

Abstract
Background
Stress alters the oxidant-antioxidant state and immune cell responses which disrupts its function to combat infection. Blastocystis hominis, a common intestinal protozoan has been reported to be opportunistic in immunocompromised patients namely cancer. B. hominis infectivity in other altered immune system conditions especially stress is unknown. We aimed to demonstrate the stress effects towards the susceptibility and pathogenicity of B. hominis infection.
Methods/Findings
Three-week-old Wistar rats were divided into four groups: (a)control; (b)stress-induced; (c)B. hominis infected; (d)stress-induced with B. hominis infection; (n = 20 respectively). Stress was induced for an hour daily (30 days) using a Belly Dancer Shaker. Weight gain was monitored, stool samples were collected for B. hominis screening and blood for the determination of differential count, levels of immunoglobulin, oxidative damage, and peripheral blood mononuclear cell (PBMC) proliferation upon induction with solubilized antigen of B. hominis (Blasto-Ag). Group (b) exhibited the highest level of weight gain. Group (d) had higher levels of parasite cyst count in stools, serum IgE, oxidized protein and lipid compared to the group (c). Levels of monocyte and antioxidant in group (d) were decreased and their PBMCs showed highest inhibition of proliferation level when exposed to Blasto-Ag. Monocyte level in Group (b) showed insignificant difference compared to group (a) but was significantly lower compared to group (c). Antioxidant levels in group (c) were generally lower compared to group (a) and (b). Inhibition level exhibited by Blasto-Ag treated PBMCs of group (c) was higher compared to group (a) and (b).
Conclusion
The pathogenicity and augmentation of B. hominis infection is enhanced when stress is present. Lifestyles today are becoming increasingly stressed and the present findings suggest that the parasite which has been reported to be one of the most common organisms seen in stool surveys, namely in developing countries, may tend to be more pathogenic in stressful situations.
Introduction
In today's world, humans are often impacted with stress in the course of pursuing success in carrier, wealth and survival. In United States of America, reports have shown that in the year 2009 stress increased 24% and 18% for both man and women respectively as compared to the year 1983 [1]. Stress is defined as a type of condition or response in a living being that is caused by various types of internal or external stimulus [2]. Stress is known to cause behavioral and psychological changes that can lead to disturbance in the body's physiological function including imbalance in the oxidant-antioxidant state [3]. The overproduction of oxidants or free radicals such as superoxides (O2 •) and hydroxyls (OH•) compared to the antioxidants will result in a state called oxidative stress. Oxidative stress is known to cause oxidation of lipid, protein and DNA of cells [4] which results in the abnormal function of tissues and organs of the body. A considerable number of studies done on humans and animals have gathered evidences to associate stress with the etiology of various psychotic and metabolic diseases such as depression, gastritis, rheumatoid arthritis, and cancer [5]–[7].
Previous studies done on rodents have also demonstrated that chronic and acute stress using vibration and restrain stressors can impair immune responses by altering the activities of peripheral blood mononuclear cells (PBMCs) such as lymphocytes, neutrophils and monocytes which will culminate in abnormal antibody productions [7], [8]. Disruption in the PBMCs function may affect its role in combating invading antigens or infections including intestinal parasitic infections. Generally, when a host's immune system is triggered by parasitic infection, a massive production of oxidants are activated by immune cells including PBMCs to eradicate the infection [9]. In long term infections, the continuous release of reactive species together with a lack of antioxidant production will result in oxidative damage, exposing the host to other illnesses.
Intestinal parasites such as Ascaris lumbricoides and Cryptosporidium parvum have been reported to cause alterations in the molecular functions of the hosts' PBMCs and immunoglobulin levels [10], [11]. In addition, antigens originating from intestinal parasites namely Ascaris lumbricoides and Trichuris trichiura have been shown to alter in vitro proliferation of PBMCs isolated from normal and parasite infected individuals [12]. The presence of oxidative stress and compromised antioxidant defence mechanisms in humans and experimental models infected with intestinal protozoans such as Entamoeba histolytica and Giardia sp. have also been reported previously [13]. However, to the best of our knowledge, there have been no reports on the role of stress in contributing to host's immunosuppression, oxidative damage, and susceptibility to intestinal parasitic infections.
Blastocystis hominis, is one of the most common intestinal protozoan parasites found in humans. It is known to show diverse morphologies and reproductive processes [14]. The prevalence of B. hominis ranges from approximately 10% in developed countries and up to 60% in developing countries [15]. The extreme dispute regarding its pathogenicity had led to the remarkable findings on both phenotypic and genotypic characteristics of asymptomatic and symptomatic human-derived B. hominis isolates [16]. Various reports have shown that certain subtype of this unicellular protozoan is coupled with intestinal disorders including Irritable Bowel Syndrome (IBS) [17]. Previously, we have reported on the elevation of oxidative damage and proinflammatory cytokines caused by B. hominis infection in animal model [18], [19]. We have also demonstrated that solubilized antigen from B. hominis (Blasto-Ag), at a certain concentration, could down-regulate PBMC responses while enhancing the growth of colorectal cancer cells in vitro [20]. Additional comparative study revealed that Blasto-Ag derived from symptomatic individual has higher inhibitory effect on PBMCs, implying a higher pathogenicity (our unpublished data). Our recent study showed that infection of B. hominis in an immunocompromised condition such as cancer is opportunistic [21]. However, the infectivity of this parasite in other immune-system-altered condition such as in stress is not known.
Therefore, the main aim of this study is to provide experimental evidence on how chronic stress influences the hosts' immune system, causes oxidative damage, and increases susceptibility towards B. hominis infection. To achive this, we used mechanical stress (by shaking) which is a simple and convenient model to induce both psychological (shocked reaction) and physical (body muscle movements) stress in rats prior to exposing them to symptomatic B. hominis infection. The effects of Blasto-Ag (isolated from symptomatic human) on the in vitro growth of PBMCs isolated from rats subjected to stress and B. hominis infection were also investigated. This experimental model is hoped to lead to a better understanding of the immune cell responses towards B. hominis infection in humans with and without stress.
Materials and Methods
Source and Isolation of B. hominis cysts
B. hominis was isolated from the stool samples of a symptomatic human and the cysts were extracted using Ficoll-Paque technique with a slight modification [22]. Isolated cysts were washed in sterile saline and incubated at room temperature for two days in saline added with 100units/ml penicillin-streptomycin. These steps were repeated two to three times prior to the inoculation stage, in order to eliminate the possible bacterial contamination. Furthermore, the cysts were further cultured in Jones' medium to examine the growth of parasite. Only purified cysts without any bacterial contamination were used for the inoculation.
Ethics statement
Prior to stool sample collection, a written informed consent was obtained from all recruited individuals. The protocol used in this study was approved by the Medical Ethics committee of the UMMC (University Malaya Medical Center) which complies with the Declaration of Helsinki. The animal care and research protocol used in this study was approved by University of Malaya Institutional Animal Care and Use Committee (UM IACUC) according to The National Research Council's Guide for the Care and Use of Laboratory Animals (8th edition). Our ethics reference number is PAR/30/03/2012/SKG(R).
Inoculation of B. hominis cysts and stress induction in animal model
Three-week-old Wistar albino rats with a mean weight of 60 grams were assigned into four groups comprising 20 rats in each group namely a) control (without stress induction and without B. hominis infection), b) stress-induced, c) B. hominis infected and d) stress-induced and B. hominis infected. Weights of all rats were recorded once a week to examine changes in their weight gain. The 20 rats in each group were housed into 4 cages of 5 rats. Throughout the experiment, it was made sure that the rats from each study group were kept separated in appropriately labelled cages. A week prior to the commencement of the experiment, the rats were pre-screened in order to ensure that they were healthy and free of B. hominis or any other intestinal parasitic infection. Ten thousand cysts in 1 ml sterile saline (without penicillin-streptomycin) were orally inoculated to each rat in the infected group. The control group was inoculated with 1 ml of sterile saline. For the stress-induced group, the respective cages of rats were placed on a Belly Dancer Shaker which was shaken at a speed of 90 rpm for 1 hour on a daily basis for a period of 30 days. The undulating motion of the Belly Dancer Shaker provided gentle agitation and induced stress to the animals especially when the speed was increased gradually (done within 5 minutes prior to shaking period). The combination of both vertical and horizontal orbital movements provided by the Belly Dancer Shaker would be able to mimic physical stress better than other types of laboratory shakers namely orbital shaker.
Detection of B. hominis in stool samples
Stool samples of all rats were examined for the presence of B. hominis daily till the 14th day and then after on alternate days till the last day of experiment. Briefly, three pellets of fresh stools were emulsified in normal saline (3–5 ml) and filtered using gauze to eliminate debris. The filtrate containing cysts were then washed 3 times with PBS. After the final washing step, supernatant was discarded and the pellet was re-suspended with minimum volume of PBS (10 µl to 50 µl). A drop of this mixture (10 µl) was mixed with 10 µl of trypan blue stain and cyst count was performed using haemocytometer and light microscope. Based on the concentration of cysts determined in this step, the number of B. hominis cysts in the PBS mixture (10 µl to 50 µl) was recalculated and expressed as cyst/ml (the final volume of the PBS mixture was adjusted to 1 ml). Concurrently, stool samples were also cultured in Jones' medium and observed after 24 hours to confirm the viability of this parasite [23].
Blood sample collection
Blood samples (from five rats per group) were collected once a week using heart-puncture technique. Prior to blood sampling, each rat was placed in a covered cylindrical jar and anesthetised with diethyl ether until loss of the righting reflex which takes approximately 75 seconds. Any possible anesthetic influence on the results was reduced by using a standardized euthanizing technique to all the animals including the control group. After collecting the blood samples, rats were euthanized by overdosing with diethyl ether. Blood samples were collected in plain and EDTA coated tubes to obtain serum and whole blood respectively. Serum sample was used for biochemical and immunoglobulin analysis whereas whole blood was used for PBMC isolation and blood differential count.
Biochemical assays
Advanced oxidation protein product (AOPP) is a biomarker used for free radical induced protein damage. It is formed by the action of chlorinated oxidants especially hypochlorous acid and chloramines. The level of AOPP was measured spectrophotometrically according to an established method [24]. The level of AOPP was calculated based on a standard curve generated using chloramine T and the results were expressed as µM chloramine T equivalents.
Lipid hydroperoxide (LHP) is the intermediary product in free radical induced lipid peroxidation. LHP level was assessed according to the method by Esterbauer and Cheeseman [25], with minor modification. In this method, the LHP present in a biological sample reacts with 1-methyl-2-phenylindole (MPI) under acidic condition to form a blue-coloured chromophore which can be measured spectrophotometrically. The level of LHP was estimated using 1,1,3,3- tetraethoxypropane as a standard and the results were expressed as µM.
The non-enzymatic antioxidants or reductants in serum samples were assessed using the ferric-reducing antioxidant power (FRAP) assay, a method previously established by Benzie and Strain [26] and further modified in our laboratory. Antioxidants present in sample reduce ferric ion-tripyridyltriazine (Fe2+-TPTZ) to ferrous ion-tripyridyltriazine (Fe3+-TPTZ) at low pH causing the formation of blue-coloured ferrous-tripyridyltriazine complex which can be measured spectrophotometrically. Concentration of reductants was determined using ferrous sulphate heptahydrate (FeSO4 7H2O) as the standard and results were expressed as µM.
Glutathione peroxidase (GPx), an enzymatic antioxidant was assayed using quantitative colorimetric assay kit purchased from BioAssay Systems, USA. Assay buffer, glutathione, NADPH, and glutathione reductase enzyme were mixed together with serum sample in a 96-well plate. Finally, hydrogen peroxide (H2O2) which is the substrate for GPx enzyme was added and the absorbance at 340 nm was taken at 0 and 4th minute. This assay directly measures the NADPH consumption in the enzyme coupled reactions. The decrease in optical density is directly proportional to the enzyme activity in the sample. GPx activity (U/L) was calculated using a NADPH standard curve as recommended by the assay kit's user manual.
Blood Differential Count
A thin blood film (from ethylenediaminetetraacetic acid or EDTA tube) was prepared on a microscopic glass slide. In order to easily visualize the differences among the various types of WBCs, the blood film was stained with Giemsa stain [27]. Briefly, one hundred WBCs were counted and the percentage of the various WBC components namely neutrophils, eosinophils, lymphocytes, and monocytes were calculated.
Isolation of Peripheral Blood Mononuclear Cells (PBMCs)
Whole blood sample (approximately 2 ml) was collected in sterile EDTA-tubes. PBMCs were harvested using Histopaque-1077 (Sigma-Aldrich, USA) according to the gradient density centrifugation technique [28]. The isolated PBMCs were washed thrice with phosphate buffer saline (PBS) and transferred into growth media (Roswell Park Memorial Institute medium or RPMI-1640 supplemented with 10% Fetal Bovine Serum or FBS, 2 mM L-glutamine, 100 units/ml penicillin-streptomycin, and 2·5 µg/ml fungizone).
Isolation of B. hominis solubilized antigen (Blasto-Ag) and introduction to PBMCs
Axenization of B. hominis and isolation of Blasto-Ag were done according to the method used in our previous study [20]. Briefly, the axenized parasites were transferred into sterile Jones' medium (without any supplements) and lysed using sonication technique before incubating overnight at 4°C. The homogenate was filter sterilized and the protein concentration was determined by Bradford assay. Freshly isolated PBMCs (5×104 cells/well) in 100 µl growth medium were seeded into 96 well plates. After an overnight incubation at 37°C in a CO2 incubator containing 5% CO2, Blasto-Ag with a final concentration of 1 µg/ml was added into each well and was further incubated for 48 hours. The concentration of Blasto-Ag used in this study was based on our previous published data [20] which caused optimum proliferation of PBMCs isolated from healthy individual. In addition to this, preliminary experiment using PBMCs of 5 normal rats exposed to various concentration of Blasto-Ag (0.001 to 10 µg/ml) showed that 1 ug/ml resulted in optimum cell proliferation (our unpublished data). The changes in proliferation percentage of cells were measured using MTT (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide) assay as described previously [29]. This colorimetric assay is based on the conversion of yellow MTT dye into purple formazan crystals by the mitochondrial enzyme activity of living cells. The formazan crystals were dissolved using DMSO and the absorbance was measured at 700 nm against 560 nm.
Measurement of Immunoglobulins IgG, IgE, and IgM in serum samples
Immunoglobulins namely IgG, IgE, and IgM were measured according to the user manual of Enzyme–Linked Immunosorbent Assay (ELISA) kit purchased from Immunology Consultants Laboratory, USA. Briefly, IgG, IgE or IgM present in serum samples combined with the respective anti-immunoglobulin antibodies that have been coated on the surface of polystyrene microtitre wells (96-well plate). The unbound proteins were removed by a washing step and the addition of anti-immunoglobulin antibodies conjugated with horseradish peroxidase (HRP), form complexes with the previously bound immunoglobulin. After another washing step, the chromogenic substrate, tetramethylbenzidine (TMB) was added resulting in the formation of coloured solution which was proportional to the quantity of respective immunoglobulin present in the serum samples. This reaction was inhibited using the provided ‘Stop Solution’ that caused a colour change from blue to yellow and the absorbance was measured at 450 nm. The level of immunoglobulins in the test sample was quantified using standard curve created according to the assay kit's user manual.
Statistical analysis
All data were analyzed using SPSS version 18. Values are expressed as mean ± SD. The significant difference between the control (normal) and other groups were analyzed using Student t-test. Comparison of parameters tested among the ‘infected’, ‘stressed’, and ‘stressed and infected’ groups were done using one-way ANOVA analysis. Correlations between the parameters for both control and parasite infected animals were identified by Pearson's correlation coefficient test and differences were considered significant when P<0·05.
Results
In present study, the control group (without stress and B. hominis infection) was labelled as ‘Normal’, stress-induced group as ‘Stressed’, B. hominis infected group (without stress) as ‘Blasto’, and both stress-induced along with B. hominis infection as ‘Blasto-Stressed’. The mean weight gained in each week for all groups were compared with week 1 (Figure 1). All groups showed a significant increase in weight gain at week 4 (P<0·05) when compared to week 2. Stressed group showed the highest level of weight gain followed by Normal, Blasto, and Blasto-Stressed. The cyst count in both Blasto and Blasto-Stressed groups showed a gradual increase over the four weeks (Figure 2). The Blasto-Stressed group had significantly higher number of cysts compared to the Blasto group at week 3 and week 4 (P<0·05 respectively). Irregular shedding of B. hominis has been reported previously [30]. It is noteworthy that we observed absence or irregular cyst shedding only in some of the rats especially during the first two weeks of the study. However, the irregular shedding effect among these two study groups were normalized by expressing the mean value of cyst count obtained from 5 rats per group/week.
Figure 1. Effects of stress and B. hominis infection on weight gain.
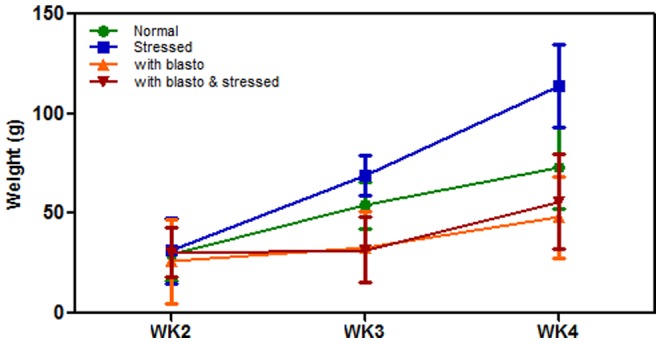
Mean weight gain is compared with their initial weight at week 1. Data shown is in mean ± SD. WK = week.
Figure 2. Cyst count in Blasto and Blasto-Stressed groups according to study duration.
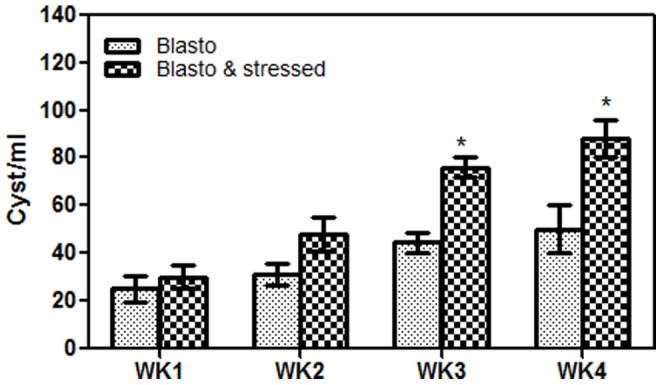
Data shown is total number of cysts recovered from 3 stool pellets from each rat (n = 5). Results are expressed as mean ± SD. *P<0.05 is the comparison done against Blasto group.
The level of IgG did not significantly differ between Normal and Stressed groups when compared between week 1 and 4 (Figure 3a). However, an increasing trend was observed in the IgG level of Blasto and Blasto-Stressed groups with a significant difference when compared to the normal group especially at week 4 (P<0·05). The levels of IgE showed an increasing trend in all four groups (Figure 3b). In the final week of experiment, Blasto-Stressed group had higher level of IgE compared to Normal and Blasto groups (P<0·05 respectively). Besides this, the level of IgM in Blasto and Blsto-Stressed groups were generally lower when compared to Normal and Stressed groups with the significant difference seen mainly during weeks 1, 2, and 3 (P<0·05 respectively, Figure 3c).
Figure 3. Levels of serum a) IgG, b) IgE, and c) IgM in the study groups according to study duration.
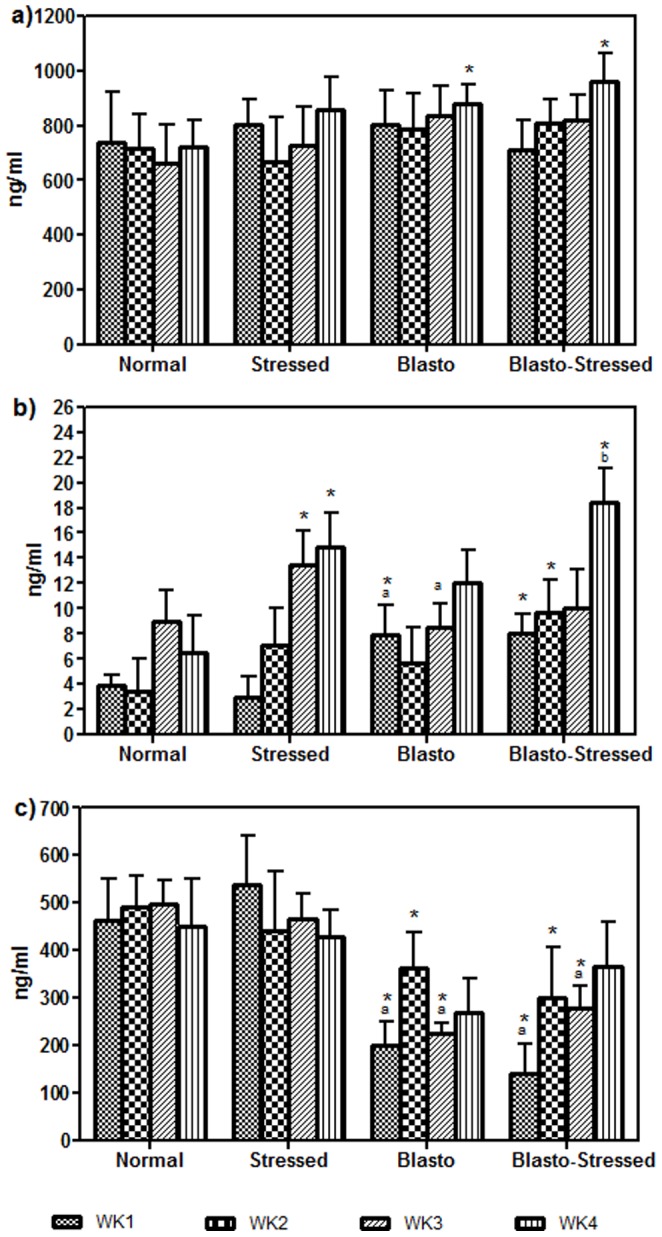
Data is given in mean ± SD. *P<0.05 is the comparison done against Normal group using Student's t-test. aP<0.05 is the significant comparison done against Stressed group using One-way ANOVA analysis.
Results of blood differential count showed that the level of neutrophils in Blasto-Stressed group was higher but not significant compared to the other three groups (Figure 4a). In contrast, the level of lymphocytes in this group was lower compared to the rest of the study groups in particular the normal group (P<0·05, Figure 4b). Level of monocytes in the Stressed group did not show any significant difference when compared to the normal group. However, there was a gradual decrease in the monocyte level of Stressed group from week 1 to week 4 (Figure 4c). During the final week, the level of monocytes in Blasto-Stressed group was significantly lower compared to Normal and Blasto groups (P<0·05). Besides this, the level of eosinophils in Blasto and Blasto-Sressed groups were significantly higher compared to normal group (P<0·05 respectively) especially in week 4 (Figure 4d).
Figure 4. Blood differential count: a) neutrophil, b) lymphocyte, c) monocyte and d) eosinophil in the respective group of rats according to study duration.

Data is given in mean ± SD. *P<0.05 is the comparison done against Normal group by Student's t-test. aP<0.05 and bP<0.05 is the comparison done against Stressed and Blasto group respectively using One-way ANOVA analysis.
Figure 5 depicts the effect of Blasto-Ag on the proliferation of PBMCs isolated from rats belonging to the respective groups. PBMCs isolated from Blasto-Stressed group showed the highest inhibition (P<0·05) when exposed to Blasto-Ag and this was followed by the Blasto and Stressed groups.
Figure 5. Proliferation of PBMCs introduced with 1 µg/ml of Blasto-Ag according to study duration.

PHA, mitogen (20 µg/ml) was used as positive control. Values are given in mean ± SD. Values are normalized against sample blank where Blasto-Ag was substituted with sterile Jones medium (without any supplements). *P<0.05 is the comparison done against Normal group by Student's t-test. aP<0.05 and bP<0.05 is the comparison done against Stressed and Blasto group respectively using One-way ANOVA analysis.
Figure 6 depicts the levels of AOPP, LHP, FRAP, and GPx in all the groups according to the weeks. Level of AOPP was significantly elevated in Stressed, Blasto, and Blasto-Stressed groups compared to the normal group especially in week 3 and 4 (P<0·05 respectively). Blasto-Stressed group showed the highest level of AOPP especially in the last two weeks of the experiment (Figure 6a). Similarly, level of LHP was the highest in Blasto-Stressed group compared to other study groups and the significant difference (P<0·05) was evident mainly in week 1 and 2 (Figure 6b). In contrast, FRAP level in all the three experimental groups was lower compared to the normal group (Figure 6c). FRAP level of Blasto-Stressed group was significantly higher than Stressed group in week 1 but decreased significantly in week 3 (P<0·05 respectively). GPx level in Blasto and Blasto-Stressed groups were generally lower compared to normal and stressed groups. During the final week of experiment, rats in the Blasto-Stressed group exhibited the lowest level of GPx compared to all other groups (P<0·05).
Figure 6. Levels of serum oxidative indices: a) AOPP, b) LHP, c) FRAP and d) GPx in the study groups according to study duration.

Data is given in mean ± SD. *P<0.05 is the comparison done against Normal by Student's t-test. aP<0.05 and bP<0.05 is the comparison done against Stressed and Blasto group respectively using One-way ANOVA analysis.
Correlation analysis of AOPP, LHP, FRAP, and GPx levels in the study groups are shown in Table 1. In the Stressed group, most of the parameters showed positive correlation. In Normal group, only FRAP and GPx correlated positively whereas in Blasto group, positive correlation existed between AOPP and GPx. Negative correlation between AOPP and FRAP was evident in Blasto-Stressed group.
Table 1. Correlation analysis of AOPP, LHP, FRAP and GPx levels in the study groups.
| Group | Parameters | Correlation |
| Normal | FRAP/GPx | r = 0·646, p<0·01 |
| Stressed | AOPP/LHP | r = 0·660, p<0·01 |
| AOPP/FRAP | r = 0·521, p<0·05 | |
| AOPP/GPx | r = 0·663, p<0·01 | |
| FRAP/GPx | r = 0·590, p<0·01 | |
| Blasto | AOPP/GPx | r = 0·550, p<0·05 |
| Blasto-Stressed | AOPP/FRAP | r = −0·591, p<0·01 |
Pearson's correlation coefficient test; differences were considered significant when p<0.05.
Discussion
Infection of B. hominis in humans is known to cause clinical symptoms such as diarrhea, anorexia, flatulence, nausea, and abdominal discomfort [31]. Its pathogenicity which is a debatable topic has stimulated interest among researchers to study the epidemiological and molecular aspects of this protozoan. Various reports have shown the association between its pathogenicity, with genotypic characteristics and serious illnesses namely Irritable Bowel Syndrome as well as colorectal cancer. To date this is the first study to report on the effect of stress on the infectivity and pathogenicity of B. hominis infection in an animal model.
In the present study, the physical stress (induced by Belly Dancer Shaker) introduced to the rats is considered a form of stress as these rats are unfamiliar to such agitation and is not their normal living condition. This could disrupt their homeostasis system which will eventually result in oxidative stress as well as immune system malfunction. Alteration in the immune cell responses will destroy the efficiency to fight against infection. Here, we studied the effects of stress against the infectivity and pathogenicity of the common intestinal parasite, B. hominis. The effects were evaluated using several parameters namely weight gain, cyst count, alteration in immunoglobulin levels, alteration in blood differential count, PBMC response in vitro and the level of oxidative indices as summarized in Figure 7a. Results obtained are discussed according to the number order (in the blue boxes) which indicates the experimental flow.
Figure 7. Illustration on the a) effects of stress on various parameters and b) overview of results obtained from this study.

Text box shaded in a) yellow represents general facts and hypothesis, b) blue represents the parameters used, c) grey represents observations obtained, and d) pink represents implications of the current study.
Stress response involves a range of biological and behavioral paths which is important for the survival or adaptations to changes in the environmental conditions. Stress triggers the hypothalamic-pituitary-adrenal axis to release glucocorticoid hormone (exist predominantly as corticosterone in rodents and cortisol in humans) that regulates the cascade of metabolic pathways in order to retain the internal physiological balance [32]. A study done on human subjects has reported that stress can cause both increase and decrease in weight or food intake [33] that has been linked with the level of glucocorticoid hormone. In current study, rats in Stressed group exhibited the highest level of weight gain compared to Normal and the other two groups (Figure 1). In contrast, researchers have also reported that restraint stress on rats resulted in decreased weight gain compared to the controls [34]. Another study has reported that only variable stressors (such as food deprivation, isolation, and swim stress) caused weight loss in rats compared to the chronic stress [35]. Similar to our findings, Badiani and colleagues [36] have reported on the significant increase in food intake in rats subjected to only a brief restraint period. However, the stress exposure period in all those studies were relatively short, ranging from 7 to 20 days and the effect of infection in the animals were not investigated. In our study, decrease in weight gain compared to the control rats were only observed when B. hominis infection was present (Blasto and Blasto-Stressed groups). Since the corticosterone level was not measured as a stress indicator we are unable to rule out the possible influence of B. hominis infection especially in the presence of stress on this glucocorticoid hormone. In view of the numerous previous studies which support the notion that the corticosterone level might not be a reliable marker of chronic stress, the exclusion of this hormone in the present study is well justified. Previous study has reported that increase in plasma corticosterone was independent of reduced food intake in rats experimentally infected with Nippostrongylus brasiliensis, a gastrointestinal nematode of rats [37]. Additionally, another study suggested that it is possible to claim that chronic stress may not directly affect the level of serum or plasma glucocorticoids [38]. The key point here is the response of target tissues or organs and the resulting oxidative damage rather than the levels of the glucocorticoid stress hormone. In view of the fact that previous evidences have shown insignificant change in the levels of glucocorticoid as well as the presence of glucocorticoid receptor resistance (GCR) in humans and(or) animals exposed to chronic stress [39], [40], it was more feasible to measure the end-products of oxidative damage as indicators of oxidative stress. Furthermore, researchers had studied the effect of chronic stress on oxidative damage parameters in rats without measuring corticosterone level [41]. Nevertheless, it may be feasible to include corticosterone level in brain or other targeted organs in future studies pertaining to chronic stress and its effect on HPA (Hypothalamic–pituitary–adrenal) axis.
Generally, a high cyst count is an indication of a very high number of trophozoites (which have reverted to become cysts) in the intestinal tract which are continuously multiplying through binary fusion. In addition to this, an increase in cyst count has been associated with the capability of a parasite to proliferate by escaping from host immune response. Studies have therefore correlated high number of cysts with susceptibility towards an infection [42]. A higher level of cyst count in the Blasto-Stressed group compared to the Blasto group observed in our study (Figure 2) has clearly suggested that stress increased the susceptibility towards B. hominis infection by suppressing the hosts' immune system. It is established that stress leads to the suppression of immune cell responses which inevitably creates a favorable condition for the growth of infectious microorganisms [43] including B. hominis. Generally, immune system in our body combats the invading antigens or parasites by two types of responses namely innate and acquired immunity [9]. The innate immunity involves natural killer (NK) cells and phagocytes or white blood cells (e.g.:monocytes, neutrophil, eosinophil) which eradicate the antigens by phagocytosis. The acquired immunity involves T helper (Th) lymphocytes that can be subdivided into Th1 and Th2 responses, also known as cellular and humoral immunity respectively. Th1 or cellular immunity is the defense against intracellular pathogens. In contrast, Th2 or humoral immunity plays the major role in defense against extracellular pathogens by triggering the production of immunoglobulin subtypes such as IgG, IgE, and IgM.
Generally, IgG makes up 80% of our circulating immunoglobulins and it is the most versatile immunoglobulin to act against invading pathogens. Its activity involves phagocytosis and activation of complement system to eradicate infection. IgE is secreted by plasma cells of mucous membrane and has been often associated with inflammatory responses. Increase in total serum IgE has been reported in humans with intestinal parasitic infection [44], [45]. Meanwhile IgM, an antibody found on the B cell surface is the first antibody produced to fight against an infection by involving humoral immunity [9].
Based on their immunological functions IgG, IgE and IgM have been previously assessed to understand the immune response of host infected with intestinal parasites [44], [46]. One of these studies [46] included the assessment of IgA which is known to be secreted by mucosal lining of gastrointestinal tracts [9]. Our study did not include the measurement of serum IgA that may reflect the adherence of B. hominis on intestinal epithelial layer. Nevertheless, a past study reported that B.hominis infection in mice resulted in predominance of IgA only in intestinal secretions while the serum immunoglobulins were dominated by IgM [47]. A low level of IgA has been reported in humans with B. hominis infection [48]. Besides this, insignificant level of serum IgA in patients infected with giardia (a type of intestinal protozoa) has also been observed when comparing with a healthy group [46]. Moving forward, future studies should include IgA in both serum and intestinal secretions in order to study the effect of stress against the pathogenesis of B. hominis in terms of damage caused to host's intestinal mucosal layer. This may be further confirmed using histopathological examination of intestinal tissues as well as assessment of protease (enzyme involved in extracellular matrix digestion) activity in intestinal secretions which have been closely associated with invasive pathogens.
In the current study, variation in the levels of total serum immunoglobulins and blood differential count (Figure 3 and 4) in Stressed, Blasto, and Blasto-Stressed groups suggest that stress and B. hominis infection independently cause dysregulation of immune system in these rats. Significantly elevated levels of IgG and inflammation associated immunoglobulin, IgE in B. hominis infected groups (regardless of stress) compared to the normal group (Figure 3a and 3b) suggest that the parasite attracts these two immunoglobulins to mediate activity of immune cells namely eosinophils in order to combat against the infection. The increased level of eosinophils observed in the Blasto and Blasto-stressed groups supports this speculation (Figure 4d). Furthermore, there are evidences to support the view that IgG [49] and IgE [50] are associated with eosinophil activity to eradicate parasitic infection. In this study, significantly elevated level of serum IgE (Figure 3b) in Blasto-Stressed group compared to Blasto group clearly suggests that stress aggravates the immune dysregulation which may assist in the infectivity of this protozoan as seen in Figure 2. To date there have been no studies done on animal model to evaluate the levels of total serum immunoglobulins in the presence of stress induced B. hominis infection. A survey done on 141 leukemic children with high prevalence of B. hominis infection has reported that 26% and 41% of them had increased level of serum IgG and IgM respectively [51]. However, the study did not include assessment of leucocytes in these patients. Conversely, in the present study, the IgM level in both Blasto and Blasto-Strssed groups was significantly decreased compared to the non-infected groups (Figure 3c). Similar to our findings, a previous research has shown that serum IgM level in both symptomatic and asymptomatic patients with B. hominis were lower than the normal range [48]. These findings suggest that the effects of B. hominis infection on total immunoglobulins may vary according to different study population and therefore requires further scrupulous investigations pertaining to immunological pathways. In addition to this, suppressed level of lymphocytes (Figure 4b) and monocytes (Figure 4c) of B. hominis infected groups especially in stressed condition (Blasto-Stressed group) leads to the speculation that stress-induced suppression of immune system facilitates the growth of B. hominis infection.
Besides this, neutrophils and monocytes are known to be mediators of oxidative stress. However, in the current study there were no changes in the levels of neutrophils especially at week 4 as observed in monocytes. Several in vitro studies have shown that the ability of neutrophils (obtained from subjects exposed to brief or maximal exercise) to generate ROS is variable whereby both increase and decrease were evident [52]–[54]. These evidences have indicated that ROS produced during a stress-induced oxidative stress condition may not be totally influenced by the level of neutrophils. In addition, a past study using parasite infected mice model reported that only inflammatory monocytes and not neutrophils were required to combat the infection [55]. In our research, significantly elevated level of monocytes were observed in Blasto-Stressed group in the first week (compared to normal group) which subsequently declined by week 4 (compared to both normal and infected groups). These observations suggest that the monocytes' response in a B. hominis infected host could be suppressed when stress is concurrently present.
Investigations on the efficiency of immune system is generally evaluated with the rate of proliferation of lymphocytes (a subset of peripheral blood mononuclear cells) upon the exposure to parasite-specific antigens and non-specific mitogens especially phytohemagglutinin (PHA) [43]. Although immunoglobulin responses are often studied using B cell mitogen namely pokeweed mitogen (PWM), past study has reported that B cells could proliferate in PHA-stimulated lymphocyte cultures [56]. Therefore, in present study PHA was used as positive control to look at the effect of Blasto-Ag towards the growth of PBMCs which consist of both B cells and T cells. Higher level of PBMC proliferation in vitro suggests a better establishment of immune responses against the infection [43]. Therefore in this study, we also investigated the effect of solubilized Blasto-Ag on the in vitro proliferation level of PBMCs isolated from all four studied groups. Comparison with controls especially in the final week revealed that PBMCs isolated from stressed rats (without infection) had significantly lower level of proliferation and inhibition when stimulated with PHA and Blasto-Ag respectively (Figure 5). This observation supports the general perception that stress is a strong suppressor of immune system thus increasing the susceptibility to infection [43]. Significant inhibition of PBMCs isolated from Blasto group upon exposure to Blasto-Ag (week 1, 2, and 3) clearly suggests that B. hominis possesses a very strong ability to suppress immune cell response in vitro. This speculation can be supported by our previous in vitro study which showed an up-regulation of pro-apoptotic gene in human PBMCs upon exposure to Blasto-Ag [20]. Compared to all other groups, PBMCs isolated from Blasto-Stressed group exhibited highest level of inhibition (week 2, 3, and 4) when stimulated with Blasto-Ag. This result strongly indicates that stress enhances the pathogenecity of B. hominis infection by suppressing the activity of host's leucocytes which could be the underlying mechanism for the increasing number of B. hominis cysts shown in Figure 2. In addition, the effect of mitogen in these PBMCs was also very minimal when compared to the control group.
As mentioned in the introduction, the process of eradicating an infection involves the host's immune system to produce active oxidants resulting in oxidative damage especially when the amount of antioxidants are lacking. Here we observed that the levels of AOPP and LHP (markers for oxidative protein and lipid damage respectively) in Stressed and Blasto groups were increased compared to the normal group (Figure 6a and 6b). This observation concurs with previous findings which showed that rats subjected to stress [57] and B. hominis infection [18] exhibited high levels of protein and lipid oxidations. Interestingly, presence of stress in B. hominis infected rats (Blasto-Stressed group) resulted in the elevation of protein and lipid oxidation compared to other groups. In contrast, the levels of both enzymatic and non-enzymatic antioxidants (indicated by FRAP and GPx levels respectively) in the Blasto-Stressed group were significantly lower compared to the normal as well as other groups (Figure 6c and 6d). The decrease of FRAP level in the infected group is in contrast to the observation in our previous study [18]. This disparity could be attributed to the nature of the B. hominis isolate used in the previous study (asymptomatic) compared to the symptomatic one used in this study. Such observations suggest that symptomatic B. hominis used here could be more pathogenic thus resulting in the suppression of this antioxidant level (vital for combating infections). Reduced levels of both FRAP and GPx in the stressed group of current study suggest that apart from immunosuppression, stress condition mediated the augmentation of B. hominis infection by down regulating oxidant-antioxidant balance in the host's system. This theory is further supported by the results obtained from correlation analysis among the oxidative indices as depicted in Table 1. In the normal group, positive correlation was present only between FRAP and GPX implicating that the levels of enzymatic and non-enzymatic antioxidants are always balanced as they are not exposed to any stressors or infections. Whereas in stressed group, correlations of AOPP/LHP, AOPP/FRAP, AOPP/GPx, and FRAP/GPx signify that stress affects the normal oxidant-antioxidant regulatory system in fighting against oxidative damage especially damage towards proteins. Existence of positive correlation of AOPP/GPx in Blasto group shows that enzymatic antioxidants play an important role to combat against the protein oxidation caused by B. hominis infection. Conversely, the negative correlation of AOPP/FRAP shows that when stress was present, the oxidative protein damage which resulted due to B. hominis infection superseded the level of antioxidant defense mechanism of the hosts.
In conclusion, the present study clearly revealed 3 strong implications: 1) Stress and B. hominis infection could independently dysregulate the immune responses as well as the oxidant-antioxidant regulatory systems; 2) Stress enhanced the susceptibility and pathogenicity of B.hominis by suppressing or dysregulating responses of PBMC and immunoglobulins; 3) Stress mediated augmentation of B. hominis by causing imbalance in the host's oxidant-antioxidant system. The summary of these implications and their relationships are depicted in Figure 7b. The parasite-stress model provided here will serve as a precedence scenario for the better understanding of the role of stress condition in humans against the susceptibility and pathogenicity of B. hominis infection.
Acknowledgments
We thank Nurul Hafefee from the Department of Biomedical Science Program, University of Malaya, for assisting in handling the animals. Special thanks to all staffs of the Department of Parasitology and Department of Biomedical Science Program for providing technical assistance in carrying out the experiments. The study was supported by UM HIR grant no:- UM.C/HIR/MOHE/MED/44.
Funding Statement
This research project was funded by The University Malaya High Impact Grant UM.C/625/1/HIR, Malaysia. The funding source did not play any role in study design, data collection, analysis, interpretation of data, or writing of the report. The corresponding author had the final decision to submit this paper for publication.
References
- 1. Cohen S, Janicki-Deverts D (2012) Who's stressed? Distributions of psychological stress in the United States in probability samples from 1983, 2006, and 2009. J Appl Soc Psychol 42: 1320–1334. [Google Scholar]
- 2. von Borell EH (2001) The biology of stress and its application to livestock housing and transportation assessment. J Anim Sci 79: E260–267. [Google Scholar]
- 3. Nadeem A, Masood A, Masood N, Gilani RA, Shah ZA (2006) Immobilization stress causes extra-cellular oxidant-antioxidant imbalance in rats: restoration by L-NAME and vitamin E. Eur Neuropsychopharmacol 16: 260–267. [DOI] [PubMed] [Google Scholar]
- 4. Halliwell B (1994) Free radicals, antioxidants, and human disease: curiosity, cause, or consequence? Lancet 344: 721–724. [DOI] [PubMed] [Google Scholar]
- 5. Zhang S, Xu Z, Gao Y, Wu Y, Li Z, et al. (2012) Bidirectional crosstalk between stress-induced gastric ulcer and depression under chronic stress. PLoS One 7(12): e51148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Herrmann M, Scholmerich J, Straub RH (2000) Stress and rheumatic diseases. Rheum Dis Clin North Am 26: : 737–763, viii. [DOI] [PubMed] [Google Scholar]
- 7. Frick LR, Arcos ML, Rapanelli M, Zappia MP, Brocco M, et al. (2009) Chronic restraint stress impairs T-cell immunity and promotes tumor progression in mice. Stress 12: 134–143. [DOI] [PubMed] [Google Scholar]
- 8. Gunasekaran R (2001) Effect of chronic vibration on the immune state of albino rats. Indian J Physiol Pharmacol 45: 487–492. [PubMed] [Google Scholar]
- 9.Roitt I, Brosto J, Male D (2001) Immunology, 6th ed. St. Louis: Mosby. 480p.
- 10. Cooper PJ, Chico ME, Sandoval C, Nutman TB (2004) Atopic phenotype is an important determinant of immunoglobulin E-mediated inflammation and expression of T helper cell type 2 cytokines to ascaris antigens in children exposed to ascariasis. J Infect Dis 190: 1338–1346. [DOI] [PubMed] [Google Scholar]
- 11. Takeda K, Omata Y, Koyama T, Ohtani M, Kobayashi Y, et al. (2003) Increase of Th1 type cytokine mRNA expression in peripheral blood lymphocytes of calves experimentally infected with Cryptosporidium parvum . Vet Parasitol 113: 327–331. [DOI] [PubMed] [Google Scholar]
- 12. Geiger SM, Massara CL, Bethony J, Soboslay PT, Carvalho OS, et al. (2002) Cellular responses and cytokine profiles in Ascaris lumbricoides and Trichuris trichiura infected patients. Parasite Immunol 24: 499–509. [DOI] [PubMed] [Google Scholar]
- 13. Tekwani BL, Mehlotra RK (1999) Molecular basis of defence against oxidative stress in Entamoeba histolytica and Giardia lamblia . Microbes Infect 1: 385–394. [DOI] [PubMed] [Google Scholar]
- 14. Govind SK, Khairul AA, Smith HV (2002) Multiple reproductive processes in Blastocystis. Trends Parasitol 18: 528. [DOI] [PubMed] [Google Scholar]
- 15. Wong KH, Ng GC, Lin RT, Yoshikawa H, Taylor MB, et al. (2008) Predominance of subtype 3 among Blastocystis isolates from a major hospital in Singapore. Parasitol Res 102: 663–670. [DOI] [PubMed] [Google Scholar]
- 16. Tan TC, Suresh KG, Smith HV (2008) Phenotypic and genotypic characterisation of Blastocystis hominis isolates implicates subtype 3 as a subtype with pathogenic potential. Parasitol Res 104: 85–93. [DOI] [PubMed] [Google Scholar]
- 17. Yakoob J, Jafri W, Jafri N, Khan R, Islam M, et al. (2004) Irritable bowel syndrome: in search of an etiology: role of Blastocystis hominis . Am J Trop Med Hyg 70: 383–385. [PubMed] [Google Scholar]
- 18. Chandramathi S, Suresh K, Shuba S, Mahmood A, Kuppusamy UR (2010) High levels of oxidative stress in rats infected with Blastocystis hominis . Parasitology 137: 605–611. [DOI] [PubMed] [Google Scholar]
- 19. Chandramathi S, Suresh KG, Mahmood AA, Kuppusamy UR (2010) Urinary hyaluronidase activity in rats infected with Blastocystis hominis—evidence for invasion? Parasitol Res 106: 1459–1463. [DOI] [PubMed] [Google Scholar]
- 20. Chandramathi S, Suresh K, Kuppusamy UR (2010) Solubilized antigen of Blastocystis hominis facilitates the growth of human colorectal cancer cells, HCT116. Parasitol Res 106: 941–945. [DOI] [PubMed] [Google Scholar]
- 21. Chandramathi S, Suresh K, Anita ZB, Kuppusamy UR (2012) Infections of Blastocystis hominis and microsporidia in cancer patients: are they opportunistic? Trans R Soc Trop Med Hyg 106: 267–269. [DOI] [PubMed] [Google Scholar]
- 22. Zaman V, Khan KZ (1994) A concentration technique for obtaining viable cysts of Blastocystis hominis from faeces. J Pak Med Assoc 44: 220–221. [PubMed] [Google Scholar]
- 23. Suresh K, Smith H (2004) Comparison of methods for detecting Blastocystis hominis . Eur J Clin Microbiol Infect Dis 23: 509–511. [DOI] [PubMed] [Google Scholar]
- 24. Witko-Sarsat V, Friedlander M, Nguyen Khoa T, Capeillere-Blandin C, Nguyen AT, et al. (1998) Advanced oxidation protein products as novel mediators of inflammation and monocyte activation in chronic renal failure. J Immunol 161: 2524–2532. [PubMed] [Google Scholar]
- 25. Esterbauer H, Cheeseman KH (1990) Determination of aldehydic lipid peroxidation products: malonaldehyde and 4-hydroxynonenal. Methods Enzymol 186: 407–421. [DOI] [PubMed] [Google Scholar]
- 26. Benzie IF, Strain JJ (1996) The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 239: 70–76. [DOI] [PubMed] [Google Scholar]
- 27. Sivanandam S, Mak JW (1974) Various Giemsa preparations and their use in filariasis. J Med Health Lab Technol Malays 1: 9–11. [Google Scholar]
- 28. Boyum A (1974) Separation of blood leucocytes, granulocytes and lymphocytes. Tissue Antigens 4: 269–274. [PubMed] [Google Scholar]
- 29. Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 65: 55–63. [DOI] [PubMed] [Google Scholar]
- 30. Vennila GD, Suresh Kumar G, Khairul Anuar A, Rajah S, Saminathan R, et al. (1999) Irregular shedding of Blastocystis hominis . Parasitol Res 85: 162–164. [DOI] [PubMed] [Google Scholar]
- 31. Sheehan DJ, Raucher BG, McKitrick JC (1986) Association of Blastocystis hominis with signs and symptoms of human disease. J Clin Microbiol 24: 548–550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Sage D, Maurel D, Bosler O (2001) Involvement of the suprachiasmatic nucleus in diurnal ACTH and corticosterone responsiveness to stress. Am J Physiol Endocrinol Metab 280: E260–269. [DOI] [PubMed] [Google Scholar]
- 33. Kivimaki M, Head J, Ferrie JE, Shipley MJ, Brunner E, et al. (2006) Work stress, weight gain and weight loss: evidence for bidirectional effects of job strain on body mass index in the Whitehall II study. Int J Obes (Lond) 30: 982–987. [DOI] [PubMed] [Google Scholar]
- 34. Roman CW, Lezak KR, Kocho-Schellenberg M, Garret MA, Braas K, et al. (2012) Excitotoxic lesions of the bed nucleus of the stria terminalis (BNST) attenuate the effects of repeated stress on weight gain: evidence for the recruitment of BNST activity by repeated, but not acute, stress. Behav Brain Res 227: 300–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Marin MT, Cruz FC, Planeta CS (2007) Chronic restraint or variable stresses differently affect the behavior, corticosterone secretion and body weight in rats. Physiol Behav 90: 29–35. [DOI] [PubMed] [Google Scholar]
- 36. Badiani A, Jakob A, Rodaros D, Stewart J (1996) Sensitization of stress-induced feeding in rats repeatedly exposed to brief restraint: the role of corticosterone. Brain Res 710: 35–44. [DOI] [PubMed] [Google Scholar]
- 37. Roberts HC, Hardie LJ, Chappell LH, Mercer JG (1999) Parasite-induced anorexia: leptin, insulin and corticosterone responses to infection with the nematode, Nippostrongylus brasiliensis . Parasitology 118 (Pt 1): 117–123. [DOI] [PubMed] [Google Scholar]
- 38. Cole SW (2008) Social regulation of leukocyte homeostasis: the role of glucocorticoid sensitivity. Brain Behav Immun 22: 1049–1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Cohen S, Janicki-Deverts D, Doyle WJ, Miller GE, Frank E, et al. (2012) Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc Natl Acad Sci U S A 109: 5995–5999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Vogel WH, Jensh R (1988) Chronic stress and plasma catecholamine and corticosterone levels in male rats. Neurosci Lett 87: 183–188. [DOI] [PubMed] [Google Scholar]
- 41. Liu J, Wang X, Shigenaga MK, Yeo HC, Mori A, et al. (1996) Immobilization stress causes oxidative damage to lipid, protein, and DNA in the brain of rats. FASEB J 10: 1532–1538. [PubMed] [Google Scholar]
- 42.McBerry C, Dias A, Shryock N, Lampe K, Gutierrez FR, et al. PD-1 modulates steady-state and infection-induced IL-10 production in vivo. Eur J Immunol. [DOI] [PMC free article] [PubMed]
- 43. Marsland AL, Bachen EA, Cohen S, Rabin B, Manuck SB (2002) Stress, immune reactivity and susceptibility to infectious disease. Physiol Behav 77: 711–716. [DOI] [PubMed] [Google Scholar]
- 44. Ehiaghe A, Agbonlahor E, Ifeanyichukwu O, Kester D, Ehiaghe oI, et al. (2013) Serum immunoglobulin E level of children infected with intestinal parasite in Okada, Nigeria. Open J Immunol 3: 123–126. [Google Scholar]
- 45. Jalalian M, Rezaiian M, Kia EB, Massoud J, Mahdavi M, et al. (2004) Relationship between serum IgE and intestinal parasites. Iranian J Publ Health 33: 18–21. [Google Scholar]
- 46. Ertan Kara P, Yereli K, Kasirga E, Kurt O, Sanlidag T, et al. (2004) The examination of link between blood levels of IgA, IgE, IgG, IgM, CRP and giardiasis in children. Ege Tip Dergisi 43: 91–94. [Google Scholar]
- 47. Santos HJ, Rivera WL (2009) Kinetic analysis of antibody responses to Blastocystis hominis in sera and intestinal secretions of orally infected mice. Parasitol Res 105: 1303–1310. [DOI] [PubMed] [Google Scholar]
- 48. Angelov I, Lukanov T, Tsvetkova N, Petkova V, Nicoloff G (2008) Clinical, immunological and parasitological parallels in patients with Blastocystosis. J of IMAB 14: 55–58. [Google Scholar]
- 49. Khalife J, Dunne DW, Richardson BA, Mazza G, Thorne KJ, et al. (1989) Functional role of human IgG subclasses in eosinophil-mediated killing of schistosomula of Schistosoma mansoni . J Immunol 142: 4422–4427. [PubMed] [Google Scholar]
- 50. Gounni AS, Lamkhioued B, Ochiai K, Tanaka Y, Delaporte E, et al. (1994) High-affinity IgE receptor on eosinophils is involved in defence against parasites. Nature 367: 183–186. [DOI] [PubMed] [Google Scholar]
- 51. Gharavi MJ, Ashraf F, Vosough P, Rokni MB (2003) Survey of intestinal parasitic infection in leukemic children and evaluation of their serum immunoglobulins. Iran J Public Health 32: 19–21. [Google Scholar]
- 52. Peake J, Suzuki K (2004) Neutrophil activation, antioxidant supplements and exercise-induced oxidative stress. Exerc Immunol Rev 10: 129–141. [PubMed] [Google Scholar]
- 53. Peake JM (2002) Exercise-induced alterations in neutrophil degranulation and respiratory burst activity: possible mechanisms of action. Exerc Immunol Rev 8: 49–100. [PubMed] [Google Scholar]
- 54. Suzuki K, Sato H, Kikuchi T, Abe T, Nakaji S, et al. (1996) Capacity of circulating neutrophils to produce reactive oxygen species after exhaustive exercise. J Appl Physiol (1985) 81: 1213–1222. [DOI] [PubMed] [Google Scholar]
- 55. Dunay IR, Fuchs A, Sibley LD (2010) Inflammatory monocytes but not neutrophils are necessary to control infection with Toxoplasma gondii in mice. Infect Immun 78: 1564–1570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Knuutila S, Kovanen PE (1987) Relative proportions of mitotic T and B cells in PHA-stimulated lymphocyte cultures. Cancer Genet Cytogenet 29: 151–154. [DOI] [PubMed] [Google Scholar]
- 57. Sahin E, Gumuslu S (2007) Immobilization stress in rat tissues: alterations in protein oxidation, lipid peroxidation and antioxidant defense system. Comp Biochem Physiol C Toxicol Pharmacol 144: 342–347. [DOI] [PubMed] [Google Scholar]