

Abstract
Background
Infection with BK virus (BKV) generally occurs early during life, but its mode of transmission has not been clearly defined. We tested the hypothesis that polyomavirus shedding in stool may be a source of BKV exposure.
Methods
Pediatric stool and rectal swab samples were tested for the presence of polyomavirus DNA by a polymerase chain reaction (PCR) assay that could detect a conserved region in the large T antigen gene of BKV, JC virus (JCV), and simian virus 40 (SV40). The specific viruses detected by this assay were confirmed by DNA sequence analysis of the PCR amplicons.
Results
Of 120 samples collected from 99 patients, 54 (45.0%) were positive for polyomavirus DNA. Of the 99 patients, 46 (46.5%) had at least 1 positive sample, with 38 (38.4%) positive for BKV and 8 (8.1%) positive for SV40. JCV was not detected. There was no association between polyomavirus fecal shedding and age, sex, race/ ethnicity, immune status, or symptoms of gastrointestinal disease in the children studied. The BKV strains detected displayed polymorphisms in the T antigen sequence.
Conclusions
Polyomaviruses are frequently present in stool samples from hospitalized children. These findings suggest that fecal-oral transmission of BKV may play a role in the ubiquity of infection.
Seroprevalence studies have documented that infection with the human polyomavirus BK virus (BKV) commonly occurs during the first decade of life [1, 2]. In contrast, JC virus (JCV) infection typically occurs during the second decade of life [2–4]. The reasons for this difference in the epidemiologic profiles of these infections are unknown but may be related to differences in the routes of transmission. The kidney is believed to be the primary reservoir for both JCV and BKV, and JCV is commonly excreted in the urine of adults—especially of those >40 years old—whereas BKV is not. Importantly, a source of polyomavirus exposure that would explain the high incidence of BKV infection during early childhood has remained elusive. Among viruses of the family Polyomaviridae that infect non-humans, the urinary tract, the respiratory tract, and the gastrointestinal tract are the principal sites of disease and are the presumed sources of exposure leading to transmission [5–7].
Although infection with BKV is common, disease due to BKV has been recognized exclusively in the setting of severe immune compromise, often in association with BKV viruria and viremia [8–10], making it unlikely that those with BKV disease are the source of exposure that would explain the high frequency of transmission early during life. Further, numerous studies have failed to detect BKV in the urine of healthy children and adults at a frequency that might account for the ubiquity of early childhood infection [11–13]. BKV excretion in the urine occurs in 5%–50% of pregnant women [8, 14–17], and, although transplacental transmission of BKV has been reported, this route of transmission is believed to be rare [18–22]. A study found that BKV excretion typically ceases shortly after parturition in women with pregnancy-associated reactivation, although this study did not use polymerase chain reaction (PCR) testing [23].
Respiratory tract tissues and secretions have been studied as a potential source of polyomavirus exposure. However, these studies failed to detect significant polyomavirus shedding—in one study, 5 of 12 tonsillar tissue samples were positive for BKV by Southern blot, but no infectious virus was detected [24]; in another study, only 2 of 201 pediatric nasopharyngeal aspirates were positive for BKV when tested by a sensitive PCR/ Southern blot procedure [25].
The available data on polyomaviruses in the gastrointestinal tract of humans is limited. Simian virus 40 (SV40) shedding in stool has been reported in infants after receipt of SV40-contaminated polio vaccine [26]. More recently, JCV DNA was detected in 26% of colon cancer biopsy samples and adjacent normal mucosa [27–29]. Quantitative PCR studies performed in regions throughout the world have detected JCV and BKV in sewage effluent, again pointing to human excrement or urine as possible sources of polyomavirus exposure [30, 31]. We tested stool and rectal swab samples from hospitalized children, to determine whether stool may be a source of BKV exposure that could account for the high frequency of BKV transmission in childhood.
PATIENTS, MATERIALS, AND METHODS
Sample collection and processing
Archived stool or rectal swab samples that had been submitted to the Diagnostic Virology Laboratory of Texas Children’s Hospital (Houston) for viral culture and that were stored at −70°C in viral transport medium (veal infusion broth in dH2O plus gentamicin and amphotericin) were identified via the laboratory computer database and retrieved. Stool and rectal swab samples were not distinguished from one another with respect to labeling, processing, or reporting in the laboratory database. Samples were thawed, and aliquots (200 μL) of each sample were transferred to the research laboratory at Baylor College of Medicine and stored at 4°C until all PCR testing was complete (1–2 days). Sample identification included the laboratory accession number, the date of collection, and the patient’s date of birth. Strict measures for avoidance of cross-contamination were maintained throughout the study, including the use of separate, plasmid-free rooms and centrifuges for processing of clinical samples, PCR setup, and PCR analysis.
PCR screening and DNA sequence determination
Samples were diluted 1:10 in sterile PBS, were boiled for 10 min, and were clarified by low-speed centrifugation before being used as templates for PCR amplifications. Ten microliters of each diluted sample was used as the template in PCR amplifications, with a total volume of 50 μL. All samples were tested with the primers PYV.for and PYV.rev [32], which amplify a 178–183-bp region in the amino terminus of the T antigen gene of JCV, BKV, and SV40. Some samples were tested with the primers AG1 and AG2, to assess the presence of human cellular DNA [33], and with the primers F-amp and R-amp, to assess the presence of bacterial DNA or plasmid contamination. Conditions for PCR amplifications and primer sequences have been published elsewhere [12]. To assess the possible presence of PCR inhibitors in diluted stool preparations, BKV-containing plasmid DNA (Dunlop strain) was added to some diluted samples, and these were used as the template for PCR amplification with the primers PYV.for and PYV.rev.
PCR products that were similar in size to the positive control amplimers (178–183 bp) produced with the primers PYV.for and PYV.rev, as determined by direct visualization on ethidium bromide–stained agarose gel, were treated with shrimp alkaline phosphatase (Amersham) and exonuclease I (New England Bio-labs) for 45 min at 37°C, followed by enzyme inactivation at 85°C for 10 min. DNA sequencing was performed by Lone Star Labs (Houston, TX) by use of the automated DNA sequencer ABI Prism 377XL and the BigDye Terminator Ready Reaction Cycle Sequencing Kit (Applied Biosystems), in accordance with the manufacturer’s directions. Consensus viral sequences were aligned and compared with known polyomavirus sequences by use of ClustalW (version 1.8; available at: http://www.embl.org) and Gene Runner (version 3.05; Hastings Software). As described elsewhere [8, 12], additional PCR primers were used to attempt amplification of the noncoding control region (NCCR) and portions of the VP1 gene from samples found to be positive for BKV.
Retrospective medical record review
To assess the demographic and medical factors that may be associated with polyomavirus excretion in this hospitalized pediatric population, we obtained approval from the Institutional Review Board for Human Subject Research to conduct a retrospective review of the medical records of patients whose stool or rectal swab samples were tested. Specific data (sex, race/ethnicity, zip code, admission and discharge diagnoses, gastrointestinal symptoms, evidence of immune compromise, complete blood cell counts, blood urea nitrogen levels, and serum creatinine levels) were sought from each chart by use of a standardized electronic data-collection form. The following were considered to be evidence of immune compromise: iatrogenic immune suppression, diagnosis of primary or secondary immune deficiency, absolute neutropenia or lymphopenia (<500 cells/μL), absolute leukopenia (<2500 white blood cells/μL), hepatic insufficiency, and renal insufficiency requiring dialysis.
RESULTS
Viral detection in stool samples
Search of the Diagnostic Virology Laboratory database identified 158 stool or rectal swab samples that had been sent for viral culture during the period from 1 October 2003 to 31 January 2004. One hundred twenty samples were randomly retrieved. When tested with the universal polyomavirus primers (PYV.for and PYV.rev), 54 (45.0%) samples yielded an amplicon of the appropriate size (figure 1A) that, by DNA sequence determination, contained unequivocal polyomavirus sequence. The DNA sequence analysis revealed that, of these 54 stool or rectal swab samples, 46 (85.2%) were positive for BKV and 8 (14.8%) were positive for SV40. JCV was not detected. Human cellular DNA was not detected in any of the 40 stool samples tested with the primers AG1 and AG2 (data not shown). When BKV-containing plasmid DNA was added to 3 samples that were negative for polyomavirus sequences, amplification of BKV sequences was accomplished (data not shown), suggesting that PCR inhibitors were not frequently present in the boiled stool preparations.
Figure 1.
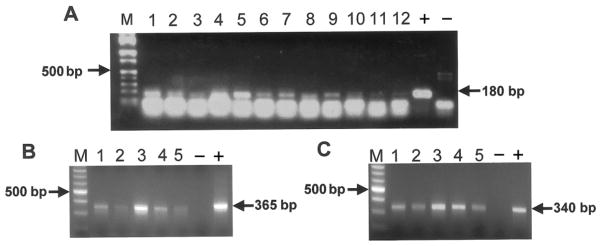
A, Ethidium bromide–stained agarose gel of polymerase chain reaction (PCR) amplicons detected with the primers PYV.for and PYV.rev. Shown are the marker lane (M), products from 12 representative PCR amplifications (lanes 1–12), the positive control (+), and the negative control (−). B and C, Ethidium bromide–stained agarose gel of PCR amplicons from the noncoding control region (B) and VP1 gene (C) of BK virus. Shown are the marker lanes (M), products from 5 representative PCR amplifications (lanes 1–5), the positive controls (+), and the negative controls (−). Note that the amplifications shown in panels B and C are from the same samples, which differ from those depicted in panel A.
Analysis of viral sequences
In addition to establishing the identity of the polyomaviruses detected with the primers PYV.for and PYV.rev, DNA sequence analysis of PCR amplicons revealed sequence heterogeneity among the BKV strains detected. Single-nucleotide polymorphisms were detected at 5 positions within the BKV amplicon (figure 2). To our knowledge, sequence heterogeneity in this genomic region of BKV has not been previously described. Among the SV40 strains detected (n =8), there were no nucleotide changes within the PYV.for/PYV.rev amplicon. This finding agrees with the 14 SV40 sequences in GenBank, each of which shows conservation at the nucleotide level in this genomic region.
Figure 2.
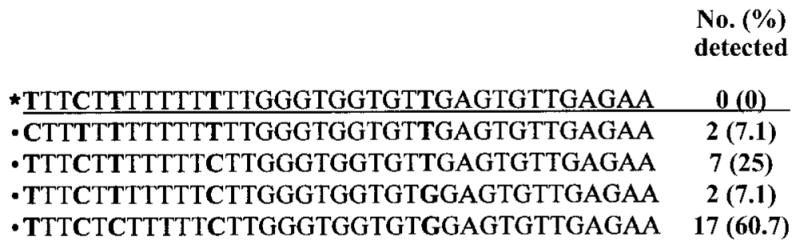
Sequence alignment of data from the PYV.for/PYV.rev amplicon from the amino terminus of the BK virus (BKV) T antigen gene and frequency of detection (in independent samples) of BKV nucleotide polymorphisms within that sequence. Data are from 28 patients. *BKV Dunlop strain, 4412–4447 nt.
None of the BKV strains detected in the stool samples were identical to the laboratory strain (figure 2). Although stringent precautions had been taken to avoid inadvertent laboratory contamination with polyomavirus-containing plasmid DNA, we addressed the possibility of plasmid contamination by testing each sample with the primers F-amp and R-amp, which amplify a 350-bp region of the ampicillin-resistance gene (AmpR) that is found in all polyomavirus-containing plasmids in our laboratory. Of the 120 samples, 5 (4.2%) produced amplicons of the expected size. Two of these 5 samples were repeatedly negative when tested with the primers PYV.for and PYV.rev, excluding a polyomavirus plasmid as the DNA template. Three samples were AmpR positive and PYV.for/PYV.rev positive. DNA sequence data established that the BKV sequences detected differed from those of the laboratory strain, BKV Dunlop. Repeat PCR assays were performed on a second dilution of the 3 original stool or rectal swab samples, and identical results were obtained. During the study period, 60% of nonurinary Escherichia coli isolates from Texas Children’s Hospital were reported to be ampicillin resistant, suggesting that the 5 stool samples that were positive for the ampicillin-resistance gene could have been contaminated by bacterial DNA.
PCR detection of other regions of the BKV genome
Additional regions of the BKV genome, including the NCCR and a 340-bp region of the VP1 gene, were amplified from 5 samples (figure 1B and 1C). These samples had PYV.for/PYV.rev amplicons that were particularly bright on ethidium bromide–stained agarose gel (e.g., lanes 1 and 5 in figure 1A), suggesting a higher titer of virus in the original sample. Amplification of the BKV NCCR and VP1 gene from 8 samples whose PYV.for/ PYV.rev amplicons had a weaker signal was unsuccessful (data not shown). This result is consistent with our experience that the primers PYV.for and PYV.rev are more sensitive than the primers for other regions of the BKV genome.
Duration of BKV shedding in stool
Multiple samples had been collected from several children in the present study on different dates. Of the 11 patients with 2 samples, 3 were found to be excreting BKV on both occasions, with a range of 3–21 days separating the collection of samples. Of the 4 patients with ≥3 samples collected on different dates, 1 was found to be excreting BKV on 3 occasions, with the interval between the collection of the first and last samples being 14 days. One patient who had polyomavirus detectable in 3 of 3 samples collected over the course of 22 days was found to be excreting BKV in the first sample, SV40 in the second sample, and, possibly, both BKV and SV40 in the third sample. The interval between the collection of the first and second sample was 10 days, and the interval between the collection of the second and third sample was 12 days. Coexcretion of SV40 and BKV was suspected because the DNA sequence chromogram appeared to reflect the presence of 2 related DNA templates, but this could not be confirmed. There was no DNA sequence variation among polyomavirus strains detected in the same patient at different sample-collection time points.
Demographics of patients shedding polyomaviruses in stool
Using available medical records, we were able to confirm the unique identities of the sources of samples with different dates of birth for each of the 120 samples studied. Polyomavirus DNA was detected in the stool of 46 (46.5%) of 99 patients, with 38 (38.4%) positive for BKV and 8 (8.1%) positive for SV40. The mean age of those excreting polyomaviruses was 4.51 years (range, 0.03–17.2 years), which was not statistically different from the mean age of those not excreting polyomaviruses (5.14 years; range, 0.03–8.3 years) (P =.71; 2-tailed Student’s t test, assuming unequal variances). Analysis of the age distribution of children with detectable polyomavirus excretion revealed a prevalence of 48.2% (27/56) among patients 0–2 years old, 42.9% (6/14) among patients 3–6 years old, 58.3% (7/12) among patients 7–11 years old, and 35.3% (6/ 17) among patients 12–17 years old (figure 3). There was no statistical difference among these subgroups (P =.64, χ2 test). Similarly, there was no statistical difference in the mean age of those patients found to be excreting SV40 and those found not to be excreting SV40 (3.77 vs. 4.29 years; P =.46, 2-tailed Student’s t test).
Figure 3.
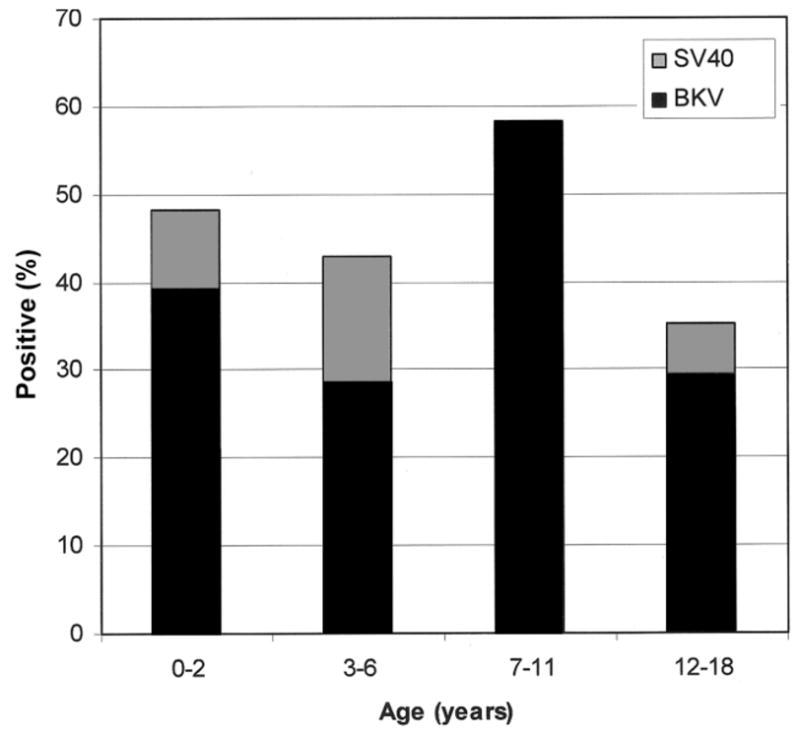
Age distribution of patients with stool samples positive for polyomavirus (n =46). BKV, BK virus; SV40, simian virus 40.
Complete medical records were available for 92 (92.9%) of the 99 patients, allowing a more-thorough exploration of the demographic and clinical characteristics that potentially influence polyomavirus excretion (table 1). There was no statistical difference in the distribution of polyomaviruses among the different race/ethnicity categories (χ2 =0.54; P =.96). Similarly, there was no statistical difference in the excretion of BKV or SV40 among the race/ethnicity categories (for BKV, P =.68; for SV40, P =.22). Polyomavirus excretion in general and BKV excretion alone were no more common in male patients than in female patients (for all polyomaviruses, P =.83; for BKV, P =.24). In contrast, SV40-excreting patients were more likely to be male than female (7/45 male patients vs. 1/54 female patients; P =.022). There was no difference in polyomavirus excretion between the patients with and the patients without immune compromise (21/41 with immune compromise vs. 25/ 58 without immune compromise; P =.33). Neither BKV nor SV40 excretion was statistically associated with immune compromise (for BKV, P =.89; for SV40, P =.26). Among the 58 children without immune compromise, there was no association between polyomavirus excretion and the presence of vomiting, diarrhea, or hematochezia (21 patients with vs. 37 patients without symptoms; P =.28). Inadequate data were available for determination of an association between polyomavirus excretion and renal function or complete blood cell counts. We questioned whether older patients in the study cohort may have been more likely to be immune compromised, because the viral culture of stool and rectal swab samples seemed unusual for older children without immune suppression, but the predominance of young patients in the present study did not allow us to evaluate this possibility. Incidentally, we noted that numerous patients in the study cohort had required a consultation from the Pediatric Nutrition and Gastroenterology Service, a group that routinely orders virus isolation tests as part of its evaluation of symptoms of gastrointestinal disease.
Table 1.
Demographic and clinical characteristics of polyomavirus-excreting and -nonexcreting patients.
| Characteristic | Polyomavirus
|
BKV excreting (n = 38) | SV40 excreting (n = 8) | |
|---|---|---|---|---|
| Nonexcreting (n = 53) | Excreting (n = 46) | |||
| Age, mean (range), years | 5.14 (0.03–18.3) | 4.51 (0.03–17.2) | 4.81 (0.03–17.2) | 3.77 (0.36–13.6) |
| Race/ethnicity | ||||
| White (n = 39) | 21 | 18 | 17 | 1 |
| African American (n = 9) | 5 | 4 | 4 | 0 |
| Hispanic (n = 41) | 21 | 20 | 14 | 6 |
| Asian (n = 4) | 2 | 2 | 2 | 0 |
| Mixed/other (n = 6) | 4 | 2 | 1 | 1 |
| Sex | ||||
| Female (n = 54) | 29 | 25 | 24 | 1 |
| Male (n = 45) | 24 | 21 | 14 | 7 |
| Immune compromise | ||||
| Yes (n = 41) | 20 | 21 | 16 | 5 |
| No (n = 58) | 33 | 25 | 22 | 3 |
NOTE. Data are no. of patients, unless otherwise noted. For statistical analyses, polyomavirus-excreting patients were compared with polyomavirus-nonexcreting patients, BK virus (BKV)–excreting patients were compared with BKV-nonexcreting patients, and simian virus 40 (SV40)–excreting patients were compared with SV40-nonexcreting patients. Excepting SV40-nonexcreting male patients vs. SV40-nonexcreting female patients (P =.022 ), for all characteristics, P ≥.05 (χ2 test).
The demographic and clinical characteristics of the 8 children found to be shedding SV40 in stool are summarized in table 2. Seven (87.5%) were male, 6 (75.0%) were Hispanic, and 5 (62.5%) were immune compromised by virtue of cancer chemotherapy or organ transplantation. Two others were severely ill and required extended hospitalization and mechanical ventilation. The one patient found to be shedding SV40 in the absence of severe disease was a 4-month-old male with gastroenteritis.
Table 2.
Demographic and clinical characteristics of the SV40-excreting patients.
| Patient (age in months, sex) | Race/ethnicity | Diagnosis/treatment | Immune compromised |
|---|---|---|---|
| 1 (4.5, male) | Hispanic | Influenza A infection, sepsis, empyema | No |
| 2 (4.5, male) | Hispanic | Gastroenteritis | No |
| 3 (27, male) | Hispanic | Bacteremia, fungemia, respiratory syncytial virus bronchiolitis | No |
| 4 (29, male) | White | Rhabdomyosarcoma (chemotherapy) | Yes |
| 5 (33, female) | Hispanic | Neuroblastoma (chemotherapy) | Yes |
| 6 (41.5, male) | Hispanic | Hemophagocytic syndrome (bone-marrow transplant) | Yes |
| 7 (60, male) | Hispanic | Acute lymphoblastic leukemia (bone-marrow transplant) | Yes |
| 8 (162, male) | Other/mixed | Cystic fibrosis (lung transplant) | Yes |
Attempted culture of viruses from clinical samples
Viral culture of stool and rectal swab samples was performed at the Diagnostic Virology Laboratory with A549 cells, rhesus monkey kidney cells, and human foreskin fibroblasts, in accordance with standard laboratory practices. Adenoviruses were detected by viral culture in 12 of the 120 samples that had been tested for polyomavirus by PCR. The samples positive for adenovirus were from 8 children, 7 of whom were bone-marrow transplant recipients. The PYV.for/PYV.rev amplicons from 6 of the 12 samples positive for adenovirus (3 patients) produced very strong bands on ethidium bromide–stained agarose gel (e.g., lanes 1 and 5 in figure 1A), suggesting high polyomavirus loads in these samples.
The cell lines used for routine culture of viruses from clinical samples are not generally useful for the cultivation of polyomaviruses. Five samples positive for SV40 were used as inocula on TC7 cells in an attempt to amplify the small amounts of SV40 present in the clinical samples. No cytopathic effects were observed during 2 cell culture passages. Cell culture lysates were tested with the primers RA1 and RA2, which amplify a portion of the SV40 regulatory region [33]. One first-passage lysate was positive for the SV40 regulatory region. After a second passage in TC7 cells, another of the 5 samples (different from the first) was positive for the SV40 regulatory region. The 2 positive samples differed in the configuration of their regulatory regions, with one having a single enhancer (archetypal regulatory region) and the other having 2 enhancers (complex regulatory region). Studies of these samples are ongoing.
DISCUSSION
A source of BKV exposure that would account for the ubiquity of infection early during life has been elusive. The detection of SV40 excretion in the stool of infants after receipt of SV40-contaminated oral polio vaccine was the earliest suggestion that fecal shedding of polyomaviruses may contribute to the spread of these agents [26]. Also, this observation is consistent with the fecal shedding of several nonprimate polyomaviruses [6, 7]. The recent detection of polyomavirus DNA in human sewage [30, 34, 35] and SV40 DNA in rhesus macaque excrement (mixed stool and urine) [36] has implicated stool as a potential source of polyomavirus exposure. This could be especially pertinent for BKV, because it is uncommon in the urine of healthy individuals.
To explore the hypothesis that BKV fecal shedding can occur in children, we used a PCR assay that detects a conserved region in the T antigen genes of JCV, BKV, and SV40. We detected polyomavirus DNA in stool samples from 46.5% of hospitalized pediatric patients, a finding consistent with the notion that fecal shedding is a common source of polyomavirus exposure. Although the detection of polyomavirus DNA suggests that fecal-oral transmission could occur, the isolation of infectious BKV or SV40 from stool would be necessary to confirm this hypothesis.
Polyomavirus excretion in the stool of neonates (<28 days old) and infants (<1 year old) is consistent with the available serologic data indicating that infection with BKV can occur in very young children [37]. Whether in utero, intrapartum, or postpartum transmission accounts for some of these early infections is unknown. The absence of JCV detection is not surprising, given the age of the patients in the present study and the serologic evidence that JCV infections occur during late childhood or adolescence. It remains unknown whether JCV is excreted in stool. The detection of SV40 in human stool samples adds to the growing body of evidence suggesting continued circulation of this pathogen in the human population. It also is consistent with our recent finding of SV40 excretion in the urine of healthy children [12]. We were not able to discern whether vertical, zoonotic, transfusion-associated, or nosocomial transmission may have contributed to the acquisition of SV40 by the children included in the present study. Whether there is a true association between male sex and SV40 infection awaits larger studies that are amenable to multivariate analysis. The apparent absence of association between BKV DNA detection in stool and host immune compromise contrasts with the presence of the virus in urine, which is closely linked to immune status [4]. If this finding is validated in larger studies, it would suggest independent immune regulation of polyomavirus replication in different physiological compartments.
The detection of polymorphisms in BKV sequences suggests that there may be more diversity among circulating BKV strains than previously realized. This diversity has implications for the development of sensitive and specific methods of polyomavirus detection in patients with polyomavirus-associated disease.
There are several strengths and limitations to this study. The strengths include the study of individual stool or rectal swab samples, the large number of samples studied, the diversity in demographic background of the patients, and the use of a PCR technique that detects all 3 human polyomaviruses in a single test. The limitations include the lack of information on polyomavirus seroprevalence in the study population, the lack of information on simultaneous viruria, the inability to determine whether excretion was associated with primary infection or reactivation, the young age of the study cohort (the majority of patients were <2 years old), the uncertainty of whether the hospitalized study patients were otherwise healthy, and the inability to assess possible seasonal variation in shedding (time of sample inclusion was <4 months). These limitations could be overcome by a prospective study of stool and rectal swab samples that includes the acquisition of blood and urine samples at multiple times throughout the year. Clearly, these important issues require further study.
In summary, we have found that BKV excretion in stool by hospitalized pediatric patients is common. This finding provides support for the hypothesis that BKV and SV40 may be spread by fecal-oral transmission and adds to our knowledge of the natural history of polyomavirus infection in humans. Further study in both children and adults will be necessary to define the sites of polyomavirus persistence in the human gastrointestinal tract and to determine the clinical significance of polyomavirus shedding in human stool.
Acknowledgments
Financial support: National Institutes of Health (grants CA104818 and DK062090).
Footnotes
Potential conflicts of interest: none reported.
References
- 1.Flaegstad T, Traavik T, Kristiansen BE. Age-dependent prevalence of BK virus IgG and IgM antibodies measured by enzyme-linked im-munosorbent assays (ELISA) J Hyg (Lond) 1986;96:523–8. doi: 10.1017/s0022172400066328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Stolt A, Sasnauskas K, Koskela P, Lehtinen M, Dillner J. Seroepidemiology of the human polyomaviruses. J Gen Virol. 2003;84:1499–504. doi: 10.1099/vir.0.18842-0. [DOI] [PubMed] [Google Scholar]
- 3.Chang H, Wang ML, Tsai RT, et al. High incidence of JC viruria in JC-seropositive older individuals. J Neurovirol. 2002;8:447–51. doi: 10.1080/13550280260422758. [DOI] [PubMed] [Google Scholar]
- 4.Shah KV. Polyomaviruses. In: Fields BN, Knipe DM, Howley PM, et al., editors. Fields virology. 3. Vol. 2. Philadelphia: Lippincott-Raven; 1996. pp. 2027–43. [Google Scholar]
- 5.Cole CN. Polyomavirinae: the viruses and their replication. In: Fields BN, Knipe DM, Howley PM, et al., editors. Fields virology. 3. Vol. 2. Philadelphia: Lippincott-Raven; 1996. pp. 1997–2025. [Google Scholar]
- 6.Guerin JL, Gelfi J, Dubois L, Vuillaume A, Boucraut-Baralon C, Pingret JL. A novel polyomavirus (goose hemorrhagic polyomavirus) is the agent of hemorrhagic nephritis enteritis of geese. J Virol. 2000;74:4523–9. doi: 10.1128/jvi.74.10.4523-4529.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.van Gorder MA, Della Pelle P, Henson JW, Sachs DH, Cosimi AB, Colvin RB. Cynomolgus polyoma virus infection: a new member of the polyoma virus family causes interstitial nephritis, ureteritis, and enteritis in immunosuppressed cynomolgus monkeys. Am J Pathol. 1999;154:1273–84. doi: 10.1016/S0002-9440(10)65379-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jin L, Gibson PE, Booth JC, Clewley JP. Genomic typing of BK virus in clinical specimens by direct sequencing of polymerase chain reaction products. J Med Virol. 1993;41:11–7. doi: 10.1002/jmv.1890410104. [DOI] [PubMed] [Google Scholar]
- 9.Nickeleit V, Hirsch HH, Binet IF, et al. Polyomavirus infection of renal allograft recipients: from latent infection to manifest disease. J Am Soc Nephrol. 1999;10:1080–9. doi: 10.1681/ASN.V1051080. [DOI] [PubMed] [Google Scholar]
- 10.Hirsch HH. Polyomavirus BK nephropathy: a (re-)emerging complication in renal transplantation. Am J Transplant. 2002;2:25–30. doi: 10.1034/j.1600-6143.2002.020106.x. [DOI] [PubMed] [Google Scholar]
- 11.Ling PD, Lednicky JA, Keitel WA, et al. The dynamics of herpesvirus and polyomavirus reactivation and shedding in healthy adults: a 14-month longitudinal study. J Infect Dis. 2003;187:1571–80. doi: 10.1086/374739. [DOI] [PubMed] [Google Scholar]
- 12.Vanchiere JA, White ZS, Butel JS. Detection of BK virus and simian virus 40 in the urine of healthy children. J Med Virol. 2005;75:447–54. doi: 10.1002/jmv.20287. [DOI] [PubMed] [Google Scholar]
- 13.Di Taranto C, Pietropaolo V, Orsi GB, Jin L, Sinibaldi L, Degener AM. Detection of BK polyomavirus genotypes in healthy and HIV-positive children. Eur J Epidemiol. 1997;13:653–7. doi: 10.1023/a:1007371320999. [DOI] [PubMed] [Google Scholar]
- 14.Bhattacharjee S, Chakraborty T. High reactivation of BK virus variants in Asian Indians with renal disorders and during pregnancy. Virus Genes. 2004;28:157–68. doi: 10.1023/B:VIRU.0000016854.37475.f3. [DOI] [PubMed] [Google Scholar]
- 15.Markowitz RB, Eaton BA, Kubik MF, Latorra D, McGregor JA, Dynan WS. BK virus and JC virus shed during pregnancy have predominantly archetypal regulatory regions. J Virol. 1991;65:4515–9. doi: 10.1128/jvi.65.8.4515-4519.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chang D, Wang M, Ou WC, Lee MS, Ho HN, Tsai RT. Genotypes of human polyomaviruses in urine samples of pregnant women in Taiwan. J Med Virol. 1996;48:95–101. doi: 10.1002/(SICI)1096-9071(199601)48:1<95::AID-JMV15>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 17.Tsai RT, Wang M, Ou WC, et al. Incidence of JC viruria is higher than that of BK viruria in Taiwan. J Med Virol. 1997;52:253–7. [PubMed] [Google Scholar]
- 18.Pietropaolo V, Di Taranto C, Degener AM, et al. Transplacental transmission of human polyomavirus BK. J Med Virol. 1998;56:372–6. doi: 10.1002/(sici)1096-9071(199812)56:4<372::aid-jmv14>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- 19.Taguchi F, Nagaki D, Saito M, Haruyama C, Iwasaki K. Transplacental transmission of BK virus in humans. Jpn J Microbiol. 1975;19:395–8. doi: 10.1111/j.1348-0421.1975.tb00897.x. [DOI] [PubMed] [Google Scholar]
- 20.Shah K, Daniel R, Madden D, Stagno S. Serological investigation of BK papovavirus infection in pregnant women and their offspring. Infect Immun. 1980;30:29–35. doi: 10.1128/iai.30.1.29-35.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Andrews CA, Daniel RW, Shah KV. Serologic studies of papovavirus infections in pregnant women and renal transplant recipients. Prog Clin Biol Res. 1983;105:133–41. [PubMed] [Google Scholar]
- 22.Brown DW, Gardner SD, Gibson PE, Field AM. BK virus specific IgM responses in cord sera, young children and healthy adults detected by RIA. Arch Virol. 1984;82:149–60. doi: 10.1007/BF01311159. [DOI] [PubMed] [Google Scholar]
- 23.Coleman DV, Wolfendale MR, Daniel RA, et al. A prospective study of human polyomavirus infection in pregnancy. J Infect Dis. 1980;142:1–8. doi: 10.1093/infdis/142.1.1. [DOI] [PubMed] [Google Scholar]
- 24.Goudsmit J, Wertheim-van DP, van Strien A, van der Noordaa J. The role of BK virus in acute respiratory tract disease and the presence of BKV DNA in tonsils. J Med Virol. 1982;10:91–9. doi: 10.1002/jmv.1890100203. [DOI] [PubMed] [Google Scholar]
- 25.Sundsfjord A, Spein AR, Lucht E, Flaegstad T, Seternes OM, Traavik T. Detection of BK virus DNA in nasopharyngeal aspirates from children with respiratory infections but not in saliva from immunodeficient and immunocompetent adult patients. J Clin Microbiol. 1994;32:1390–4. doi: 10.1128/jcm.32.5.1390-1394.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Melnick JL, Stinebaugh S. Excretion of vacuolating SV-40 virus (papova virus group) after ingestion as a contaminant of oral poliovaccine. Proc Soc Exp Biol Med. 1962;109:965–8. doi: 10.3181/00379727-109-27392. [DOI] [PubMed] [Google Scholar]
- 27.Ricciardiello L, Chang DK, Laghi L, Goel A, Chang CL, Boland CR. Mad-1 is the exclusive JC virus strain present in the human colon, and its transcriptional control region has a deleted 98-base-pair sequence in colon cancer tissues. J Virol. 2001;75:1996–2001. doi: 10.1128/JVI.75.4.1996-2001.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ricciardiello L, Laghi L, Ramamirtham P, et al. JC virus DNA sequences are frequently present in the human upper and lower gastrointestinal tract. Gastroenterology. 2000;119:1228–35. doi: 10.1053/gast.2000.19269. [DOI] [PubMed] [Google Scholar]
- 29.Laghi L, Randolph AE, Chauhan DP, et al. JC virus DNA is present in the mucosa of the human colon and in colorectal cancers. Proc Natl Acad Sci USA. 1999;96:7484–9. doi: 10.1073/pnas.96.13.7484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bofill-Mas S, Pina S, Girones R. Documenting the epidemiologic patterns of polyomaviruses in human populations by studying their presence in urban sewage. Appl Environ Microbiol. 2000;66:238–45. doi: 10.1128/aem.66.1.238-245.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bofill-Mas S, Formiga-Cruz M, Clemente-Casares P, Calafell F, Girones R. Potential transmission of human polyomaviruses through the gastrointestinal tract after exposure to virions or viral DNA. J Virol. 2001;75:10290–9. doi: 10.1128/JVI.75.21.10290-10299.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bergsagel DJ, Finegold MJ, Butel JS, Kupsky WJ, Garcea RL. DNA sequences similar to those of simian virus 40 in ependymomas and choroid plexus tumors of childhood. N Engl J Med. 1992;326:988–93. doi: 10.1056/NEJM199204093261504. [DOI] [PubMed] [Google Scholar]
- 33.Lednicky JA, Stewart AR, Jenkins JJ, III, Finegold MJ, Butel JS. SV40 DNA in human osteosarcomas shows sequence variation among T-antigen genes. Int J Cancer. 1997;72:791–800. doi: 10.1002/(sici)1097-0215(19970904)72:5<791::aid-ijc15>3.0.co;2-c. [DOI] [PubMed] [Google Scholar]
- 34.Bofill-Mas S, Girones R. Role of the environment in the transmission of JC virus. J Neurovirol. 2003;9:54–8. doi: 10.1080/13550280390195306. [DOI] [PubMed] [Google Scholar]
- 35.Bofill-Mas S, Girones R. Excretion and transmission of JCV in human populations. J Neurovirol. 2001;7:345–9. doi: 10.1080/13550280152537210. [DOI] [PubMed] [Google Scholar]
- 36.Bofill-Mas S, Albinana-Gimenez N, Pipkin PA, Minor PD, Girones R. Isolation of SV40 from the environment of a colony of cynomolgus monkeys naturally infected with the virus. Virology. 2004;330:1–7. doi: 10.1016/j.virol.2004.09.007. [DOI] [PubMed] [Google Scholar]
- 37.Knowles WA. The epidemiology of BK virus and the occurrence of antigenic and genomic subtypes. In: Khalili K, Stoner GL, editors. Human polyomaviruses: molecular and clinical perspectives. New York: Wiley-Liss; 2001. pp. 527–59. [Google Scholar]