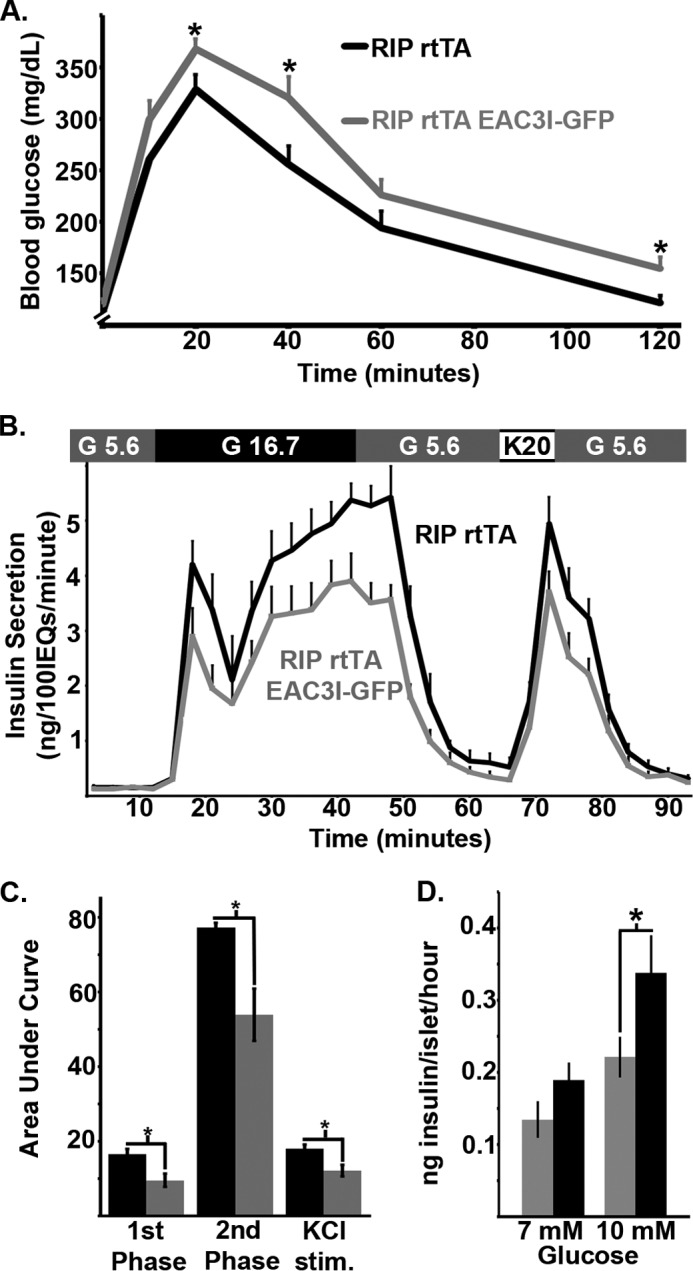
β-Cell CaMKII enhances glucose tolerance by amplifying glucose-stimulated insulin secretion in mice. A, glucose tolerance test on mice with β-cell CaMKII inhibition (gray line) and control (black line) animals. B, insulin secretion from islets isolated from β-cell EAC3I-GFP-expressing or (gray line) control (black line) animals, treated as indicated in above with 5.6 mm glucose, 16.7 mm glucose, and 20 mm KCl (n = 4 islet sets from independent animals per group). C, total islet insulin secretion (area under the curve) during 1st (15–24 min) and 2nd phase (24–42 min) insulin secretion as well as following KCl stimulation (66–72 min). Error bars represent ± S.E. D, doxycyline-treated RIP rtTA EAC3I (gray bars) and doxycyline-treated RIP rtTA (black bars) static islet insulin secretion under 7 and 10 mm glucose as indicated (n = 3 male islet sets for each animal type). Error bar, means ± S.E.; *, p < 0.05.
CaMKII Inhibition Does Not Affect Glucose Metabolism or β-Cell K+ Channel Activity
GSIS involves multiple steps that may be affected by CaMKII inhibition. First, we examined the potential effect of CaMKII inhibition on β-cell glucose metabolism by monitoring NAD(P)H fluorescence under varying glucose conditions (Fig. 3). No significant difference in glucose- stimulated changes in NAD(P)H fluorescence was observed between intact islet β-cells or whole islets that were obtained from mice with β-cell expression of EAC3I-GFP or GFP alone (Fig. 3, A and B). Thus, β-cell glucose metabolism is not affected by chronic CaMKII inhibition.
FIGURE 3.
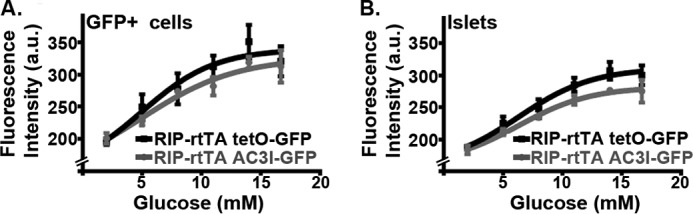
β-Cell CAMKII does not influence glucose metabolism. A, glucose-induced NAD(P)H fluorescence in GFP+ β-cells of an intact islet from RIP rtTA tetO-GFP (black line, 10 islets each from three independent preparations) or RIP rtTA EAC3I-GFP (gray line, 10 islets each from four independent preparations). a.u., arbitrary units. B, glucose-induced NAD(P)H fluorescence from all cells in an islet plane 30 μm deep from RIP rtTA tetO-GFP (black line, 10 islets each from three independent preparations) or RIP rtTA EAC3I-GFP (gray line, 10 islets each from four independent preparations). a.u., arbitrary units.
We next investigated potential effects of CaMKII inhibition on the two major β-cell K+ channels, the delayed rectifier voltage-gated K+ channel (KV) and ATP-sensitive K+ channel (KATP). The voltage step induction of KV currents was equivalent in control (RIP-rtTA, Fig. 4A) and CaMKII inhibited (EAC3I-GFP, Fig. 4B) β-cells, with similar overall current-voltage relationships (Fig. 4C). Similarly, reducing intracellular ATP concentrations induced equivalent KATP currents in control (RIP-rtTA) and EAC3I-GFP expressing β-cells in response to a voltage ramp (from −120 to 60 mV, Fig. 4D). Thus, CaMKII inhibition has no detectable affect on K+ channel activities in β-cells.
FIGURE 4.
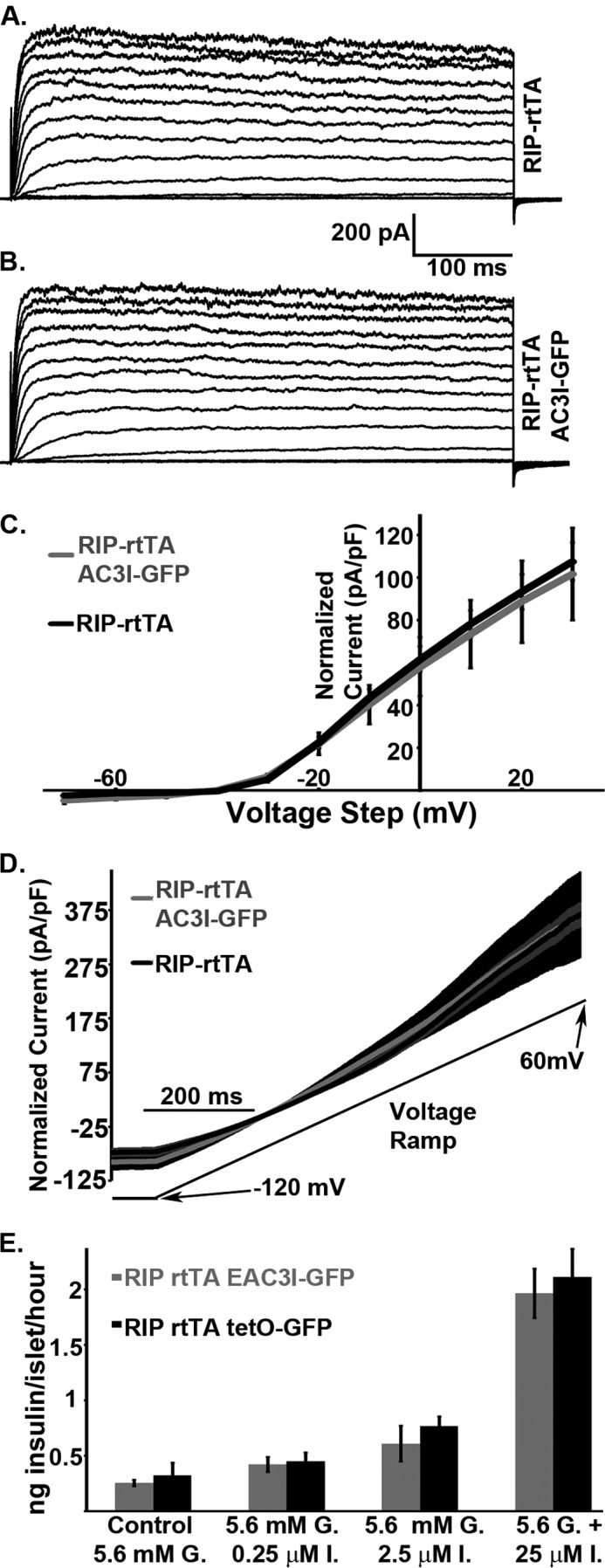
CaMKII does not significantly modulate β-cell KV or KATP channels. A, representative KV currents from a control (RIP-rtTA) β-cell in response to voltage steps in 10-mV increments from −80 to +80 mV. B, representative KV currents from an EAC3I-GFP expressing β-cell in response to voltage steps in 10-mV increments from −80 to +80 mV. C, current density versus voltage plots of RIP-rtTA (black line) and EAC3I-GFP β-cells (gray line) recorded in steps from −80 mV to the indicated voltage ± S.E. values (n = 5 for each condition). D, KATP currents recorded in response to a voltage ramp from −120 to 60 mV (black line below traces) from RIP-rtTA (black trace is the average of 11 recordings with gray S.E. bars) or EAC3I-GFP β-cells (gray trace is the average of 22 recordings with black S.E. bars). E, doxycyline-treated RIP rtTA EAC3I-GFP (gray bars) and doxycyline-treated RIP rtTA tetO-GFP (black bars) static islet insulin secretion with 5.6 mm glucose (G.) alone or in combination with 0.25, 2.5, or 25 μm ionomycin (I.; n = 3 male islet sets for each animal type). Error bars represent means ± S.E.
Consistent with these findings, insulin secretion induced by 20 mm potassium chloride in the presence of basal glucose, which bypasses β-cell glucose metabolism and K+ channels by directly depolarizing β-cells, was significantly lower from EAC3I-GFP expressing islets compared with controls (Fig. 2, B and C). Taken together, these data indicate that CaMKII modulates GSIS downstream of glucose metabolism and K+ channels to modulate GSIS.
CaMKII Modulates Glucose-stimulated Ca2+ Entry in β-Cells
Although CaMKII inhibition reduces Ca2+-dependent insulin secretion induced by high glucose or direct depolarization (see above), there was no difference in insulin secretion from EAC3I-GFP expressing and control islets in response to the Ca2+ ionophore ionomycin (Fig. 4E) (18), suggesting that CaMKII modulates β-cell Ca2+ homeostasis at some level. Consistent with this hypothesis, islet β-cells expressing EAC3I-GFP showed reduced basal Ca2+ levels compared with control islets (Fig. 5). Moreover, the changes in Ca2+ following glucose stimulation were also affected: CaMKII inhibition shortened the delay in initiation of Ca2+ influx but suppressed peak Ca2+ concentrations.
FIGURE 5.

β-Cell CaMKII increases cytoplasmic and endoplasmic reticulum Ca2+ levels. A, relative islet Ca2+ levels following glucose-stimulation (14 mm); islets (54 from RIP rtTA and 58 from RIP rtTA EAC3I) isolated from four independent animals. B, relative islet Ca2+ influx during 6 min post initiation of glucose-stimulated islet Ca2+ (IC) influx. C, relative islet Ca2+ levels from islet cell clusters (∼20 cells per cluster) following glucose stimulation(14 mm); AIP2 (pretreated with 20 μm cell permeable AIP2 for 20 min), vehicle (pretreated with vehicle for 20 min). D, relative islet Ca2+ influx (AUC) during 6 min post initiation of glucose-stimulated islet Ca2+ influx. E, relative islet Ca2+ levels from GFP positive islet cells with (gray trace, RIP rtTA EAC3I-GFP) or without EAC3I (black trace, RIP rtTA tetO-GFP) in 5.6 mm glucose KRB in response to 20 mm KCl stimulation (black bar). F, relative KCl induced Ca2+ increase (AUC) determined from β-cell Ca2+ influx during KCl treatment. G, relative islet Ca2+ levels during ER Ca2+ release with 2 μm thapsigargin (with no extracellular Ca2+). H, relative ER Ca2+ levels (AUC) determined from islet Ca2+ influx during thapsigargin treatment. I, relative islet Ca2+ levels during carbachol-induced (CCh) ER Ca2+ release (with no extracellular Ca2+). J, relative carbachol-induced ER Ca2+ release (AUC) determined from islet Ca2+ influx during carbachol treatment. Error bars represent means ± S.E.; ***, p < 0.001.
Acute CaMKII inhibition with a cell-permeable inhibitor peptide, AIP2, in dispersed wild-type C57/BL6 islet cell clusters also significantly affected Ca2+ homeostasis (Fig. 5, C and D). In contrast to the effects of chronic CaMKII inhibition (see above), acute CaMKII inhibition using AIP2 increased basal Ca2+ levels (2 mm glucose) and delayed the initiation of Ca2+ influx following glucose stimulation (Fig. 5C). However, acute CaMKII inhibition with AIP2 suppressed total glucose-induced increases in Ca2+, similar to the effects of chronic CaMKII inhibition. Notably, Ca2+ homeostasis in EAC3I-GFP β-cells treated with AIP2 was similar to EAC3I-GFP islet Ca2+ without AIP2 (Fig. 5C), indicating that these acute changes in basal β-cell Ca2+ are specifically caused by inhibition of CaMKII activity. The somewhat different effects of chronic in vivo and acute in vitro CaMKII inhibition on β-cell Ca2+ homeostasis presumably reflect long term adaptive responses to CaMKII inhibition that may be influenced by the in vivo environment. Nevertheless, the peak glucose-stimulated increase in Ca2+ concentration was significantly reduced by both acute and chronic CaMKII inhibition.
CaMKII Modulates ER Ca2+ Storage in β-Cells
CaMKII modulates ER Ca2+ uptake and release in cardiomyocytes (19–23). Therefore, we investigated whether the alterations in basal Ca2+ levels following β-cell CaMKII inhibition are due to changes in the intracellular Ca2+ stores. β-Cell ER Ca2+ levels were determined by inhibiting the sarco/endoplasmic reticulum ATPase (SERCA) with thapsigargin in the absence of extracellular Ca2+ and measuring the resulting elevation of cytosolic Ca2+ (Fig. 5, G and H). We found that ER Ca2+ stores were significantly reduced in islets with chronic β-cell inhibition of CaMKII compared with controls (RIP-rtTA islets; Fig. 5, G and H). To confirm that the reduction in thapsigargin-stimulated ER Ca2+ release was due to diminished ER Ca2+ levels, islets were also stimulated with the insulin secretagogue carbachol in the absence of extracellular Ca2+ to induce release of ER Ca2+ via IP3 receptors. ER Ca2+ release induced by carbachol treatment was found to be significantly reduced in islets with β-cell CaMKII inhibition (EAC3I-GFP) compared with control islets (RIP-rtTA) (Fig. 5, I and J). Therefore, CaMKII regulates β-cell cytoplasmic Ca2+ at least in part through modulation of ER Ca2+ handling.
CaMKII Activity Is Required for β-Cell l-type Ca2+ Channel Facilitation
GSIS is driven by Ca2+ influx via LTCCs. CaMKII facilitates voltage-induced Ca2+ entry via LTCCs in myocytes and neurons by multiple mechanisms involving different LTCC subunits (24–29). Therefore, we tested whether β-cell LTCCs also show voltage-induced facilitation that is dependent on CaMKII activity. Ca2+ currents were measured in response to a 100-ms voltage step to 30 mV before and after a strong positive voltage prepulse for 200 ms (160 mV, Fig. 6, A and B). Control β-cells (RIP-rtTA-cells) show significant Ca2+ channel facilitation following the positive voltage prepulse (current increase of 42.2 ± 4.8%, n = 19; Fig. 6, B and C), whereas β-cells with CaMKII inhibition show minimal voltage-induced Ca2+ channel facilitation (current increase of 11.1 ± 3.7%, n = 25; Fig. 6, B and C). This indicates that CaMKII activity facilitates β-cell Ca2+ channels following depolarization. To determine whether this change in LTCC facilitation influenced β-cell Ca2+ currents, LTCC currents were recorded with voltage steps from −70 to +70 mV in 10-mV increments. To prevent Ca2+ induced changes in LTCC currents before the first recording, the whole-cell configuration was obtained with a constant holding potential of −80 mV in low glucose (2 mm) and only the first set of voltage step recordings for each cell was utilized for subsequent analysis. Interestingly, under these conditions, β-cells with CaMKII inhibition show significantly reduced Ca2+ currents in response to voltage steps between 0 and +40 mV when compared with control β-cells (RIP-rtTA-cells; Fig. 6, C and D). However, there is no change in VDCC activation or inactivation with CaMKII inhibition (Fig. 6, E and F). Thus, CaMKII enhances Ca2+ entry into β-cells via LTCCs.
FIGURE 6.

CaMKII facilitates β-cell Ca2+ channels augmenting VDCC current amplitude. A, triple voltage step protocol used to elicit Ca2+ channel facilitation (left). β-Cell Ca2+ channel currents following the voltage clamp protocol are shown in A (right). The black Ca2+ current trace is during the first voltage step (P1), and the gray trace is during the third voltage step (P3). B, normalized change in β-cell Ca2+ current between the first voltage step and the third voltage step. C, β-cell Ca2+ channel currents in response to a voltage step to 10 mV. D, current voltage plot of β-cell Ca2+ channel currents in response to the indicated voltage steps. Error bars represent means ± S.E.; *, p < 0.05; **, p < 0.01. E, activation of β-cell VDCCs in response to the indicated voltage steps. F, fast (τfast) and slow (τslow) time constants of inactivation kinetics of β-cell VDCCs in response to the indicated voltage steps.
Glucose-stimulated activation of LTCCs is responsible for the upstroke of the action potential (AP), resulting in β-cell firing (30, 31). Therefore, we measured mouse β-cell membrane potentials to test whether CaMKII modulation of LTCCs affects β-cell AP firing. The AP firing frequency after a 10-min exposure to 14 mm glucose was significantly reduced in islet β-cells expressing EAC3I-GFP (1.39 ± 0.17 Hz) compared with control β-cells (RIP-rtTA-cells, 1.9 ± 0.16 Hz) (Fig. 7, A, B, and E). Furthermore, inhibition of β-cell KATP channels by a 2-min exposure to tolbutamide increases the AP firing frequency in control β-cells (RIP-rtTA, 2.35 ± 0.10 Hz), but this increase is significantly attenuated in EAC3I-GFP expressing β-cells (1.78 ± 0.21 Hz) (Fig. 7, C–E). Thus, CaMKII has an important role in glucose-induced and LTCC-dependent increase in AP-firing frequency, which further increases Ca2+ influx.
FIGURE 7.
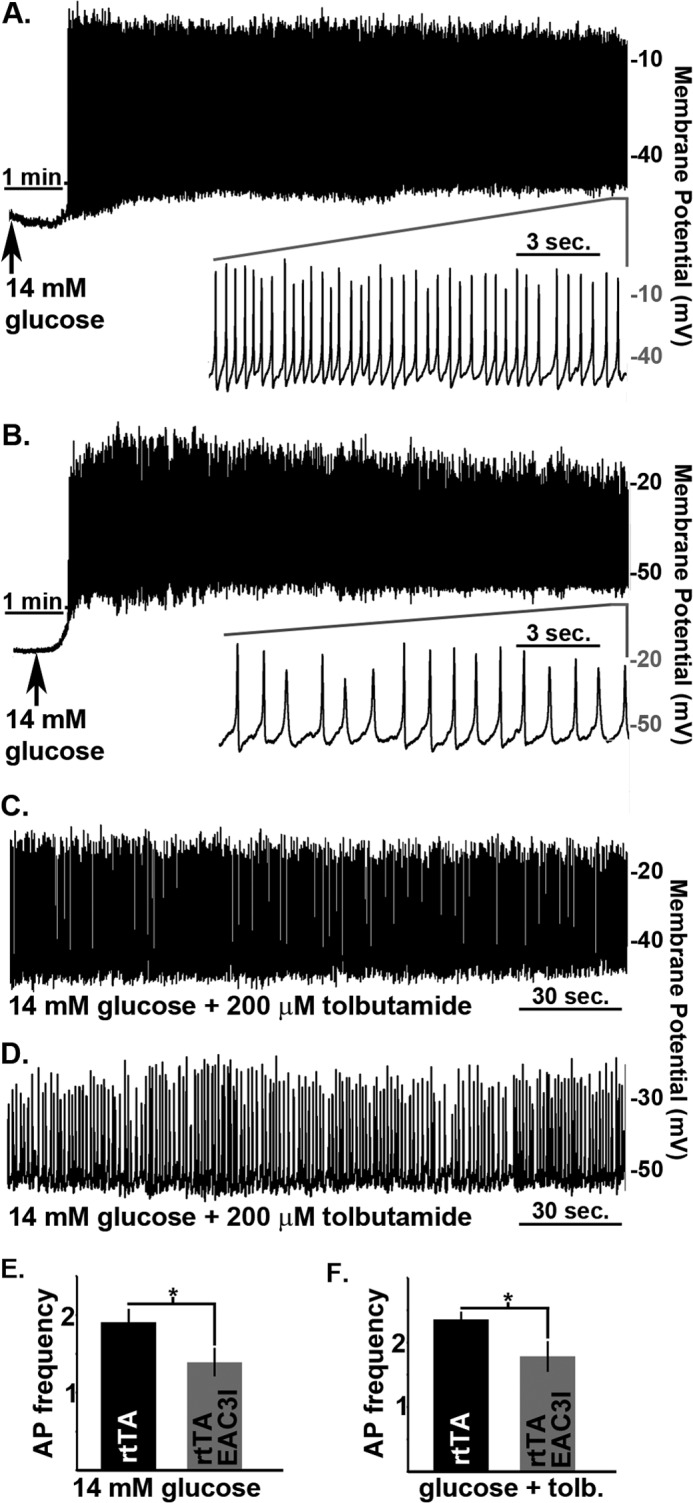
CaMKII activity increases β-cell action potential firing frequency. A, control β-cell electrical activity in response to 14 mm glucose. Inset shows the APs from the segment of activity indicated by the horizontal gray bar. B, electrical activity from a β-cell expressing EAC3I in response to 14 mm glucose. Inset shows the APs from the segment of activity indicated by the horizontal gray bar. C, control β-cell electrical activity in response to glucose (14 mm) and tolbutamide (200 μm). D, electrical activity from a β-cell expressing EAC3I in response to glucose (14 mm) and tolbutamide (200 μm). E, β-cell action potential firing frequency 10 min post glucose (14 mm) treatment, rtTA (n = 16) and EAC3I (n = 15). F, β-cell action potential firing frequency 10 min post glucose (14 mm) and 2 min post glucose (14 mm) with tolbutamide (200 μm) treatment, rtTA (n = 7) and EAC3I (n = 9). Error bars represent means ± S.E.; *, p < 0.05.
CaMKII Enhances β-Cell Function in the Presence of Insulin Resistance
To determine the role of CaMKII in augmenting Ca2+ influx and insulin secretion during periods of increased insulin demand, RIP-rtTA and EAC3I-GFP mice (n = 5 each) were placed on a HFD at 3 weeks of age to induce systemic insulin resistance. GTTs revealed that both groups of mice exhibited comparable glucose tolerance, as expected because the eAC3I-GFP transgene is not expressed under these conditions (Fig. 8, A and C). Both cohorts of animals were then placed on doxycycline while the HFD was continued. Reassessment of glucose tolerance after 2 weeks on doxycycline revealed a substantial impairment of glucose tolerance in animals with EAC3I-GFP expression in β-cells when compared with RIP-rtTA animals (Fig. 8, B and C, n = 5 per group). Because increased Ca2+ influx augments insulin secretion during conditions of insulin resistance (32), glucose-stimulated Ca2+ influx was also monitored in islets of HFD-treated animals. Glucose-stimulated Ca2+ influx was significantly diminished in EAC3I-GFP expressing islets compared with RIP-rtTA islets, which were both isolated from HFD-treated mice (Fig. 8, D and E). This demonstrates an important role for CaMKII in augmenting glucose-stimulated β-cell Ca2+ influx and insulin secretion under conditions of systemic insulin resistance.
FIGURE 8.

β-Cell CaMKII protects animals from glucose intolerance following exposure to a high fat diet by enhancing glucose stimulated Ca2+ entry. A, glucose tolerance test on mice (black line, RIP rtTA; gray line, RIP rtTA EAC3I) following 2 months on a high fat diet. B, glucose tolerance test on the mice shown in A 2 weeks post doxycycline (Dox.) treatment (gray line, β-cell CaMKII inhibition; black line, controls) following 2.5 months on a high fat diet. C, area under the curve data for the GTT of RIPrtTA (black bars) and EAC3I-GFP (gray bars) mice before and after doxycycline treatment. D, relative islet Ca2+ levels following glucose-stimulation (14 mm); islets (n > 70 islets from each group, RIP rtTA and RIP rtTA EAC3I) isolated from four independent animals. E, relative islet Ca2+ influx (AUC) during 6 min following the initiation of islet glucose-stimulated Ca2+ entry. Error bars represent means ± S.E.; *, p < 0.05.
DISCUSSION
It is well established that Ca2+ entry into pancreatic β-cells stimulates insulin secretion and that CaMKII activity influences Ca2+ homeostasis in many cell types (20, 33–35). Although CaMKII has been implicated in regulating islet insulin secretion, mechanisms underlying CaMKII action in β-cells are poorly understood (2). The results presented here demonstrate that CaMKII modulates Ca2+ handling and insulin secretion in a mouse model with conditional β-cell expression of a CaMKII inhibitory peptide. The results suggest that CaMKII enhances glucose tolerance by amplifying glucose-stimulated Ca2+ entry and insulin secretion.
Ca2+-dependent amplification and/or reduction of the channels required for cytosolic Ca2+ entry dynamically modulates Ca2+ signals in many cell types. Amplification can occur through many mechanisms, including Ca2+-dependent activation of ER Ca2+ release via ryanodine receptor (RYR) channels, as well as through Ca2+-dependent facilitation of plasma membrane LTCCs (33, 36, 37). Although some of these mechanisms involve direct binding of Ca2+-binding proteins (e.g. calmodulin) to the channels, others involve additional Ca2+-activated proteins such as CaMKII (33). In cardiac cells, CaMKII regulates multiple proteins involved in Ca2+ homeostasis, tightly modulating excitation-contraction and excitation-transcription coupling (33, 36, 37). Similarly, we find that CaMKII controls Ca2+ homeostasis in pancreatic β-cell, in this case, modulating excitation-secretion coupling. When activated during glucose-stimulation, β-cell CaMKII facilitates LTCCs and amplifies glucose-stimulated islet Ca2+ entry. Although LTCC facilitation by CaMKII can be induced by depolarization, it has also been shown to result from ER Ca2+ release in neurons as well as sarcoplasmic reticulum Ca2+ release in myocytes (26, 38, 39). Therefore, the CaV1.2 and/or CaV1.3 LTCCs in β-cells may undergo CaMKII-dependent facilitation via multiple mechanisms under different physiological conditions, which influence insulin secretion. The results suggest that β-cell CaMKII provides a Ca2+-sensitive feedback loop for enhancing GSIS secretion by amplifying Ca2+ influx.
CaMKII can also indirectly regulate LTCCs by regulating the plasma membrane potential via K+ channel modulation (40). Data presented here suggest that CaMKII does not modulate β-cell KV and KATP currents. We also find that CaMKII enhances AP firing frequency during KATP inhibition. Our data suggest that CaMKII regulates β-cell electrical activity and Ca2+ entry independently of any effects on KATP or KV channel activity. However, CaMKII can inhibit the inward rectifier subunit of the β-cell KATP channel complex (Kir6.2) (41). Moreover, CaMKII modulates H2O2 activation of sarcolemmal KATP channels and oxidative stress activates KV2.1 channels (42, 43). Thus, it is important to note that our data do not exclude potential effects of CaMKII on β-cell K+ channels under specific conditions not assessed in this study such as oxidative stress.
Endoplasmic reticulum Ca2+ stores also play an important role in regulating β-cell excitability and GSIS (44–47). CaMKII modulates ER Ca2+ levels by regulating both RYR and SERCA (21–23). RYR2 is phosphorylated by CaMKII in islets and mutations of RYR2 that mimic CaMKII phosphorylation increase β-cell ER Ca2+ release (23). Therefore, diminished ER Ca2+ leak through RYRs following chronic inhibition of β-cell CaMKII may cause the reductions in both thapsigargin-induced ER Ca2+ release and basal cytoplasmic Ca2+ levels. However, carbachol-induced ER Ca2+ release through IP3 receptors is also decreased in islets with β-cell CaMKII inhibition. Therefore, β-cell ER Ca2+ levels appear to be reduced by CaMKII inhibition, possibly due to SERCA inhibition. A brief reduction in SERCA activity with acute inhibition of CaMKII would account for the increase of basal Ca2+ (Fig. 5C) due to the inability of ER to take up cytoplasmic Ca2+. A sustained reduction of SERCA activity with chronic CaMKII inhibition would ultimately decrease β-cell ER Ca2+ stores (Fig. 5A). The exact mechanism(s) responsible for CaMKII modulation of β-cell ER Ca2+ handling are currently being investigated.
CaMKII-dependent modulation of Ca2+ influx likely causes the changes in GSIS reported here because ionomycin-stimulated insulin secretion is unaffected by CaMKII inhibition. However, other studies suggest roles for CaMKII in regulating components of insulin granule priming and or fusion. For example, one study found that CaMKII inhibition decreased insulin secretion even when islet intracellular calcium levels are clamped (48). These effects may involve the direct phosphorylation of synapsin 1 and microtubule-associated protein, MAP2, by CaMKII (49, 50). The studies presented here confirm that synapsin1 phosphorylation is indeed regulated by CaMKII. Synapsin1 controls the targeting of synaptic-like vesicles in primary pancreatic β-cells and rat insulinoma cells (17) and is localized to insulin granules in mouse insulinoma cells (51). This indicates the possibility that β-cell synapsin1 either directly influences insulin granule release via association with the insulin granule or indirectly influences insulin secretion through release of neurotransmitters from synaptic like vesicles. MAP2 is also phosphorylated by CaMKII and may influence insulin granule priming through insulin granule trafficking to the membrane on microtubules. This would be predicted to influence second phase insulin secretion, and interestingly, the studies presented here identify significant reductions in second phase insulin secretion when CaMKII is inhibited in islet β-cells. Thus, MAP2 may be important in regulating trafficking of insulin granules in response to CaMKII activation during GSIS. Future studies will determine the roles of synapsin 1 and MAP2 regulation of insulin secretion and islet function in response to glucose activation of CaMKII. In summary, although the changes in Ca2+ induced by β-cell CaMKII inhibition are likely to impair first phase insulin secretion, our data cannot exclude additional roles for CaMKII during insulin secretion such as granule trafficking and priming during second phase insulin secretion.
Although CaMKII physiologically modulates islet function, not much is known about the influence of CaMKII signaling during the pathogenesis of diabetes (2). Perturbations in CaMKII signaling are observed following palmitate treatment of insulinoma β-cells (52). Therefore, changes in CaMKII signaling may contribute to aberrant β-cell function following HFD feeding, during the progression of diabetes (52, 53). Indeed, we found that the inhibition of CaMKII in mice maintained on a HFD results in a rapid exacerbation of glucose intolerance. These data indicate that β-cell CaMKII activity is critical for the maintenance of glucose-stimulated islet Ca2+ influx and thus insulin secretion in the face of chronic hyperglycemia and the initial progression of insulin resistance induced by a high fat diet, thereby protecting mice from glucose intolerance. This indicates the exciting possibility that augmenting CaMKII activity may also increase human glucose-stimulated β-cell Ca2+ entry and enhance GSIS in a diabetic setting. Future studies will determine how CaMKII signaling influences human β-cell GSIS during the pathogenesis of diabetes.
In conclusion, this study suggests that CaMKII plays an important role as a β-cell Ca2+ sensor that modulates Ca2+ handling under both high energy/glucose and low energy/glucose conditions. CaMKII amplifies Ca2+ influx and insulin secretion during glucose stimulation and maintains cytoplasmic Ca2+ levels under fasting conditions. Thus, CaMKII is a key Ca2+ sensor that controls pancreatic β-cell Ca2+ homeostasis to dynamically modulate insulin secretion.
Acknowledgment
We thank Danny Winder for the EAC3I-GFP mouse model as well as helpful discussions and suggestions.
This work was supported by National Institutes of Health Grant DK096122 (to D. A. J.), DK081666 (to D. A. J.) as well as Pilot and Feasibility Grant P60 DK20593 through the Vanderbilt University Diabetes Research Training Center. The Vanderbilt Islet Procurement and Analysis Core was also supported by National Institutes of Health Grant DK20593.
- CaMKII
- Ca2+/calmodulin-dependent protein kinase II
- AIP2
- autocamtide-2 related inhibitory peptide II
- ER
- endoplasmic reticulum
- GSIS
- glucose stimulated insulin secretion
- HFD
- high fat diet
- KRB
- Krebs-Ringer buffer
- LTCC
- L-type Ca2+ channel
- RYR
- ryanodine receptor
- SERCA
- sarco/endoplasmic reticulum ATPase
- tetO
- tetracycline operator
- VDCC
- voltage dependent Ca2+ channel
- GTT
- glucose tolerance test
- AUC
- area under the curve
- AP
- action potential.
REFERENCES
- 1. Ammälä C., Eliasson L., Bokvist K., Berggren P. O., Honkanen R. E., Sjöholm A., Rorsman P. (1994) Activation of protein kinases and inhibition of protein phosphatases play a central role in the regulation of exocytosis in mouse pancreatic beta cells. Proc. Natl. Acad. Sci. U.S.A. 91, 4343–4347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Easom R. A. (1999) CaM kinase II: a protein kinase with extraordinary talents germane to insulin exocytosis. Diabetes 48, 675–684 [DOI] [PubMed] [Google Scholar]
- 3. Ashcroft S. J. (1994) Protein phosphorylation and beta-cell function. Diabetologia 37, S21–29 [DOI] [PubMed] [Google Scholar]
- 4. Norling L. L., Colca J. R., Kelly P. T., McDaniel M. L., Landt M. (1994) Activation of calcium and calmodulin dependent protein kinase II during stimulation of insulin secretion. Cell Calcium 16, 137–150 [DOI] [PubMed] [Google Scholar]
- 5. Wenham R. M., Landt M., Easom R. A. (1994) Glucose activates the multifunctional Ca2+/calmodulin-dependent protein kinase II in isolated rat pancreatic islets. J. Biol. Chem. 269, 4947–4952 [PubMed] [Google Scholar]
- 6. Ammälä C., Eliasson L., Bokvist K., Larsson O., Ashcroft F. M., Rorsman P. (1993) Exocytosis elicited by action potentials and voltage-clamp calcium currents in individual mouse pancreatic B-cells. J. Physiol. 472, 665–688 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Babb E. L., Tarpley J., Landt M., Easom R. A. (1996) Muscarinic activation of Ca2+/calmodulin-dependent protein kinase II in pancreatic islets. Temporal dissociation of kinase activation and insulin secretion. Biochem. J. 317, 167–172 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Gromada J., Høy M., Renström E., Bokvist K., Eliasson L., Göpel S., Rorsman P. (1999) CaM kinase II-dependent mobilization of secretory granules underlies acetylcholine-induced stimulation of exocytosis in mouse pancreatic B-cells. J. Physiol. 518, 745–759 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Milo-Landesman D., Surana M., Berkovich I., Compagni A., Christofori G., Fleischer N., Efrat S. (2001) Correction of hyperglycemia in diabetic mice transplanted with reversibly immortalized pancreatic beta cells controlled by the tet-on regulatory system. Cell Transplant. 10, 645–650 [PubMed] [Google Scholar]
- 10. Klug J. R., Mathur B. N., Kash T. L., Wang H. D., Matthews R. T., Robison A. J., Anderson M. E., Deutch A. Y., Lovinger D. M., Colbran R. J., Winder D. G. (2012) Genetic inhibition of CaMKII in dorsal striatal medium spiny neurons reduces functional excitatory synapses and enhances intrinsic excitability. PLoS One 7, e45323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Jacobson D. A., Kuznetsov A., Lopez J. P., Kash S., Ammälä C. E., Philipson L. H. (2007) Kv2.1 ablation alters glucose-induced islet electrical activity, enhancing insulin secretion. Cell Metab. 6, 229–235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Philipson L. H., Rosenberg M. P., Kuznetsov A., Lancaster M. E., Worley J. F., 3rd, Roe M. W., Dukes I. D. (1994) Delayed rectifier K+ channel overexpression in transgenic islets and beta-cells associated with impaired glucose responsiveness. J. Biol. Chem. 269, 27787–27790 [PubMed] [Google Scholar]
- 13. Rae J., Cooper K., Gates P., Watsky M. (1991) Low access resistance perforated patch recordings using amphotericin B. J. Neurosci. Methods 37, 15–26 [DOI] [PubMed] [Google Scholar]
- 14. Schindelin J., Arganda-Carreras I., Frise E., Kaynig V., Longair M., Pietzsch T., Preibisch S., Rueden C., Saalfeld S., Schmid B., Tinevez J. Y., White D. J., Hartenstein V., Eliceiri K., Tomancak P., Cardona A. (2012) Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Nir T., Melton D. A., Dor Y. (2007) Recovery from diabetes in mice by beta cell regeneration. J. Clin. Invest. 117, 2553–2561 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Pullen T. J., Sylow L., Sun G., Halestrap A. P., Richter E. A., Rutter G. A. (2012) Overexpression of monocarboxylate transporter-1 (SLC16A1) in mouse pancreatic beta-cells leads to relative hyperinsulinism during exercise. Diabetes 61, 1719–1725 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Krueger K. A., Ings E. I., Brun A. M., Landt M., Easom R. A. (1999) Site-specific phosphorylation of synapsin I by Ca2+/calmodulin-dependent protein kinase II in pancreatic betaTC3 cells: synapsin I is not associated with insulin secretory granules. Diabetes 48, 499–506 [DOI] [PubMed] [Google Scholar]
- 18. Bernal-Mizrachi E., Fatrai S., Johnson J. D., Ohsugi M., Otani K., Han Z., Polonsky K. S., Permutt M. A. (2004) Defective insulin secretion and increased susceptibility to experimental diabetes are induced by reduced Akt activity in pancreatic islet beta cells. J. Clin. Invest. 114, 928–936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Anthony D. F., Beattie J., Paul A., Currie S. (2007) Interaction of calcium/calmodulin-dependent protein kinase IIδC with sorcin indirectly modulates ryanodine receptor function in cardiac myocytes. J. Mol. Cell Cardiol. 43, 492–503 [DOI] [PubMed] [Google Scholar]
- 20. Wehrens X. H., Lehnart S. E., Reiken S. R., Marks A. R. (2004) Ca2+/calmodulin-dependent protein kinase II phosphorylation regulates the cardiac ryanodine receptor. Circ. Res. 94, e61–70 [DOI] [PubMed] [Google Scholar]
- 21. Currie S., Loughrey C. M., Craig M. A., Smith G. L. (2004) Calcium/calmodulin-dependent protein kinase IIδ associates with the ryanodine receptor complex and regulates channel function in rabbit heart. Biochem. J. 377, 357–366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Xu A., Hawkins C., Narayanan N. (1993) Phosphorylation and activation of the Ca2+-pumping ATPase of cardiac sarcoplasmic reticulum by Ca2+/calmodulin-dependent protein kinase. J. Biol. Chem. 268, 8394–8397 [PubMed] [Google Scholar]
- 23. Dixit S. S., Wang T., Manzano E. J., Yoo S., Lee J., Chiang D. Y., Ryan N., Respress J. L., Yechoor V. K., Wehrens X. H. (2013) Effects of CaMKII-mediated phosphorylation of ryanodine receptor type 2 on islet calcium handling, insulin secretion, and glucose tolerance. PLoS One 8, e58655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Grueter C. E., Abiria S. A., Dzhura I., Wu Y., Ham A. J., Mohler P. J., Anderson M. E., Colbran R. J. (2006) L-type Ca2+ channel facilitation mediated by phosphorylation of the beta subunit by CaMKII. Molecular Cell 23, 641–650 [DOI] [PubMed] [Google Scholar]
- 25. Hudmon A., Schulman H., Kim J., Maltez J. M., Tsien R. W., Pitt G. S. (2005) CaMKII tethers to L-type Ca2+ channels, establishing a local and dedicated integrator of Ca2+ signals for facilitation. J. Cell Biol. 171, 537–547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Gao L., Blair L. A., Salinas G. D., Needleman L. A., Marshall J. (2006) Insulin-like growth factor-1 modulation of CaV1.3 calcium channels depends on Ca2+ release from IP3-sensitive stores and calcium/calmodulin kinase II phosphorylation of the α1 subunit EF hand. J. Neurosci. 26, 6259–6268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Jenkins M. A., Christel C. J., Jiao Y., Abiria S., Kim K. Y., Usachev Y. M., Obermair G. J., Colbran R. J., Lee A. (2010) Ca2+-dependent facilitation of Cav1.3 Ca2+ channels by densin and Ca2+/calmodulin-dependent protein kinase II. J. Neurosci. 30, 5125–5135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Abiria S. A., Colbran R. J. (2010) CaMKII associates with CaV1.2 L-type calcium channels via selected beta subunits to enhance regulatory phosphorylation. J. of Neurochem. 112, 150–161 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Lee T. S., Karl R., Moosmang S., Lenhardt P., Klugbauer N., Hofmann F., Kleppisch T., Welling A. (2006) Calmodulin kinase II is involved in voltage-dependent facilitation of the L-type Cav1.2 calcium channel: Identification of the phosphorylation sites. J. Biol. Chem. 281, 25560–25567 [DOI] [PubMed] [Google Scholar]
- 30. Braun M., Ramracheya R., Bengtsson M., Zhang Q., Karanauskaite J., Partridge C., Johnson P. R., Rorsman P. (2008) Voltage-gated ion channels in human pancreatic beta-cells: electrophysiological characterization and role in insulin secretion. Diabetes 57, 1618–1628 [DOI] [PubMed] [Google Scholar]
- 31. Matthews E. K., Sakamoto Y. (1975) Electrical characteristics of pancreatic islet cells. J. Physiol. 246, 421–437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Gonzalez A., Merino B., Marroquí L., Ñeco P., Alonso-Magdalena P., Caballero-Garrido E., Vieira E., Soriano S., Gomis R., Nadal A., Quesada I. (2013) Insulin hypersecretion in islets from diet-induced hyperinsulinemic obese female mice is associated with several functional adaptations in individual beta-cells. Endocrinology 154, 3515–3524 [DOI] [PubMed] [Google Scholar]
- 33. Dzhura I., Wu Y., Colbran R. J., Balser J. R., Anderson M. E. (2000) Calmodulin kinase determines calcium-dependent facilitation of L-type calcium channels. Nat. Cell Biol. 2, 173–177 [DOI] [PubMed] [Google Scholar]
- 34. Malinow R., Schulman H., Tsien R. W. (1989) Inhibition of postsynaptic PKC or CaMKII blocks induction but not expression of LTP. Science 245, 862–866 [DOI] [PubMed] [Google Scholar]
- 35. Kranias E. G., Gupta R. C., Jakab G., Kim H. W., Steenaart N. A., Rapundalo S. T. (1988) The role of protein kinases and protein phosphatases in the regulation of cardiac sarcoplasmic reticulum function. Mol. Cell Biochem. 82, 37–44 [DOI] [PubMed] [Google Scholar]
- 36. Zhang R., Khoo M. S., Wu Y., Yang Y., Grueter C. E., Ni G., Price E. E., Jr., Thiel W., Guatimosim S., Song L. S., Madu E. C., Shah A. N., Vishnivetskaya T. A., Atkinson J. B., Gurevich V. V., Salama G., Lederer W. J., Colbran R. J., Anderson M. E. (2005) Calmodulin kinase II inhibition protects against structural heart disease. Nat. Med. 11, 409–417 [DOI] [PubMed] [Google Scholar]
- 37. Wu Y., Colbran R. J., Anderson M. E. (2001) Calmodulin kinase is a molecular switch for cardiac excitation-contraction coupling. Proc. Natl. Acad. Sci. U.S.A. 98, 2877–2881 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Xiao R. P., Cheng H., Lederer W. J., Suzuki T., Lakatta E. G. (1994) Dual regulation of Ca2+/calmodulin-dependent kinase II activity by membrane voltage and by calcium influx. Proc. Natl. Acad. Sci. U.S.A. 91, 9659–9663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Wu Y., Kimbrough J. T., Colbran R. J., Anderson M. E. (2004) Calmodulin kinase is functionally targeted to the action potential plateau for regulation of L-type Ca2+ current in rabbit cardiomyocytes. J. Physiol. 554, 145–155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Griffith L. C., Wang J., Zhong Y., Wu C. F., Greenspan R. J. (1994) Calcium/calmodulin-dependent protein kinase II and potassium channel subunit eag similarly affect plasticity in Drosophila. Proc. Natl. Acad. Sci. U.S.A. 91, 10044–10048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Kline C. F., Wright P. J., Koval O. M., Zmuda E. J., Johnson B. L., Anderson M. E., Hai T., Hund T. J., Mohler P. J. (2013) betaIV-Spectrin and CaMKII facilitate Kir6.2 regulation in pancreatic beta cells. Proc. Natl. Acad. Sci. U.S.A. 110, 17576–17581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. McCord M. C., Aizenman E. (2013) Convergent Ca2+ and Zn2+ signaling regulates apoptotic Kv2.1 K+ currents. Proc. Natl. Acad. Sci. U.S.A. 110, 13988–13993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Zhang D. M., Chai Y., Erickson J. R., Brown J. H., Bers D. M., Lin Y. F. (2014) Modulation of sarcolemmal ATP-sensitive potassium channels by nitric oxide via sGC/PKG/ROS/ERK1/2/CaMKII signaling in ventricular cardiomyocytes. J. Physiol. 592, 971–990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Worley J. F., 3rd, McIntyre M. S., Spencer B., Mertz R. J., Roe M. W., Dukes I. D. (1994) Endoplasmic reticulum calcium store regulates membrane potential in mouse islet beta-cells. J. Biol. Chem. 269, 14359–14362 [PubMed] [Google Scholar]
- 45. Roe M. W., Mertz R. J., Lancaster M. E., Worley J. F., 3rd, Dukes I. D. (1994) Thapsigargin inhibits the glucose-induced decrease of intracellular Ca2+ in mouse islets of Langerhans. Am. J. Physiol. 266, E852–862 [DOI] [PubMed] [Google Scholar]
- 46. Wollheim C. B., Janjic D., Siegel E. G., Kikuchi M., Sharp G. W. (1981) Importance of cellular calcium stores in glucose-stimulated insulin release. Ups. J. Med. Sci. 86, 149–164 [DOI] [PubMed] [Google Scholar]
- 47. Prentki M., Wollheim C. B. (1984) Cytosolic free Ca2+ in insulin secreting cells and its regulation by isolated organelles. Experientia 40, 1052–1060 [DOI] [PubMed] [Google Scholar]
- 48. Bhatt H. S., Conner B. P., Prasanna G., Yorio T., Easom R. A. (2000) Dependence of insulin secretion from permeabilized pancreatic beta-cells on the activation of Ca2+/calmodulin-dependent protein kinase II: a re-evaluation of inhibitor studies. Biochem. Pharmacol. 60, 1655–1663 [DOI] [PubMed] [Google Scholar]
- 49. Matsumoto K., Fukunaga K., Miyazaki J., Shichiri M., Miyamoto E. (1995) Ca2+/calmodulin-dependent protein kinase II and synapsin I-like protein in mouse insulinoma MIN6 cells. Endocrinology 136, 3784–3793 [DOI] [PubMed] [Google Scholar]
- 50. Krueger K. A., Bhatt H., Landt M., Easom R. A. (1997) Calcium-stimulated phosphorylation of MAP-2 in pancreatic βTC3-cells is mediated by Ca2+/calmodulin-dependent kinase II. J. Biol. Chem. 272, 27464–27469 [DOI] [PubMed] [Google Scholar]
- 51. Yamamoto H., Matsumoto K., Araki E., Miyamoto E. (2003) New aspects of neurotransmitter release and exocytosis: involvement of Ca2+/calmodulin-dependent phosphorylation of synapsin I in insulin exocytosis. J. Pharmacol. Sci. 93, 30–34 [DOI] [PubMed] [Google Scholar]
- 52. Watson M. L., Macrae K., Marley A. E., Hundal H. S. (2011) Chronic effects of palmitate overload on nutrient-induced insulin secretion and autocrine signaling in pancreatic MIN6 beta cells. PLoS One 6, e25975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Prentki M., Nolan C. J. (2006) Islet beta cell failure in type 2 diabetes. J. Clin. Invest. 116, 1802–1812 [DOI] [PMC free article] [PubMed] [Google Scholar]