

Abstract
Background
Breast milk contains complex nutrients and facilitates the maturation of various biological systems in infants. Exosomes, membranous vesicles of endocytic origin found in different body fluids such as milk, can mediate intercellular communication. We hypothesized that microRNAs (miRNAs), a class of non-coding small RNAs of 18–25 nt which are known to be packaged in exosomes of human, bovine and porcine milk, may play important roles in the development of piglets.
Results
In this study, exosomes of approximately 100 nm in diameter were isolated from porcine milk through serial centrifugation and ultracentrifugation procedures. Total RNA was extracted from exosomes, and 5S ribosomal RNA was found to be the major RNA component. Solexa sequencing showed a total of 491 miRNAs, including 176 known miRNAs and 315 novel mature miRNAs (representing 366 pre-miRNAs), which were distributed among 30 clusters and 35 families, and two predicted novel miRNAs were verified targeting 3’UTR of IGF-1R by luciferase assay. Interestingly, we observed that three miRNAs (ssc-let-7e, ssc-miR-27a, and ssc-miR-30a) could be generated from miRNA-offset RNAs (moRNAs). The top 10 miRNAs accounted for 74.5% (67,154 counts) of total counts, which were predicted to target 2,333 genes by RNAhybrid software. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses using DAVID bioinformatics resources indicated that the identified miRNAs targeted genes enriched in transcription, immunity and metabolism processes, and 14 of the top 20 miRNAs possibly participate in regulation of the IgA immune network.
Conclusions
Our findings suggest that porcine milk exosomes contain a large number of miRNAs, which potentially play an important role in information transfer from sow milk to piglets. The predicted miRNAs of porcine milk exosomes in this study provide a basis for future biochemical and biophysical function studies.
Electronic supplementary material
The online version of this article (doi:10.1186/1471-2164-15-100) contains supplementary material, which is available to authorized users.
Keywords: Porcine milk exosomes, Solexa sequencing, miRNA
Background
Milk, as the sole source of nutrition for infants, contains a potent mixture of diverse components such as milk fat globules (MFG) [1], immune competent cells and soluble proteins, for instance IgA, cytokines and antimicrobial peptides [2], which can provide protection against infections in newborns [3]. In addition, breast milk may have a role in tolerance induction [1] and may protect infants from developing allergies [4].
Exosomes are small (30–100 nm) membrane vesicles of endocytic origin that are released into the extracellular environment upon fusion of multivesicular bodies (MVB) with the plasma membrane [5]. Many cells have the capacity to release exosomes, including reticulocytes [6], dendritic cells [7], B cells [8], T cells [9], mast cells [10], epithelial cells [11] and tumor cells [12]. In addition, exosomes have been found in physiological fluids, such as saliva [13, 14], plasma [15], urine [16], amniotic fluid [17], malignant ascites [18], bronchoalveolar lavage fluid [19] and synovial fluids [20]. Several studies have suggested that exosomes, which contain proteins, mRNA and microRNA (miRNA), stimulate and transfer surface receptors to target cells [21–23], as well as serve as novel vehicles for genetic exchange between cells [24]. As with other biological fluids, microvesicle-like particles are also present in mouse milk [25] and human milk [26]. Recent published studies have isolated mRNAs and miRNAs from bovine milk-derived microvesicles [27]. One study via deep sequencing technology identified 602 unique miRNAs originating from 452 miRNA precursors (pre-miRNAs) in human breast milk exosomes and found that, out of 87 well-characterized immune-related pre-miRNAs, 59 (67.82%) were enriched in breast milk exosomes [28]. Recently, porcine milk was reported to contain 180 pre-miRNAs, including 140 known and 40 novel porcine pre-miRNAs, altogether encoding 237 mature miRNAs [29].
MiRNAs are widespread among eukaryotes and represent key components of a conserved system of RNA-based gene regulation [30–33]. Many studies have demonstrated that miRNAs are key post-transcriptional regulators of gene expression and play important roles in a wide range of physiological and pathological processes [34], including development, differentiation, proliferation and immune responses. It is believed that about 60% of mammalian genes are regulated by miRNAs [35–39].
Aside from being important farm livestock, pigs are also model animals for medical research. In the present study, we investigated miRNAs in milk exosomes of Landrace pigs in order to provide new information for investigations into the physiological functions of porcine milk.
Methods
Milk samples
Porcine milk samples were collected between day 1 to 5 after parturition from healthy lactating Landrace female pigs bred in the breeding farm of the Livestock Research Institute (Guangzhou, China). Milk samples were frozen immediately after milking and were kept at-80°C until use.
Preparation of exosomes from milk
Porcine milk samples were centrifuged first at 2,000 × g for 30 min at 4°C to remove MFGs as well as mammary gland-derived cells. Defatted samples were then subjected to centrifugations at 4°C for 30 min at 12,000 × g to remove residual MFGs, casein and other debris. Subsequently, from the final supernatant (so-called whey or milk serum), the membrane fraction was prepared by ultracentrifugation at 110,000 × g for 2 h in an SW41T rotor (Beckman Coulter Instruments, Fullerton, CA) [40].
Transmission electron microscopy (TEM)
The final fraction obtained as described above was diluted with 0.01 M PBS and ultracentrifuged again to recover microvesicles as pellets. Following fixation in 2% glutaraldehyde, microvesicles were negatively stained with uranyl acetate and observed by TEM (JEOL JEM2000EX, Tokyo, Japan).
RNA isolation and Solexa sequencing
Total RNA was isolated from samples collected after ultracentrifugation using Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s protocol. The quality of RNA was examined by 2% agarose gel electrophoresis and with a Biophotometer 6131 (Eppendorf, Germany), as well as further confirmed by using a Bioanalyzer (Agilent Technologies, Santa Clara, CA). Small RNAs (18–30 nt) were obtained from the total RNA, 5’ and 3’ adaptors were ligated to the small RNAs, then the adaptor-ligated RNAs were subsequently transcribed into cDNA by RT-PCR, and the samples were amplified by PCR using primers complementary to the two adaptors. The PCR products were purified and subjected to Solexa sequencing (Illumina, CA) at the Beijing Genomics Institute (BGI, Shenzhen, China).
Sequence data analysis
The raw reads obtained from Solexa sequencing were processed to obtain clean reads by summarizing data production, evaluating sequencing quality, calculating the length distribution of small RNA reads, removing low quality reads and adaptor sequences as described in previous paper [41]. All the clean reads were aligned against non-coding RNAs from the GenBank and Rfam (11.0) (ftp.sanger.ac.uk/pub/databases/Rfam) database to annotate and classify rRNA, tRNA, snRNA and other ncRNA sequences using tag2 annotation software (developed by BGI). Then the selected sequences were mapped to the pig genome (sscrofa9, http://www.ensembl.org/Sus_scrofa/) using SOAPv1.11 software [42] to analyze their expression and distribution. Subsequently, the miRNA candidates were further analyzed by miRDeep 2 against all known miRNAs and porcine miRNA precursors (miRBase 20.0). All remaining candidates who did not map to any miRNAs in miRBase 20.0 were considered as potential novel miRNAs. To further identify these potential novel miRNA candidates, software MIREAPv0.2 (http://sourceforge.net/projects/mireap) [43] developed by BGI was used to predict novel miRNA by exploring the secondary structure, the Dicer cleavage site and the minimum free energy of the annotated small RNAs which could be mapped to genome. In briefly, the sequence length should be between 18–26 nt, maximal free energy allowed for a miRNA precursor was -18 kcal/mol, maximal space between miRNA and miRNA* was 35 nt, and flank sequence length of miRNA precursor should be 10 nt. Finally, all remaining novel miRNA candidates were further subjected to MiPred (http://www.bioinf.seu.edu.cn/miRNA/) to filter out pseudo-pre-miRNAs. The minimum free energy must be > -20 kcal/mol or P-value was >0.05 [44], and their secondary structures were also checked using the Mfold3.2 software [45]. All data for analysis in this study have been deposited in https://mynotebook.labarchives.com/share/allinchen/MTkuNXwxMzMxMS8xNS0yL1RyZWVOb2RlLzE1NzEyODU2fDQ5LjU= with a DOI:10.6070/H4DN432G.
PCR and qRT-PCR identification of known and novel miRNAs
Total RNA (identical sample to that of the Solexa sequencing sample) was first digested with DNase I (Invitrogen), and 2 μg of total RNA was reverse transcribed to poly (A) tail-added cDNA using the One Step PrimeScript miRNA cDNA Synthesis Kit (TaKaRa, Dalian) according to the manufacturer’s instructions. Briefly, a 60-nt adaptor containing a poly (A) structure was added to the 3’ sequence of miRNAs, which were then reverse transcribed to an 80-bp cDNA sequence [46]. The cDNA was diluted 5-fold with ddH2O, and PCR was performed on a Bio-Rad system (BIO-RAD,USA )in a final 20 μL volume reaction containing 2 μl PCR cDNA, 10 μL of 2× PCR Mix (Toyobo, Osaka, Japan) and 1 mM of each primer. The real-time PCR thermal profile was as follows: 5 min at 95°C, 40 cycles of 30 s at 94°C, 30 s at the corresponding annealing temperature (Tm) and 72°C for 30 s, followed by 72°C at 10 min. PCR products were examined on an agarose gel to confirm that a single PCR product was generated, and 5S ribosomal RNA was used as an internal control for the PCR. The miRNA forward primer was designed with Primer 5.0 ( Table 1), and the reverse primer for miRNAs was the Uni-miR qPCR Primer offered by the kit One Step PrimeScript miRNA cDNA Synthesis Kit (TaKaRa, Dalian).
Table 1.
PCR primers for miRNAs
| miRNAs name | Primer sequence | Renaturation temperature |
|---|---|---|
| ssc-let-7e | TGAGGTAGGAGGTTGTATAGTT | 59.5°C |
| ssc-miR-21 | GCTAGCTTATCAGACTGATGTTG | 59.5°C |
| ssc-miR-206 | TGGAATGTAAGGAAGTGTGTG | 59.5°C |
| ssc- let-7i | GCCGCTGAGGTAGTAGTTTGTGCT | 59.5°C |
| ssc-miR-140 | GACAGTGGTTTTACCCTATGGTA | 59.5°C |
| ssc-miR-92b-5p | TTATAGGGACGGGACGCGGTG | 59.5°C |
| ssc-miR-22b-3p | AAGCTGCCAGTTGAAGAACTG | 59.5°C |
| ssc-miR-28-5p | GAAGGAGCTCACACTCTATTGA | 59.5°C |
| ssc-miR-205 | TCCTTCATTCCACCGGAGTCT | 59.5°C |
| ssc-miR-451 | AAACCGTTACCATTACTGAGTT | 59.5°C |
| ssc-miR-125b | TCCCTGAGACCCTAACTTGTG | 59.5°C |
| ssc-miR-9 | GCGGTCTTTGGTTATCTAGCTGT | 59.5°C |
| ssc-let-7c | TGAGGTAGTAGGTTGTATGGT | 59.5°C |
| P-m0281-5p(PS-16) | TCTCCCAACCCTTGTACCA | 58°C |
| P-m0124-3p(PC-280) | TGTTCCGAGATTGGGCTG | 58°C |
| P-m0227-5p(PC-291) | TTCCTGAGTCGGACTGGG | 58°C |
| P-m0355-5p(PC-82) | CCCAGGATCAGAGGATGG | 58°C |
| P-m0338-3p(PC-241) | TCTGTGAACTAGAAACCTCTGG | 58°C |
| P-m0105-3p(PC-72) | CATTTGATTCAGTTGGACACT | 58°C |
| P-m0113-3p(PC-130) | CTATGGATCTAGGAGGACGC | 58°C |
| P-m0129-5p(PC-129) | CTATGGATCTAAGAGGACACCC | 58°C |
| P-m0058-5p(PC-276) | TGTGTGTGATCGTTAATGTGC | 58°C |
| P-m0279-5p(PC-192) | GTCCTTGGTGAGTCGGATG | 58°C |
| P-m0103-3p(PC-70) | CATTGCTTTGATCGTCTGG | 58°C |
| P-m0265-3p(PC-139) | CTGGAAGGATTTGGGTAGG | 58°C |
| P-m0210-5p(PC-277) | TGTGTGTTCTGTCGGATGAG | 58°C |
| P-m0186-5p(PC-73) | CATTTTAAGGATCGTGTGGG | 58°C |
| P-m0070-3p(PC-63) | CAGTAGGGATGAGAGGACACT | 58°C |
miRNAs target prediction and plasmid construction
Two predicted novel miRNAs, named miR-PC-86 and miR-PC-263, were selected to predict their target genes in pig genome using the RNAhybrid software (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/) with its own algorithm. The 3’-UTR sequences of porcine transcripts in whole genome were obtained from ensemble gene 66 (sscorfa 9, http://www.ensembl.org/Sus_scrofa/). The 3’-UTR of IGF-1R contains the highly conserved binding sites for the two miRNAs, and the sequence (104 bp) is as follows:TCCTGGATCCCGATCCCGTGCAAACAGTACCGTGCGCACGCGGGCGGGCGGGGGGAGAGTTTTAACAATCTATTCACAAGCCTCCTGTACCTCAGTGGATCTTC. Further, the 3’-UTR sequence was inserted into pmirGLO Vector (Promega) with XhoI and XbaI double digestion to construct recombinant Dual-Luciferase reporter vector, named as pGLO-IGF-1R-3’UTR (Figure 1A). Meanwhile, a plasmid containing mutant IGF-1R 3′-UTR, named as pGLO-IGF-1R-3’UTR-delete (Figure 1A), was generated by deleting the core sequence of the two miRNA binding sites through DNA synthesis (Sangon Biotech (Shanghai) Co., Ltd.), the sequence is as follows: TCCTGGATCCCGATCCCGTGCAAACAGTACCGTGCGCACGCGGGCGGGCGGGGGGAGAGTTTTAACAATCTATTCACAAGCCTCCTGTACCC.
Figure 1.
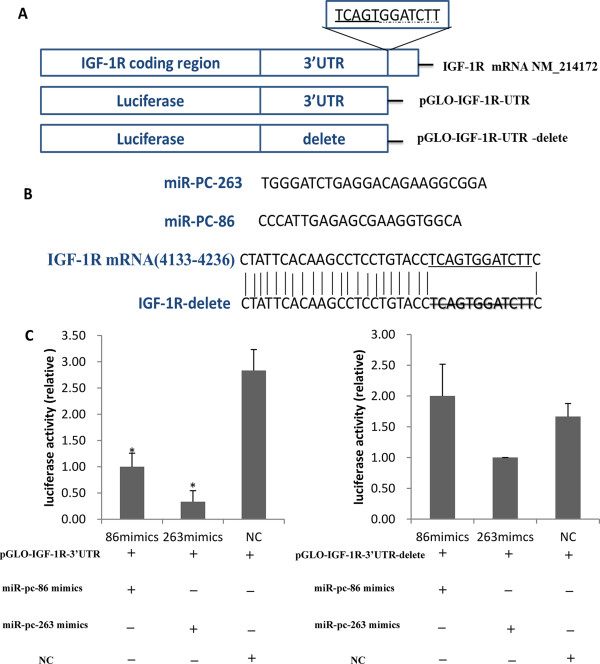
MiR-PC-86 and MiR-PC-263 directly regulate IGF-1R expression via 3’ UTR sites. (A) Schematic of IGF-1R mRNA and the luciferase reporter plasmids containing the miR-PC-86 and miR-PC-263 binding sites of IGF-1R mRNA. The 3’ UTR sites were inserted downstream of the luciferase reporter, as indicated. TCAGTGG was the predicted target site of miR-PC-86, GGATCTT was the predicted target site of miR-PC-263. (B) miR-PC-86 and miR-PC-263 sequences and predicted binding site between miR-PC-86 and miR-PC-263 and IGF-1R mRNA. IGF-1R mRNA has one putative binding site for miR-PC-86/ miR-PC-263 on the 3’ UTR. Twelve nucleotides TCAGTGGATCTT of IGF-1R 3’ UTR (underlined) were delete in order to disrupt the binding with miR-PC-86 and miR-PC-263 seed regions. (C) IPEC-J2 cells were transfected with each of the constructed plasmids, together with miR-PC-86/ miR-PC-263and Renilla luciferase reporter plasmid (*P < 0.05, n = 6).
Leuciferase reporter assay
IPEC-J2 cells were maintained in DMEM/F12 (1:1) (GIBCO) and supplemented with 10% fetal bovine serum (FBS, GIBCO), 5 ng/ml EGF (peprotech, USA) and 5 ug/ml insulin (Sigma, USA). Lipofectamine 2000 (Invitrogen) was used for transfection. Cells (10,000) were plated in a 96-well plate. After 24 h cultivation, cells were transfected with a mixture including 500 ng pGLO-IGF-1R-3’UTR or pGLO-IGF-1R-3’UTR-delete construct and 30pM of miR-PC-86 or miR-PC-263 mimics (GenePharma, Shanghai, China). For control, 500 ng of pmirGLO-scramble including a scrambled sequence of the miRNA target sequence was used. Cells were collected 48 h after transfection, and luciferase activity was measured using a Dual-GLO luciferase reporter assay system (Promega). Statistical differences between treatment and control groups were determined using Student’s t-test, at P < 0.05.
Bioinformatics analysis
Chromosomal localization and cluster analysis of miRNAs
Pre-miRNAs of all miRNAs (known miRNAs and novel miRNAs) were mapped to the porcine genome (sscrofa9, http://www.ensembl.org/Sus_scrofa/) according to their positions on the chromosomes. Pre-miRNA positions less than 10 kb apart were considered to belong to the same miRNA cluster.
Target prediction and Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses
Porcine miRNA targets were obtained at the genome level. In brief, miRNA targets were predicted using the RNAhybrid software algorithm (http://bibiserv.techfak.uni-bielefeld.de/rnahybrid/) in 3’-UTR sequences of transcripts from the whole pig genome obtained from Ensembl Gene 66 database (sscrofa9, http://www.ensembl.org/Sus_scrofa/). Strict criteria (perfect match of 2–8 nt in the seed region; no more than -25 kcal/mol low free energy of miRNAs-mRNA binding) were applied to the target prediction procedure. GO and KEGG pathway analyses were performed using DAVID bioinformatics resources (http://david.abcc.ncifcrf.gov/).
Results
Identification of exosomes
Exosomes were obtained from porcine milk by ultracentrifugation. After negative staining, approximately round-shaped porcine milk exosomes with diameters of ~50-100 nm were observed by TEM, showing a greater density at the center than at the boundary (Figure 2A, B).
Figure 2.
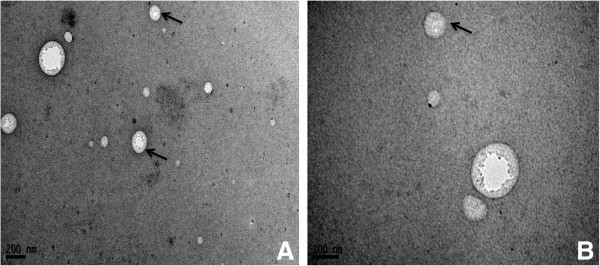
Exosomes detected by TEM. The exosomes appeared as round or oval microvesicles (A, B), with a diameter of 50–150 nm and heavier density at the center than on the margin.
Porcine milk exosomes contain RNA
To further ascertain whether the porcine milk exosomes collected by ultracentrifugation contained RNA, we extracted the samples using Trizol reagent and then examined the recovered product by electrophoresis on a 2% agarose gel. To exclude the possibility of DNA contamination, total RNA was incubated at 37°C for 30 min with DNase I or RNase. The extracted RNA could not be digested by DNase I, while it could be degraded by RNase (Figure 3A). These results demonstrated that the porcine milk exosomes contained RNA. More interestingly, total RNA of porcine milk exosomes were enriched with 5S rRNA (Figure 3B), consistent with previous studies [27, 28, 47].
Figure 3.
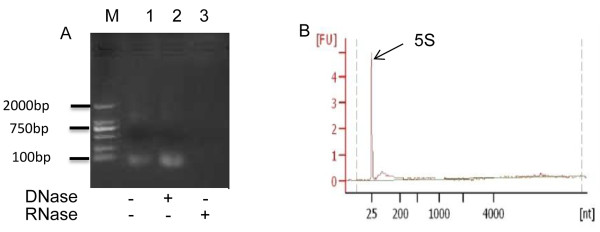
Milk-derived exosomes containing RNA. (A) Total RNA was extracted from porcine exosomes. M, 1, 2 and 3 represent the marker (DL 2000), RNA without any treatment, RNA treated with DNase I and RNA treated with RNase, respectively. (B) RNA sample analyzed by the Agilent Bioanalyzer 2100.
Solexa sequencing and analysis
Solexa sequencing
The sRNAs were enriched from porcine exosomes to construct a library for Solexa sequencing. We obtained 9,033,167 raw reads and 6,013,724 high qualities reads after removal of low quality and contaminant reads. Among the high quality reads, there were 4,964,542 clean reads (82.55%), representing 1,691,655 unique sRNAs. The majority of the sRNAs in porcine milk exosomes were 18–25 nt in length (74.89%, Figure 4), with 2,458,894 reads (49.53%) representing 872,096 unique sRNAs (51.55%), including miRNAs and other sRNAs, such as rRNA, tRNA, snRNA, snoRNA, scRNA, small recognition particle RNA (srpRNA), repetitive sequence elements and unannotated sequences, which could be mapped to the pig genome. BLAST searching with the miRbase 20.0, identified a total of 230,216 reads, representing 1,555 unique known miRNAs. Due to RNA bias editing, 5’ modifications and 3’ modifications, many pre-miRNAs produce multiple mature miRNA isoforms, namely isomiRNAs, as described in many studies [48–50]. In the subsequent analysis, all isomiRNAs generated from the same precursor were considered one type of miRNA. Consequently, these 1,555 unique miRNAs corresponded to 176 known mature miRNAs (205 pre-miRNAs, all the detail were listed in Additional file 1: Table S1). In addition, we identified 315 novel mature miRNAs (generated from 366 pre-miRNAs, detail in Additional file 2: Table S2). Among the 315 novel miRNAs, 18 have not been deposited as porcine miRNAs in miRbase 20.0, but had very high similarity with miRNA sequences of other species. These 18 miRNAs are labeled “PS” (Table 2), while 298 miRNAs that have not been deposited in miRbase 20.0 for any organism are labeled as “PC” and presented in Additional file 2: Table S2.
Figure 4.
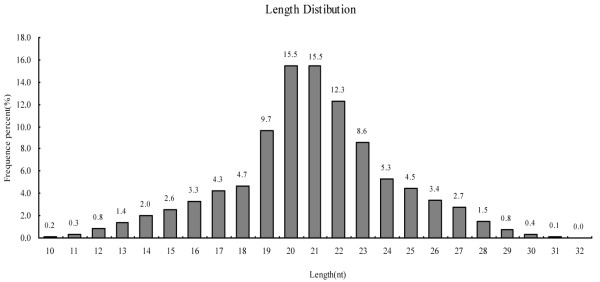
Length distribution of miRNAs reads from Solexa sequencing. A total of 4,964,542 clean reads were obtained, ranging from 10 to 32 nt, most of which were 18–25 nt in length (accounting for 74.89%).
Table 2.
Porcine novel miRNAs conserved in other species (miRBase release 20.0)
| Unique ID | miRNAs name | Count | Sequence | Size | Conservation | Match |
|---|---|---|---|---|---|---|
| PS-1 | miR-290-5p | 14 | ACTCAAACTGTGGGGGCACTTT | 22 | mmu(#) | 1nt sub(#) |
| PS-2 | miR-378c | 33 | ACTGGACTTGGAGTCAGAAGT | 21 | hsa | 4nt delete |
| PS-3 | miR-20b-3p | 4 | ACTGTAGTGTGGGCACTTCCAGT | 23 | hsa | 1nt add, 1nt sub |
| PS-4 | miR-219-3p | 11 | AGAATTGTGGCTGGACATCT | 20 | bta | 1nt delete |
| PS-6 | miR-138-5p | 42 | AGCTGGTGTTGTGAATCAGGCCG | 23 | mmu | perfect |
| PS-7 | miR-31-5p | 5 | AGGCAAGATGCTGGCATAGCT | 21 | has | perfect |
| PS-9 | let-7f-1-3p | 2 | CTATACAATCTATTGCCTTCC | 21 | rno | perfect |
| PS-11 | miR-874-3p | 25 | CTGCCCTGGCCCGAGGGACCGA | 22 | mmu | perfect |
| PS-12 | miR-551a | 225 | GCGACCCACTCTTGGTTTCC | 20 | hsa | 1nt delete |
| PS-13 | miR-138-3p | 1 | GCTACTTCACAACACCAGGGT | 21 | hsa | 1nt sub, 1delete |
| PS-14 | miR-182-3p | 1 | GGTGGTTCTAGACTTGCCAACT | 22 | mmu | 1nt insert |
| PS-15 | miR-5003-3p | 58 | TATTTAATAGGTTGTTGGGA | 20 | hsa | 2nt sub, 2nt delete |
| PS-16 | miR-150-5p | 164 | TCTCCCAACCCTTGTACCAGT | 21 | mmu | 1nt delete |
| PS-17 | miR-2411-3p | 7 | TGAACTGTCATACTCCCACATC | 22 | bta | 3nt delete, 1nt sub |
| PS-18 | let-7f-5p | 20 | TGAGGTAGTAGATTGTATAGTTG | 23 | hsa | 1nt insert |
| PS-19 | miR-31-3p | 3 | TGCTATGCCAACATATTGCCA | 21 | has | 1nt delete |
| PS-20 | miR-182 | 46 | TTTGGCAATGGTAGAACTCACA | 22 | dre | perfect |
| PS-21 | miR-96-5p | 65 | TTTGGCACTAGCACATTTTTGCT | 23 | hsa | perfect |
#: mmu, Mus musculus; hsa, Homo sapiens; bta, Bos taurus; dre, Danio rerio; rno, Rattus norvegicus;
“sub”, “delete”, “add” represents nucleotide substitution, deletion, addition (at 5-end of miRNAs), respectively perfect stands for perfect match with reference miRNAs.
There were 73 miRNAs with more than 100 counts and 264 miRNAs with less than 10 counts. The top 10, top 20, top 50 and top 100 miRNAs accounted for 74.5%, 85.2%, 95.3% and 98.3% of the reads, respectively (Figure 5A). The top 10 miRNAs were ssc-miR-193a-3p, ssc-miR-423-5p, ssc-miR-320, ssc-miR-181a, ssc-miR-30a-3p, ssc-miR-378, ssc-miR-191, ssc-let-7a, ssc-let-7f and ssc-let-7c. With 67,154 counts (29.6%, average count: 460.6) (Figure 5B), ssc-miR-193a-3p ranked first among all miRNAs reads.
Figure 5.
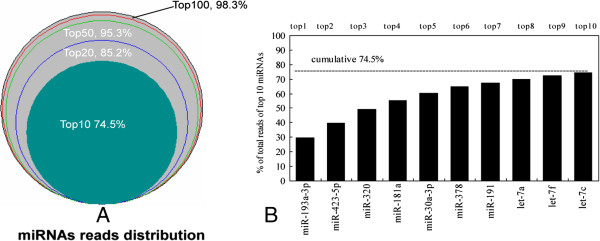
Distribution of miRNA reads and top 10 miRNAs. (A) The distribution of miRNA reads showed that the top 10, top 20, top 50 and top 100 miRNAs accounted for 74.5%, 85.2% and 95.3% and 98.3% of total reads. (B) Cumulative proportions of top 10 miRNAs. MiR-193-3p ranked first, accounting for 29.6% of total reads.
Identification of miRNAs by PCR and sequencing
To verify the deep sequencing results, we selected randomly 13 known miRNAs and 15 predicted novel miRNAs for PCR amplification (Figure 6A, B). Subsequently, the 15 newly predicted miRNAs were cloned and sequenced. The results showed that 8 sequences were fully matched, while 7 sequences had one or two mismatched nucleotides (Table 3). However their seed sequences remained unchanged. Simultaneously, the abundance of some novel miRNAs predicted by Solexa sequencing was confirmed by quantitative real-time PCR. The abundance of most miRNAs observed by qPCR of the sample pool and by sequencing were generally consistent (Figure 7).
Figure 6.
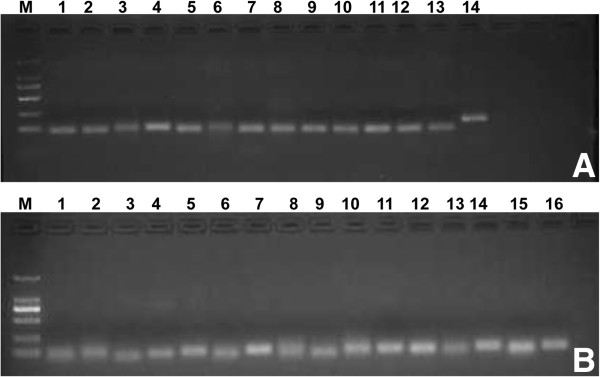
MiRNAs detected randomly in porcine milk. (A) Known miRNAs from miRBase (18.0), from M to 14, respectively: marker (DL 2000), ssc-let-7e, ssc-miR-21, ssc-miR-206, ssc-let-7i, ssc-miR-140, ssc-miR-92b-5p, ssc-miR-22-3p, ssc-miR-28-5p, ssc-miR-205, ssc-miR-451, ssc-miR-125b, ssc-miR-9, ssc-let-7c and 5 s (control). (B) Top 15 predicted novel miRNAs, from M to 16, respectively: marker (DL 2000), P-m0227-5p, P-m0338-3p, P-m0105-3p, P-m0058-5p, P-m0281-5p, P-m0265-3p, P-m0279-5p, P-m0103-3p, P-m0113-3p, P-m0129-5p, P-m0355-5p, P-m0210-5p, P-m0070-3p, P-m0124-3p, P-m0186-5p and 5 s (control).
Table 3.
miRNAs matched to sequecing
| Predict new miRNA | Matched sequence |
|---|---|
| P-m0281-5p |
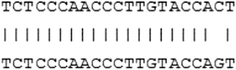
|
| P-m0124-3p |

|
| P-m0227-5p |

|
| P-m0355-5p |

|
| P-m0338-3p |

|
| P-m0105-3p |

|
| P-m0113-3p |
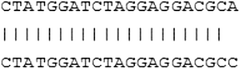
|
| P-m0129-5p |

|
| P-m0058-5p |

|
| P-m0279-5p |

|
| P-m0103-3p |

|
| P-m0265-3p |

|
| P-m0210-5p |

|
| P-m0186-5p |

|
| P-m0070-3p |

|
Figure 7.
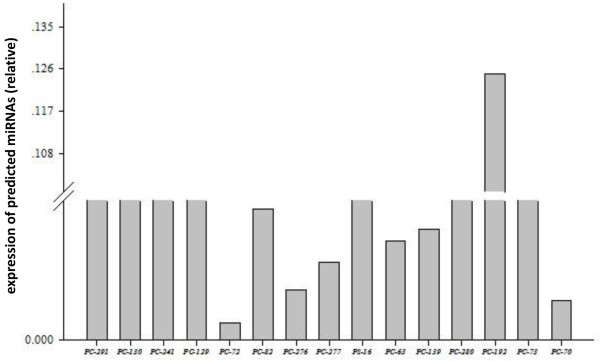
Expression of 15 predicted novel miRNAs in the sample pool detected by qRT-PCR. Trends in relative expression by qRT-PCR and counts from Solexa sequencing of miRNAs, except for PC-192, were consistent.
Target verification of miR-PC-86 and miR-PC-263 against 3’UTR of IGF-1R using luciferase report assay
To investigate whether the predicted miR-PC-86 and miR-PC-263 (Figure 1) were functional novel miRNAs, target genes were predicted, and miR-PC-86/ miR-PC-263 were found to directly target IGF-1R 3’UTR sequence. The full-length 3’UTR of IGF-1R mRNA was inserted downstream of the luciferase gene in the pmirGLO Dual-Luciferase miRNA Target Expression Vector reporter plasmid, and the seed sequence was also delete to disrupt miR-PC-86/ miR-PC-263 binding (Figure 1B). The wild-type (pGLO-IGF-1R-3’UTR) or delete (pGLO-IGF-1R-3’UTR-delete) plasmid was co-transfected with the miR-PC-86 and miR-PC-263 mimics into IPEC-J2 cells. Forty-eight hours after transfection, the luciferase activity of the miR-PC-86 and miR-PC-263 group were significantly lower than that of the NC group (P < 0.05) respectively, and the reduction was rescued in the delete group (Figure 1C). Thus, IGF-1R was initially confirmed as the target of miR-PC-86 and miR-PC-263.
Genomic localization of pre-miRNAs
To further establish the presence of miRNA precursors in the genome, all mature miRNAs (176 known and 315 novels) were mapped to the S. scrofa genome (Figure 8). As a result, 176 known mature miRNAs were mapped to 205 pre-miRNAs, and 315 novel miRNAs were mapped to 366 pre-miRNAs on the chromosomes. Our analysis revealed that the genomic density distribution of porcine milk pre-miRNAs (number of pre-miRNAs per Mb of each chromosome) was heterogeneous (Figure 8), ranging from 0.45 to 0.11 pre-miRNAs for 1 M of genomic sequence. Chromosomes with the highest and lowest densities of pre-miRNAs were chromosome 12 (29 pre-miRNAs per 64 Mbp) and chromosome 13 (25 pre-miRNAs per 219 Mbp), respectively. Interestingly, the medium-length X chromosome (144 Mbp, ranking 10th in length among the 19 chromosomes in pigs) was an exception by encoding an intermediate number (25 out of 366, 6.8%) of pre-miRNAs, corresponding to 0.17 pre-miRNAs for 1 M of genome sequence, but yet contained the most clusters.
Figure 8.
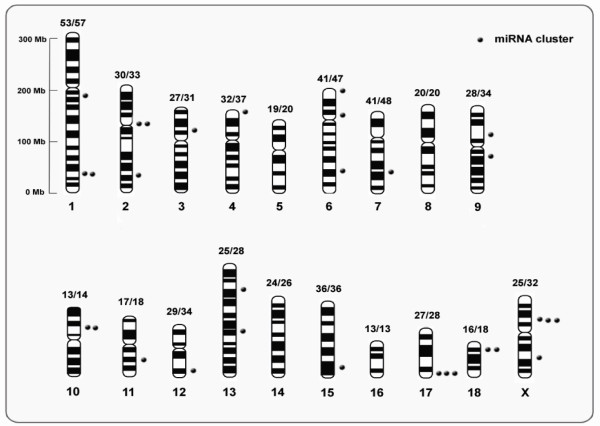
Distribution of 30 miRNA clusters. The number of base points near different bars indicates the number of clusters in the chromosome. The relative vertical dimension of the point on the bar represents the location of cluster. The label “number1 /number2” above every bar indicates the value of “pre-miRNAs/mature miRNA”.
In addition, we observed many mature miRNAs having multiple miRNA precursors located in the same or different chromosomes. Of the novel predicted miRNAs, 40 pre-miRNAs had two copies in the genome, 7 pre-miRNAs had 3 copies, 2 pre-miRNAs had 4 copies, 1 pre-miRNA had 8 copies and 249 pre-miRNAs were unique. With regard to known miRNAs, 4 pre-miRNAs had 3 copies, 22 pre-miRNAs had two copies and 149 pre-miRNAs had only one copy.
Mature miRNAs
It is well accepted that only one of two strands generated from a precursor is preferentially incorporated into RNA-induced silencing complexes (RISC), whereas the complementary strand (miR*) may be degraded. Closer examination of mature miRNAs generated from pre-miRNAs showed that precursor miRNAs could be divided into three groups (Table 4): pre-miRNAs only with the left-arm sequence (5p), pre-miRNAs only with the right-arm sequence or both. Most pre-miRNAs seemed to be single-arm miRNAs (5p or 3p), while 50 pre-miRNAs possessed both 5p and 3p sequences (40 coupled mature miRNAs, all details were listed in Additional file 3: Table S3). Further analysis of the 40 coupled mature miRNAs indicated that most of the pre-miRNAs had no significant difference in abundance between the 5p-arm and 3p-arm sequences (Figure 9). Some miRNAs had different expression levels between the two strands. For example, ssc-miR-193a-3p had 67,154 counts, ranking first among all miRNAs, while ssc-miR-193a-5p had 2,538 counts. Conversely, miR-423-5p had 22,588 counts, while its complementary strand, miR-423-3p, had only 654 reads, and which was shared by miR-22, miR-30a, miR-339, miR-17, miR-24, miR-331, miR-27b and let-7d (detail in Additional file 3: Table S3).
Table 4.
Pre-miRNAs and their corresponding mature miRNAs type
| Pre-miRNAs | miRNA-5p | miRNA-3p | Both | Mature miRNAs | |
|---|---|---|---|---|---|
| known | 205 | 67 | 57 | 33(26)# | 176 |
| novel | 366 | 150 | 139 | 17(13) | 315 |
| total | 571 | 217 | 196 | 50(39) | 491 |
#Number in bracket represents mature miRNAs couple number.
Figure 9.
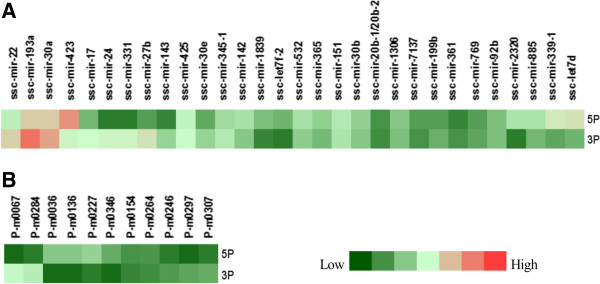
5p and 3p arm expression of 46 pre-miRNAs. (A) 30 known pre-miRNAs. (B) 11 novel pre-miRNAs (representing 15 coupled 3p and 5p arm sequences).
As described in other studies [51], some small RNAs were generated from the loop or the region between the loop and stem (ssc-let-7e, ssc-miR-27a and ssc-miR-30a) (Figure 10A–C). In addition, we detected another interesting type of small RNAs known as miRNA-offset RNAs, or “moRNAs”, which are derived from the ends of pre-miRNAs, predominantly from the 5’ end, independent of the mature miRNA. A good example of moRNAs and small RNAs generated from the loop was pre-miR-30a (Figure 10C). At the 5’ end of pre-miR-30, a 18 nt RNA sequence was found to be generated from the loop, downstream of ssc-miR-30a-5p (Figure 10C). The findings suggest that these RNAs may be only byproducts of Drosha and Dicer processing, or these small RNAs alternatively may take part in other important regulatory functions different from those of miRNAs.
Figure 10.
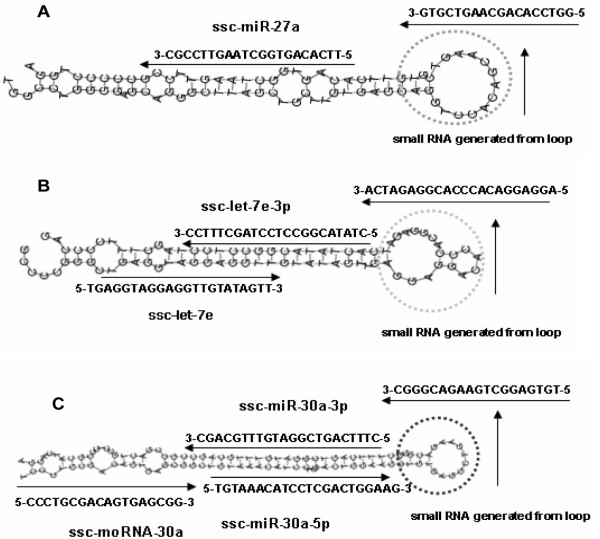
Three distinctive pre-miRNAs identified in porcine milk exosomes. (A) The ssc-miR-27a precursor produced a 3p-arm miRNA sequence and a loop-derived small RNA. (B) The ssc-let-7e precursor produced a 5p-arm sequence, a 3p-arm miRNA sequence and a loop-derived small RNA. (C) The ssc-miR-30a precursor produced a 5p-arm sequence, a 3p-arm miRNA sequence and a loop-derived small RNA. In addition, ssc-moRNA-3, belonging to new type of miRNA termed moRNA, was found at the 5’ end of pre-miR-30.
MiRNA clusters
According to criteria for classification in miRbase, pre-miRNAs located on a chromosome with an interval of less than 10 kb are defined as belonging to an miRNA cluster. In our analysis, 30 (including 11 novel and 19 known) (Figure 8) clusters were detected (Table 5). Among all miRNAs clusters, there were several pre-miRNAs with intervening sequences of less than 1 kb, including 10 known clusters (miR-99b/let-7e/125a, miR-24-2/27b/23b, miR-99a/let-7c, miR-29b/29a, miR-221/222, miR-98/let-7f, miR-181c/d, miR-363/92a/19b-2/106a, miR-363/92a/19b-2, miR-181b-1/181a-1 and miR-17/18a/19b-1/92a-1) and 4 novel miRNAs clusters (cluster 3, 9, 12, 22). We identified a typical polycistronic miRNA cluster, miR-363/92a/19b-2/20b, on chromosome X. Interestingly, the homologous cluster, miR-363/92a/19b-2/20b/106a on chromosome X, was located 33.5 kb downstream of miR-363/92a/19b-2/20b (Figure 11A, C), and a paraologous cluster miR-17/18a/19b-1/92a-1 was found on chromosome 11 (Figure 11B, C). The organization of miRNA precursors in the genome may account for variable levels of expression and regulation of mature miRNAs.
Table 5.
miRNAs cluster
| Cluster No. | Cluster name | members | chromosome |
|---|---|---|---|
| cluster 1 | new | p-m0165, p-m0166 | 1 |
| cluster 2 | mir-181a-2/181b-2 | ssc-mir-181a-2, ssc-mir-181b-2 | 1 |
| cluster 3 | new | ssc-miR-199b, P-m0179 | 1 |
| cluster 4 | miR-24-1/27a/23a | ssc-mir-24-1, ssc-miR-27a, ssc-mir-23a | 2 |
| cluster 5 | miR-181c/d | ssc-miR-181c, ssc-mir-181d | 2 |
| cluster 6 | miR-143/145 | ssc-mir-143, ssc-miR-145 | 2 |
| cluster 7 | let-7a/let-7f-2/let-7d | ssc-let-7a-2, P-m0204, ssc-let-7f-2, ssc-let-7d | 3 |
| cluster 8 | miR-30d/30b | ssc-mir-30d, ssc-mir-30b | 4 |
| cluster 9 | new | P-m0263, P-m0264 | 6 |
| cluster 10 | mir-99b/let-7e/125a | ssc-mir-99b, ssc-let-7e, ssc-mir-125a | 6 |
| cluster 11 | mir-30c-1/ mir-30e | ssc-mir-30c-1,ssc-mir-30e | 6 |
| Cluster 12 | new | P-m0302,P-m0318 | 7 |
| cluster 13 | let-7a-1/miR-100 | ssc-let-7a-1, ssc-mir-100 | 9 |
| cluster 14 | new | P-m0346, P-m0347, P-m0348,P-m0349 | 9 |
| cluster 15 | miR-181b-1/181a-1 | ssc-mir-181b-1, ssc-mir-181a-1 | 10 |
| cluster 16 | miR-24-2/27b/23b | ssc-mir-24-2, ssc-mir-27b, ssc-mir-23b | 10 |
| cluster 17 | miR-17-92a-1 | ssc-mir-17, ssc-mir-18a,ssc-mir-19b-1, ssc-mir-92a-1 | 11 |
| cluster 18 | new | P-m0024, P-m0025, P-m0026 | 12 |
| cluster 19 | miR-425/191 | ssc-mir-425, ssc-mir-191 | 13 |
| cluster 20 | miR-99a/let-7c | ssc-mir-99a, ssc-let-7c | 13 |
| cluster 21 | new | P-m0099, P-m0083 | 15 |
| cluster 22 | new | P-m0124, P-m0113 | 17 |
| cluster 23 | new | P-m0125, P-m0126, P-m0127, P-m0128, P-m0114 | 17 |
| cluster 24 | new | P-m0130, P-m0131 | 17 |
| cluster 25 | miR-29b-1/29a | ssc-mir-29b-1, ssc-mir-29a | 18 |
| cluster 26 | new | P-m0136, P-m0137, ssc-mir-183 | 18 |
| cluster 27 | miR-221/222 | ssc-mir-221, ssc-mir-222 | X |
| cluster 28 | miR-98/let-7f-1 | ssc-mir-98, ssc-let-7f-1 | X |
| cluster 29 | miR-363~20b | ssc-mir-363-1, ssc-mir-92a-2, ssc-mir-19b-2, P-m0370, sc-mir-20b-1/ssc-mir-20b-2 | X |
| Cluster 30 | miR-363~20b | ssc-mir-363-1, ssc-mir-92a-2, ssc-mir-19b-2, P-m0371, ssc-mir-20b-1/ssc-mir-20b-2, ssc-miR-106a | X |
Figure 11.
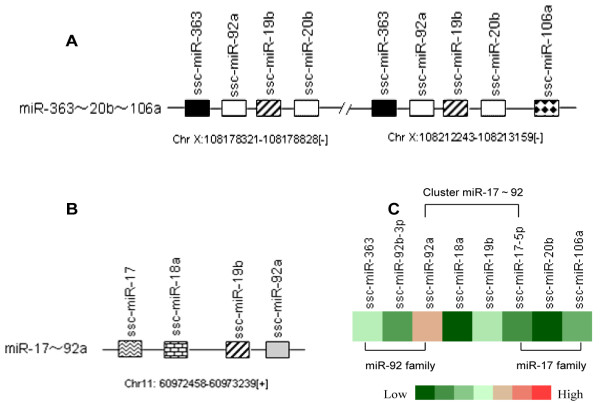
MiR-363~20b~106a homologous or paralogous cluster and expression level. (A) The miR-363/92a/19b/20b cluster and its homologous cluster miR-363/92a/19b/20b/106a on chromosome X were separated by a 33.5 kb DNA fragment. (B) The miR-17/18a/19/92a cluster was located on chromosome 11. In the genome, miR-92a/19b showed three copies; miR-363 and miR-20b had two copies; while miR-17, miR-18a and miR-106a had one copy. (C) Expression of mature miRNAs produced from the miR-363~20b~106a cluster and miR-17~92 cluster.
MiRNA families
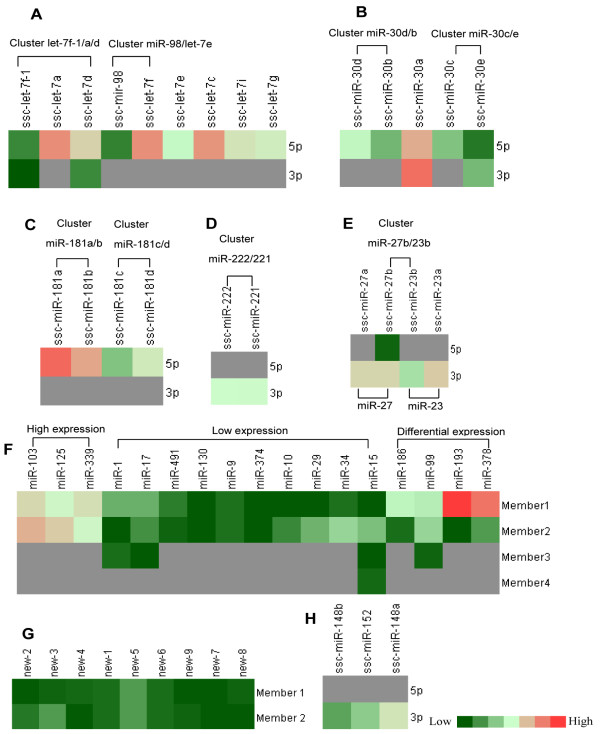
It is widely believed that the members of a given miRNA family regulate very similar sets of target genes. Apart from miRNA clusters, miRNA families were also recognized in the miRNAs of exosomes. Based on seed sequences, 35 miRNA families were identified (26 known and 9 novel miRNA families) to contain at least two members and the identification of novel miRNAs added new members to 5 known families (Table 6). In our study, 8 miRNA families (let-7, mir-1, mir-17, mir-181, mir-148, mir-30, mir-92 and mir-99) were found with at least 3 members among all exosome miRNAs. The let-7 family had 9 members, miR-181 family had 4 members (miR-181a/b/c/d) and miR-30 family had 5 members (miR-30a/b/c/d/e). Most importantly, these miRNAs were highly expressed, and let-7 family members (let-7a/f/c), miR-181a and miR-30a-3p were enriched among the top 10 miRNAs. However, members in the same family were highly differentially expressed. In the miR-181 family, miR-181a and miR-181b were dominant types with 13,345 reads and 3,333 reads, respectively. Similarly, miR-30a was the most abundant in the miR-30 family. The differential expression of members in the same family may be partly due to regulation of their precursors [52]. On the other hand, combined with the cluster analysis, we also observed that some miRNAs shared not only the same cluster but also the same families. These miRNAs included 181a/b, let-7f/miR-98, 181c/d, let-7a/f-5p/d-5p, 30b/d, 30c/e and miR-221/222. More interestingly, family members of the same cluster seemed to share expression patterns (Figure 12A–E). As mentioned above, miR-17-5p, miR-363, miR-106a, miR-18a, miR-19b, miR-92a, miR-20b and miR-92b formed a complex cluster and family network, and they also showed different expression patterns. MiR-92a, miR-19b and miR-363 were found to be highly expressed, while miR-17-5p, miR-18a, miR-20b and miR-106a were lowly expressed. The difference in abundance of the homologous or paralogous clusters may be attributed to the copy number of miRNA precursor itself or to the post-transcriptional regulation of the process of generating a mature miRNA from the precursor miRNA.
Table 6.
miRNAs family
| Family | Seed | Number | Members | |
|---|---|---|---|---|
| Family 1 | let-7 | GAGGTAG | 9 | ssc-let-7i, ssc-miR-98, ssc-let-7a,ssc-let-7f, ssc-let-7c, ssc-let-7 g, ssc-let-7e, ssc-let7d-5p |
| Family 2 | mir-1 | GGAATGT | 3 | ssc-miR-1, ssc-miR-206, PC-117 |
| Family 3 | mir-10 | ACCCTGT | 2 | ssc-miR-10b, ssc-miR-10a |
| Family 4 | mir-103 | GCAGCAT | 2 | ssc-miR-107, ssc-miR-103 |
| Family 5 | mir-125 | CCCTGAG | 2 | ssc-miR-125a, ssc-miR-125b |
| Family 6 | mir-130 | AGTGCAA | 2 | ssc-miR-130b, ssc-miR-130a |
| Family 7 | mir-148 | CAGTGCA | 3 | ssc-miR-148b, ssc-miR-152, ssc-miR-148a |
| Family 8 | mir-17 | AAAGTGC | 3 | ssc-miR-106a, ssc-miR-17-5p, ssc-mir-20b-1/ssc-mir-20b-2 |
| Family 9 | mir-181 | ACATTCA | 4 | ssc-miR-181a, ssc-miR-181b, ssc-miR-181c, ssc-miR-181d-5p |
| Family 10 | mir-186 | AAAGAAT | 2 | ssc-miR-186, PC-36 |
| Family 11 | mir-193 | ACTGGCC | 2 | ssc-miR-193a-3p, PC-3 |
| Family 12 | mir-221 | GCTACAT | 2 | ssc-miR-222, ssc-miR-221 |
| Family 13 | mir-23 | TCACATT | 2 | ssc-miR-23b, ssc-miR-23a |
| Family 14 | mir-27 | TCACAGT | 2 | ssc-miR-27a,ssc-miR-27b |
| Family 15 | mir-29 | AGCACCA | 2 | ssc-miR-29b, ssc-miR-29c |
| Family 16 | mir-30 | GTAAACA | 5 | ssc-miR-30e-5p, ssc-miR-30c, ssc-miR-30d, ssc-miR-30b-5p, ssc-miR-30a-5p |
| Family 17 | mir-30(#) | TTTCAGT | 2 | ssc-miR-30a-3p, ssc-miR-30e-3p |
| Family 18 | mir-339 | CCCTGTC | 2 | ssc-miR-339-5p, ssc-miR-4334-3p |
| Family 19 | mir-34 | GGCAGTG | 2 | ssc-miR-34c, ssc-miR-34a, |
| Family 20 | mir-363 | ATTGCAC | 3 | ssc-miR-363, ssc-miR-92b-3p, ssc-miR-92a |
| Family 21 | mir-374 | TATAATA | 2 | ssc-miR-374b-5p, ssc-miR-374a, |
| Family 22 | mir-378 | CTGGACT | 2 | ssc-miR-378, PS-2 |
| Family 23 | mir-491 | GTGGGGA | 2 | ssc-miR-491, PC-122 |
| Family 24 | mir-497 | AGCAGCA | 4 | ssc-miR-497, ssc-miR-15b, ssc-miR-16, ssc-miR-15a |
| Family 25 | mir-9 | CTTTGGT | 2 | ssc-miR-9-2, ssc-miR-9-1, |
| Family 26 | mir-99 | ACCCGTA | 3 | ssc-miR-99a, ssc-miR-99b, ssc-miR-100 |
| Family 27 | new-1 | CATGATT | 2 | P-m0040-3p(PC-9), P-m0240-5p(PC-232), |
| Family 28 | new-2 | AGAGGGA | 2 | P-m0048-3p(PC-152), P-m0064-3p(PC-217) |
| Family 29 | new-3 | ATTTGAT | 2 | P-m0084-5p(PC-228), P-m0105-3p(PC-72), |
| Family 30 | new-4 | GCTAGGA | 2 | P-m0110-5p(PC-112), P-m0139-5p(PC-111) |
| Family 31 | new-5 | TATGGAT | 2 | P-m0113-3p(PC-130), P-m0129-5p(PC-129) |
| Family 32 | new-6 | CCTGGAT | 2 | P-m0142-5p(PC-164), P-m0179-3p(PC-165) |
| Family 33 | new-7 | CATATTT | 2 | P-m0161-3p(PC-231), P-m0244-3p(PC-162) |
| Family 34 | new-8 | GTTTGGA | 2 | P-m0183-3p(PC-286), P-m0325-5p(PC-128) |
| Family 35 | new-9 | CTTTGGG | 2 | P-m0229-5p(PC-15), P-m0342-3p(PC-105) |
The underline indicated this family contains novel miRNAs. #: due to miRNAs classification by seed sequence, 3p and 5p of miR-30 represent different miRNAs families. PC is unique ID for porcine miRNAs candidate.
Figure 12.
Expression of miRNA families. (A) Expression of the let-7f family; let-7f-1, let-7a and let-7d formed a cluster, while miR-98 and let-7e formed a cluster. (B) Expression of miR-30 family; miR-30d clustered with miR-30b, and miR-30c clustered with miR-30e. (C) Expression of miR-181 family; miR-181a clustered with miR-181b, and miR-181c clustered with miR-181d. (D) MiR-222 and miR-221; miR-222 and miR-221 belonged to the same family and the same cluster. (E) MiR-27 family and miR-23 family; miR-27b and miR-23b formed a cluster in the genome. (F) Expression and clustering of remaining miRNA families; members 1, 2, 3 and 4 represent members of the indicated family. (G) Novel miRNA family with low expression. (H) Expression of the miR-148 family containing milk “marker” miRNAs.
In addition, many miRNA families showed low expression (count number <100) in milk exosomes, such as the miR-1, miR-130, miR-17, miR-10, miR-29, miR-374, mir-9, miR-15 and miR-491 families (Figure 12F), which are routinely expressed in specific tissues [53–56]. Interestingly, all 9 novel miRNAs families showed extremely low expression levels (Figure 12G, count number <50), indicating that these miRNAs may only be expressed in certain physiology processes and may be the reason for why these miRNAs have not been detected until now. MiR-148a was reported to be an important biomarker for milk exosome miRNAs [28, 57]. In this study, three members of this family (miR-148a, miR-148b and miR-152, Figure 12H) showed modest expression levels, suggesting that miR-148 may be a stably expressed miRNA in exosomes of most mammals including pigs.
GO and KEGG pathway analyses
To better understand the role of miRNAs in pig milk exosomes, potential targets of miRNAs at the pig genome level were explored by using our previous method [58]. We selected the top 10 miRNAs for target prediction using RNAhybrid, which found 2,333 potential transcripts. GO analysis showed that those targets were enriched in many processes (Table 7), including antigen processing and presentation, MHC class II protein complex and transcription. Pathways analysis revealed these targets were enriched in 7 KEGG pathways (Table 8), which are essential for piglet metabolism (glycerophospholipid metabolism and citrate cycle), immunity (cell adhesion molecules, asthma and intestinal immune network for IgA production, antigen processing and presentation) and development (Notch signaling pathway). The results suggest that these milk miRNAs likely take part in regulation of important protective functions in piglets, including the intestinal immune network for IgA production and antigen processing and presentation.
Table 7.
Gene ontology analysis of potential targets of top10 miRNAs
| Category | Term | Count | P-Value |
|---|---|---|---|
| Biological process | transcription | 15 | 6.70E-03 |
| Biological process | regulation of transcription | 21 | 1.00E-02 |
| Biological process | antigen processing and presentation | 7 | 1.50E-02 |
| Biological process | regulation of RNA metabolic process | 17 | 1.70E-02 |
| Biological process | regulation of transcription, DNA-dependent | 17 | 1.70E-02 |
| Biological process | antigen processing and presentation of peptide or polysaccharide antigen via MHC class II | 4 | 2.20E-02 |
| Cellular Component | MHC class II protein complex | 6 | 3.70E-04 |
| Cellular Component | MHC protein complex | 7 | 5.00E-03 |
| Cellular Component | large ribosomal subunit | 3 | 2.90E-02 |
| Molecular Function | transcription regulator activity | 20 | 1.80E-03 |
| Molecular Function | sequence-specific DNA binding | 13 | 2.60E-03 |
| Molecular Function | ligand-dependent nuclear receptor activity | 7 | 5.80E-03 |
| Molecular Function | transcription factor activity | 15 | 1.20E-02 |
| Molecular Function | steroid hormone receptor activity | 6 | 2.10E-02 |
| Molecular Function | DNA binding | 20 | 2.40E-02 |
| Molecular Function | phosphatase regulator activity | 4 | 2.90E-02 |
| Molecular Function | protein phosphatase regulator activity | 4 | 2.90E-02 |
Table 8.
KEGG pathway analysis of potential targets of top10 miRNAs
| Term | Count | gene | p-value |
|---|---|---|---|
| ssc04514: Cell adhesion molecules (CAMs) | 14 | CADM3, CD4, CD40, F11R, LOC100521555, SELE, SELL, SELP, SLA, SLA-DMA, SLA-DOA, SLA-DOB, SLA-DRA, SLA-DRB1 | 1.65E-03 |
| ssc05310: Asthma | 7 | CD40, SLA, SLA-DMA, SLA-DOA, SLA-DOB, SLA-DRA, SLA-DRB1 | 5.55E-03 |
| ssc04672: Intestinal immune network for IgA production | 8 | CD40, SLA, SLA-DMA, SLA-DOA, SLA-DOB, SLA-DRA, SLA-DRB1, TGFB3 | 2.15 E-02 |
| ssc04330: Notch signaling pathway | 5 | DLL4, PTCRA, LOC733643, APH1A, NOTCH4 | 2.33E-02 |
| ssc04612: Antigen processing and presentation | 10 | CD4, LOC100152370, NFYB, PSME1, SLA, SLA-DMA, SLA-DOA, SLA-DOB, SLA-DRA, SLA-DRB1 | 3.00E-02 |
| ssc00564:Glycerophospholipid metabolism | 6 | AGPAT1, AGPAT4, |AGPAT6, GNPAT, LOC100152491, PCYT1B | 4.25E-02 |
| ssc00020:Citrate cycle (TCA cycle) | 5 | ACO2,DLST, IDH2, LOC100157889, PCK1 | 4.57E-02 |
Targets of miRNAs in IgA immune network
As shown by GO and KEGG analysis, many of the identified miRNAs were predicted to participate in immunity, similar to findings of other studies [28]. To further understand those miRNAs, the top 20 predicted miRNAs were selected for in-depth analysis of the IgA immune network. The results showed that 14 of the top 20 miRNAs are likely involved in the IgA network and may target about 20 immune-related genes (APRIL, CCL25, CD40, CD80, CD86, ICOS, IL10, IL-5, ITGB7, LOC100621559, LTBR, MADCAM1, SLA, SLA-DMA, SLA-DMB, SLA-DOB, SLA-DQA1, SLA-DRA, SLA-DRB1 and TGFB3) (Figure 13). These results suggest that porcine milk miRNAs take part in regulation of the IgA network and immunity of piglets. In addition, several miRNAs shared the same target gene. Interestingly, all of the let-7 family members (let-7a, let-7c and let-7f) could target CCL25. Those miRNAs are proposed to play a key role in IgA production in the piglet digestive tract and deserve further exploration, as mucosal immunity is critically important for the protection of newborn piglets.
Figure 13.
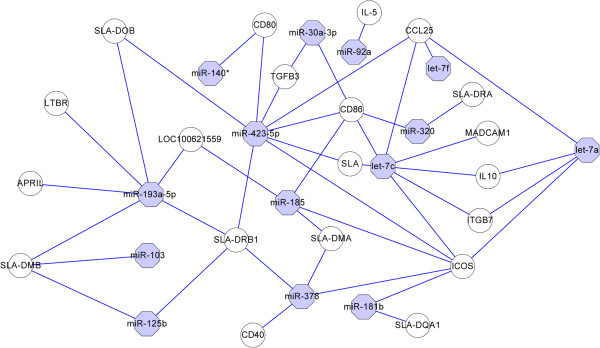
MiRNAs targeting the IgA immune network. The top 20 miRNAs were analyzed, and 14 of them were found to participate in the IgA immune network, involving 20 target genes. Different colors and shapes represent various relationships between miRNAs and genes.
Discussion
In the present study, a comprehensive miRNA expression profile of porcine breast milk exosomes was explored via a deep sequencing approach. We found in total 176 known miRNAs (miRBase 20.0) and 366 pre-miRNAs producing 315 mature miRNAs. Luciferase reporter assay was used to explore the targets of two predicted novel miRNAs in this study. Results indicated both of them down-regulated the luciferase expression by targeting 3’UTR of IGF-1R. All these pre-miRNAs were distributed in 30 clusters (11 novel and 19 known clusters), and the mature miRNAs could be assigned to 35 families (26 known and 9 unknown families). GO and KEGG pathway analyses show that those miRNAs may participate in many different immune-related processes. An analysis of the top 20 miRNAs showed that 14 of them may be involved in many regulatory aspects of the IgA immune network.
A recent study of exosome miRNAs in Yorkshire sow milk discovered 180 pre-miRNAs, including 140 known porcine pre-miRNAs and 40 novel pre-miRNAs, which encode 237 mature miRNAs (234 unique miRNAs) [29] In the current study, we discovered 205 known porcine pre-miRNAs (176 mature miRNAs) and 366 novel pre-miRNAs (315 mature miRNAs), approximately 254 more mature miRNAs than were revealed in the former report [29]. Therefore, our results substantially supplement the known pig miRNAs, particularly milk exosome miRNAs. Interestingly, most of the novel miRNAs were low in abundance (312 miRNAs with less than 100 reads and only 3 miRNAs with >100 reads), which is possibly the reason for why these miRNAs were not detected in a previous study by Gu et al.[29]. Further comparison revealed that miR-191 and let-7a, which potentially play a vital role in immunity, were found in both that study and in the top 10 miRNAs of our study. Other miRNAs identified previously (miR-30a-5p, miR-25-3p, miR-182-5p, miR-200c-3p and miR-375-3p) were not detected in our study. Furthermore, miR-148a, a potential biomarker for quality control in bovine milk [57] and human milk [28], which was found to be highly expressed throughout the lactation period of Yorkshire sows [29], was only detected at a moderate level in Landrace pigs in our study. MiR-148a has been reported to be a tumor metastasis suppressor in gastric cancer [59], and ectopic expression of miR-148a was shown to induce apoptosis and silence Bcl-2 in colorectal cancer cells [60]. By bioinformatics analysis, miR-148a was determined to be possibly related to immunity and gastrointestinal health, but the underlying regulatory mechanism remains unclear.
MiR-92a belongs to the miR-17 ~92 cluster with seven miRNAs (miR-17-5p, miR-17-3p, miR-18a, miR-19a, miR-19b, miR-20a and miR-92a) and was first described as an oncogenic miRNA cluster involved in B-cell lymphoma [61]. Recent studies indicated that the miRNA-17-92 (miR-17-92) cluster directly targets the TGFB pathway in cancer cell lines in the mouse embryo stage [62]. In addition, the miR-17-92 cluster also participates in normal development of the heart, lungs and immune system [63]. MiR-19 can promote leukemogenesis in Notch1-induced T-cell acute lymphoblastic leukemia (T-ALL) in vivo[64]. Overexpression of the mir-17–mir-18a–mir-19b-1 cluster was shown to accelerate Myc-induced tumor development in a mouse B-cell lymphoma model [61]. The combined results above imply that members of the cluster miR-363/92a/19b-2/20b/106a may be related to cell proliferation and development. In porcine milk, miR-363/92a/19b-2/20b (miR-363/92a/19b-2/20b/106a) and miR-17/18a/19b-1/92-1 were also detected. The miR-181 (181a/b/c/d) family is related to the development of different cells. It was reported that miR-181c/d can inhibit cell cycle and proliferation and that miR-181c regulates TNF-α[65]. The miR-30(b/c/d/e) family regulates kidney development by targeting the transcription factor Xlim1/Lhx1 in Xenopus[66]. The well-known let-7(a/b/d/f) family is involved in oncogene expression [67], and let-7/miR-98 family members are expressed late in mammalian embryonic development [68]. Thus, these miRNAs mentioned above may participate in development of the piglet digestive tract.
Notably, some miRNAs among the top 10 identified here have been reported to be related to immunity (miR-320, miR-181a, miR-30a-3p, let-7a, let-7f and let-7c) and development (miR-193a-3p, miR-378 and miR-191). MiR-193a-3p was demonstrated to regulate cell proliferation, cell cycle progression in vitro and in nude mice [69]. MiR-378 promotes osteoblast differentiation by targeting polypeptide N-acetylgalactosaminyltransferase 7 (GalNAc-T7 or GalNT7) [70], and miR-191 regulates erythroid differentiation in mammals by up-regulating erythroid-enriched genes Riok3 and Mxi1[71]. Meanwhile, miR-320 is able to inhibit HL-60 cell proliferation by suppressing receptor 1 (TfR-1; CD71) [72], and miR-181a was believed to act as an intrinsic antigen sensitivity “rheostat” during T cell development [73]. MiR-320, miR-181a, miR-30a-3p and let-7 were shown to be downregulated in colorectal cancer [74]. Of course, further experimental evidence is needed to verify that these miRNAs are indeed related to immunity of the piglet digestive tract.
IgA is a major immunoglobulin in milk [75]. Expression of the polymeric IgA receptor (pIgR) in mammary epithelial cells contribute much to the development of the immune system at the early stage of lactation [76]. In the present study, some miRNAs were predicted to target genes (CD40, SLA, SLA-DMA, SLA-DOA, SLA-DOB, SLA-DRA, SLA-DRB1 and TGFB3) involved in processes of the intestinal immune network for IgA production in porcine milk. CD40 is a B-cell antigen activated during immune responses [77]. CD40 and CD40 ligand (CD40L) expressed on activated T cells are essential to B cell proliferation [78] and secretion of IgG, IgA and IgE [79]. SLA Class I were found to be expressed in the epithelial and lamina propria cells of the intestine in adult pigs and to be involved in mother-newborn interactions [80]. A study in humans showed that TGF-β acts as a specific switch for IgA present at early stages of development of B cells [81].
In the present study, the top 20 miRNAs were used for IgA network analysis. APRIL was the predicted target of miR-193a-5p, which is essential to triggering IgA2 class switch in human B cells. Intestinal epithelial cells (IECs) release APRIL after sensing bacteria through Toll-like receptors, and mucosal vaccines activate IECs to induce more effective IgA2 responses [82]. The let7 family and miR-423-5p were predicted to target CCL25, a potent and selective chemoattractant for IgA antibody-secreting cells [83]. CCL25 is known to selectively modulate immune responses, specifically the localization of T lymphocytes to the small-intestinal mucosa [84]. CD80 and CD86, which are costimulators of T lymphocytes [85], were identified as possible targets of five miRNAs in our study. Let-7a, let-7c, miR-181b, miR-185, miR-378 and miR-423-5p were predicted to target the inducible co-stimulatory molecule (ICOS), which plays a key role in regulating T-cell differentiation, T-cell proliferation, and secretion of lymphokines, providing effective help for antibody secretion by B cells [86]. We hypothesize that some miRNAs identified here in porcine milk regulate IgA production in the intestine of piglets, which may play an important role in mucosa immunity. However, their regulatory mechanisms warrant further study.
Conclusions
In conclusion, the present study revealed 176 known miRNAs and 366 (315 mature miRNAs) novel pre-miRNAs in porcine milk, most of which were predicted to be involved in regulation of digestive tract development and immunity of newborn piglets. These findings contribute to an increased understanding of the roles of miRNAs in porcine (S. scrofa) milk exosomes and to building the foundation for understanding their physiological functions and regulatory mechanisms.
Availability of supporting data
All the supporting data has been deposited in https://mynotebook.labarchives.com/share/allinchen/MTkuNXwxMzMxMS8xNS0yL1RyZWVOb2RlLzE1NzEyODU2fDQ5LjU= with a DOI:10.6070/H4DN432G.
Electronic supplementary material
Additional file 1: Table S1: 176 known mature miRNAs.xls. (XLS 109 KB)
Additional file 2: Table S2: 315 novel miRNAs.xls. (XLS 196 KB)
Additional file 3: Table S3: Pre-miRNAs with 3p and 5p sequence.xls. (XLS 77 KB)
Acknowledgements
This work was supported by grants from the Key Project of Guangdong Provincial Nature Science Foundation (S2013020012766), National Basic Research Program of China (973 Program, 2011CB944200, 2009CB941600 and 2013CB127304), Natural Science Foundation of China program (31272529) and the Natural Science Foundation of Guangdong Province (S2013010013215). We thank the breeding farm of the Livestock Research Institute (Guangzhou, China) for providing milk samples.
Footnotes
Competing interests
We declared this manuscript have no competing interests.
Authors’ contributions
TC, QX carried out the miRNAs Solexa Sequencing and data analysis, and participated in drafted the manuscript. XC, QQ carried out the PCR, qPCR. GS, SW participated in the sample collected. XZ, NW performed the Transmission electron microscopy. RY, QJ YZ conceived of the study, and participated in its design and coordination and helped to draft the manuscript. All authors read and approved the final manuscript.
Contributor Information
Ting Chen, Email: 36391538@qq.com.
Qian-Yun Xi, Email: Xqy0228@163.com.
Rui-Song Ye, Email: yiguangnian2004@163.com.
Xiao Cheng, Email: 21739942@qq.com.
Qi-En Qi, Email: qiqien1987@163.com.
Song-Bo Wang, Email: songbowang@gmail.com.
Gang Shu, Email: shugang@scau.edu.cn.
Li-Na Wang, Email: wanglinatracy@163.com.
Xiao-Tong Zhu, Email: xtzhu@scau.edu.cn.
Qing-Yan Jiang, Email: qyjiang@scau.edu.cn.
Yong-Liang Zhang, Email: Zhangyl@scau.edu.cn.
References
- 1.Strobel S. Immunity induced after a feed of antigen during early life: oral tolerance v. sensitisation. Proc Nutr Soc. 2001;60(4):437–442. doi: 10.1079/PNS2001119. [DOI] [PubMed] [Google Scholar]
- 2.Armogida SA, Yannaras NM, Melton AL, Srivastava MD. Identification and quantification of innate immune system mediators in human breast milk. Allergy Asthma Proc. 2004;25(5):297–304. [PubMed] [Google Scholar]
- 3.Kramer MS, Chalmers B, Hodnett ED, Sevkovskaya Z, Dzikovich I, Shapiro S, Collet JP, Vanilovich I, Mezen I, Ducruet T. Promotion of breastfeeding intervention trial (PROBIT) JAMA. 2001;285(4):413–420. doi: 10.1001/jama.285.4.413. [DOI] [PubMed] [Google Scholar]
- 4.Høst A, Koletzko B, Dreborg S, Muraro A, Wahn U, Aggett P, Bresson J, Hernell O, Lafeber H, Michaelsen K. Dietary products used in infants for treatment and prevention of food allergy. Arch Dis Child. 1999;81(1):80–84. doi: 10.1136/adc.81.1.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Van Niel G, Porto-Carreiro I, Simoes S, Raposo G. Exosomes: a common pathway for a specialized function. J Biochem. 2006;140(1):13–21. doi: 10.1093/jb/mvj128. [DOI] [PubMed] [Google Scholar]
- 6.Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33(3):967–978. doi: 10.1016/0092-8674(83)90040-5. [DOI] [PubMed] [Google Scholar]
- 7.Théry C, Regnault A, Garin J, Wolfers J, Zitvogel L, Ricciardi-Castagnoli P, Raposo G, Amigorena S. Molecular characterization of dendritic cell-derived exosomes. J Cell Biol. 1999;147(3):599–610. doi: 10.1083/jcb.147.3.599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Raposo G, Nijman HW, Stoorvogel W, Liejendekker R, Harding CV, Melief C, Geuze HJ. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–1172. doi: 10.1084/jem.183.3.1161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Blanchard N, Lankar D, Faure F, Regnault A, Dumont C, Raposo G, Hivroz C. TCR activation of human T cells induces the production of exosomes bearing the TCR/CD3/ζ complex. J Immunol. 2002;168(7):3235–3241. doi: 10.4049/jimmunol.168.7.3235. [DOI] [PubMed] [Google Scholar]
- 10.Raposo G, Tenza D, Mecheri S, Peronet R, Bonnerot C, Desaymard C. Accumulation of major histocompatibility complex class II molecules in mast cell secretory granules and their release upon degranulation. Mol Biol Cell. 1997;8(12):2631–2645. doi: 10.1091/mbc.8.12.2631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Van Niel G, Raposo G, Candalh C, Boussac M, Hershberg R, Cerf-Bensussan N, Heyman M. Intestinal epithelial cells secrete exosome-like vesicles. Gastroenterology. 2001;121(2):337–349. doi: 10.1053/gast.2001.26263. [DOI] [PubMed] [Google Scholar]
- 12.Mears R, Craven RA, Hanrahan S, Totty N, Upton C, Young SL, Patel P, Selby PJ, Banks RE. Proteomic analysis of melanoma‒derived exosomes by two‒dimensional polyacrylamide gel electrophoresis and mass spectrometry. Proteomics. 2004;4(12):4019–4031. doi: 10.1002/pmic.200400876. [DOI] [PubMed] [Google Scholar]
- 13.Gonzalez-Begne M, Lu B, Han X, Hagen FK, Hand AR, Melvin JE, Yates JR., III Proteomic analysis of human parotid gland exosomes by multidimensional protein identification technology (MudPIT) J Proteome Res. 2009;8(3):1304–1314. doi: 10.1021/pr800658c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ogawa Y, Kanai-Azuma M, Akimoto Y, Kawakami H, Yanoshita R. Exosome-like vesicles with dipeptidyl peptidase IV in human saliva. Biol Pharm Bull. 2008;31(6):1059–1062. doi: 10.1248/bpb.31.1059. [DOI] [PubMed] [Google Scholar]
- 15.García JM, García V, Peña C, Domínguez G, Silva J, Diaz R, Espinosa P, Citores MJ, Collado M, Bonilla F. Extracellular plasma RNA from colon cancer patients is confined in a vesicle-like structure and is mRNA-enriched. RNA. 2008;14(7):1424–1432. doi: 10.1261/rna.755908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Pisitkun T, Shen RF, Knepper MA. Identification and proteomic profiling of exosomes in human urine. Proc Natl Acad Sci USA. 2004;101(36):13368. doi: 10.1073/pnas.0403453101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Keller S, Rupp C, Stoeck A, Runz S, Fogel M, Lugert S, Hager H, Abdel-Bakky M, Gutwein P, Altevogt P. CD24 is a marker of exosomes secreted into urine and amniotic fluid. Kidney Int. 2007;72(9):1095–1102. doi: 10.1038/sj.ki.5002486. [DOI] [PubMed] [Google Scholar]
- 18.Runz S, Keller S, Rupp C, Stoeck A, Issa Y, Koensgen D, Mustea A, Sehouli J, Kristiansen G, Altevogt P. Malignant ascites-derived exosomes of ovarian carcinoma patients contain CD24 and EpCAM. Gynecol Oncol. 2007;107(3):563–571. doi: 10.1016/j.ygyno.2007.08.064. [DOI] [PubMed] [Google Scholar]
- 19.Prado N, Marazuela EG, Segura E, Fernández-García H, Villalba M, Théry C, Rodríguez R, Batanero E. Exosomes from bronchoalveolar fluid of tolerized mice prevent allergic reaction. J Immunol. 2008;181(2):1519–1525. doi: 10.4049/jimmunol.181.2.1519. [DOI] [PubMed] [Google Scholar]
- 20.Simpson RJ, Jensen SS, Lim JWE. Proteomic profiling of exosomes: current perspectives. Proteomics. 2008;8(19):4083–4099. doi: 10.1002/pmic.200800109. [DOI] [PubMed] [Google Scholar]
- 21.Deregibus MC, Cantaluppi V, Calogero R, Iacono ML, Tetta C, Biancone L, Bruno S, Bussolati B, Camussi G. Endothelial progenitor cell–derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood. 2007;110(7):2440–2448. doi: 10.1182/blood-2007-03-078709. [DOI] [PubMed] [Google Scholar]
- 22.Lakkaraju A, Rodriguez-Boulan E. Itinerant exosomes: emerging roles in cell and tissue polarity. Trends Cell Biol. 2008;18(5):199–209. doi: 10.1016/j.tcb.2008.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schorey JS, Bhatnagar S. Exosome function: from tumor immunology to pathogen biology. Traffic. 2008;9(6):871–881. doi: 10.1111/j.1600-0854.2008.00734.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–659. doi: 10.1038/ncb1596. [DOI] [PubMed] [Google Scholar]
- 25.Nakatani H, Aoki N, Nakagawa Y, Jin-No S, Aoyama K, Oshima K, Ohira S, Sato C, Nadano D, Matsuda T. Weaning-induced expression of a milk-fat globule protein, MFG-E8, in mouse mammary glands, as demonstrated by the analyses of its mRNA, protein and phosphatidylserine-binding activity. Biochem J. 2006;395(Pt 1):21. doi: 10.1042/BJ20051459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Admyre C, Johansson SM, Qazi KR, Filén JJ, Lahesmaa R, Norman M, Neve EPA, Scheynius A, Gabrielsson S. Exosomes with immune modulatory features are present in human breast milk. J Immunol. 2007;179(3):1969–1978. doi: 10.4049/jimmunol.179.3.1969. [DOI] [PubMed] [Google Scholar]
- 27.Hata T, Murakami K, Nakatani H, Yamamoto Y, Matsuda T, Aoki N. Isolation of bovine milk-derived microvesicles carrying mRNAs and microRNAs. Biochem Biophys Res Commun. 2010;396(2):528–533. doi: 10.1016/j.bbrc.2010.04.135. [DOI] [PubMed] [Google Scholar]
- 28.Zhou Q, Li M, Wang X, Li Q, Wang T, Zhu Q, Zhou X, Gao X, Li X. Immune-related microRNAs are abundant in breast milk exosomes. Int J Biol Sci. 2012;8(1):118. doi: 10.7150/ijbs.8.118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Gu Y, Li M, Wang T, Liang Y, Zhong Z, Wang X, Zhou Q, Chen L, Lang Q, He Z, et al. Lactation-related microRNA expression profiles of porcine breast milk exosomes. PLoS One. 2012;7(8):e43691. doi: 10.1371/journal.pone.0043691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cullen BR. RNA interference: antiviral defense and genetic tool. Nat Immunol. 2002;3(7):597–599. doi: 10.1038/ni0702-597. [DOI] [PubMed] [Google Scholar]
- 31.Hutvágner G, Zamore PD. A microRNA in a multiple-turnover RNAi enzyme complex. Science. 2002;297(5589):2056–2060. doi: 10.1126/science.1073827. [DOI] [PubMed] [Google Scholar]
- 32.Carrington JC, Ambros V. Role of microRNAs in plant and animal development. Science. 2003;301(5631):336–338. doi: 10.1126/science.1085242. [DOI] [PubMed] [Google Scholar]
- 33.Cerutti H. RNA interference: traveling in the cell and gaining functions? Trends Genet. 2003;19(1):39–46. doi: 10.1016/S0168-9525(02)00010-0. [DOI] [PubMed] [Google Scholar]
- 34.Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi: 10.1016/j.cell.2009.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Friedman RC, Farh KKH, Burge CB, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009;19(1):92–105. doi: 10.1101/gr.082701.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rodriguez A, Vigorito E, Clare S, Warren MV, Couttet P, Soond DR, van Dongen S, Grocock RJ, Das PP, Miska EA. Requirement of bic/microRNA-155 for normal immune function. Science. 2007;316(5824):608–611. doi: 10.1126/science.1139253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hyun S, Lee JH, Jin H, Nam J, Namkoong B, Lee G, Chung J, Kim VN. Conserved MicroRNA miR-8/miR-200 and its target USH/FOG2 control growth by regulating PI3K. Cell. 2009;139(6):1096–1108. doi: 10.1016/j.cell.2009.11.020. [DOI] [PubMed] [Google Scholar]
- 38.Lal A, Navarro F, Maher CA, Maliszewski LE, Yan N, O’Day E, Chowdhury D, Dykxhoorn DM, Tsai P, Hofmann O. miR-24 Inhibits cell proliferation by targeting E2F2, MYC, and other cell-cycle genes via binding to “seedless” 3’ UTR microRNA recognition elements. Mol Cell. 2009;35(5):610–625. doi: 10.1016/j.molcel.2009.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen CZ, Li L, Lodish HF, Bartel DP. MicroRNAs modulate hematopoietic lineage differentiation. Science. 2004;303(5654):83–86. doi: 10.1126/science.1091903. [DOI] [PubMed] [Google Scholar]
- 40.Lässer C, Alikhani VS, Ekström K, Eldh M, Paredes PT, Bossios A, Sjöstrand M, Gabrielsson S, Lötvall J, Valadi H. Human saliva, plasma and breast milk exosomes contain RNA: uptake by macrophages. J Transl Med. 2011;9(1):9. doi: 10.1186/1479-5876-9-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ji Z, Wang G, Xie Z, Wang J, Zhang C, Dong F, Chen C. Identification of novel and differentially expressed microRNAs of dairy goat mammary gland tissues using Solexa sequencing and bioinformatics. PLoS ONE. 2012;7(11):e49463. doi: 10.1371/journal.pone.0049463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Li R, Li Y, Kristiansen K, Wang J. SOAP: short oligonucleotide alignment program. Bioinformatics. 2008;24(5):713–714. doi: 10.1093/bioinformatics/btn025. [DOI] [PubMed] [Google Scholar]
- 43.Li Y, Zhang Z, Liu F, Vongsangnak W, Jing Q, Shen B. Performance comparison and evaluation of software tools for microRNA deep-sequencing data analysis. Nucleic Acids Res. 2012;40(10):4298–4305. doi: 10.1093/nar/gks043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jiang P, Wu H, Wang W, Ma W, Sun X, Lu Z. MiPred: classification of real and pseudo microRNA precursors using random forest prediction model with combined features. Nucleic Acids Res. 2007;35(suppl 2):W339–W344. doi: 10.1093/nar/gkm368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zuker M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003;31(13):3406–3415. doi: 10.1093/nar/gkg595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fu H, Tie Y, Xu C, Zhang Z, Zhu J, Shi Y, Jiang H, Sun Z, Zheng X. Identification of human fetal liver miRNAs by a novel method. FEBS Lett. 2005;579(17):3849–3854. doi: 10.1016/j.febslet.2005.05.064. [DOI] [PubMed] [Google Scholar]
- 47.Kosaka N, Izumi H, Sekine K, Ochiya T. microRNA as a new immune-regulatory agent in breast milk. Silence. 2010;1(1):7. doi: 10.1186/1758-907X-1-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ebhardt HA, Fedynak A, Fahlman RP. Naturally occurring variations in sequence length creates microRNA isoforms that differ in argonaute effector complex specificity. Silence. 2010;1(1):12. doi: 10.1186/1758-907X-1-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guo L, Lu Z. Global expression analysis of miRNA gene cluster and family based on isomiRs from deep sequencing data. Comput Biol Chem. 2010;34(3):165–171. doi: 10.1016/j.compbiolchem.2010.06.001. [DOI] [PubMed] [Google Scholar]
- 50.Naya L, Khan GA, Sorin C, Hartmann C, Crespi M, Lelandais-Brière C. Cleavage of a non-conserved target by a specific miR156 isoform in root apexes of Medicago truncatula. Plant Signal Behav. 2010;5(3):328–331. doi: 10.4161/psb.5.3.11190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Li M, Xia Y, Gu Y, Zhang K, Lang Q, Chen L, Guan J, Luo Z, Chen H, Li Y. MicroRNAome of porcine pre-and postnatal development. PLoS One. 2010;5(7):e11541. doi: 10.1371/journal.pone.0011541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet. 2004;5(7):522–531. doi: 10.1038/nrg1379. [DOI] [PubMed] [Google Scholar]
- 53.Lagos-Quintana M, Rauhut R, Yalcin A, Meyer J, Lendeckel W, Tuschl T. Identification of tissue-specific microRNAs from mouse. Curr Biol. 2002;12(9):735–739. doi: 10.1016/S0960-9822(02)00809-6. [DOI] [PubMed] [Google Scholar]
- 54.Tran N, O’Brien CJ, Clark J, Rose B. Potential role of micro‒RNAs in head and neck tumorigenesis. Head Neck. 2010;32(8):1099–1111. doi: 10.1002/hed.21356. [DOI] [PubMed] [Google Scholar]
- 55.Morales Prieto DM, Markert UR. MicroRNAs in pregnancy. J Reprod Immunol. 2011;88(2):106–111. doi: 10.1016/j.jri.2011.01.004. [DOI] [PubMed] [Google Scholar]
- 56.Xie S, Huang T, Shen Y, Li X, Zhang X, Zhu M, Qin H, Zhao S. Identification and characterization of microRNAs from porcine skeletal muscle. Anim Genet. 2010;41(2):179–190. doi: 10.1111/j.1365-2052.2009.01991.x. [DOI] [PubMed] [Google Scholar]
- 57.Chen X, Gao C, Li H, Huang L, Sun Q, Dong Y, Tian C, Gao S, Dong H, Guan D. Identification and characterization of microRNAs in raw milk during different periods of lactation, commercial fluid, and powdered milk products. Cell Res. 2010;20(10):1128–1137. doi: 10.1038/cr.2010.80. [DOI] [PubMed] [Google Scholar]
- 58.Ye R-S, Xi Q-Y, Qi Q, Cheng X, Chen T, Li H, Kallon S, Shu G, Wang S-B, Jiang Q-Y. Differentially Expressed miRNAs after GnRH Treatment and Their Potential Roles in FSH Regulation in Porcine Anterior Pituitary Cell. PLoS ONE. 2013;8(2):e57156. doi: 10.1371/journal.pone.0057156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zheng B, Liang L, Wang C, Huang S, Cao X, Zha R, Liu L, Jia D, Tian Q, Wu J. MicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in gastric cancer. Clin Cancer Res. 2011;17(24):7574–7583. doi: 10.1158/1078-0432.CCR-11-1714. [DOI] [PubMed] [Google Scholar]
- 60.Zhang H, Li Y, Huang Q, Ren X, Hu H, Sheng H, Lai M. MiR-148a promotes apoptosis by targeting Bcl-2 in colorectal cancer. Cell Death Differ. 2011;18(11):1702–1710. doi: 10.1038/cdd.2011.28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.He L, Thomson JM, Hemann MT, Hernando-Monge E, Mu D, Goodson S, Powers S, Cordon-Cardo C, Lowe SW, Hannon GJ. A microRNA polycistron as a potential human oncogene. Nature. 2005;435(7043):828–833. doi: 10.1038/nature03552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Li L, Shi JY, Zhu GQ, Shi B. MiR‒17‒92 cluster regulates cell proliferation and collagen synthesis by targeting TGFB pathway in mouse palatal mesenchymal cells. J Cell Biochem. 2012;113(4):1235–1244. doi: 10.1002/jcb.23457. [DOI] [PubMed] [Google Scholar]
- 63.Mendell JT. miRiad roles for the miR-17-92 cluster in development and disease. Cell. 2008;133(2):217–222. doi: 10.1016/j.cell.2008.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mavrakis KJ, Wolfe AL, Oricchio E, Palomero T, De Keersmaecker K, McJunkin K, Zuber J, James T, Khan AA, Leslie CS. Genome-wide RNA-mediated interference screen identifies miR-19 targets in Notch-induced T-cell acute lymphoblastic leukaemia. Nat Cell Biol. 2010;12(4):372–379. doi: 10.1038/ncb2037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Río P, Agirre X, Garate L, Baños R, Álvarez L, San José-Enériz E, Badell I, Casado JA, Garín M, Prósper F. Down-regulated expression of hsa-miR-181c in Fanconi anemia patients: implications in TNFα regulation and proliferation of hematopoietic progenitor cells. Blood. 2012;119(13):3042–3049. doi: 10.1182/blood-2011-01-331017. [DOI] [PubMed] [Google Scholar]
- 66.Agrawal R, Tran U, Wessely O. The miR-30 miRNA family regulates Xenopus pronephros development and targets the transcription factor Xlim1/Lhx1. Development. 2009;136(23):3927–3936. doi: 10.1242/dev.037432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Akao Y, Nakagawa Y, Naoe T. let-7 microRNA functions as a potential growth suppressor in human colon cancer cells. Biol Pharm Bull. 2006;29(5):903–906. doi: 10.1248/bpb.29.903. [DOI] [PubMed] [Google Scholar]
- 68.Shell S, Park SM, Radjabi AR, Schickel R, Kistner EO, Jewell DA, Feig C, Lengyel E, Peter ME. Let-7 expression defines two differentiation stages of cancer. Proc Natl Acad Sci. 2007;104(27):11400–11405. doi: 10.1073/pnas.0704372104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ma K, He Y, Zhang H, Fei Q, Niu D, Wang D, Ding X, Xu H, Chen X, Zhu J. DNA methylation-regulated miR-193a-3p dictates resistance of hepatocellular carcinoma to 5-fluorouracil via repression of SRSF2 expression. J Biol Chem. 2012;287(8):5639–5649. doi: 10.1074/jbc.M111.291229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kahai S, Lee SC, Lee DY, Yang J, Li M, Wang CH, Jiang Z, Zhang Y, Peng C, Yang BB. MicroRNA miR-378 regulates nephronectin expression modulating osteoblast differentiation by targeting GalNT-7. PLoS One. 2009;4(10):e7535. doi: 10.1371/journal.pone.0007535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Zhang L, Flygare J, Wong P, Lim B, Lodish HF. miR-191 regulates mouse erythroblast enucleation by down-regulating Riok3 and Mxi1. Genes Dev. 2011;25(2):119–124. doi: 10.1101/gad.1998711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Schaar DG, Medina DJ, Moore DF, Strair RK, Ting Y. miR-320 targets transferrin receptor 1 (CD71) and inhibits cell proliferation. Exp Hematol. 2009;37(2):245–255. doi: 10.1016/j.exphem.2008.10.002. [DOI] [PubMed] [Google Scholar]
- 73.Li QJ, Chau J, Ebert PJR, Sylvester G, Min H, Liu G, Braich R, Manoharan M, Soutschek J, Skare P. miR-181a is an intrinsic modulator of T cell sensitivity and selection. Cell. 2007;129(1):147–161. doi: 10.1016/j.cell.2007.03.008. [DOI] [PubMed] [Google Scholar]
- 74.Aslam MI, Patel M, Singh B, Jameson JS, Pringle JH. MicroRNAs are Novel Biomarkers for Detection of Colorectal Cancer. In: Khan TK, editor. Biomarker. Croatia, Rijeka: In Tech; 2012. pp. 1–18. [Google Scholar]
- 75.Curtis J, Bourne F. Immunoglobulin quantitation in sow serum, colostrum and milk and the serum of young pigs. Biochimica et Biophysica Acta (BBA)-Protein Structure. 1971;236(1):319–332. doi: 10.1016/0005-2795(71)90181-4. [DOI] [PubMed] [Google Scholar]
- 76.Kumura H, Sone T, Shimazaki K, Kobayashi E. Sequence analysis of porcine polymeric immunoglobulin receptor from mammary epithelial cells present in colostrum. J Dairy Res. 2000;67(04):631–636. doi: 10.1017/S0022029900004404. [DOI] [PubMed] [Google Scholar]
- 77.Castigli E, Alt FW, Davidson L, Bottaro A, Mizoguchi E, Bhan AK, Geha RS. CD40-deficient mice generated by recombination-activating gene-2-deficient blastocyst complementation. Proc Natl Acad Sci. 1994;91(25):12135–12139. doi: 10.1073/pnas.91.25.12135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kawabe T, Naka T, Yoshida K, Tanaka T, Fujiwara H, Suematsu S, Yoshida N, Kishimoto T, Kikutani H. The immune responses in CD40-deficient mice: impaired immunoglobulin class switching and germinal center formation. Immunity. 1994;1(3):167. doi: 10.1016/1074-7613(94)90095-7. [DOI] [PubMed] [Google Scholar]
- 79.Durandy A, Schiff C, Bonnefoy JY, Forveille M, Rousset F, Mazzei G, Milili M, Fischer A. Induction by anti‒CD40 antibody or soluble CD40 ligand and cytokines of IgG, IgA and IgE production by B cells from patients with X‒linked hyper IgM syndrome. Eur J Immunol. 2005;23(9):2294–2299. doi: 10.1002/eji.1830230936. [DOI] [PubMed] [Google Scholar]
- 80.Le Jan C, Chevaleyre C. Reduced expression of SLA Class 1 antigens by intestinal epithelium of newborn piglets. Vet Immunol Immunopathol. 1996;50(1):167–172. doi: 10.1016/0165-2427(95)05486-3. [DOI] [PubMed] [Google Scholar]
- 81.Van Vlasselaer P, Punnonen J, De Vries J. Transforming growth factor-beta directs IgA switching in human B cells. J Immunol. 1992;148(7):2062–2067. [PubMed] [Google Scholar]
- 82.He B, Xu W, Santini PA, Polydorides AD, Chiu A, Estrella J, Shan M, Chadburn A, Villanacci V, Plebani A. Intestinal Bacteria Trigger T Cell-Independent Immunoglobulin A < sub > 2</sub > Class Switching by Inducing Epithelial-Cell Secretion of the Cytokine APRIL. Immunity. 2007;26(6):812–826. doi: 10.1016/j.immuni.2007.04.014. [DOI] [PubMed] [Google Scholar]
- 83.Bowman EP, Kuklin NA, Youngman KR, Lazarus NH, Kunkel EJ, Pan J, Greenberg HB, Butcher EC. The intestinal chemokine thymus-expressed chemokine (CCL25) attracts IgA antibody-secreting cells. J Exp Med. 2002;195(2):269–275. doi: 10.1084/jem.20010670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Svensson M, Marsal J, Ericsson A, Carramolino L, Brodén T, Márquez G, Agace WW. CCL25 mediates the localization of recently activated CD8alphabeta^+ lymphocytes to the small-intestinal mucosa. J Clin Investig. 2002;110(8):1113–1122. doi: 10.1172/JCI0215988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Lanier LL, O’Fallon S, Somoza C, Phillips JH, Linsley PS, Okumura K, Ito D, Azuma M. CD80 (B7) and CD86 (B70) provide similar costimulatory signals for T cell proliferation, cytokine production, and generation of CTL. J Immunol. 1995;154(1):97–105. [PubMed] [Google Scholar]
- 86.Hutloff A, Dittrich AM, Beier KC, Eljaschewitsch B, Kraft R, Anagnostopoulos I, Kroczek RA. ICOS is an inducible T-cell co-stimulator structurally and functionally related to CD28. Nature. 1999;402:21–24. doi: 10.1038/46909. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1: Table S1: 176 known mature miRNAs.xls. (XLS 109 KB)
Additional file 2: Table S2: 315 novel miRNAs.xls. (XLS 196 KB)
Additional file 3: Table S3: Pre-miRNAs with 3p and 5p sequence.xls. (XLS 77 KB)