

Abstract
HLA-G is a nonclassical major histocompatibility complex molecule first described at the maternal-fetal interface, on extravillous cytotrophoblasts. Its expression is restricted to some tissues in normal conditions but increases strongly in pathological conditions. The expression of this molecule has been studied in detail in cancers and is now also beginning to be described in infectious diseases. The relevance of studies on HLA-G expression lies in the well known inhibitory effect of this molecule on all cell types involved in innate and adaptive immunity, favoring escape from immune control. In this review, we summarize the features of HLA-G expression by type of infections (i.e, bacterial, viral, or parasitic) detailing the state of knowledge for each pathogenic agent. The polymorphism, the interference of viral proteins with HLA-G intracellular trafficking, and various cytokines have been described to modulate HLA-G expression during infections. We also discuss the cellular source of HLA-G, according to the type of infection and the potential role of HLA-G. New therapeutic approaches based on synthetic HLA-G-derived proteins or antibodies are emerging in mouse models of cancer or transplantation, and these new therapeutic tools may eventually prove useful for the treatment of infectious diseases.
1. Introduction
HLA-G was first described by Geraghty et al. in 1987 [1] as a member of the nonclassical human leukocyte antigen (HLA) family, which also includes HLA-E and F [2, 3]. The HLA-G gene is located within the major histocompatibility complex on the p21.31 region of chromosome 6. It has eight exons and seven introns, and its sequence is about 86% identical to the consensus sequence of the HLA-A, -B, and -C genes. Unlike classical class I molecules, HLA-G has a short cytoplasmic tail of six amino acids, due to premature stop codon in exon 6 [1]. Alternative splicing of the primary transcript generates four membrane-bound isoforms and three soluble forms. HLA-G1 has a structure similar to that of classical HLA class I molecules: a heavy chain consisting of three extracellular globular domains (α1, α2, α3) noncovalently associated with the β-2 microglobulin and a monomer peptide. The membrane-bound isoforms, HLA-G2, -G3, and -G4, are truncated isoforms lacking the α2 and/or α3 domains of the heavy chain [4, 5] and they should not, therefore, bind β-2 microglobulin [6]. Soluble HLA-G isoforms are generated either by alternative splicing of the HLA-G primary transcript (HLA-G5, -G6, and -G7) or by proteolysis of the HLA-G1 isoform (HLA-G1s) [7–9]. Indeed, the HLA-G5, -G6, and -G7 isoforms are highly unusual, as they are spliced variants of the HLA-G mRNA retaining introns 4 and 2 [7, 9, 10].
HLA-G is structurally diverse, with (i) different isoforms resulting from alternative splicing, (ii) some β2 M-free molecules [11], and (iii) all isoforms other than HLA-G3 being able to form homomultimers [12]. Indeed, HLA-G isoforms can also form homotrimers and homodimers, through the establishment of disulfide bridges between cysteine residues located in positions 42 and 147 [13]. Truncated isoforms of HLA-G can also carry out the biological functions of this molecule. Indeed, multimeric structures of HLA-G isoforms function by differential binding to LILRB receptors [12]. Thus, HLA-G has specific features not found in other HLA class I molecules, such as (i) limited polymorphism [14, 15], (ii) restricted expression in physiological conditions [16], (iii) a shorter cytoplasmic tail region due to a stop codon in exon 6, (iv) unusual regulatory mechanisms due to the use of a promoter unique among HLA class I genes [17–20], and (v) numerous immunomodulatory properties, as described below.
HLA-G expression was initially described as restricted to the maternal-fetal interface, on extravillous cytotrophoblasts [21]. In healthy, nonfetal subjects, the HLA-G protein is found only on the cornea [22], thymic medulla [23], nail matrix [24], beta cells of the islets of Langerhans [25], mesenchymal stem cells [26], and endothelial precursors [27].
Levels of this protein are upregulated in many diseases and this upregulation may modulate the immune response.
The immunosuppressive properties of HLA-G have been thoroughly described. Indeed, the role of this molecule in immunotolerance was first described following its detection at the maternal-fetal interface, in in vitro studies, and has recently been confirmed by in vivo studies in mice. HLA-G can inhibit all types of immune competent cells (Figure 1). This effect is mediated by the direct binding of both completely soluble and membrane-bound isoforms to inhibitory receptors via the α3 domain. Indeed, B and T lymphocytes, NK cells, and monocytes of the myeloid lineage express the immunoglobulin-like transcript ILT2 (CD85j, ILIRB1) [28]; monocytes, macrophages, and dendritic cells express ILT-4 (CD85d, LILRB2) [29]. The killer cell immunoglobulin-like receptor (KIR2DL4/p49) is specific for HLA-G and is expressed by decidual NK cells. Unlike other inhibitory receptors, it may also mediate activation [30, 31]. In addition, soluble HLA-G triggers the apoptosis of T and NK cells via CD8-like classical class I soluble molecules [32]. HLA-G modulates adaptive and innate immunity by interacting with T or B lymphocytes and NK cells or polymorphonuclear cells (Figure 1).
Figure 1.
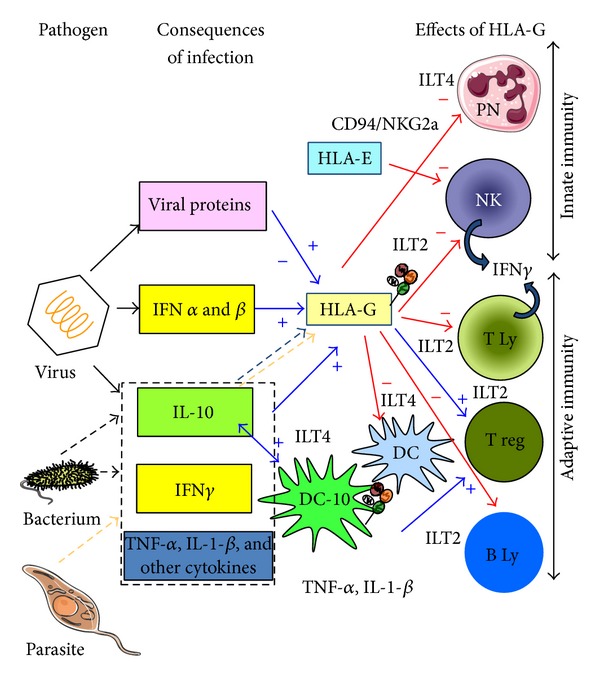
Causes and consequences of HLA-G modulation in infectious diseases. Positive and negative effects of HLA-G are shown in blue and red, respectively. Parasites, bacteria, or viruses induce the secretion of various cytokines, including IL-10 and interferon (-γ for bacterium and IFN-α and -β for virus). These cytokines upregulate the expression or secretion of HLA-G. In addition, IL-10 induces IL-10-producing human dendritic cells (DCs), termed DC-10, expressing HLA-G and ILT4. HLA-G induces tolerogenic DC in addition to DC-10 and regulatory cells via direct interaction with ILT2 and/or ILT4. HLA-G, through direct interaction with ILT2, inhibits the function of T and NK cells and B cells, whereas it inhibits the function of granulocytes and myeloid DC via direct interaction with ILT4. Indirect effects of HLA-G are mediated by the induction of HLA-E cell surface expression, which inhibits CD94/NKG2a on NK and T cells. The consequence of HLA-G action is a downregulation of innate and adaptive immunity.
HLA-G can inhibit all steps in the immune response: differentiation, proliferation, cytolysis, cytokine secretion, and immunoglobulin production. It can also alter antigen presentation to T lymphocytes, by inhibiting dendritic cell function and maturation [33–36] and by specific effects on T and B lymphocytes during effector activities. Indeed, this molecule inhibits the cytolytic activity of T and NK cells [37, 38] and the proliferation of B lymphocytes, together with the differentiation of these cells and their immunoglobulin secretion [39]. It also affects cooperation between B and T lymphocytes, by inhibiting T4 alloproliferation [36, 40] and inducing different types of regulatory T cells [41, 42]. Trogocytosis can generate different types of temporary regulatory cells in situ, accounting for the immunosuppressive effect of HLA-G-positive cells, despite their small numbers [43]. In addition, HLA-G inhibits the function of neutrophils, key cells in host immune defense against pathogens. Indeed, its interaction with its receptor, ILT4, on neutrophils impairs phagocytosis and the respiratory burst of neutrophils responsible for reactive oxygen species production [44].
Many studies have focused on HLA-G in tumoral processes, highlighting its role in tumor escape from the immune response [45]. The expression of this molecule is also beginning to be reported in other diseases, including infectious diseases. As host immune defense mechanisms efficiently eliminate most infections, studies on HLA-G in infections are based on the rationale that this molecule decreases the efficacy of the immune response through wide-ranging effects on all cell types involved in the immune response.
2. Features of HLA-G Expression by Infection Type
The main studies on HLA-G and infectious diseases are summarized in Table 1.
Table 1.
Summary of the main studies on HLA-G and infectious diseases.
| HLA-G level changes | Model or patients | Characteristics | References | |||
|---|---|---|---|---|---|---|
| Cell surface | Cell surface | Blood sHLA-G | ||||
| Increase | Decrease | Increase | ||||
| Infectious nonviral diseases | ||||||
| Septic shock | ↗ | n = 64 | Marked early and persistent increase: predictor of survival | [47] | ||
|
| ||||||
| Parasitic infections due to Plasmodium falciparum |
||||||
| ↘ | Malaria-infected placenta (n = 15) | Extravillous trophoblast cells (42% versus 90% in controls) Association with an increase in NK cells |
[118] |
|||
|
| ||||||
|
Leishmania infantum
visceral leishmania (VL) |
↗ |
n = 94 31 HIV+ 24 VL (7 HIV+ and 17 HIV−) 39 healthy subjects |
HIV and VL: 57% HIV alone: 81% VL-HIV seronegative: 35% |
[50] | ||
|
| ||||||
| Toxoplasma gondii | ↗ | In vitro infection of human trophoblast and BeWo cells | At mRNA and protein levels. Treatment with IL-10 decreases HLA-G expression. |
[53] | ||
| ↗ | Amniotic fluid: 58 women infected 24 noninfected |
Significantly higher levels when the fetus is congenitally infected. | [52] | |||
|
| ||||||
| Viral infections | ||||||
| HIV | ↘ | Cotransfection experiments on glioma cell line and macrophages | Nef-independent, Vpu-dependent |
[119] | ||
| ↗ | Before HAART, correlated with viral clearance and increase in CD4+ T-cell levels. Decrease after treatment (36 months) Role of interferons and cytokines Increase in shedding |
[54] | ||||
| ↗ | Infection treated (20) or not treated (3) |
Indirect induction by viral products and/or cytokines (IL-10) T lymphocytes and monocytes |
[57] | |||
| ↗ | Treated by HAART (n = 12) | Monocytes in treated patients Expression on monocytes decreases after treatment to block cytokines (IL-10) |
[56] | |||
| ↗ | Patients treated (n = 7) with HAART or with a protease inhibitor regime or after HAART stopped | Monocytes (50%) on HAART Increase with nucleoside reverse transcriptase inhibitors but not with protease inhibitors Decrease when HAART removed |
[58] | |||
| ↗ | Longitudinal study in 24 infected patients | In early phases, restored to normal level in chronic phases of untreated normal progressors and long-term nonprogressors Secretion by monocytes, dendritic cells Role of IL-10 |
[120] | |||
|
| ||||||
| hCMV | ↗ | hCMV reactivation in in vitro activated macrophages (n = 10) Patients with HCMV pneumonitis |
Day 20 poststimulation: expression in 45% of macrophages Bronchoalveolar macrophages Cooperative action of pp72 and pp86 |
[61] | ||
| ↗ | ↗ | Patients (n = 75) | Increase on peripheral monocytes (6.3% versus 1.6%) Association with increase in plasma IL-10 concentration and no significant increase in IFN-γ concentration |
[62] | ||
|
| ||||||
| Neurotropic virus (HSV-1 and RABV) | ↗ | In vitro infection of human neuron cell line (NT2-N) | Activation of HLA-G transcription Cell surface expression during RABV infection but not during HSV-1 infection |
[63, 64] | ||
|
| ||||||
| Influenza virus (IAV) H1N1 | ↗ | In vitro IAV infection of human alveolar epithelial cell line A549 | Upregulation of HLA-G m RNA and proteins | [65] | ||
| ↗ | (101) HIN1 patients (58 pandemic and 43 seasonal H1N1) | Monocytes and T lymphocytes (T reg CD4CD25FOXP3) | [66] | |||
|
| ||||||
| HPV | ↘ | Biopsies of invasive cervical carcinoma (n = 79) | Low HLA-G5 expression in all HPV-related cases | [67] | ||
|
| ||||||
| Hepatitis B virus | ↗ | ↗ | 90 acute, 131 chronic, and 152 resolved cases of hepatitis B | Chronic > acute Resolved = normal Expression on monocytes and Treg |
[68] | |
| ↗ | Chronic hepatitis B (n = 74) | Hepatocytes and biliary epithelial cells | [70] | |||
|
| ||||||
| Hepatitis C virus | ↗ | Chronic hepatitis C (n = 67) | sHLA-G = sHLA-G1 and-G5 Increase in plasma IL-10 and IFN-γ concentrations |
[69] | ||
| ↗ | Liver biopsies of patients with chronic hepatitis C (n = 89) | Hepatocytes More frequent in milder stages |
[121] | |||
| ↗ | Liver biopsies of patients with chronic hepatitis C (n = 20) | Significant correlation with the area of liver fibrosis HLA-G-positive cells are mast cells Soluble HLA-G secretion by human mast cells regulated by class I interferons |
[71] | |||
(a) Bacterial Infections. Septic shock is characterized by high mortality (40–50%) despite adequate initial treatment. Indeed, during septic shock, the initial huge systemic inflammatory response is immediately followed by an anti-inflammatory process, acting as negative feedback. However, this compensatory inhibitory response may subsequently become deleterious, as nearly all immune functions are compromised [46].
Monneret et al. [47] reported that marked, persistent HLA-G5 expression in septic shock was predictive of survival. The exocytosis-mediated upregulation of ILT4 expression on neutrophils is inhibited in conditions of sepsis, so the large amounts of HLA-G5 found in the plasma samples of patients surviving sepsis may have allowed them to control neutrophil inflammatory activity [44]. However, soluble HLA-G concentration was not found to be predictive of the detection of bacteremia and sepsis in pediatric oncology patients with chemotherapy-induced febrile neutropenia [48].
(b) Parasitic Infections. Few clinical data for parasitic infections are available, and those published relate mostly to plasma concentrations of sHLA-G, with the exception of one study of the protective role of HLA-G polymorphism in malaria [49]. We previously reported an increase in soluble HLA-G levels in 35% of cases of visceral leishmaniasis (Leishmania infantum) (VL) in HIV-seronegative patients and 57% of patients coinfected with HIV and Leishmania infantum [50]. However, the percentage of HLA-G-positive patients and the mean sHLA-G value were significantly lower in patients with both HIV infection and VL than in the patients with HIV infection alone. These results suggest that the increase in sHLA-G levels in HIV-infected patients with VL may contribute to a general tolerogenic environment, favoring the persistence of Leishmania and shortening the life expectancy of HIV-infected patients. sHLA-G may also be an immune biomarker of successful treatment. Thus, levels of sHLA-G with indoleamine 2,3 dioxygenase (IDO) activity may thus constitute, together with Th1/Th2 cytokine levels, surrogate markers for the resolution of VL, at least in immunocompetent patients [51]. High levels of sHLA-G are found in the amniotic fluid in women acquiring toxoplasmosis during pregnancy. The levels of this protein are the highest when the fetus is congenitally infected. However, all fetuses were born alive in our small series of patients, consistent with adequate downregulation of the inflammatory response. HLA-G may, therefore, play an immunomodulatory role that is necessary to avoid fetal loss but that may lead to the maternal-fetal transmission of Toxoplasma gondii [52]. HLA-G expression increases upon the in vitro infection of primary human trophoblasts and BeWo cells with Toxoplasma gondii, probably due to the secretion of proinflammatory cytokines in response to the parasite [53].
(c) Viral Infections. Many extensive studies have been carried out on cancers, but HLA-G expression has also been studied in many viral infections, with HIV infections being the most extensively studied (at least 30 published studies).
2.1. HIV Infection
Levels of sHLA-G are significantly higher in HIV-infected patients before treatment than in healthy controls [54]. The increase in plasma sHLA-G concentration in these patients has been attributed to an increase in HLA-G secretion from intracellular stores in monocytes and dendritic cells [55]. Indeed, a longitudinal study of plasma sHLA-G concentration in HIV-infected individuals with different rates of clinical progression showed that sHLA-G expression was associated with HIV disease progression [56]. HLA-G levels are high early in infection and remain high in rapid progressors. However, these concentrations return to normal levels in the chronic phase of infection, in both untreated normal progressors and long-term nonprogressors, when the infection is controlled. Cell surface expression of HLA-G is also detected on 93% of monocytes and 34% of T lymphocytes in patients [57]. Serum concentrations of HLA-G, like those of the other classical class I molecules (sHLA-A, -B, -C), also increase in HIV-infected patients and are significantly decreased by antiretroviral therapy (highly active antiretroviral therapy, or HAART), in cases in which HIV-1 replication is strongly inhibited.
Moreover, HAART significantly decreases the concentration of circulating soluble HLA-G molecules, this decrease being correlated with viral clearance and an increase in CD4+ T cells, as reported for classical class I molecules. The decrease in sHLA-G levels after HAART reported by Cabello et al. is not consistent with the finding of an increase in HLA-G expression on monocytes following HAART in another study [56]. The agents responsible for this increase are nucleoside reverse transcriptase inhibitors rather than protease inhibitors [58]. Murdaca et al. explain these conflicting findings in terms of the membrane expression of HLA-G inducing an increase in soluble HLA-G molecule shedding [59]. However, high levels of HLA-G in peripheral monocytes were also observed in two of the 12 untreated patients, suggesting other causes unrelated to HAART [56]. High levels of HLA-G molecules are also found in the monocytes of untreated HIV-positive patients [57], possibly due to the pathogenesis of infection.
HLA-G expression may allow these cells to evade the immune system, because the protective function of HLA-G occurs after the induction of this molecule in HAART-treated HIV-1 patients, accounting for both the consistently defective function of monocytes in HIV-1-infected patients and the role of the viral reservoir present in monocytes during infection [56]. It inhibits myeloid dendritic antigen-presenting capacity via ILT4 and enhances the secretion of inflammatory cytokines [55]. HLA-G+ regulatory T cells decrease in both absolute numbers and relative proportions during progressive HIV-1 infection. Their levels are thus inversely correlated to those of phenotypic markers of immune activation. HLA-G+ T regulatory cells can decrease harmful bystander activation and may protect against HIV-1-associated immune activation and HIV-1 disease progression [60].
2.2. Human CMV (hCMV)
Both membrane-bound and soluble plasma HLA-G concentrations increase during hCMV infection. The induction of HLA-G protein in macrophages has been observed after the generation of these cells ex vivo from latently infected monocytes and after the reactivation of hCMV infection [61]. HLA-G protein has also been detected ex vivo on bronchoalveolar macrophages from patients suffering from acute hCMV pneumonitis, on peripheral monocytes and in plasma [62]. Blood sHLA-G concentration has been shown to be correlated with blood IL-10 and IFN-γ concentrations.
2.3. Neurotropic Virus
HLA-G protein has been reported to be expressed in human neurons after infection with rabies virus or herpes simplex type I, following the activation of gene transcription [63, 64].
2.4. Influenza A Virus (IAV)
HLA-G expression was first demonstrated in vitro in an alveolar epithelial cell line, at the mRNA and protein levels, after treatment with various IAV strains [65]. HLA-G expression has been detected in vivo in patients infected with the pandemic H1N1 or seasonal H1N1 [66] viruses. It has been detected on monocytes and T lymphocytes, including T4 regulatory cells in particular. This cellular HLA-G expression contrasts with the absence of an increase in the plasma concentration of this protein.
2.5. Human Papilloma Virus (HPV)
Low levels of HLA-G5 expression are observed in all HPV-related cases of invasive cervical cancer [67]. Indeed, HPV E5 may be involved in the decrease in HLA-G expression at the cell surface, because high-risk HPV oncoproteins may inhibit the promoters of HLA class I heavy chain genes and may modulate the levels of the transporter associated with antigen processing (TAP1) protein.
2.6. Hepatitis B and C Viruses (HBV and HCV)
Plasma HLA-G concentration is higher during hepatitis infection than in healthy subjects without HBV infection. It is higher in cases of chronic hepatitis B than in acute hepatitis B and it returns to normal after resolution of the infection. In addition, an increase in HLA-G cell surface expression is observed on peripheral monocytes and regulatory T cells [68]. Similarly, an increase in blood sHLA-G concentration has been reported in patients with chronic hepatitis infection [69], associated with an increase in blood IL-10 and IFN-γ concentrations.
HLA-G expression in the liver has been detected by immunohistochemical methods, in hepatocytes and biliary epithelial cells from patients with chronic hepatitis B, by Souto et al. [70]. We [71] found that the number of HLA-G+ cells was significantly correlated with the area of tissue affected by fibrosis. This led to the first demonstration that HLA-G+ cells were mast cells. HLA-G secretion was significantly induced in human mast cells stimulated with IL-10 or class I interferons.
3. Mechanisms of HLA-G Modulation during Infection
These mechanisms (polymorphism, interference of infectious proteins with HLA-G intracellular trafficking and shedding, and cytokines) are summarized in Figure 1 and Tables 2 and 3.
Table 2.
Influence of HLA-G polymorphism on susceptibility to infectious diseases.
| Pathogens | Protection | Susceptibility | Vertical transmission (mother-to-child) | References |
|---|---|---|---|---|
| HIV | HLA-G*0105N (null allele) |
[72, 73] | ||
| G*010108 allele G*010108/010401 G*010101/010108 |
[72] | |||
| G*0105N 14 bp (ins) allele +3142G (insG) haplotype |
[74] [75] [75] |
|||
| G*01:01:01 genotype | G*01:04:04 genotype | [122] | ||
| Differences in the HLA-G gene DNA sequence between mother and child | [76] | |||
| 14 bp insertion allele 14 bp + 3142G (insG) haplotype |
[75] | |||
| insG/insG diplotype in HCV coinfected |
[75] | |||
|
| ||||
| HCV | insG/insG diplotype in HIV coinfected |
[75] | ||
| +3142C allele in sickle cell disease patients | [79] | |||
| −14 bp/−14 bp genotype | [81] | |||
| HLA-G*0105N | G*010401 homozygosity for HLA-G 14 bp deletion |
[80] | ||
|
| ||||
| HPV | 14 bp ins allele | 14 bp del allele del/C haplotype with ICC development |
[82] | |
|
| ||||
| Plasmodium falciparum | +3187G allele and haplotype UTR1 |
Haplotype UTR3 | [83] | |
| +3010G and +3142C +3010G and +3196G |
[49] | |||
Table 3.
Interference of viral proteins with HLA-G intracellular trafficking; comparison with classical HLA class I molecules.
| Virus | Viral protein | Classical HLA class I | HLA-G | References | |||
|---|---|---|---|---|---|---|---|
| Mechanism | Downregulation | Downregulation | No change | Upregulation | |||
| HIV | Nef | Interacts directly with class I domain Redirects to endolysosomal pathway |
↘ | → truncated cytoplasmic domain |
[85] | ||
| Vpu | Redirects to degradation pathway Affects early step in biosynthesis |
↘ | ↘ | [119] | |||
|
| |||||||
| HCMV | US2 US11 |
Exports for cytosolic degradation |
↘ | → truncated cytoplasmic domain |
[84, 86] | ||
| US3 US6 |
Retention in endoplasmic reticulum | ↘ | ↘ | [87] | |||
| US10 |
↘
cytoplasmic tail |
[88] | |||||
| pp72 and pp86 | ↗ | [99] | |||||
|
| |||||||
| Herpes Virus | ICP47 | Inhibits TAP (transporter associated with antigen processing) | ↘ | ↘ | [90] | ||
(a) Polymorphism, Alleles, and Single-Nucleotide Polymorphisms. Firstly, HLA-G polymorphism, although limited with 40 alleles identified [15], is involved in susceptibility to viral infections, particularly those caused by HIV and HCV (Table 1). Indeed, the G*010108 allele has been reported to be associated with an increase in the risk of HIV-1 infection, whereas the G*0105N allele (null allele) has been shown to be associated with protection from infection in African women [72, 73] but a greater risk of infection in a population from north-eastern Italy [74]. Da Silva et al. have shown that HLA-G variants influence the horizontal transmission of HIV horizontal in African-derived HIV-infected patients, with a higher frequency of alleles and genotypes associated with low levels of HLA-G expression (i.e., a higher frequency of the 14 bp insertion allele) in African-derived HIV-infected individuals and a higher frequency of the 14 bp insertion +3142G (insG) haplotype and the insG/insG diplotype. In addition, a higher frequency of the ins/ins genotype is found among African-derived HIV-infected patients also infected with HCV [75].
Thus, the transmission of HIV-1 from infected mothers to their infants may be influenced by dissimilarities in their HLA-G sequences [76]. HLA-G*01:03+ mothers have recently been shown to be less likely to transmit HIV-1 to their children during the perinatal period [77]. The polymorphic sites may affect miRNA binding to the HLA-G mRNA, thereby influencing HLA-G translation [19, 78].
HLA-G polymorphism may also affect susceptibility to HCV infection in patients with sickle cell disease, because the C allele seems to confer protection against HCV, by a mechanism associated with an increase in HLA-G expression [79]. Homozygosity for the 14 bp deletion and the allele containing this deletion (010401) seems to be a risk factor for the vertical transmission of HCV, whereas the 0105N allele confers protection [80]. The HLA-G 14 bp insertion/deletion polymorphism is also a putative susceptibility factor for active hCMV infection in children [81]. Two polymorphisms in the 3′ untranslated region of the HLA-G gene (3′UTR) (14 bp ins/del, +3142C>G) are involved in susceptibility to HPV infection; indeed, the 14 bp del allele is associated with a high risk of HPV infection, and the del/C haplotype is associated with the development of invasive cervical cancer [82].
An association of HLA-G 3′UTR polymorphisms with the antibody response to Plasmodium falciparum has also recently been reported [49, 83].
(b) Interference of Viral Proteins with the Intracellular Trafficking of HLA-G. Viral proteins have generally been reported to decrease HLA class I expression, but their effect on HLA-G expression at the cell surface is more ambiguous (Table 3). Indeed, they may have no effect [84–86] or an inhibitory effect [87–90], and one study, carried out by Onno et al. [61], even reported HLA-G induction after viral reactivation in activated macrophages, through the cooperative action of the early HCMV proteins pp72 and pp86. By contrast, another team showed that HLA-G1 levels at the cell surface were downregulated and that this downregulation was dependent on hCMV short viral US glycoproteins [89]. Some US proteins have differential effects on the expression of classical HLA class I and HLA-G molecules at the cell surface, due to the shorter cytoplasmic tail of HLA-G [91] and other structural characteristics [91].
These conflicting results for hCMV may be accounted for by differences between the cell types studied (monocytes, trophoblasts, or the U373-MG astrocytoma cell line). The effects of viral proteins differ with the infected cell target, the type (classical or otherwise) of HLA class I molecules, and the membrane-bound or soluble nature of the HLA-G protein. These conclusions are illustrated by the following examples. US10 downregulates the cell surface expression of HLA-G but not that of classical class I MHC molecules [88], because the short cytoplasmic tail of HLA-G (RKKSSD) acts as a US10 substrate. On the other hand, the US2 protein decreases levels of HLA class I molecules by supporting proteasome-mediated degradation, unlike HLA-G1, which lacks the residues essential for interaction with US2 [84]. Moreover, HLA-G1 has also been reported to be targeted for degradation, independently of the cytoplasmic tail [84].
For HIV infections, the short cytoplasmic tail of HLA-G confers resistance to Nef-induced downregulation [85], whereas Nef downregulates MHC class I molecules [92].
(c) Cytokines. Many viruses have also developed other strategies for escaping host immune surveillance, such as a deregulation of the host cytokine network through the secretion of cytokines. Cytokines are also important in bacterial infections.
The interleukin- (IL-) 10 family of cytokines and the related interferon (IFN) family form the larger class II cytokine family [93]. The IL-10 family consists of three subgroups, defined on the basis of biological functions: IL-10, the IL-20 subfamily cytokines (including IL-19, IL-20, IL-22, IL-24, and IL-26), and the type III IFN group (IFNλs).
Several viruses have been shown to upregulate the expression of cellular IL-10, which is produced by monocytes and, to a letter extent, by lymphocytes and, possibly, mast cells. Other viruses, such as the Epstein-Barr virus and HCMV, have functional orthologs of IL-10. Indeed, blood IL-10 and IFN-γ concentrations are high in hCMV infection [62] and in chronic hCMV infection [69]. In bacterial infections, IL-10 is also produced during sepsis [94]. High IL-10 levels are associated with bacteremia and sepsis in febrile pediatric cancer patients with neutropenia [95].
IL-10 is a pleiotropic cytokine with both immunostimulatory and immunosuppressive properties [96]. HLA-G expression is induced following IL-10 stimulation in experiments in vitro and is associated with IL-10 expression in vivo in a context of cancer. IL-10 selectively induces HLA-G expression, at both the mRNA and protein levels, in human trophoblasts and monocytes [97]. By contrast, Zhao et al. [53] have reported that IL-10 downregulates HLA-G expression in an in vitro model based on the infection of human trophoblasts with Toxoplasma gondii.
Interferons trigger important antiviral effects during viral infections. They can be classified into three classes: (i) class I (IFN-α, -β), produced by NK cells, lymphocytes, macrophages and fibroblasts, and other molecules, such as IFN-ω and -ζ, produced by leukocytes, (ii) class II, consisting solely in IFN-γ produced by NK and T cells, and (iii) class III, recently described and including IFN-λ1 (IL-29), -λ2 (IL-28A), and -λ3 (IL-28B), produced by numerous cell types, including plasmacytoid dendritic cells. Types I and III interferons are produced by virus-infected cells. In these cells, double-stranded RNA activates the signaling cascades leading to the transcription of the IFN-α and -β genes. Following their secretion, these interferons interact with a specific IFNα/β receptor on neighboring uninfected cells and on the initial infected cells, activating a signaling cascade that produces antiviral proteins that act on viruses and upregulate HLA class I expression. IFN-γ is involved in both innate and adaptive immunity. Type III IFNs signal through a receptor complex consisting of IL10R2 and IFNL-R1 (IL-28RA). HLA-G induction by interferons has been reported in numerous studies. Indeed, different types of IFN (α, β, and γ) can induce HLA-G expression in different cell types. Yang et al. [98] reported the induction of HLA-G on Jeg 3 cells by different interferons. The induction of HLA-G on monocytes has also been reported [99]. IFN-β and -γ have recently been shown to activate HLA-G expression in a human neuron cell line infected with rabies virus [63]. HLA-G expression after IFN treatment has also been demonstrated in several tumor models, including a melanoma cell line. Thus, treatment with IFN-β or -γ increases the dimer/monomer ratio and, subsequently, affinity for the ILT2 receptor [100]. An increase in HLA-G expression, in monocytes and serum, is also observed in patients treated systemically with IFN-α [101]. Similar effects have also been reported after treatment with IFN-β1 [102]. Interferons are known to induce HLA class I expression by binding to the interferon-stimulated response element (IRSE) motif in the proximal promoter region of class I genes. This motif is absent from the HLA-G promoter [15], so the upregulation of HLA-G expression by interferons was unexpected. This upregulation was accounted for by the identification of another specific functional IRSE in the distal promoter, at a position −744 bp upstream from the ATG [103].
The early phase of septic shock is characterized by a massive release of inflammatory mediators, causing organ dysfunction and hypoperfusion. These cytokines include tumor necrosis factor-alpha (TNF-alpha), interleukin-1beta (IL-1beta), and IFN-γ. Like IFN-γ, TNF-α and IL-1 can also induce HLA-G. Indeed, TNF-α has been shown to induce a moderate increase in steady-state levels of HLA-G mRNA in human trophoblast cell lines [98]. IL-1β increases the expression of HLA-G and Toll-like receptor 4 (TLR4) in an HIF-1α-dependent manner [104].
Protease levels generally increase during bacterial and viral infections and this may lead to the proteolytic shedding of membrane-bound HLA-G in a soluble form, resulting in an increase in blood HLA-G concentration.
4. General Discussion
An upregulation of HLA-G expression has been reported in most studies of viral infection. Reported discrepancies in the results concerning HLA-G expression in hCMV or HIV infections may reflect differences in the models used or in infection status or stage between studies. This upregulation of HLA-G expression results principally from an increase in the secretion of cytokines, such as IL-10 and class I interferons. HLA-G levels increase, either at the cell surface or in the blood (sHLA-G). Indeed, concentrations of soluble HLA-G in the blood increase in some viral infections caused by HIV, hCMV, HCV, and HBV viruses, similar to classical soluble class I antigens. The increase in the secretion of cytokines, including interferons in particular, during the course of viral infection, and the use of interferons as therapeutic agents may account for the increase in HLA-G levels. Shedding, due to metalloprotease digestion, is favored by interferons and also contributes to the increase in soluble HLA-G concentration in the blood. The peripheral cells expressing HLA-G during viral infections are monocytes and T lymphocytes (HIV, influenza). Neurons and bronchoalveolar macrophages have been shown to express HLA-G in infected tissues. In HCV hepatitis, Souto et al. [70] found that hepatocytes and biliary epithelial cells expressed HLA-G, whereas we identified HLA-G-positive cells as mast cells [71]. This discrepancy can also be accounted for a difference in the definition of positivity, because we also observed a weak staining of hepatocytes but took only strong staining into account. These findings were confirmed by our findings for a human mast cell line showing that this cell line expressed HLA-G and secreted class I interferons. Moreover, mast cells may promote liver fibrosis [105] by stimulating collagen synthesis and fibroblast chemotaxis. Cytokines involved in liver fibrosis, such as IL-4 or IL-33 [106], act as chemoattractants, driving the activation of mast cells [107, 108]. In addition, mast cells secrete tryptase and many cytokines involved in fibroblast proliferation [109] and fibrogenesis [110], including IL-10 [111]. However, the role of HLA-G in viral infections remains unclear, because two hypotheses are possible. It may promote virus immune escape, as in cancers. This hypothesis is supported by the immunosuppressive properties of HLA-G, which act on all the cells involved in the immune response. In addition, sHLA-G downregulates CXCR3 levels on peripheral blood and tonsil CD56 cells [112]. This dysregulation of CXCR3 signaling due to CXCL10 deficiency impairs antiviral responses in vivo, including the antiviral response to herpes simplex virus 1 infection [113].
Alternatively, HLA-G expression or secretion may reflect an appropriate and efficient response to the inflammatory process occurring during viral infection or septic shock. Indeed, HLA-G may be beneficial during viral infection, because an increase in HLA-G concentration occurs following the secretion or therapeutic administration of interferons, classes I and III IFNs are secreted as physiologic antiviral responses, and IFN-α is an effective treatment for chronic HCV infection. We can hypothesize that the antiviral effect of classes I and III IFNs may be mediated by the properties of HLA-G, which is induced by IFN, as described above.
The immunosuppressive properties of HLA-G have been clearly demonstrated in vitro, and the role of this protein has now been elucidated in vivo. Indeed, two studies have demonstrated the involvement of this protein in tumor progression in a mouse model in vivo. In a xenograft model, the HLA-G1 isoform promotes tumor progression in immunocompetent Balb/c mice, affecting both innate and adaptive immunity. By contrast, no tumor development is observed when HLA-G is blocked by a specific antibody, demonstrating the specificity of the effect [114]. HLA-G plays a role in tumor escape, through expansion of the population of myeloid-derived suppressor cells and an alteration of the cytokine balance in favor of a Th2 response rather than a Th1/Th17 response. HLA-G expression is associated with tumor metastasis and poor survival in the Balb/c nu/nu mouse model of ovarian cancer [115]. In another model used to assess the efficacy of synthetic HLA-G proteins for therapeutic purposes in a context of transplantation, it was shown that a single treatment of skin allograft recipient mice with these proteins was sufficient to prolong graft survival significantly and that four weekly treatments were sufficient to ensure graft survival [116].
The feasibility of synthesizing effective HLA-G-derived molecules opens up new possibilities in the fields of tumor diseases and infection. For example, HCV infections are a worldwide public health problem and may be suitable for treatment with such molecules, because HLA-G expression is correlated with the area of fibrosis.
In the future, it may be possible to modulate HLA-G transcription with a miRNA, such as the hsa mir-148a and mir-152, which bind to the 3′ untranslated region of the HLA-G gene (3′UTR) [19], downregulating its mRNA levels. Indeed a polymorphism of the binding site for this miRNA (the 263del/ins SNP) has been associated with poor control of HIV infection [117].
5. Conclusions
As in cancers, HLA-G expression is upregulated in infectious diseases, in response to changes in the cytokine microenvironment, relating principally to increases in the levels of IL-10 and interferons. HLA-G expression may occur in infected tissues and/or, more frequently, in peripheral blood, in the form of sHLA-G or a membrane-bound form on monocytes or different types of T cells (CD4, T reg). This molecule may have deleterious effects, promoting pathogen escape from immune control, as reported in cancers, or it may be beneficial, as in septic shock [47], reflecting appropriate and effective feedback control of inflammatory process. The role of this protein in parasitic and viral infections remains to be elucidated. Thus, HLA-G may be a single marker of infectious diseases, related to pathogens and/or to the immune response, or it may constitute a therapeutic target, once its function has been clarified in particular types of infections.
Acknowledgments
This work was supported by grants from Inserm (Institut national de la santé et de la recherche médicale), University of Rennes 1, ANRS 2010, Ligue Nationale Contre le Cancer (Comité d'Ille et Vilaine) 2009, IFR 140 2010, and BioSit 2012. The authors thank Dr. Butterfield for generously providing the HMC1.1 human mast cell line, the Biological Resource Center (BRC) of Rennes University Hospital, and the H2P2 platform of BioSit.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Geraghty DE, Koller BH, Orr HT. A human major histocompatibility complex class I gene that encodes a protein with a shortened cytoplasmic segment. Proceedings of the National Academy of Sciences of the United States of America. 1987;84(24):9145–9149. doi: 10.1073/pnas.84.24.9145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mizuno S, Trapani JA, Koller BH, Dupont B, Yang SY. Isolation and nucleotide sequence of a cDNA clone encoding a novel HLA class I gene. Journal of Immunology. 1988;140(11):4024–4030. [PubMed] [Google Scholar]
- 3.Geraghty DE, Wei X, Orr HT, Koller BH. Human leukocyte antigen F (HLA-F). An expressed HLA gene composed of a class I coding sequence linked to a novel transcribed repetitive element. The Journal of Experimental Medicine. 1990;171(1):1–18. doi: 10.1084/jem.171.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ishitani A, Geraghty DE. Alternative splicing of HLA-G transcripts yields proteins with primary structures resembling both class I and class II antigens. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(9):3947–3951. doi: 10.1073/pnas.89.9.3947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kirszenbaum M, Moreau P, Gluckman E, Dausset J, Carosella E. An alternatively spliced form of HLA-G mRNA in human trophoblasts and evidence for the presence of HLA-G transcript in adult lymphocytes. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(10):4209–4213. doi: 10.1073/pnas.91.10.4209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Carosella ED, Favier B, Rouas-Freiss N, Moreau P, Lemaoult J. Beyond the increasing complexity of the immunomodulatory HLA-G molecule. Blood. 2008;111(10):4862–4870. doi: 10.1182/blood-2007-12-127662. [DOI] [PubMed] [Google Scholar]
- 7.Fujii T, Ishitani A, Geraghty DE. A soluble form of the HLA-G antigen is encoded by a messenger ribonucleic acid containing intron 4. Journal of Immunology. 1994;153(12):5516–5524. [PubMed] [Google Scholar]
- 8.Park GM, Lee S, Park B, et al. Soluble HLA-G generated by proteolytic shedding inhibits NK-mediated cell lysis. Biochemical and Biophysical Research Communications. 2004;313(3):606–611. doi: 10.1016/j.bbrc.2003.11.153. [DOI] [PubMed] [Google Scholar]
- 9.Paul P, Adrian Cabestre F, Ibrahim EC, et al. Identification of HLA-G7 as a new splice variant of the HLA-G mRNA and expression of soluble HLA-G5, -G6, and -G7 transcripts in human transfected cells. Human Immunology. 2000;61(11):1138–1149. doi: 10.1016/s0198-8859(00)00197-x. [DOI] [PubMed] [Google Scholar]
- 10.Moreau P, Carosella E, Teyssier M, et al. Soluble HLA-G molecule: an alternatively spliced HLA-G mRNA form candidate to encode it in peripheral blood mononuclear cells and human trophoblasts. Human Immunology. 1995;43(3):231–236. doi: 10.1016/0198-8859(95)00009-s. [DOI] [PubMed] [Google Scholar]
- 11.Morales PJ, Pace JL, Platt JS, Langat DK, Hunt JS. Synthesis of β2-microglobulin-free, disulphide-linked HLA-G5 homodimers in human placental villous cytotrophoblast cells. Immunology. 2007;122(2):179–188. doi: 10.1111/j.1365-2567.2007.02623.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Howangyin KY, Loustau M, Wu J, et al. Multimeric structures of HLA-G isoforms function through differential binding to LILRB receptors. Cellular and Molecular Life Sciences. 2012;69:4041–4049. doi: 10.1007/s00018-012-1069-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gonen-Gross T, Achdout H, Gazit R, et al. Complexes of HLA-G protein on the cell surface are important for leukocyte Ig-like receptor-1 function. Journal of Immunology. 2003;171(3):1343–1351. doi: 10.4049/jimmunol.171.3.1343. [DOI] [PubMed] [Google Scholar]
- 14.Kirszenbaum M, Djoulah S, Hors J, Prost S, Dausset J, Carosella ED. Polymorphism of HLA-G gene and protein. Journal of Reproductive Immunology. 1999;43(2):105–109. doi: 10.1016/s0165-0378(99)00025-x. [DOI] [PubMed] [Google Scholar]
- 15.Donadi EA, Castelli EC, Arnaiz-Villena A, Roger M, Rey D, Moreau P. Implications of the polymorphism of HLA-G on its function, regulation, evolution and disease association. Cellular and Molecular Life Sciences. 2011;68(3):369–395. doi: 10.1007/s00018-010-0580-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Carosella ED, Moreau P, LeMaoult J, Rouas-Freiss N. HLA-G: from biology to clinical benefits. Trends in Immunology. 2008;29(3):125–132. doi: 10.1016/j.it.2007.11.005. [DOI] [PubMed] [Google Scholar]
- 17.Gobin SJP, van den Elsen PJ. Transcriptional regulation of the MHC class Ib genes HLA-E, HLA-F and HLA-G. Human Immunology. 2000;61(11):1102–1107. doi: 10.1016/s0198-8859(00)00198-1. [DOI] [PubMed] [Google Scholar]
- 18.Gonzalez A, Rebmann V, leMaoult J, Horn PA, Carosella ED, Alegre E. The immunosuppressive molecule HLA-G and its clinical implications. Critical Reviews in Clinical Laboratory Sciences. 2012;49:63–84. doi: 10.3109/10408363.2012.677947. [DOI] [PubMed] [Google Scholar]
- 19.Manaster I, Goldman-Wohl D, Greenfield C, et al. MiRNA-mediated control of HLA-G expression and function. PLoS ONE. 2012;7(3) doi: 10.1371/journal.pone.0033395.e33395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Moreau P, Flajollet S, Carosella ED. Non-classical transcriptional regulation of HLA-G: an update. Journal of Cellular and Molecular Medicine. 2009;13(9):2973–2989. doi: 10.1111/j.1582-4934.2009.00800.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kovats S, Main EK, Librach C, Stubblebine M, Fisher SJ, Demars R. A class I antigen, HLA-G, expressed in human trophoblasts. Science. 1990;248(4952):220–223. doi: 10.1126/science.2326636. [DOI] [PubMed] [Google Scholar]
- 22.le Discorde M, Moreau P, Sabatier P, Legeais J-M, Carosella ED. Expression of HLA-G in Human Cornea, an Immune-Privileged Tissue. Human Immunology. 2003;64(11):1039–1044. doi: 10.1016/j.humimm.2003.08.346. [DOI] [PubMed] [Google Scholar]
- 23.Mallet V, Fournel S, Schmitt C, Campan A, Lenfant F, le Bouteiller P. Primary cultured human thymic epithelial cells express both membrane-bound and soluble HLA-G translated products. Journal of Reproductive Immunology. 1999;43(2):225–234. doi: 10.1016/s0165-0378(99)00036-4. [DOI] [PubMed] [Google Scholar]
- 24.Ito T, Ito N, Saathoff M, et al. Immunology of the human nail apparatus: the nail matrix is a site of relative immune privilege. The Journal of Investigative Dermatology. 2005;125(6):1139–1148. doi: 10.1111/j.0022-202X.2005.23927.x. [DOI] [PubMed] [Google Scholar]
- 25.Cirulli V, Zalatan J, McMaster M, et al. The class I HLA repertoire of pancreatic islets comprises the nonclassical class Ib antigen HLA-G. Diabetes. 2006;55(5):1214–1222. doi: 10.2337/db05-0731. [DOI] [PubMed] [Google Scholar]
- 26.Selmani Z, Naji A, Zidi I, et al. Human leukocyte antigen-G5 secretion by human mesenchymal stem cells is required to suppress T lymphocyte and natural killer function and to induce CD4+ CD25highFOXP3+ regulatory T cells. Stem Cells. 2008;26(1):212–222. doi: 10.1634/stemcells.2007-0554. [DOI] [PubMed] [Google Scholar]
- 27.Menier C, Rabreau M, Challier J-C, le Discorde M, Carosella ED, Rouas-Freiss N. Erythroblasts secrete the nonclassical HLA-G molecule from primitive to definitive hematopoiesis. Blood. 2004;104(10):3153–3160. doi: 10.1182/blood-2004-03-0809. [DOI] [PubMed] [Google Scholar]
- 28.Colonna M, Navarro F, Bellón T, et al. A common inhibitory receptor for major histocompatibility complex class I molecules on human lymphoid and myelomonocytic cells. The Journal of Experimental Medicine. 1997;186(11):1809–1818. doi: 10.1084/jem.186.11.1809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Colonna M, Samaridis J, Cella M, et al. Cutting edge: human myelomonocytic cells express an inhibitory receptor for classical and nonclassical MHC class I-molecules. Journal of Immunology. 1998;160(7):3096–3100. [PubMed] [Google Scholar]
- 30.Faure M, Long EO. KIR2DL4 (CD158d), an NK cell-activating receptor with inhibitory potential. Journal of Immunology. 2002;168(12):6208–6214. doi: 10.4049/jimmunol.168.12.6208. [DOI] [PubMed] [Google Scholar]
- 31.Rajagopalan S, Long EO. A human histocompatibility leukocyte antigen (HLA)-G-specific receptor expressed on all natural killer cells. The Journal of Experimental Medicine. 1999;189(7):1093–1099. doi: 10.1084/jem.189.7.1093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fournel S, Aguerre-Girr M, Huc X, et al. Cutting edge: soluble HLA-G1 triggers CD95/CD95 ligand-mediated apoptosis in activated CD8+ cells by interacting with CD8. Journal of Immunology. 2000;164(12):6100–6104. doi: 10.4049/jimmunol.164.12.6100. [DOI] [PubMed] [Google Scholar]
- 33.Gros F, Cabillic F, Tourirais O, le Maux A, Sebti Y, Amiot L. Soluble HLA-G molecules impair natural killer/dendritic cell crosstalk via inhibition of dendritic cells. European Journal of Immunology. 2008;38(3):742–749. doi: 10.1002/eji.200736918. [DOI] [PubMed] [Google Scholar]
- 34.Horuzsko A, Lenfant F, Munn DH, Mellor AL. Maturation of antigen-presenting cells is compromised in HLA-G transgenic mice. International Immunology. 2001;13(3):385–394. doi: 10.1093/intimm/13.3.385. [DOI] [PubMed] [Google Scholar]
- 35.le Friec G, Gros F, Sebti Y, et al. Capacity of myeloid and plasmacytoid dendritic cells especially at mature stage to express and secrete HLA-G molecules. Journal of Leukocyte Biology. 2004;76(6):1125–1133. doi: 10.1189/jlb.0104015. [DOI] [PubMed] [Google Scholar]
- 36.Le Friec G, Laupèze B, Fardel O, et al. Soluble HLA-G inhibits human dendritic cell-triggered allogeneic T-cell proliferation without altering dendritic differentiation and maturation processes. Human Immunology. 2003;64(8):752–761. doi: 10.1016/s0198-8859(03)00091-0. [DOI] [PubMed] [Google Scholar]
- 37.le Gal F-A, Riteau B, Sedlik C, et al. HLA-G-mediated inhibition of antigen-specific cytotoxic T lymphocytes. International Immunology. 1999;11(8):1351–1356. doi: 10.1093/intimm/11.8.1351. [DOI] [PubMed] [Google Scholar]
- 38.Rouas-Freiss N, Gonçalves RM-B, Menier C, Dausset J, Carosella ED. Direct evidence to support the role of HLA-G in protecting the fetus from maternal uterine natural killer cytolysis. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(21):11520–11525. doi: 10.1073/pnas.94.21.11520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Naji A, Menier C, Maki G, Carosella ED, Rouas-Freiss N. Neoplastic B-cell growth is impaired by HLA-G/ILT2 interaction. Leukemia. 2012;26:1889–1892. doi: 10.1038/leu.2012.62. [DOI] [PubMed] [Google Scholar]
- 40.Lila N, Rouas-Freiss N, Dausset J, Carpentier A, Carosella ED. Soluble HLA-G protein secreted by allo-specific CD4+ T cells suppresses the allo-proliferative response: a CD4+ T cell regulatory mechanism. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(21):12150–12155. doi: 10.1073/pnas.201407398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Naji A, Durrbach A, Carosella ED, Rouas-Freiss N. Soluble HLA-G and HLA-G1 expressing antigen-presenting cells inhibit T-cell alloproliferation through ILT-2/ILT-4/fasL-mediated pathways. Human Immunology. 2007;68(4):233–239. doi: 10.1016/j.humimm.2006.10.017. [DOI] [PubMed] [Google Scholar]
- 42.Carosella ED, Gregori S, LeMaoult J. The tolerogenic interplay(s) among HLA-G, myeloid APCs, and regulatory cells. Blood. 2011;118(25):6499–6505. doi: 10.1182/blood-2011-07-370742. [DOI] [PubMed] [Google Scholar]
- 43.LeMaoult J, Caumartin J, Daouya M, et al. Immune regulation by pretenders: cell-to-cell transfers of HLA-G make effector T cells act as regulatory cells. Blood. 2007;109(5):2040–2048. doi: 10.1182/blood-2006-05-024547. [DOI] [PubMed] [Google Scholar]
- 44.Baudhuin J, Migraine J, Faivre V, et al. Exocytosis acts as a modulator of the ILT4-mediated inhibition of neutrophil functions. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:17957–17962. doi: 10.1073/pnas.1221535110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Amiot L, Ferrone S, Grosse-Wilde H, Seliger B. Biology of HLA-G in cancer: a candidate molecule for therapeutic intervention? Cellular and Molecular Life Sciences. 2011;68(3):417–431. doi: 10.1007/s00018-010-0583-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Reddy RC, Chen GH, Tekchandani PK, Standiford TJ. Sepsis-induced immunosuppression: from bad to worse. Immunologic Research. 2001;24(3):273–287. doi: 10.1385/IR:24:3:273. [DOI] [PubMed] [Google Scholar]
- 47.Monneret G, Voirin N, Krawice-Radanne I, et al. Soluble human leukocyte antigen-G5 in septic shock: marked and persisting elevation as a predictor of survival. Critical Care Medicine. 2007;35(8):1942–1947. doi: 10.1097/01.CCM.0000277039.84372.1C. [DOI] [PubMed] [Google Scholar]
- 48.Urbonas V, Eidukaite A, Tamuliene I. The predictive value of soluble biomarkers (CD14 subtype, interleukin-2 receptor, human leucocyte antigen-G) and procalcitonin in the detection of bacteremia and sepsis in pediatric oncology patients with chemotherapy-induced febrile neutropenia. Cytokine. 2013;62:34–37. doi: 10.1016/j.cyto.2013.02.030. [DOI] [PubMed] [Google Scholar]
- 49.Sabbagh A, Courtin D, Milet J, et al. Association of HLA-G 3′ untranslated region polymorphisms with antibody response against Plasmodium falciparum antigens: preliminary results. Tissue Antigens. 2013;82:53–58. doi: 10.1111/tan.12140. [DOI] [PubMed] [Google Scholar]
- 50.Donaghy L, Gros F, Amiot L, et al. Elevated levels of soluble non-classical major histocompatibility class I molecule human leucocyte antigen (HLA)-G in the blood of HIV-infected patients with or without visceral leishmaniasis. Clinical and Experimental Immunology. 2007;147(2):236–240. doi: 10.1111/j.1365-2249.2006.03268.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Gangneux JP, Poinsignon Y, Donaghy L, et al. Indoleamine 2,3-dioxygenase activity as a potential biomarker of immune suppression during visceral leishmaniasis. Innate Immunity. 2013;19:564–568. doi: 10.1177/1753425912473170. [DOI] [PubMed] [Google Scholar]
- 52.Robert-Gangneux F, Gangneux J-P, Vu N, Jaillard S, Guiguen C, Amiot L. High level of soluble HLA-G in amniotic fluid is correlated with congenital transmission of Toxoplasma gondii . Clinical Immunology. 2011;138(2):129–134. doi: 10.1016/j.clim.2010.12.004. [DOI] [PubMed] [Google Scholar]
- 53.Zhao M, Zhang R, Xu X, et al. IL-10 reduces levels of apoptosis in Toxoplasma gondii-infected trophoblasts. PloS ONE. 2013;8 doi: 10.1371/journal.pone.0056455.e56455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Murdaca G, Contini P, Setti M, et al. Behavior of non-classical soluble HLA class G antigens in human immunodeficiency virus 1-infected patients before and after HAART: comparison with classical soluble HLA-A, -B, -C antigens and potential role in immune-reconstitution. Clinical Immunology. 2009;133(2):238–244. doi: 10.1016/j.clim.2009.08.002. [DOI] [PubMed] [Google Scholar]
- 55.Huang J, Burke P, Yang Y, et al. Soluble HLA-G inhibits myeloid dendritic cell function in HIV-1 infection by interacting with leukocyte immunoglobulin-like receptor B2. Journal of Virology. 2010;84(20):10784–10791. doi: 10.1128/JVI.01292-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cabello A, Rivero A, Garcia MJ, et al. HAART induces the expression of HLA-G on peripheral monocytes in HIV-1 infected individuals. Human Immunology. 2003;64(11):1045–1049. doi: 10.1016/j.humimm.2003.08.353. [DOI] [PubMed] [Google Scholar]
- 57.Lozano JM, González R, Kindelán JM, et al. Monocytes and T lymphocytes in HIV-1-positive patients express HLA-G molecule. AIDS. 2002;16(3):347–351. doi: 10.1097/00002030-200202150-00005. [DOI] [PubMed] [Google Scholar]
- 58.Rivero A, Lozano JM, González R, et al. Nucleoside reverse transcriptase inhibitors are able and protease inhibitors unable to induce the tolerogenic molecule HLA-G1 on monocytes from HIV-1 infected patients. Human Immunology. 2007;68(4):303–306. doi: 10.1016/j.humimm.2006.10.014. [DOI] [PubMed] [Google Scholar]
- 59.Murdaca G, Contini P, Setti M, et al. Soluble human leukocyte antigen-G serum levels in patients with acquired immune deficiency syndrome affected by different disease-defining conditions before and after antiretroviral treatment. Human Immunology. 2011;72(9):712–716. doi: 10.1016/j.humimm.2011.05.008. [DOI] [PubMed] [Google Scholar]
- 60.Li C, Toth I, Schulze zur Wiesch J, et al. Functional characterization of HLA-G(+) regulatory T cells in HIV-1 infection. PLoS Pathogens. 2013;9 doi: 10.1371/journal.ppat.1003140.e1003140 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Onno M, Pangault C, le Friec G, Guilloux V, André P, Fauchetz R. Modulation of HLA-G antigens expression by human cytomegalovirus: specific induction in activated macrophages harboring human cytomegalovirus infection. Journal of Immunology. 2000;164(12):6426–6434. doi: 10.4049/jimmunol.164.12.6426. [DOI] [PubMed] [Google Scholar]
- 62.Yan W-H, Lin A, Chen B-G, Chen S-Y. Induction of both membrane-bound and soluble HLA-G expression in active human cytomegalovirus infection. The Journal of Infectious Diseases. 2009;200(5):820–826. doi: 10.1086/604733. [DOI] [PubMed] [Google Scholar]
- 63.Lafon M, Prehaud C, Megret F, et al. Modulation of HLA-G expression in human neural cells after neurotropic viral infections. Journal of Virology. 2005;79(24):15226–15237. doi: 10.1128/JVI.79.24.15226-15237.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Mégret F, Prehaud C, Lafage M, et al. Modulation of HLA-G and HLA-E expression in human neuronal cells after rabies virus or herpes virus simplex type 1 infections. Human Immunology. 2007;68(4):294–302. doi: 10.1016/j.humimm.2006.12.003. [DOI] [PubMed] [Google Scholar]
- 65.LeBouder F, Khoufache K, Menier C, et al. Immunosuppressive HLA-G molecule is upregulated in alveolar epithelial cells after influenza A virus infection. Human Immunology. 2009;70(12):1016–1019. doi: 10.1016/j.humimm.2009.07.026. [DOI] [PubMed] [Google Scholar]
- 66.Chen H-X, Chen B-G, Shi W-W, et al. Induction of cell surface human leukocyte antigen-G expression in pandemic H1N1 2009 and seasonal H1N1 influenza virus-infected patients. Human Immunology. 2011;72(2):159–165. doi: 10.1016/j.humimm.2010.11.009. [DOI] [PubMed] [Google Scholar]
- 67.Guimarães MCM, Soares CP, Donadi EA, et al. Low expression of human histocompatibility soluble leukocyte antigen-G (HLA-G5) in invasive cervical cancer with and without metastasis, associated with papilloma virus (HPV) The Journal of Histochemistry and Cytochemistry. 2010;58(5):405–411. doi: 10.1369/jhc.2009.954131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Shi W-W, Lin A, Xu D-P, et al. Plasma soluble human leukocyte antigen-G expression is a potential clinical biomarker in patients with hepatitis B virus infection. Human Immunology. 2011;72(11):1068–1073. doi: 10.1016/j.humimm.2011.06.012. [DOI] [PubMed] [Google Scholar]
- 69.Weng P-J, Fu Y-M, Ding S-X, Xu D-P, Lin A, Yan W-H. Elevation of plasma soluble human leukocyte antigen-G in patients with chronic hepatitis C virus infection. Human Immunology. 2011;72(5):406–411. doi: 10.1016/j.humimm.2011.02.008. [DOI] [PubMed] [Google Scholar]
- 70.Souto FJD, Crispim JCO, Ferreira SC, et al. Liver HLA-G expression is associated with multiple clinical and histopathological forms of chronic hepatitis B virus infection. Journal of Viral Hepatitis. 2011;18(2):102–105. doi: 10.1111/j.1365-2893.2010.01286.x. [DOI] [PubMed] [Google Scholar]
- 71.Amiot L, Vu N, Rauch M, et al. Expression of HLA-G by mast cells is associated with hepatitis C virus-induced liver fibrosis. Journal of Hepatology. 2014;60:245–252. doi: 10.1016/j.jhep.2013.09.006. [DOI] [PubMed] [Google Scholar]
- 72.Matte C, Lajoie J, Lacaille J, Zijenah LS, Ward BJ, Roger M. Functionally active HLA-G polymorphisms are associated with the risk of heterosexual HIV-1 infection in African women. AIDS. 2004;18(3):427–431. doi: 10.1097/00002030-200402200-00008. [DOI] [PubMed] [Google Scholar]
- 73.Lajoie J, Hargrove J, Zijenah LS, Humphrey JH, Ward BJ, Roger M. Genetic variants in nonclassical major histocompatibility complex class I human leukocyte antigen (HLA)-E and HLA-G molecules are associated with susceptibility to heterosexual acquisition of HIV-1. The Journal of Infectious Diseases. 2006;193(2):298–301. doi: 10.1086/498877. [DOI] [PubMed] [Google Scholar]
- 74.Segat L, Catamo E, Fabris A, et al. HLA-G*0105N allele is associated with augmented risk for HIV infection in white female patients. AIDS. 2010;24(12):1961–1964. doi: 10.1097/QAD.0b013e32833c3324. [DOI] [PubMed] [Google Scholar]
- 75.da Silva GK, Vianna P, Veit TD, et al. Influence of HLA-G polymorphisms in human immunodeficiency virus infection and hepatitis C virus co-infection in Brazilian and Italian individuals. Infection, genetics and evolution. Journal of Molecular Epidemiology and Evolutionary Genetics in Infectious Diseases. 2014;21:418–423. doi: 10.1016/j.meegid.2013.12.013. [DOI] [PubMed] [Google Scholar]
- 76.Aikhionbare FO, Kumaresan K, Shamsa F, Bond VC. HLA-G DNA sequence variants and risk of perinatal HIV-I transmission. AIDS Research and Therapy. 2006;3(1, article 28) doi: 10.1186/1742-6405-3-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Luo M, Czarnecki C, Ramdahin S, Embree J, Plummer FA. HLA-G and mother-child perinatal HIV transmission. Human Immunology. 2013;74:459–463. doi: 10.1016/j.humimm.2012.11.023. [DOI] [PubMed] [Google Scholar]
- 78.Castelli EC, Moreau P, Chiromatzo AOE, et al. In silico analysis of microRNAS targeting the HLA-G 3′ untranslated region alleles and haplotypes. Human Immunology. 2009;70(12):1020–1025. doi: 10.1016/j.humimm.2009.07.028. [DOI] [PubMed] [Google Scholar]
- 79.Cordero EAA, Veit TD, da Silva MAL, Jacques SMC, Silla LMDR, Chies JAB. HLA-G polymorphism influences the susceptibility to HCV infection in sickle cell disease patients. Tissue Antigens. 2009;74(4):308–313. doi: 10.1111/j.1399-0039.2009.01331.x. [DOI] [PubMed] [Google Scholar]
- 80.Martinetti M, Pacati I, Cuccia M, et al. Hierarchy of baby-linked immunogenetic risk factors in the vertical transmission of hepatitis C virus. International Journal of Immunopathology and Pharmacology. 2006;19(2):369–378. doi: 10.1177/039463200601900213. [DOI] [PubMed] [Google Scholar]
- 81.Zheng X-Q, Zhu F, Shi W-W, Lin A, Yan W-H. The HLA-G 14 bp insertion/deletion polymorphism is a putative susceptible factor for active human cytomegalovirus infection in children. Tissue Antigens. 2009;74(4):317–321. doi: 10.1111/j.1399-0039.2009.01312.x. [DOI] [PubMed] [Google Scholar]
- 82.Bortolotti D, Gentili V, Rotola A, di Luca D, Rizzo R. Implication of HLA-G 3′ untranslated region polymorphisms in human papillomavirus infection. Tissue Antigens. 2014;83:113–118. doi: 10.1111/tan.12281. [DOI] [PubMed] [Google Scholar]
- 83.Garcia A, Milet J, Courtin D, et al. Association of HLA-G 3′UTR polymorphisms with response to malaria infection: a first insight. Infection, genetics and evolution. Journal of Molecular Epidemiology and Evolutionary Genetics in Infectious Diseases. 2013;16:263–269. doi: 10.1016/j.meegid.2013.02.021. [DOI] [PubMed] [Google Scholar]
- 84.Barel MT, Ressing M, Pizzato N, et al. Human cytomegalovirus-encoded US2 differentially affects surface expression of MHC class I locus products and targets membrane-bound, but not soluble HLA-G1 GL for degradation. Journal of Immunology. 2003;171(12):6757–6765. doi: 10.4049/jimmunol.171.12.6757. [DOI] [PubMed] [Google Scholar]
- 85.Pizzato N, Derrien M, Lenfant F. The short cytoplasmic tail of HLA-G determines its resistance to HIV-1 Nef-mediated cell surface downregulation. Human Immunology. 2004;65(11):1389–1396. doi: 10.1016/j.humimm.2004.07.239. [DOI] [PubMed] [Google Scholar]
- 86.Schust DJ, Tortorella D, Seebach J, et al. Trophoblast class I major histocompatibility complex (MHC)4 products are resistant to rapid degradation imposed by the human cytomegalovirus (HCMV) gene products US2 and US11. The Journal of Experimental Medicine. 1998;188:497–503. doi: 10.1084/jem.188.3.497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Jun Y, Kim E, Jin M, et al. Human cytomegalovirus gene products US3 and US6 down-regulate trophoblast class I MHC molecules. Journal of Immunology. 2000;164(2):805–811. doi: 10.4049/jimmunol.164.2.805. [DOI] [PubMed] [Google Scholar]
- 88.Park B, Spooner E, Houser BL, Strominger JL, Ploegh HL. The HCMV membrane glycoprotein US10 selectively targets HLA-G for degradation. The Journal of Experimental Medicine. 2010;207(9):2033–2041. doi: 10.1084/jem.20091793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Pizzato N, Garmy-Susini B, le Bouteiller P, Lenfant F. Down-regulation of HLA-G1 cell surface expression in human cytomegalovirus infected cells. American Journal of Reproductive Immunology. 2003;50(4):328–333. doi: 10.1034/j.1600-0897.2003.00087.x. [DOI] [PubMed] [Google Scholar]
- 90.Schust DJ, Hill AB, Ploegh HL. Herpes simplex virus blocks intracellular transport of HLA-G in placentally derived human cells. Journal of Immunology. 1996;157(8):3375–3380. [PubMed] [Google Scholar]
- 91.Pizzato N, Garmy-Susini B, Bouteiller PL, Lenfant F. Differential down-modulation of HLA-G and HLA-A2 or -A3 cell surface expression following human cytomegalovirus infection. Journal of Reproductive Immunology. 2004;62(1-2):3–15. doi: 10.1016/j.jri.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 92.le Gall S, Buseyne F, Trocha A, Walker BD, Heard J-M, Schwartz O. Distinct trafficking pathways mediate Nef-induced and clathrin-dependent major histocompatibility complex class I down-regulation. Journal of Virology. 2000;74(19):9256–9266. doi: 10.1128/jvi.74.19.9256-9266.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ouyang W, Rutz S, Crellin NK, Valdez PA, Hymowitz SG. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annual Review of Immunology. 2011;29:71–109. doi: 10.1146/annurev-immunol-031210-101312. [DOI] [PubMed] [Google Scholar]
- 94.Marie C, Muret J, Fitting C, Losser M-R, Payen D, Cavaillon J-M. Reduced ex vivo interleukin-8 production by neutrophils in septic and nonseptic systemic inflammatory response syndrome. Blood. 1998;91(9):3439–3446. [PubMed] [Google Scholar]
- 95.Urbonas V, Eidukaitë A, Tamulienë I. Increased interleukin-10 levels correlate with bacteremia and sepsis in febrile neutropenia pediatric oncology patients. Cytokine. 2012;57(3):313–315. doi: 10.1016/j.cyto.2011.11.012. [DOI] [PubMed] [Google Scholar]
- 96.Moore KW, de Waal Malefyt R, Coffman RL, O’Garra A. Interleukin-10 and the interleukin-10 receptor. Annual Review of Immunology. 2001;19:683–765. doi: 10.1146/annurev.immunol.19.1.683. [DOI] [PubMed] [Google Scholar]
- 97.Moreau P, Adrian-Cabestre F, Menier C, et al. IL-10 selectively induces HLA-G expression in human trophoblasts and monocytes. International Immunology. 1999;11(5):803–811. doi: 10.1093/intimm/11.5.803. [DOI] [PubMed] [Google Scholar]
- 98.Yang Y, Geraghty DE, Hunt JS. Cytokine regulation of HLA-G expression in human trophoblast cell lines. Journal of Reproductive Immunology. 1995;29(3):179–195. doi: 10.1016/0165-0378(95)00942-e. [DOI] [PubMed] [Google Scholar]
- 99.Onno M, le Friec G, Pangault C, et al. Modulation of HLA-G antigens expression in myelomonocytic cells. Human Immunology. 2000;61(11):1086–1094. doi: 10.1016/s0198-8859(00)00191-9. [DOI] [PubMed] [Google Scholar]
- 100.Zilberman S, Schenowitz C, Agaugue S, et al. HLA-G1 and HLA-G5 active dimers are present in malignant cells and effusions: the influence of the tumor microenvironment. European Journal of Immunology. 2012;42:1599–1608. doi: 10.1002/eji.201141761. [DOI] [PubMed] [Google Scholar]
- 101.Ugurel S, Rebmann V, Ferrone S, Tilgen W, Grosse-Wilde H, Reinhold U. Soluble human leukocyte antigen—G serum level is elevated in melanoma patients and is further increased by interferon-alpha immunotherapy. Cancer. 2001;92:369–376. doi: 10.1002/1097-0142(20010715)92:2<369::aid-cncr1332>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- 102.Mitsdoerffer M, Schreiner B, Kieseier BC, et al. Monocyte-derived HLA-G acts as a strong inhibitor of autologous CD4 T cell activation and is upregulated by interferon-βin vitro and in vivo: rationale for the therapy of multiple sclerosis. Journal of Neuroimmunology. 2005;159(1-2):155–164. doi: 10.1016/j.jneuroim.2004.09.016. [DOI] [PubMed] [Google Scholar]
- 103.Lefebvre S, Berrih-Aknin S, Adrian F, et al. A specific interferon (IFN)-stimulated response element of the distal HLA-G promoter binds IFN-regulatory factor 1 and mediates enhancement of this nonclassical class I gene by IFN-β . Journal of Biological Chemistry. 2001;276(9):6133–6139. doi: 10.1074/jbc.M008496200. [DOI] [PubMed] [Google Scholar]
- 104.Gupta P, Ghosh S, Nagarajan A, Mehta VS, Sen E. Beta-defensin-3 negatively regulates TLR4-HMGB1 axis mediated HLA-G expression in IL-1beta treated glioma cells. Cellular Signalling. 2013;25:682–689. doi: 10.1016/j.cellsig.2012.12.001. [DOI] [PubMed] [Google Scholar]
- 105.Gruber BL, Kew RR, Jelaska A, et al. Human mast cells activate fibroblasts: tryptase is a fibrogenic factor stimulating collagen messenger ribonucleic acid synthesis and fibroblast chemotaxis. Journal of Immunology. 1997;158(5):2310–2317. [PubMed] [Google Scholar]
- 106.Marvie P, Lisbonne M, L’Helgoualc’h A, et al. Interleukin-33 overexpression is associated with liver fibrosis in mice and humans. Journal of Cellular and Molecular Medicine. 2010;14(6 B):1726–1739. doi: 10.1111/j.1582-4934.2009.00801.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Allakhverdi Z, Smith DE, Comeau MR, Delespesse G. Cutting edge: the ST2 ligand IL-33 potently activates and drives maturation of human mast cells. Journal of Immunology. 2007;179(4):2051–2054. doi: 10.4049/jimmunol.179.4.2051. [DOI] [PubMed] [Google Scholar]
- 108.Iikura M, Suto H, Kajiwara N, et al. IL-33 can promote survival, adhesion and cytokine production in human mast cells. Laboratory Investigation. 2007;87(10):971–978. doi: 10.1038/labinvest.3700663. [DOI] [PubMed] [Google Scholar]
- 109.Gruber BL, Marchese MJ, Kew RR. Transforming growth factor-β1 mediates mast cell chemotaxis. Journal of Immunology. 1994;152(12):5860–5867. [PubMed] [Google Scholar]
- 110.Hsu C-L, Neilsen CV, Bryce PJ. IL-33 is produced by mast cells and regulates IgE-dependent inflammation. PLoS ONE. 2010;5(8) doi: 10.1371/journal.pone.0011944.e11944 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Grimbaldeston MA, Nakae S, Kalesnikoff J, Tsai M, Galli SJ. Mast cell-derived interleukin 10 limits skin pathology in contact dermatitis and chronic irradiation with ultraviolet B. Nature Immunology. 2007;8(10):1095–1104. doi: 10.1038/ni1503. [DOI] [PubMed] [Google Scholar]
- 112.Morandi F, Ferretti E, Castriconi R, et al. Soluble HLA-G dampens CD94/NKG2A expression and function and differentially modulates chemotaxis and cytokine and chemokine secretion in CD56bright and CD56dim NK cells. Blood. 2011;118(22):5840–5850. doi: 10.1182/blood-2011-05-352393. [DOI] [PubMed] [Google Scholar]
- 113.Wuest TR, Carr DJJ. Dysregulation of CXCR3 signaling due to CXCL10 deficiency impairs the antiviral response to herpes simplex virus 1 infection. Journal of Immunology. 2008;181(11):7985–7993. doi: 10.4049/jimmunol.181.11.7985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Agaugué S, Carosella ED, Rouas-Freiss N. Role of HLA-G in tumor escape through expansion of myeloid-derived suppressor cells and cytokinic balance in favor of Th2 versus Th1/Th17. Blood. 2011;117(26):7021–7031. doi: 10.1182/blood-2010-07-294389. [DOI] [PubMed] [Google Scholar]
- 115.Lin A, Zhang X, Xu H-H, Xu D-P, Ruan Y-Y, Yan W-H. HLA-G expression is associated with metastasis and poor survival in the Balb/c nu/nu murine tumor model with ovarian cancer. International Journal of Cancer. 2012;131(1):150–157. doi: 10.1002/ijc.26375. [DOI] [PubMed] [Google Scholar]
- 116.LeMaoult J, Daouya M, Wu J, Loustau M, Horuzsko A, Carosella ED. Synthetic HLA-G proteins for therapeutic use in transplantation. FASEB Journal. 2013;27:3643–3651. doi: 10.1096/fj.13-228247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Celsi F, Catamo E, Kleiner G, Tricarico PM, Vuch J, Crovella S. HLA-G/C, miRNAs, and their role in HIV infection and replication. BioMed Research International. 2013;2013:13 pages. doi: 10.1155/2013/693643.693643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Sartelet H, Schleiermacher D, Le-Hesran JY, et al. Less HLA-G expression in Plasmodium falciparum-infected third trimester placentas is associated with more natural killer cells. Placenta. 2005;26(6):505–511. doi: 10.1016/j.placenta.2004.08.006. [DOI] [PubMed] [Google Scholar]
- 119.Derrien M, Pizzato N, Dolcini G, et al. Human immunodeficiency virus 1 downregulates cell surface expression of the non-classical major histocompatibility class I molecule HLA-G1. The Journal of General Virology. 2004;85(7):1945–1954. doi: 10.1099/vir.0.79867-0. [DOI] [PubMed] [Google Scholar]
- 120.Lajoie J, Fontaine J, Tremblay C, Routy J-P, Poudrier J, Roger M. Persistence of high levels of blood soluble human leukocyte antigen-G is associated with rapid progression of HIV infection. AIDS. 2009;23(11):1437–1440. doi: 10.1097/QAD.0b013e32832d0825. [DOI] [PubMed] [Google Scholar]
- 121.de Oliveira Crispim JC, Silva TGA, Souto FJD, et al. Upregulation of soluble and membrane-bound human leukocyte antigen G expression is primarily observed in the milder histopathological stages of chronic hepatitis C virus infection. Human Immunology. 2012;73(3):258–262. doi: 10.1016/j.humimm.2011.12.004. [DOI] [PubMed] [Google Scholar]
- 122.Turk WJ, Kimani J, Bielawny T, et al. Associations of human leukocyte antigen-G with resistance and susceptibility to HIV-1 infection in the Pumwani sex worker cohort. AIDS. 2013;27:7–15. doi: 10.1097/QAD.0b013e32835ab1f2. [DOI] [PubMed] [Google Scholar]