

Abstract
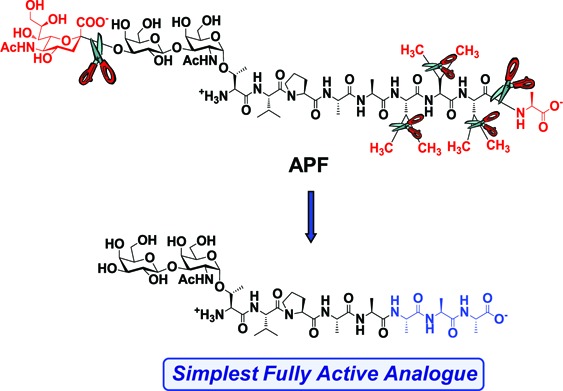
Antiproliferative factor (APF), a sialylated glycopeptide secreted by explanted bladder epithelial cells from interstitial cystitis/painful bladder syndrome (IC/PBS) patients, and its unsialylated analogue (as-APF) significantly decrease proliferation of bladder epithelial cells and/or certain carcinoma cell lines in vitro. We recently reported a structure−activity relationship profile for the peptide portion of as-APF and revealed that truncation of the C-terminal alanine did not significantly affect antiproliferative activity. To better understand the structural basis for the maintenance of activity of this truncated eight amino acid as-APF (as-APF8), we synthesized several amino acid-substituted derivatives and studied their ability to inhibit bladder epithelial cell proliferation in vitro as well as their solution conformations by CD and NMR spectroscopy. While single amino acid changes to as-APF8 often strongly reduced activity, full potency was retained when the trivaline tail was replaced with three alanines. The Ala6−8 derivative 9 is the simplest, fully potent APF analogue synthesized to date.
Keywords: Interstitial cystitis/painful bladder syndrome, antiproliferative, glycopeptide, hydrophobicity, peptide conformation
Interstitial cystitis/painful bladder syndrome (IC/PBS) is a devastating disease of the bladder that is characterized by thinning or even focal obliteration of the bladder epithelium.1−3 Approximately 1 million Americans suffer from IC/PBS, and it occurs 8−9 times more frequently in women than in men.1,3 Although a bladder condition resembling IC/PBS was first described about 200 years ago,2 the cause of this disease has remained unknown. However, urine from approximately 95% of IC/PBS patients who fulfill modified NIDDK criteria for IC4 contains an antiproliferative factor (APF) that potently decreases the proliferation of human bladder epithelial cells in vitro5 and elicits changes in normal bladder epithelium also seen in bladder biopsy specimens and/or explanted bladder epithelial cells from IC/PBS patients (including specific changes in growth factor expression,6 increased paracellular permeability, and aberrations in tight junction protein expression and tight junction formation).7−16 APF has been identified as a short trisaccharide-containing glycopeptide—viz., Neu5Acα2-3Galβ1-3GalNAcα-O-TVPAAVVVA17 (1, Figure 1) with an intriguing structure−activity relationship (SAR).18 Although the peptide sequence is identical to a segment of the sixth transmembrane domain of the frizzled-8 protein,17,19 a Wnt ligand receptor, the details of its biosynthesis and mechanism of action in IC/PBS patients remain unknown. An important advance into understanding the mechanism of APF function was identification of cytoskeletal-associated protein 4 (CKAP4/ERGIC-63/CLIMP-63) as a cellular receptor for APF in bladder epithelium.20
Figure 1.

Structures of APF (1), as-APF (2), and as-APF8 (3).
A preliminary analysis of APF showed that the trisaccharide is not an absolute requirement as the desialylated native APF as well as the asialo synthetic derivative (as-APF, 2, Figure 1) maintained full potency; however, two proximal sugar residues are required for activity17,18 (the unglycosylated peptide as well as the α-GalNAc monosaccharide derivatives were both inactive). A recent SAR study18 of the peptide portion of APF revealed certain amino acid requirements for maintenance of as-APF activity. Greater understanding of the SAR for active APF derivatives potentially will allow for more rational design of APF mimics/inhibitors.
As the 8-residue derivative (as-APF8, 3, Figure 1) was equipotent to as-APF, we synthesized several analogues of as-APF8 and qualitatively compared their activity and solution structures as determined from circular dichroism (CD) and NMR spectroscopies. We concentrated primarily on C-terminal amino acid substitutions, because our previous data indicated that the C-terminal A-XXX motif was important for activity, and modifications to this segment tended to have negative effects on potency (with some exceptions). During initial17 structural characterization and subsequent SAR18 work on APF, it was discovered that compounds 2 and 3 (Figure 1) were essentially equipotent in antiproliferation assays with normal bladder epithelial cells; further truncation to a 7-mer yielded a completely inactive compound. On the basis of these results, the potency of each as-APF8 analogue described in this report was compared simultaneously to that of 2.
General procedures for glycopeptide synthesis, their structures, and activity data are outlined in the Supporting Information (Table S1). Generally, Fmoc-protected amino acids were coupled using HATU and HOAt in the presence of DIPEA in dry NMP. Incorporation of the valine residue in position 2 was affected by two rounds of coupling (double coupling) in the synthesis of all glycopeptides due to the lower reactivity of the secondary prolyl amino group. Coupling of Fmoc-Thr(Ac4Galβ1-3Ac2GalNAcα-O)-OH (1.1 equiv) was performed without any added base to prevent epimerization. All compounds were determined to be >95% pure by analytical high-performance liquid chromatography (HPLC) and electrospray ionization liquid chromatography/mass spectrometry (LC/MS) analysis (Supporting Information).
Inhibition of [3H]thymidine incorporation was used to evaluate antiproliferative activity.5 The activity data for the synthetic APF derivative glycopeptides described herein along with previously prepared compounds 1−4 are listed in Table S1 in the Supporting Information. We were aware that a disaccharide was a requirement for antiproliferative activity of the as-APF glycopeptide;17,18 unglycosylated peptide 5 was also devoid of activity. A representative set of assay curves for compounds 2, 7, 9, and 14 are shown in Figure 2.
Figure 2.

Antiproliferative activity of compounds 2, 7, 9, and 14 in normal bladder epithelial cell explant cultures. Explanted normal bladder epithelial cells were treated with varying concentrations of as-APF 2 (⧫), trileucine analogue 7 (●), trialanine analogue 9 (◻), or Val-Ala-Val analogue 14 (△) for 48 h prior to the determination of 3[H]-thymidine incorporation. The assay was performed in triplicate twice; data are expressed as percent inhibition of thymidine incorporation as compared to control cells incubated with medium alone.
The modifications made to the three C-terminal amino acids were either “single” (compounds 6, 13, and 14) or “global” involving all three C-terminal amino acids (compounds 7−12). It was previously determined that ionization at either the C- or the N-amino acid terminus of as-APF was important for full potency as “end-capping” by N-acetylation or carboxamidation caused a 2.5 order of magnitude drop in potency.18 This behavior was even more dramatic for as-APF8 derivatives as the C-terminal carboxamide analogue 6 was completely inactive (acetylation of the N-terminal amino group also resulted in inactivation, data not shown).
Compounds 7−12 were prepared to evaluate the importance of the hydrophobicity and β-branching in the A-XXX motif of APF. Previous data showed that this unit might allow the C-terminal tail of active APF analogues to populate some form of defined secondary structure.18,21 Because as-APF8 maintained full antiproliferative potency, we systematically tested as-APF8 analogues incorporating amino acids with varying hydrophobicities, by globally changing the trivaline segment to one with amino acids with one added methylene unit (trileucine replacement, designated Leu6−8, compound 7) to one with amino acids having no carbon-containing side chains (Gly6−8, compound 10). The octanol/water partition coefficient (logP) values of each new compound were calculated22 and are listed in Table S1 in the Supporting Information to compare with as-APF8. The slightly more hydrophobic Leu6−8 analogue 7 maintained a high degree of potency (10% of full activity), while removal of one methyl group from valines 6−8 by substitution with unnatural aminobutyric acid (Abu, 8) resulted in inactivation. The even less hydrophobic Ala6−8 derivative 9 yielded a compound that retained full potency. This stands in contrast to replacement of the valines 6−8 in the 9-mer as-APF with alanines, as this congener showed a reduction in potency of 2 orders of magnitude as compared to as-APF.18 This result made it difficult to rationalize the lack of the activity of Galβ1-3GalNAcα-O-TVPAA-Abu-Abu-Abu analogue 8, as its ethyl side chain is “between” alanine and valine in both hydrophobicity and steric bulk. Lyu et al.23 found that alanine and Abu possess equal (but greater than valine) ability to stabilize helical content. Assuming that stabilization of helical content is also important in the C terminus of APF, it would seem reasonable to predict that 8 should show some potency in our assay. However, other factors may complicate this hypothesis (see below). As expected, compound 10 (Gly6−8 substitution) resulted in complete loss of activity. Potency was partially recovered (0.1%) when Val6−8 was substituted with β-alanine (compound 11), a replacement used in many instances to induce helical content in small peptides.24 Adding back an α-methyl group by way of β3-homoalanine replacements (compound 12) once more inactivated the glycopeptides.
Two single replacements used previously in as-APF were also used in as-APF8, viz. replacement of Val7 with d-valine or alanine. As it had for as-APF, d-valine substitution inactivated as-APF8, illustrating that non-natural amino acid derivatives (compounds 8, 12, and 13) are not tolerated. The single Ala7-substituted analogue 14 was also prepared to offer a compound with a logP that was midway between the two active derivatives 3 and 9. However, this also resulted in complete inactivation of as-APF8.
CD spectroscopy25 was initially used to study each analogue with comparisons to the spectra for either 2 and/or 3. The CD spectrum of 3 revealed no organized structure of the glycopeptide in water (similar to previous data for 2(18)), as was the case for all other new analogues. However, 3 appeared to adopt some secondary structure in a 1:1 H2O:TFE mixture, (Figure 3) the milieu that was used for all subsequent CD spectra. The spectrum, with two minima centered at ca.199 and 220 nm, resembled that previously obtained for 2 in neat TFE (unpublished results). In 1:1 H2O:TFE, nearly identical CD spectra for the three derivatives 2, 3, and 4 suggested that truncation does not significantly affect conformation in this milieu. Surprisingly, the structure of unglycosylated 8-mer 5 appeared to be significantly more organized than the structure of 3 (Figure 3). Other notable CD results (see Figure 4) showed that (1) compound 7 (10% potency) was significantly less organized than structure 3, (2) compounds 8 (inactive) and 9 (full activity) had almost superimposable spectra, and (3) substitution with non-natural amino acids resulted in no organized structures (compounds 11, 12, and 13; Figure S1 in the Supporting Information).
Figure 3.
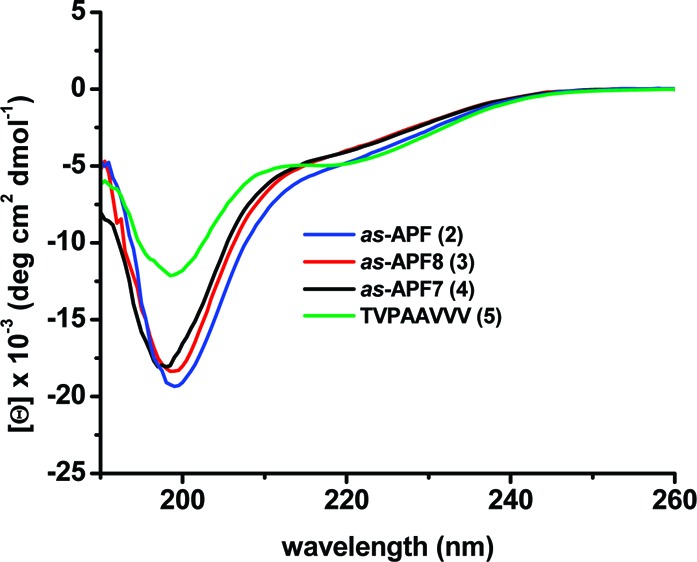
CD spectra of a 50 μM solution of 2 (blue line), 3 (red line), 4 (black line), and 5 (green line) in H2O:TFE (1:1) recorded at 25 °C.
Figure 4.
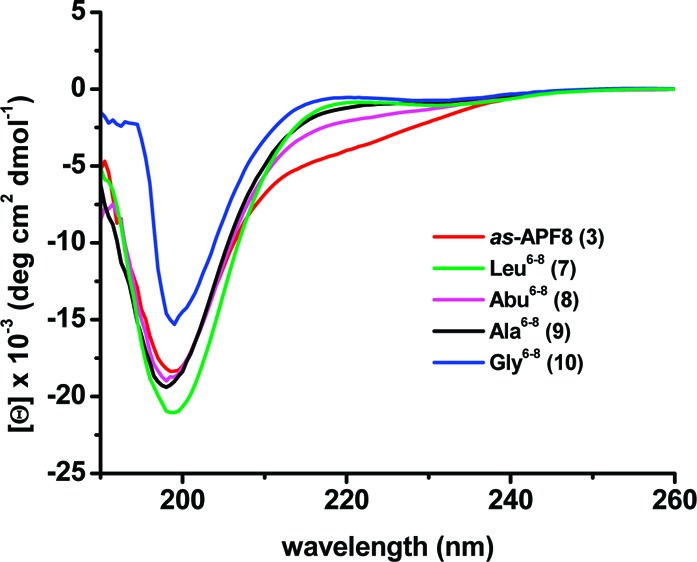
CD spectra of 50 μM solutions of 3 (red line), 7 (green line), 8 (pink line), 9 (black line), and 10 (blue line) in H2O:TFE (1:1) mixture recorded at 25 °C.
The glycopeptides synthesized in this study were also examined by NMR spectroscopy in aqueous solution. All 8-mer peptides from Table S1 in the Supporting Information were fully assigned by exhaustive NMR experiments (a representative comparison of chemical shifts and temperature coefficients data for compounds 3 and 5 can be found in Table S2 and Figure S3 in the Supporting Information). Spectra of APF derivatives in 9:1 H2O:D2O, in general, gave well-dispersed and sharp signals, suggesting little if any slow chemical exchange on the NMR time scale. Buffered solutions at pH 4.6 gave the highest quality spectra as compared with several pH values tested. A stacked plot of the amide region of several 8-residue analogues synthesized here is shown in Figure 5. Most NH protons were fully resolved in several derivatives, suggesting preferred conformations in solution. Some more notable observations were (1) consistent chemical shift values across all derivatives for amide protons for GalNAc, Val2, Ala4, and Ala5, indicating, little “cross-talk” between the N and the C termini; (2) very distinct chemical shift differences for the two C-terminal carboxamide protons for “inactive” derivative 6; and (3) a higher population of the cis amide bond for Pro3 when the sugar is removed. In addition, amide temperature coefficient data showed that the Δδ/ΔT value for Val2 decreased from −7.3 to −6.0 ppb/K, indicating a possible shielding or H-bonding effect upon glycosylation. Our data thus far (refs (17) and (18) and unpublished results) have shown that a distinct sugar disaccharide profile is important for the maintenance of activity. The sugars have important and also remote conformational effects, which in turn can be closely related to changes in antiproliferative activity.
Figure 5.

One-dimensional NMR spectra of the amide region of 8-mer APF derivatives in 9:1 H2O:D2O in acetate buffer, pH 4.6. Amino acid assignments are labeled over the peaks. Compound labels for each spectrum (right of spectrum) identify the substitution employed and the compound number from Table S1 in the Supporting Information.
With the exception of values for alanine residues, three-bond amide-Hα (3JNH-α) coupling constants were found to be between 7.0 and 8.8 Hz, indicating an extended conformation in the backbone (representative 3JNH-α values for compounds 3 and 5 are listed in the Supporting Information). However, 3JNH-α values were between 4.8 and 6.0 Hz for nearly all alanine residues, consistent with backbone angles that resemble twisted (helical) motifs. Interestingly, values of 5.2−5.9 Hz were observed for residues Ala4-Ala7 for the fully active trialanine derivative 9, suggesting an “alanine-extended” short stretch of folded type secondary structure. In contrast, the 3JNH-α values for Val6-Val8 in both 2 and 3 were between 7.9 and 8.7 Hz. Further analysis showed that chemical shift indices26,27 for backbone carbon, nitrogen, and Hα protons (data not shown) suggested that, contrary to our original hypothesis,18 the C-terminal tail may be structured more closely to a β-motif and not α-disposed. Looking at these combined data lead us to conclude that there is little correlation between NMR observables in the C-terminal residues and activity.
The only receptor known for as-APF is CKAP4, a reversibly palmitoylated protein of ∼600 residues that connects the endoplasmic reticulum to microtubules.20,28 Structural studies on CKAP4 reported that the large extracellular/luminal domain of this protein (putatively where APF binds) is a helical coiled coil that forms large aggregates.29 Interestingly, the only other protein isolated from solubilized bladder epithelial cell membrane proteins with APF was vimentin, an intermediate filament protein that also forms helical coiled coils. It is possible that as-APF disrupts protein−protein interactions during multimerization of CKAP4. Alternatively, APF could function by disruption of CKAP4/vimentin coiled−coil interactions. These disruptions may prevent subsequent signaling or internalization of the CKAP4 chaperone protein. The varied “conformations” of active APF analogues also suggest that APF may act at more than one motif on the protein's surface, a tentative explanation for the confounding activity swings with minor changes to the peptide backbone.
In summary, our lead compound, as-APF (2), has a unique and distinct SAR profile that is maintained in Ala9-truncated derivatives. In the current study, we identified two quite potent derivatives of as-APF and determined that Ala6−8 analogue 9 is the “simplest” fully potent derivative of APF synthesized to date. NMR data are beginning to uncover some important clues to the structural implications of specific features of the APF molecule. These data will assist in designing glycopeptidemimetics that could be useful tools in the therapy of IC/PBS and cancer.
Acknowledgments
We thank Toby Chai for providing bladder biopsy specimens from which the explanted bladder epithelial cells were propagated. We also gratefully acknowledge the assistance of the Biophysics Resource in the Structural Biophysics Laboratory, at the Center for Cancer Research, National Cancer Institute.
Supporting Information Available
Experimental details, structures, activity, purity, mass spectroscopy, HPLC, and selected CD and NMR data for new glycopeptides. This material is available free of charge via the Internet at http://pubs.acs.org.
NOTE ADDED AFTER ASAP PUBLICATION
This paper was published on the Web on July 19, 2010 with errors in Table S3 of the Supporting Information file. The revised version was published on July 28, 2010.
Particle Sciences, Inc., 3894 Courtney St., Suite 180, Bethlehem, PA 18017.
Deceased in 2007.
This work was supported by funding from the National Institutes of Health (NIDDK R01 DK52596), as well as funding from the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research.
Supplementary Material
References
- Hanno P. M.Interstitial Cystitis; Springer-Verlag: London, New York, 1990; p xvi, 218 pp. [Google Scholar]
- Parsons J. K.; Parsons C. L. The historical origins of interstitial cystitis. J. Urol. 2004, 171, 20–22. [DOI] [PubMed] [Google Scholar]
- Parsons C. L. Interstitial cystitis: A chronic disease with ever increasing recognition and prevalence. J. Urol. 2005, 173, 98–102. [DOI] [PubMed] [Google Scholar]
- Gillenwater J. Y.; Wein A. J. Summary of the National Institute of Arthritis Diabetes Digestive and Kidney Diseases workshop on Interstitial Cystitis, National Institutes of Health, Bethesda, Maryland, August 28−29, 1987. J. Urol. 1988, 140, 203–206. [DOI] [PubMed] [Google Scholar]
- Keay S.; Zhang C. O.; Trifillis A. L.; Hise M. K.; Hebel J. R.; Jacobs S. C.; Warren J. W. Decreased 3[H]-thymidine incorporation by human bladder epithelial cells following exposure to urine from interstitial cystitis patients. J. Urol. 1996, 156, 2073–2078. [PubMed] [Google Scholar]
- Keay S.; Zhang C. O.; Kagan D.; Hise M. K.; Jacobs S. C.; Hebel R. J.; Gordon D.; Whitmore K.; Bodison S.; Warren J. W. Alterations in HB-EGF, EGF, IGF1 and IGFBP3 levels in the urine of interstitial cystitis patients. J. Urol. 1998, 159, 307–307. [Google Scholar]
- Zhang C. O.; Wang J. Y.; Koch K.; Keay S. Antiproliferative factor (APF) decreases occludin and ZO-1 production and increases paracellular permeability in bladder epithelial cell monolayers. J. Urol. 2005, 173, 81–81. [Google Scholar]
- Zhang C. O.; Wang J. Y.; Koch K. R.; Keay S. Regulation of tight junction proteins and bladder epithelial paracellular permeability by an antiproliferative factor from patients with interstitial cystitis. J. Urol. 2005, 174, 2382–2387. [DOI] [PubMed] [Google Scholar]
- Slobodov G.; Feloney M.; Gran C.; Kyker K. D.; Hurst R. E.; Culkin D. J. Abnormal expression of molecular markers for bladder impermeability and differentiation in the urothelium of patients with interstitial cystitis. J. Urol. 2004, 171, 1554–1558. [DOI] [PubMed] [Google Scholar]
- Johansson S. L.; Fall M. Clinical-Features and Spectrum of Light Microscopic Changes in Interstitial Cystitis. J. Urol. 1990, 143, 1118–1124. [DOI] [PubMed] [Google Scholar]
- Southgate J.; Varley C. L.; Garthwaite M. A. E.; Hinley J.; Marsh F.; Stahlschmidt J.; Trejdosiewicz L. K.; Eardley I. Differentiation potential of urothelium from patients with benign bladder dysfunction. BJU Int. 2007, 99, 1506–1516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith B. H.; Dehner L. P. Chronic Ulcerating Interstitial Cystitis (Hunners Ulcer)—Study of 28 Cases. Arch. Pathol. 1972, 93, 76–81. [PubMed] [Google Scholar]
- Parsons C. L.; Lilly J. D.; Stein P. Epithelial Dysfunction in Nonbacterial Cystitis (Interstitial Cystitis). J. Urol. 1991, 145, 732–735. [DOI] [PubMed] [Google Scholar]
- Fowler J. E.; Lynes W. L.; Lau J. L. T.; Ghosh L.; Mounzer A. Interstitial Cystitis Is Associated with Intraurothelial Tamm-Horsfall Protein. J. Urol. 1988, 140, 1385–1389. [DOI] [PubMed] [Google Scholar]
- Tomaszewski J. E.; Landis J. R.; Russack V.; Williams T. M.; Wang L. P.; Hardy C.; Brensinger C.; Matthews Y. L.; Abele S. T.; Kusek J. W.; Nyberg L. M.; Stu I. C. D. Biopsy features are associated with primary symptoms in interstitial cystitis: Results from the Interstitial Cystitis Database Study. Urology 2001, 57, 67–81. [DOI] [PubMed] [Google Scholar]
- Eldrup J.; Thorup J.; Nielsen S. L.; Hald T.; Hainau B. Permeability and Ultrastructure of Human Bladder Epithelium. Br. J. Urol. 1983, 55, 488–492. [DOI] [PubMed] [Google Scholar]
- Keay S. K.; Szekely Z.; Conrads T. P.; Veenstra T. D.; Barchi J. J.; Zhang C. O.; Koch K. R.; Michejda C. J. An antiproliferative factor from interstitial cystitis patients is a frizzled 8 protein-related sialoglycopeptide. Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 11803–11808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaczmarek P.; Keay S. K.; Tocci G. M.; Koch K. R.; Zhang C. O.; Barchi J. J.; Grkovic D.; Guo L.; Michejda C. J. Structure-activity relationship studies for the peptide portion of the bladder epithelial cell antiproliferative factor from interstitial cystitis patients. J. Med. Chem. 2008, 51, 5974–5983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saitoh T.; Hirai M.; Katoh M. Molecular cloning and characterization of human Frizzled-8 gene on chromosome 10p11.2. Int. J. Oncol. 2001, 18, 991–996. [DOI] [PubMed] [Google Scholar]
- Conrads T. P.; Tocci G. M.; Hood B. L.; Zhang C. O.; Guo L.; Koch K. R.; Michejda C. J.; Veenstra T. D.; Keay S. K. CKAP4/p63 is a receptor for the frizzled-8 protein-related antiproliferative factor from interstitial cystitis patients. J. Biol. Chem. 2006, 281, 37836–37843. [DOI] [PubMed] [Google Scholar]
- Liff M. I.; Lyu P. C.; Kallenbach N. R. Analysis of Asymmetry in the Distribution of Helical Residues in Peptides by H-1 Nuclear-Magnetic-Resonance. J. Am. Chem. Soc. 1991, 113, 1014–1019. [Google Scholar]
- Cheng T. J.; Zhao Y.; Li X.; Lin F.; Xu Y.; Zhang X. L.; Li Y.; Wang R. X.; Lai L. H. Computation of octanol-water partition coefficients by guiding an additive model with knowledge. J. Chem. Inf. Model. 2007, 47, 2140–2148. [DOI] [PubMed] [Google Scholar]
- Lyu P. C.; Sherman J. C.; Chen A.; Kallenbach N. R. Alpha-Helix Stabilization by Natural and Unnatural Amino-Acids with Alkyl Side-Chains. Proc. Natl. Acad. Sci. U.S.A. 1991, 88, 5317–5320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seebach D.; Beck A. K.; Bierbaum D. J. The world of beta- and gamma-peptides comprised of homologated proteinogenic amino acids and other components. Chem. Biodiversity 2004, 1, 1111–1239. [DOI] [PubMed] [Google Scholar]
- Greenfield N. J. Analysis of the kinetics of folding of proteins and peptides using circular dichroism. Nat. Protoc. 2006, 1, 2891–2899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berjanskii M. V.; Wishart D. S. A simple method to predict protein flexibility using secondary chemical shifts. J. Am. Chem. Soc. 2005, 127, 14970–14971. [DOI] [PubMed] [Google Scholar]
- Shen Y.; Delaglio F.; Cornilescu G.; Bax A. TALOS plus: A hybrid method for predicting protein backbone torsion angles from NMR chemical shifts. J. Biomol. NMR 2009, 44, 213–223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Planey S. L.; Keay S. K.; Zhang C. O.; Zacharias D. A. Palmitoylation of Cytoskeleton Associated Protein 4 by DHHC2 Regulates Antiproliferative Factor-mediated Signaling. Mol. Biol. Cell 2009, 20, 1454–1463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klopfenstein D. R.; Klumperman J.; Lustig A.; Kammerer R. A.; Oorschot V.; Hauri H. P. Subdomain-specific localization of CLIMP-63 (p63) in the endoplasmic reticulum is mediated by its luminal alpha-helical segment. J. Cell Biol. 2001, 153, 1287–1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.