

Abstract
The present study was conducted to determine whether and how store-operated Ca2+ entry (SOCE) in glomerular mesangial cells (MCs) was altered by high glucose (HG) and diabetes. Human MCs were treated with either normal glucose or HG for different time periods. Cyclopiazonic acid-induced SOCE was significantly greater in the MCs with 7-day HG treatment and the response was completely abolished by GSK-7975A, a selective inhibitor of store-operated Ca2+ channels. Similarly, the inositol 1,4,5-trisphosphate-induced store-operated Ca2+ currents were significantly enhanced in the MCs treated with HG for 7 days, and the enhanced response was abolished by both GSK-7975A and La3+. In contrast, receptor-operated Ca2+ entry in MCs was significantly reduced by HG treatment. Western blotting showed that HG increased the expression levels of STIM1 and Orai1 in cultured MCs. A significant HG effect occurred at a concentration as low as 10 mM, but required a minimum of 7 days. The HG effect in cultured MCs was recapitulated in renal glomeruli/cortex of both type I and II diabetic rats. Furthermore, quantitative real-time RT-PCR revealed that a 6-day HG treatment significantly increased the mRNA expression level of STIM1. However, the expressions of STIM2 and Orai1 transcripts were not affected by HG. Taken together, these results suggest that HG/diabetes enhanced SOCE in MCs by increasing STIM1/Orai1 protein expressions. HG upregulates STIM1 by promoting its transcription but increases Orai1 protein through a posttranscriptional mechanism.
Keywords: store-operated Ca2+ entry, STIM1, Orai1, mesangial cells, high glucose, diabetic nephropathy
ubiquitous store-operated Ca2+ entry (SOCE), Ca2+ entry through store-operated Ca2+ channels (SOC) driven by depletion of endoplasmic reticulum (ER) Ca2+, is critical to the primary Ca2+ signaling pathway in a variety of cell types (59). This Ca2+ entry pathway plays an essential role in a wide variety of physiological functions including exocytosis, enzymatic activity, gene transcription, cell proliferation, and apoptosis (59). Although SOCE was originally described over two decades ago (64), its molecular mediators were unknown until recently. By high through-put RNAi screening, two protein families, STIM (45, 67) and Orai (22, 78, 87), were identified as required components of SOCE. STIM1 is a single-pass transmembrane protein located primarily in the ER membrane and functions as an ER Ca2+ sensor to sense ER luminal Ca2+ concentration. Orai1 is a small plasma membrane protein and is believed to be the pore-forming unit of SOC. Upon depletion of ER Ca2+, STIM1 aggregates and translocates to ER-plasma membrane junctions, where it physically associates and subsequently activates Orai1 and causes Ca2+ entry into the cytosol (12, 80).
It is well known that Ca2+ signals regulate functions of the renal microvasculature (10, 16, 18–21, 26). Glomerular mesangial cells (MCs) sit between glomerular capillary loops, a special part of the renal microvasculature, and maintains the structural architecture of the capillary networks. These cells play important roles in mesangial matrix homeostasis, regulation of glomerular filtration rate, and phagocytosis of apoptotic cells in the glomerulus (1, 68, 74). MC dysfunction is closely associated with several glomerular diseases, such as diabetic nephropathy (37, 39, 69). Like many other cell types, MC function is largely regulated by intracellular Ca2+ signals and MC Ca2+ homeostasis is, to a great extent, attributed to Ca2+ channels in the plasma membrane (47, 74). Over past decades, we and others have demonstrated that SOCE mediates MC Ca2+ responses to a variety of circulating and locally produced hormones, such as angiotensin II (ANG II), endothelin 1, and epidermal growth factor (9, 43, 48, 55, 58). We further demonstrated that STIM1 was required for activation of SOCE in human MCs (73). However, the association of SOCE in MCs with diabetic nephropathy remains unknown. The aim of the present study was to determine what and how diabetes affects SOCE in MCs using cultured MCs in combination with animal models of diabetes.
MATERIALS AND METHODS
MC culture.
Human MCs (HMC) belong to the Clonetics renal MC system and were purchased from Lonza (Walkersville, MD). MCs in a 75-cm2 flask were cultured in normal glucose (5.6 mM; NG) DMEM (GIBCO, Carlsbad, CA) supplemented with 25 mM HEPES, 4 mM glutamine, 1.0 mM sodium pyruvate, 0.1 mM nonessential amino acids, 100 U/ml penicillin, 100 μg/ml streptomycin, and 20% FBS. When MCs reached ∼90% confluence, the cells were split into 60-mm cell culture plates for various treatments as specified in figure legends. For NG treatment, 20 mM α-mannitol was added to the media as an osmotic control. For HG treatment, the concentration of d-glucose was 25 mM. Cells were growth-arrested with 0.5% FBS media during treatments. Culture media was replaced with fresh media every other day. Only sub-passage 4–9 MCs were used in the present study.
Transient transfection of human MCs.
In Fig. 5, E and F, small interfering (si) RNA against human Orai1 or scramble control siRNA were transiently transfected into human MCs using Lipofectamine and Plus reagent (Invitrogen-BRL, Carlsbad, CA) following the protocols provided by the manufacturer. Cells were harvested for Western blotting 72 h after transfection.
Fig. 5.

HG effect on Orai1 protein expression in HMCs. A: HMCs were cultured in 0.5% FBS NG or HG media for different time periods. B: HMCs were cultured in 0.5% FBS media with different concentrations of d-glucose for 7 days. Appropriate concentrations of α-mannitol were used as the osmotic control. In both A and B, Orai1 is shown as 2 bands, a lower band at ∼35 kDa and an upper band at ∼42 kDa. Actin was used as a loading control. C and D: summary data from experiments presented in A and B, respectively. In C, *P < 0.05, NG vs. HG for the same time period. n, No. of independent Western blots. In D, *P ≤ 0.05, vs. 5.6 mM glucose. E: Orai1 protein expression in human MCs with and without transfection of small interfering (si) RNA against human Orai1 (Orai1-siRNA) or scrambled siRNA (Scramble). UT, untransfected MCs; L, protein ladder. Actin was used as a loading control. F: summary data from experiments presented in E. n, No. of independent experiments. †P < 0.05, compared with both scrambled and Orai1 siRNA. G: fura 2 fluorescence ratiometry showing SOCE in HMCs with NG or HG treatment for 7 days with transfection of scrambled siRNA or Orai1 siRNA. Transfections were conducted on day 4 of HG treatment, and the Ca2+ imaging experiments were performed after 3 days of continuous HG treatment after transfection. n, No. of cells analyzed in each group. *P < 0.05, compared with NG. †P < 0.05, compared with HG and HG+Scramble.
Western blotting.
The whole-cell lysates, glomerular extracts, or renal cortical extracts were fractionated by 10% SDS-PAGE, transferred to polyvinylidene difluoride membranes, and probed with primary STIM1, Orai1, actin, and tubulin antibodies. Bound antibodies were visualized with Super Signal West Femto or Pico Luminol/Enhancer Solution (Thermo Scientific, Rockford, IL). The specific protein bands were visualized and captured using the AlphaEase FC Imaging System (Alpha Innotech, San Leandro, CA). The integrated density value (IDV) of each band was measured by drawing a rectangle outlining the band using AlphaEase FC software with auto background subtraction. If a protein had double bands, a total IDV by summation of each band IDV was used, The expression levels of STIM1 and Orai1 proteins were quantified by normalization of the IDVs of those protein bands to that of actin bands on the same blot.
Quantitative real-time RT-PCR.
Total RNA was isolated from cultured human MCs using a PerfectPure RNA cultured cell kit (5 Prime, Hamburg, Germany) following the manufacturer's protocol. All primers used in the present study were listed in Table 1. The primers were synthesized by IDT (Coralville, IA). A total of 1.0 μg RNA in a final volume of 20 μl was used for RT reactions using an iScript cDNA synthesis kit (Bio-Rad, Hercules, CA) following the manufacturer's reaction protocol. A total of 0.2 μg RT product and 100 nM primer was used for real-time PCR which was performed using iQ SYBR green supermix (Bio-Rad) in a final volume of 20 μl. The PCR mix was denatured at 95°C for 10 min, followed by 45 cycles of melting at 95°C for 15 s, annealing at 57°C for 10 s, and elongation at 72°C for 15 s. After amplification, a melting curve analysis from 65 to 95°C with a heating rate of 0.02°C/s with a continuous fluorescence acquisition was made. The assay was run on a C1000 Thermal Cycler (Bio-Rad). The average Ct (threshold cycle) of each fluorescence unit was used to analyze the mRNA levels. The STIM1, STIM2, and Orai1 mRNA levels were normalized by their corresponding β-actin mRNA levels. Quantification was calculated as follows: mRNA levels = 2ΔCt, where ΔCt = Ct, STIM1 or Ct, STIM2 or Ct, orai1 − Ct, actin.
Table 1.
Primers used for real-time PCR
| Gene Name | Primer (5′-3′) | GenBank Accession Number |
|---|---|---|
| STIM1 | F: ACAGGGACTGTGCTGAAGATGACA | NM_003156.2 |
| R: ACCAGCATGAAGTCCTTGAGGTGA | ||
| STIM2 | F: CAACACACACCACACTCCTT | NM_001169117.1 |
| R: GCCTCTTCCTCCTCTTCATTTC | ||
| Orai1 | F: CAGAGCATGGAGGGAAGAGGATTT | NM_032790.2 |
| R: ACCTGGAGCTGGAAGAACAGGAAA |
F, forward; R, reverse.
Fura 2 fluorescence ratiometry.
Measurement of intracellular Ca2+ concentration ([Ca2+]i) in MCs was performed using fura 2 fluorescence ratiometry as described elsewhere (81). Briefly, MCs, grown on a coverslip (22 × 22 mm), were loaded with 2 μM acetoxymethyl ester of fura 2 (fura 2-AM) plus 0.018 g/dl Pluronic F-127 (Invitrogen, Grand Island, NY) for 50 min at room temperature followed by an additional 20-min incubation in fura 2-free physiological saline solution. The coverslip was then placed in a perfusion chamber (model RC-2OH, Warner Instruments, Hamden, CT) mounted on the stage of a Nikon Diaphot inverted microscope. Fura 2 fluorescence was monitored at 340- and 380-nm excitation wavelengths and at 510-nm emission wavelength using NIS Elements AR software (Nikon Instruments, Melville, NY) at room temperature. [Ca2+]i was calculated using the software following the manufacturer's instructions. Calibrations were performed at the end of each experiment, and conditions of high [Ca2+]i were achieved by addition of 5 μM ionomycin, whereas conditions of low [Ca2+]i were obtained by addition of 5 mM EGTA.
Electrophysiology.
The conventional whole-cell voltage-clamp configuration was performed in single human MCs at room temperature with a Warner PC-505B amplifier (Warner Instruments) and Clampex 9.2 (Axon Instruments, Foster City, CA). Glass pipettes (plain, Fisher Scientific) with resistances of 3–5 MΩ were prepared with a pipette puller and polisher (PP-830 and MF-830, respectively, Narishige, Tokyo, Japan). When the whole-cell configuration was achieved, cell capacitance and series resistance were immediately compensated. The whole-cell currents were continuously recorded at a holding potential of −80 mV until the end of each experiment (∼10–15 min). Current traces were filtered at 1 kHz and analyzed off-line with Clampfit 9.2 (Axon Instruments). The compositions of the pipette solution were (in mM) 140 Cs-aspartate, 6 MgCl2, 10 BAPTA, 10 EGTA, and 10 HEPES. 1,4,5-Trisphosphate (IP3; 10 μM) was included in the solution for depleting internal Ca2+ stores (33, 44). There were two type of bathing solutions: Ca2+ solution and divalent-free solution (DVF). The compositions of the Ca2+ solution were (in mM) 130 NaCl, 5 KCl, 10 CaCl2, 1 MgCl2, and 10 HEPES, and the compositions of the DVF solution were (in mM) 150 NaCl, 10 Na2EDTA, and 10 HEPES.
Animals.
The study protocol was approved by the University of North Texas Health Science Center Institutional Animal Care and Use Committee. Two rat models of diabetes were used in this study. In the first, the streptozotocin (STZ)-type I diabetes model, a total of 12 male Sprague-Dawley rats at an age of 8 wk were purchased from Harlan (Indianapolis, IN). Diabetes (7 rats) was induced by intraperitoneal injection of STZ at 65 mg/kg body wt in sodium citrate buffer (0.01 M, pH 4.5) as we described previously (26, 46). An equivalent amount of sodium citrate buffer alone was used as a vehicle control (5 rats). Blood glucose levels were monitored 24 h later and periodically thereafter (LifeScan One Touch glucometer, Johnson&Johnson, Milpitas, CA) by rat-tailed blood sampling. STZ-injected rats with sustained elevation of blood glucose above 300 mg/dl were designated as diabetic rats. Four STZ-injected diabetic rats were euthanized at 2 wk after injection, and the remaining three were euthanized at 4 wk after injection. In the second model, the high-fat diet (HFD) plus STZ (HFD/STZ)-type II diabetes model (11, 24, 65, 75), a total of 10 male Sprague-Dawley rats at an age of 6 wk were evenly and randomly distributed to two groups, one group fed with a low-fat diet (LFD), which served as controls, and the other group fed with a HFD. In the LFD, fat, protein, carbohydrates, and ethanol represent 10, 20, 67, and 1.8% of the total calories, respectively, while in the HFD, fat, protein, carbohydrates, and ethanol represent 44, 20, 34, and 1% of the total calories, respectively (Research Diets, Brunswick, NJ). In the HFD group, after 5 wk of HFD, the rats were given STZ at 35 mg/kg via the tail vein. Fifteen weeks later, when these HFD/STZ rats manifested overt type II diabetic phenotypes (Table 2), all LFD and HFD/STZ rats were euthanized for biochemical assays.
Table 2.
Diabetic parameters in LFD and HFD rats
| Index | LFD | HFD |
|---|---|---|
| Albumin/creatinine ratio, μg:mg | 508 ± 137 | 4,522 ± 697* |
| Blood glucose, mg/dl | 85 ± 2 | 350 ± 15* |
| Body weight, g | 484 ± 14 | 440 ± 13 |
| Urine output, ml·100 g body wt−1·24 h−1 | 1.4 ± 0.1 | 36.9 ± 2.7* |
| Serum insulin, ng/ml | 0.04 ± 0.01 | 0.84 ± 0.01* |
| Triglyceride, mg/dl | 61 ± 7 | 333 ± 31* |
| Cholesterol, mg/dl | 105 ± 4 | 289 ± 64* |
Values are means ± SE; n = 5/group. LFD, low-fat diet; HFD, high-fat diet.
P < 0.01, HFD vs. LFD (Student's t-test).
Isolation of renal cortex and glomeruli and extracting cortical and glomerular proteins.
The protocol of isolating glomeruli was described elsewhere (57) with modifications. Briefly, rats were euthanized and both kidneys were quickly removed. The renal cortex was separated from the other region of kidney using a sharp blade, and the cortical tissue was minced using two sharp blades. For STZ-injected rats, glomeruli were isolated by differential sieving of minced renal cortex. Finely chopped kidney cortex in Hanks' balanced salt solution (pH 7.4) was pressed through sequentially smaller metal sieves and collected on a final sieve of 63-μm pore size (minisieve set, Scienceware, Pequannock, NJ). Glomeruli were pelleted by centrifugation at 500 g for 10 min at 4°C (5810R, Eppendorf). We have previously shown that the purity of the glomerulus preparation was ∼99% (26). Both cortical tissues and glomeruli were sonicated in a lysis buffer followed by centrifugation at 20,817 g for 15 min at 4°C. The supernatants were collected for Western blotting.
Materials.
Cyclopiazonic acid (CPA) was purchased from Alomone Laboratories (Har Hotzvim Hi-Tech Park, Jerusalem, Israel). GSK-7975A was kindly donated by GSK (Stevenage, UK). The rabbit polyclonal anti-STIM1 antibody was purchased from ProteinTech (Chicago, IL). Orai1 antibody was purchased from Abcam (for rat tissues, Cambridge, MA) and Sigma (for human MCs, Sigma, St. Louis, MO). All other chemicals and antibodies were purchased from Sigma-Aldrich unless indicated.
Statistical analysis.
Data were reported as means ± SE. One-way ANOVA plus Student-Newman-Keuls post hoc analysis and Student's unpaired t-test were used to analyze the differences among multiple groups and between two groups, respectively. P < 0.05 was considered statistically significant. Statistical analysis was performed using SigmaStat (Jandel Scientific, San Rafael, CA).
RESULTS
SOCE in HMCs was enhanced by prolonged HG treatment.
In HMCs cultured in NG and HG at different time periods, we measured the fura 2 fluorescence-indicated Ca2+ entry response using a classic “Ca2+ add-back” protocol described in our previous publications (15, 16, 26, 27). CPA was used to activate SOCE. As shown in Fig. 1, a Ca2+ entry response was observed in both NG and HG cultured MCs upon readdition of 2 mM Ca2+ to a Ca2+-free bathing solution. There was no difference in the CPA-induced SOCE between NG and HG treatments for the time periods of 1 and 3 days (Fig. 1, A, B, and E). However, this Ca2+ entry response was significantly greater in the cells with 7- and 14-day HG treatments compared with NG treatment for the same time periods (Fig. 1, C–E). The augmented response to 7-day HG treatment was completely abolished by GSK-7975A, a selective SOC inhibitor which directly acts on the pore region of SOC (2, 13, 25, 66), verifying the Ca2+ entry being mediated by SOC, i.e., SOCE in Fig. 1D. Furthermore, in the absence of CPA, readdition of 2 mM Ca2+ also produced Ca2+ entry in both NG and HG 7-day-treated HMCs (Fig. 1F). However, the tonic Ca2+ entry responses were much weaker compared with the responses in the presence of CPA (Fig. 1, F and C) and were not significantly different with NG and HG treatments (Fig. 1G). These results suggest that prolonged HG treatment enhanced SOCE, but did not alter tonic Ca2+ entry.
Fig. 1.

High glucose (HG) effect on store-operated Ca2+ entry (SOCE) in human mesangial cells (HMCs). Fura 2 fluorescence ratiometry was used to assess the intracellular Ca2+ concentration ([Ca2+]i). MCs were initially bathed in physiological saline solution (containing 1 mM Ca2+). After [Ca2+]i was stable (∼2 min), 25 μM cyclopiazonic acid (CPA) was applied to the bath to deplete the internal Ca2+ stores, indicated by the first transient. When the CPA-induced initial Ca2+ spike declined steadily, the bathing solution was replaced with a Ca2+-free solution in the presence of CPA, which was followed by readdition of a 2 mM Ca2+ solution containing CPA. SOCE was estimated as the increase in [Ca2+]i upon switching of the bathing solution from a Ca2+-free to a 2 mM Ca2+ solution, indicated by a dashed rectangle. A–D: representative traces showing CPA (25 μM)-evoked Ca2+ response in HMCs with normal glucose (NG) and HG treatment for 1, 3, 7, and 14 days. [Ca2+]B represents the Ca2+ concentration in the bathing solution. E: summary data showing time course effect of HG on SOCE in HMCs. Numbers inside the bars indicate the number of cells analyzed in each group. GSK, GSK-A7975. *P < 0.05, compared with NG at the same time period. †P < 0.05, compared with HG for 7 days. F: representative traces showing Ca2+ response to readdition of Ca2+, in the absence of CPA, in HMCs with NG and HG treatment for 7 days. G: summary data from experiments shown in F. n, No. of cells analyzed in each group.
SOC activity in HMCs was promoted by prolonged HG treatment.
SOCE is mediated by SOC in the plasma membrane. Since chronic treatment with HG increased SOCE in MCs, we would expect a stimulatory effect on SOC by HG. As expected, whole-cell patch-clamp experiments showed that store-operated Ca2+ currents were significantly augmented by HG treatment for 7 days, but not for 1 day (Fig. 2, A–E). Because SOC is more conductive to monovalent cations over divalent cations (4, 5, 34, 62), we also measured DVF currents in the cells with NG and HG treatments. As shown in Fig. 2, A–D and F, the DVF currents were much more robust compared with Ca2+ currents, a characteristic of SOC. Consistent with Ca2+ current response, the DVF currents were significantly augmented by 7-day, but not 1-day HG treatment (Fig. 2, A–D and F). Both the Ca2+ and DVF current responses to the prolonged HG treatment were significantly inhibited by GSK-7975A (10 μM), but not by DMSO, a vehicle control (Fig. 2G). Furthermore, a low concentration of La3+ can selectively block SOC (7, 8, 49, 77). We examined how much currents were reduced by La3+ (2 μM) as an indication of SOC activity in NG- and HG-treated MCs. As shown in Fig. 2H, the La3+-sensitive decrease in DVF current was significantly greater in MCs with 7-day HG treatment compared with NG treatment for the same time period. There was no significant difference in the La3+ response between NG and HG treatments for 1 day.
Fig. 2.

HG effect on SOC currents in HMCs. Whole-cell currents in response to depletion of intracellular Ca2+ stores in HMCs with NG and HG treatment for 1 and 7 days are shown. The whole-cell current was recorded at a holding potential of −80 mV. A–D: representative traces. Arrows, membrane breaking-in; horizontal bars, type of bathing solution [Ca2+ or divalent-free (DVF) solution] and the time of application of La3+. E–G: store depletion-induced membrane currents in Ca2+ solution (E) and DVF solution (F) and blockade of currents by GSK-7975A (10 μM) in HMCs with 7-day HG treatment (G). The response was calculated as the difference between the basal membrane current (before breaking-in) and the peak current after breaking-in. H: membrane currents reduced by 2 μM La3+ in DVF solution. Numbers under each bar, no. of cells analyzed in each group. *P < 0.05.
HG reduced receptor-operated Ca2+ entry in HMCs.
In addition to SOC, we and others have demonstrated that receptor operated Ca2+ channels (ROC) or canonical transient receptor potential channels (TRPC) also participate in Ca2+ signaling in MCs (16, 47, 58, 72). To be distinguished from SOC, ROC is defined as the channel activated by the G protein-coupled receptor signaling pathway through a mechanism bypassing the IP3-induced internal Ca2+ store depletion. It has been debated for ∼20 years whether the HG-impaired Ca2+ response in MCs was mediated by SOC or ROC (23, 26, 27, 52, 58, 71). To dissect SOCE from receptor-operated Ca2+ entry (ROCE) in response to HG treatment, we performed an additional Ca2+ imaging study in which HMCs with NG or HG treatment for 7 days were incubated with CPA (25 μM) for ∼7 min to deplete the internal Ca2+ stores (activation of SOC). As described above, SOCE was estimated by the elevation of [Ca2+]i upon switching the bathing solution from Ca2+-free to a 2 mM Ca2+ solution. This was followed by addition of ANG II (1 μM) to the bathing solution in the presence of CPA. Since the intracellular Ca2+ stores had been depleted by CPA, the additional increase in [Ca2+]i by ANG II would be attributed to ROCE. As shown in Fig. 3, although SOCE was significantly greater in the MCs with 7-day HG treatment (consistent with Fig. 1), ROCE was significantly reduced. The contribution of ROCE to the additional response by ANG II was verified by an inhibitory effect of SKF96365 (an inhibitor of ROC) on the response (Fig. 3, C and D). This ANG II-stimulated, SOCE-independent Ca2+ response was also significantly attenuated by blocking voltage-gated Ca2+ channels with diltiazem (31, 49) (Fig. 3, C and D). Activation of voltage-gated Ca2+ channels has been proposed as one mechanism in the ROC-dependent Ca2+ response because of membrane depolarization by Na+ influx through ROC (70, 83). In summary, these results are consistent with the notion that the ANG II-induced Ca2+ entry after CPA treatment was mediated by ROC.
Fig. 3.

HG effect on receptor-operated Ca2+ entry (ROCE) in HMCs. Fura 2 fluorescence ratiometry shows SOCE and ROCE in HMCs with NG or HG treatment for 7 days. MCs were treated with 25 μM CPA for ∼7 min to deplete the internal Ca2+ stores. SOCE was estimated by the elevation of [Ca2+]i upon switching of the bathing solution from a Ca2+-free to a 2 mM Ca2+ solution in the presence of CPA. ROCE was defined as the additional increase in [Ca2+]i by adding ANG II (1 μM) in the presence of CPA. A–C: representative traces recorded ∼4 min after CPA application. [Ca2+]B represents the Ca2+ concentration in the bathing solution. Thick arrows, application of ANG II with and without diltiazem (Dil) or SKF96365 (SKF) Traces were smoothed using SigmaPlot software (version 11.0). D: summary data averaged from NG groups with ANG II treatment (n = 29), HG with ANG II treatment (n = 18), NG with ANG II+Dil treatment (n = 16), and NG with ANG+SKF treatment (n = 15). *P < 0.05, compared with SOCE in NG group. †P < 0.01, compared with ROCE in NG group.
HG increased STIM1 protein expression in cultured HMCs.
STIM1 has been demonstrated as a key component of SOCE by gating SOC (45, 67). Our previous study has also shown that STIM1 was required for activation of SOCE in HMCs (73). Since HG-promoted SOCE in MCs was a chronic effect (Figs. 1 and 2), we thought that an increase in abundance of one or more key proteins in the SOCE pathway, such as STIM1, contributed to the HG effect. Thereby, Western blotting was performed to determine the HG effect on STIM1 protein expression in cultured HMCs. As shown in Fig. 4, the expression level of STIM1 was remarkably increased by HG treatment. In agreement with the results from functional studies (Figs. 1 and 2), the HG effect on STIM1 expression was also a chronic process, and a minimum of 7 days was required for a significant increase (Fig. 4, A and C). We also examined the STIM1 response to different concentrations of glucose and showed that a concentration of 10 mM was sufficient to significantly increase STIM1 protein expression (Fig. 4, B and D). Although there was a trend toward an increase in STIM1 protein expression with increases in concentrations of glucose, no significant difference in STIM1 expression levels was observed among different concentrations of glucose in a range of 10–30 mM (Fig. 4, B and D).
Fig. 4.
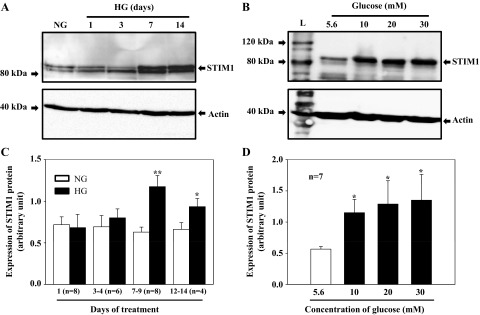
HG effect on STIM1 protein expression in HMCs. A: HMCs were cultured in 0.5% FBS NG media for 3 days or in 0.5% FBS HG media for different time periods. B: HMCs were cultured in 0.5% FBS media with different concentration of d-glucose for 7 days. Appropriate concentrations of α-mannitol were used as the osmotic control. C and D: summary data showing the time course and dose dependence of HG effect on STIM1 protein expression. In C, **P < 0.01 and *P < 0.05, NG vs. HG for the same time period. n, No. of independent Western blots. In D, *P ≤ 0.05, vs. 5.6 mM glucose.
HG increased Orai1 protein expression in cultured HMCs.
Orai1 is the pore-forming protein of SOC (12, 12, 80). To determine whether HG-stimulated SOCE involved upregulation of Orai1 protein expression, Western blotting was performed in cultured HMCs. Orai1 protein was indicated by the two bands at sizes of ∼37 and ∼43 kDa (Fig. 5, A and B), which was verified by knockdown of Orai1 using siRNA (Fig. 5, E and F). Similar to the STIM1 response, HG significantly increased Orai1 protein level after an ∼7-day treatment (Fig. 5, A and C), and a concentration of 10 mM was high enough to induce a significant increase (Fig. 5, B and D).
To determine whether Orai1 contributed to the enhanced SOCE, we examined the effect of knocking down Orai1 using siRNA on the HG-enhanced Ca2+ response. As shown in Fig. 5G, HG treatment for 7 days significantly increased SOCE and this enhanced response was abolished by Orai1-siRNA, but not by scrambled siRNA.
Diabetes increased abundance of STIM1 and Orai1 proteins in glomeruli and renal cortex.
The in vitro effects of HG on expressions of the two key SOCE proteins in cultured MCs were further examined in intact animals. Several rat models of diabetes were used to detect the diabetic effect on STIM1 and Orai1 protein expression in glomeruli/cortex, where MCs are located. In STZ type I diabetic rats, the model we have used before (26), the amount of STIM1 protein was dramatically increased in glomeruli from the rats 4 wk after STZ injection. Although glomerular STIM1 protein in the rats with 2 wk of STZ showed a tendency to increase, this response did not reach a significant level compared with nondiabetic control glomeruli (Fig. 6, A and B). The upregulation of STIM1 protein by diabetes was further verified in the renal cortex of the rats with a HFD followed by STZ treatment (Fig. 6, C and D), which is a well-established nongenetic type II diabetes model (11, 24, 65, 75). As shown in Table 2, these HFD/STZ rats manifested overt type II diabetic phenotypes, characterized by albuminuria, hyperlipidemia, hyperglycemia, and hyperinsulinemia.
Fig. 6.

Effect of diabetes on STIM1 protein expression in rat kidney. Western blotting shows STIM1 protein expression in extracts of glomeruli freshly isolated from rats 2 and 4 wk after streptozotocin (STZ) injection (STZ-2W and STZ-4W, respectively) and 4 wk after vehicle injection (control; A and B) and in renal cortex of low-fat diet (LFD) and high-fat diet (HFD) rats (C and D). In A and C, actin was used as a loading control. B and D: summary data from experiments presented in A and C, respectively. *P < 0.05, compared with control in B and LFD in D.
In agreement with the STIM1 response to diabetes, Orai1 protein expression was also increased in the glomeruli from the rats with 4 wk of STZ treatment and in the renal cortex of rats with the HFD followed by STZ injection (Fig. 7).
Fig. 7.

Effect of diabetes on Orai1 protein expression in rat kidney. Western blotting shows Orai1 protein expression in extracts of glomeruli freshly isolated from rats 4 wk after STZ injection (STZ-4W) and 4 wk after vehicle injection (control; A and B) and in renal cortex of LFD and HFD rats (C and D). In A and C, β-integrin and tubulin were used as loading controls, respectively. B and D: summary data from experiments presented in A and C, respectively. **P < 0.01, compared with control in B and LFD in D.
mRNA expression level of STIM1, but not Orai1 was increased by HG in HMCs.
To determine whether the diabetes/HG-associated upregulation of STIM1 and Orai1 in MCs occurred at the transcriptional or posttranscriptional level, we conducted quantitative real-time RT-PCR. Total RNA was isolated from MCs treated with 5.6 and 25 mM glucose for 1, 3, and 6 days. As shown in Fig. 8A, HG treatment for 6 days significantly increased the mRNA expression level of STIM1. The HG effect was specific because STIM2, which is another member of the STIM family and shares protein domain organization and biochemical features with STIM1 (85), did not change with HG treatment (Fig. 8B). Different from the STIM1 response, Orai1 mRNA expression level did not increase at all with HG in a time period ranging from 1-day to 6-day treatment (Fig. 8C). These data suggest that HG and hyperglycemia in diabetes increased STIM1 protein expression by a positive regulation of STIM1 gene expression, but increased Orai1 protein expression through a posttranscriptional mechanism.
Fig. 8.
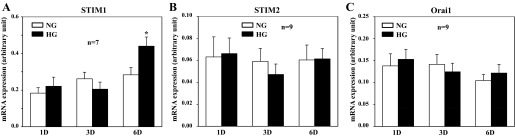
Quantitative real-time RT-PCR, showing HG effect on STIM1, STIM2, and Orai1 mRNA expressions in HMCs. HMCs were cultured in 0.5% FBS NG or HG media for different days (D). In A, *P < 0.05, HG vs. NG for 6D treatment.
DISCUSSION
The HG effect on SOCE in MCs has been debated for over 20 years. In an earlier study by Mene et al. (52), arginine vasopressin-induced Ca2+ influx was significantly reduced in rat MCs treated with HG (30 mM) for 5 days (52). Because this vasoconstrictor can release Ca2+ from the ER via the classic Gq-coupled receptor pathway, the authors speculated that the impaired Ca2+ response was due to a suppressed SOCE. However, their further data showing that the HG effect was mediated by protein kinase C, a mechanism bypassing internal Ca2+ store depletion, contradicted with the SOCE mechanism. Several studies from other groups did not support SOCE as responsible for HG-induced impairment of the Ca2+ response (23, 58). For instance, Nutt and O'Neil (58) in a later study demonstrated that it was ROC, but not SOC that was responsible for a reduced Ca2+ response to endothelin 1 in rat MCs treated with HG for 5–7 days. Their findings are consistent with our recent reports that HG reduced the abundance of TRPC6 (26, 27), which is a well-known ROC (14). The present study provided Ca2+ imaging and electrophysiological evidence that HG enhanced SOCE in HMCs. This conclusion was further supported by biochemical data which showed a significant increase in the expression level of STIM1 and Orai1 proteins, two key components of the SOCE pathway (3, 32, 42, 45, 61, 63). Importantly, there is a good time correlation between increases in the two-protein expression and enhancement of SOCE in response to HG, suggesting a STIM1/Orai1-dependent mechanism for HG-promoted SOCE in MCs.
Like in many other studies (23, 52, 53, 58), HG-induced impairment of Ca2+ signaling in MCs is a chronic process in a range from 2 to 7 days. In our case, a significant increase in SOCE and protein expression levels of STIM1/Orai1 occurred 7 days after HG treatment. The prolonged response indicates multiple intracellular processes are involved. Apparently, further exploration of the intermediators linking HG with SOCE would be important for delineating the molecular mechanisms for HG-promoted SOCE. Interestingly, our study suggests that the mechanism for the HG-induced STIM1 response is different from the mechanism mediating the Orai1 response. The STIM1 response was apparently attributed to elevated transcriptional activity, as indicated by a significant increase in mRNA expression level by HG treatment. Although the present study did not determine how HG activated STIM1 gene transcription, multiple transcription factors, such as NF-κB and activated protein 1 (AP-1), which have been known to play a critical role in the development of DN (6, 28, 30, 36, 56, 82), are speculated to be downstream molecules of the HG-signaling pathway. Particularly, NF-κB has been reported to promote stim1 transcription by binding to its promoter region (17). Different from STIM1, the Orai1 response to HG may be through a posttranscription mechanism because HG treatment did not change the steady-state level of Orai1 mRNA. Although the exact underlying mechanism is unknown from the present study, ubiquitination and lysosomal degradation of Orai1 have been reported recently (41).
The pathological relevance of HG-regulated STIM1/Orai1 protein expressions was indicated by an increase in abundance of the two proteins in the renal glomeruli or cortex of both type I and II diabetic rats. While the pathogenesis of diabetes is different in type I and type II diabetes, the natural history of DN is similar. Thus it is not surprising that STIM1/Orai1 proteins had a consistent change in abundance in both type I and II diabetic rats. MCs play a critical role in the development of DN in both early and late stages of diabetes (37, 38, 40, 88). A question raised from the ex vivo study is whether the increases in STIM1/Orai1 proteins were derived from the MC response. Although contributions from other cell types, such as podocytes, endothelial cells, and tubular epithelial cells (in renal cortex) cannot be ruled out, MCs constitute one-third of the glomerular cell population (54), and thereby the changes in STIM1/Orai1 protein expression in MCs are expected to be detectable at the tissue level. Another question from the animal experiments is whether the increased STIM1/Orai1 proteins in diabetic kidneys were due to hyperglycemia. Indeed, hyperglycemia is the main determinant of initiation and progression of diabetic microvascular complications including nephropathy (29, 76). In vitro and in vivo studies have demonstrated that HG or hyperglycemia directly stimulates MCs, which subsequently results in mesangial dysfunction or malfunction (40, 51, 52, 58, 84). Combined with the data from cultured MCs and the fact that all diabetic rats used in the present study were hyperglycemic, we reason that the elevated cellular glucose level contributed, at least partially if not fully, to the STIM1/Orai1 response.
In addition to the kidney, a recent study reported that STIM1/Orai1 protein expression levels were also upregulated in platelets isolated from patients with type II diabetes mellitus (86), suggesting that diabetes may have a general effect on SOCE.
What is the physiological and pathological relevance of the enhancement of SOCE in diabetic MCs? Ca2+ signaling in MCs involves multiple Ca2+ channels, each of which may have a specified downstream pathway and thus have distinct functional consequences (47). For instance, our previous study demonstrated that among four subtypes of TRPC channels endogenously expressed in MCs, HG/diabetes did not either alter (TRPC1, 3, and 4) or decrease (TRPC6) the abundance of TRPC proteins (26). The downregulation of TRPC6 impairs the contractile function of MCs by attenuating the agonist-stimulated Ca2+ response, which may contribute to hyperfiltration at the early stage of diabetes (26). Different from TRPC channel (likely function as ROC)-mediated Ca2+ entry, SOCE is more related to sustained and chronic changes in cell function, such as gene transcription, cell proliferation, and apoptosis (59). Recently, SOCE was found to contribute to cardiac hypertrophy (35, 79), and STIM1-regulated Ca2+ homeostasis is crucial for smooth muscle cell proliferation, development, and growth response to injury (50). In contrast, SOCE constrains tuberous sclerosis complex-related tumor development in mice (60). Thus the effect of SOCE on cell growth and protein synthesis may depend on cell types and downstream signaling pathways. Although the importance of SOCE in renal function and renal disease development has not been examined in the present study, it is certainly noteworthy to address this issue in the near future.
In summary, our findings from the present study imply that HG/diabetes enhanced SOCE in MCs by increasing STIM1/Orai1 protein expressions. HG upregulates STIM1 by promoting its transcription, but increases Orai1 protein through a posttranscriptional mechanism.
GRANTS
The work was supported by National Institutes of Health (NIH/NIDDK) Grant RO1 (5RO1DK079968-01 to R. Ma) and by a Grant-in-Aid (11GRNT7560013) from the American Heart Association Southwestern Affiliate (to R. Ma).
DISCLOSURES
Malcolm Begg is employed by GSK.
AUTHOR CONTRIBUTIONS
Author contributions: S.C., P.W., Y.W., and Y.D. performed experiments; S.C., P.W., Y.W., Y.D., and R.M. analyzed data; S.C., P.W., Y.W., Y.D., j.Y., and R.M. interpreted results of experiments; S.C., P.W., Y.W., and R.M. prepared figures; S.C., P.W., j.Y., M.B., and R.M. edited and revised manuscript; S.C., P.W., Y.W., Y.D., j.Y., M.B., and R.M. approved final version of manuscript; R.M. provided conception and design of research; R.M. drafted manuscript.
ACKNOWLEDGMENTS
We thank GlaxoSmithKline (Stevenage, UK) for providing GSK-7975A compound.
REFERENCES
- 1.Abboud HE. Mesangial cell biology. Exp Cell Res 318: 979–985, 2012 [DOI] [PubMed] [Google Scholar]
- 2.Ashmole I, Duffy SM, Leyland ML, Morrison VS, Begg M, Bradding P. CRACM/Orai ion channel expression and function in human lung mast cells. J Allergy Clin Immunol 129: 1628–1635, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Baba Y, Hayashi K, Fujii Y, Mizushima A, Watarai H, Wakamori M, Numaga T, Mori Y, Lino M, Hikida M, Kurosaki T. Coupling of STIM1 to store-operated Ca2+ entry through its constitutive and inducible movement in the endoplasmic reticulum. Proc Natl Acad Sci USA 103: 16704–16709, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bakowski D, Parekh AB. Monovalent cation permeability and Ca2+ block of the store-operated calcium current ICRAC in rat basophilic leukaemia cells. Pflügers Arch 443: 892–902, 2002 [DOI] [PubMed] [Google Scholar]
- 5.Bakowski D, Parekh AB. Permeation through store-operated CRAC channels in divalent-free solution: potential problems and implications for putative CRAC channel genes. Cell Calcium 32: 379–391, 2002 [DOI] [PubMed] [Google Scholar]
- 6.Banday AA, Fazili FR, Lokhandwala MF. Oxidative stress causes renal dopamine D1 receptor dysfunction and hypertension via mechanisms that involve nuclear factor-kB and protein kinase C. J Am Soc Nephrol 18: 1446–1457, 2007 [DOI] [PubMed] [Google Scholar]
- 7.Bird GS, Dehaven WI, Smyth JT, Putney JW., Jr Methods for studying store-operated calcium entry. Methods 46: 204–212, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Broad LM, Cannon TR, Taylor CW. A non-capacitive pathway activated by arachidonic acid is the major Ca2+ entry mechanism in rat A7r5 smooth muscle cells stimulated with low concentrations of vasopressin. J Physiol (Lond) 517: 121–134, 1999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Campos AH, Calixto JB, Schor N. Bradykinin induces a calcium-store-dependent calcium influx in mouse mesangial cells. Nephron 91: 308–315, 2002 [DOI] [PubMed] [Google Scholar]
- 10.Che Q, Carmines PK. Src family kinase involvement in rat preglomerular microvascular contractile and [Ca2+]i responses to ANG II. Am J Physiol Renal Physiol 288: F658–F664, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Danda RS, Habiba NM, Rincon-Choles H, Bhandari BK, Barnes JL, Abboud HE, Pergola PE. Kidney involvement in a nongenetic rat model of type 2 diabetes. Kidney Int 68: 2562–2571, 2005 [DOI] [PubMed] [Google Scholar]
- 12.Deng X, Wang Y, Zhou Y, Soboloff J, Gill DL. STIM and Orai-dynamic intermembrane coupling to control cellular calcium signals. J Biol Chem 284: 22501–22505, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Derler I, Schindl R, Fritsch R, Heftberger P, Riedl MC, Begg M, House D, Romanin C. The action of selective CRAC channel blockers is affected by the Orai pore geometry. Cell Calcium 53: 139–151, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dietrich A, Gudermann T. TRPC6. Handb Exp Pharmacol 179: 125–141, 2007 [DOI] [PubMed] [Google Scholar]
- 15.Du J, Ding M, Sours-Brothers S, Graham S, Ma R. Mediation of angiotensin II-induced Ca2+ signaling by polycystin 2 in glomerular mesangial cells. Am J Physiol Renal Physiol 294: F909–F918, 2008 [DOI] [PubMed] [Google Scholar]
- 16.Du J, Sours-Brothers S, Coleman R, Ding M, Graham S, Kong D, Ma R. Canonical transient receptor potential 1 channel is involved in contractile function of glomerular mesangial cells. J Am Soc Nephrol 18: 1437–1445, 2007 [DOI] [PubMed] [Google Scholar]
- 17.Eylenstein A, Schmidt S, Gu S, Yang W, Schmid E, Schmidt EM, Alesutan I, Szteyn K, Regel I, Shumilina E, Lang F. Transcription factor NF-κB regulates the expressions of a pore-forming unit, Orai1, and its activator, STIM1, to control Ca2+ entry and affect cellular functions. J Biol Chem 287: 2719–2730, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fallet RW, Ikenaga H, Bast JP, Carmines PK. Relative contributions of Ca2+ mobilization and influx in renal arteriolar contractile responses to arginine vasopressin. Am J Physiol Renal Physiol 288: F545–F551, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fellner S, Arendshorst WJ. Angiotensin II-stimulated calcium entry mechanism in afferent arterioles: role of transient receptor potential canonical channels and reverse Na+/Ca2+ exchange. Am J Physiol Renal Physiol 294: F212–F219, 2008 [DOI] [PubMed] [Google Scholar]
- 20.Fellner SK, Arendshorst WJ. Angiotensin II Ca2+ signaling in rat afferent arterioles: stimulation of cyclic ADP ribose and IP3 pathways. Am J Physiol Renal Physiol 288: F785–F791, 2005 [DOI] [PubMed] [Google Scholar]
- 21.Fellner SK, Arendshort WJ. Store-operated Ca2+ entry is exaggerated in fresh preglomerular vascular smooth muscle cells of SHR. Kidney Int 61: 2132–2141, 2002 [DOI] [PubMed] [Google Scholar]
- 22.Feske S, Gwack Y, Prakriya M, Srikanth S, Puppel SH, Tanasa B, Hogan PG, Lewis RS, Daly M, Rao A. A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 441: 179–185, 2006 [DOI] [PubMed] [Google Scholar]
- 23.Frecker H, Munk S, Wang H, Whiteside C. Mesangial cell reduced Ca2+ signaling in high glucose is due to inactivation of phospholipase C-β3 by protein kinase C. Am J Physiol Renal Physiol 289: F1078–F1087, 2005 [DOI] [PubMed] [Google Scholar]
- 24.Gaikwad AB, Gupta J, Tikoo K. Epigenetic changes and alteration of Fbn1 and Col3A1 gene expression under hyperglycaemic and hyperinsulinaemic conditions. Biochem J 432: 333–341, 2010 [DOI] [PubMed] [Google Scholar]
- 25.Gerasimenko JV, Gryshchenko O, Ferdek PE, Stapleton E, Hebert TO, Bychkova S, Peng S, Begg M, Gerasimenko OV, Petersen OH. Ca2+ release-activated Ca2+ channel blockade as a potential tool in antipancreatitis therapy. Proc Natl Acad Sci USA 110: 13186–13191, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Graham S, Ding M, Sours-Brothers S, Yorio T, Ma JX, Ma R. Downregulation of TRPC6 protein expression by high glucose, a possible mechanism for the impaired Ca2+ signaling in glomerular mesangial cells. Am J Physiol Renal Physiol 293: F1381–F1390, 2007 [DOI] [PubMed] [Google Scholar]
- 27.Graham S, Gorin Y, Abboud HE, Ding M, Lee DY, Shi H, Ding Y, Ma R. Abundance of TRPC6 protein in glomerular mesangial cells is decreased by ROS and PKC in diabetes. Am J Physiol Cell Physiol 301: C304–C315, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gruden G, Setti G, Hayward A, Sugden D, Duggan S, Burt D, Buckingham RE, Gnudi L, Viberti G. Mechanical stretch induces monocyte chemoattractant activity via an NF-kappaB-dependent monocyte chemoattractant protein-1-mediated pathway in human mesangial cells: inhibition by rosiglitazone. J Am Soc Nephrol 16: 688–696, 2005 [DOI] [PubMed] [Google Scholar]
- 29.Ha H, Yu M, Kim KH. Melatonin and taurine reduce early glomerulopathy in diabetic rats. Free Radic Biol Med 26: 944–950, 1999 [DOI] [PubMed] [Google Scholar]
- 30.Ha H, Yu MR, Choi YJ, Kitamura M, Lee HB. Role of high glucose-induced nuclear factor-κB activation in monocyte chemoattractant protein-1 expression by mesangial cells. J Am Soc Nephrol 13: 894–902, 2002 [DOI] [PubMed] [Google Scholar]
- 31.Hall D, Carmines PK, Sansom SC. Dihydropyridine-sensitive Ca2+ channels in human glomerular mesangial cells. Am J Physiol Renal Physiol 278: F97–F103, 2000 [DOI] [PubMed] [Google Scholar]
- 32.Hewavitharana T, Deng X, Soboloff J, Gill DL. Role of STIM and Orai proteins in the store-operated calcium signaling pathway. Cell Calcium 42: 173–182, 2007 [DOI] [PubMed] [Google Scholar]
- 33.Hoth M, Penner R. Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature 355: 353–356, 1992 [DOI] [PubMed] [Google Scholar]
- 34.Hoth M, Penner R. Calcium release-activated calcium current in rat mast cells. J Physiol (Lond) 465: 359–386, 1993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Hulot JS, Fauconnier J, Ramanujam D, Chaanine A, Aubart F, Sassi Y, Merkle S, Cazorla O, Ouillé A, Dupuis M, Hadri L, Jeong D, Mühlstedt S, Schmitt J, Braun A, Bénard L, Saliba Y, Laggerbauer B, Nieswandt B, Lacampagne A, Hajjar RJ, Lompré AM, Engelhardt S. Critical role for stromal interaction molecule 1 in cardiac hypertrophy. Circulation 124: 796–805, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Iwamoto M, Mizuiri S, Arita M, Hemmi H. Nuclear factor-κB activation in diabetic rat kidney: evidence for involvement of P-selectin in diabetic nephropathy. Tohoku J Exp Med 206: 163–171, 2005 [DOI] [PubMed] [Google Scholar]
- 37.Kanwar YS, Akagi S, Sun L, Nayak B, Xie P, Wada J, Chugh SS, Danesh FR. Cell biology of diabetic kidney disease. Nephron Exp Nephrol 101: e100–e110, 2005 [DOI] [PubMed] [Google Scholar]
- 38.Kanwar YS, Wada J, Sun L, Xie P, Wallner EI, Chen S, Chugh S, Danesh FR. Diabetic nephropathy: mechanisms of renal disease progression. Exp Biol Med 233: 4–11, 2008 [DOI] [PubMed] [Google Scholar]
- 39.Kashgarian M, Sterzel RB. The pathobiology of the mesangium. Kidney Int 41: 524–529, 1992 [DOI] [PubMed] [Google Scholar]
- 40.Kikkawa R, Kitamura E, Fujiwara Y, Arimura T, Haneda M, Shigeta Y. Impaired contractile responsiveness of diabetic glomeruli to angiotensin II: a possible indication of mesangial dysfunction in diabetes mellitus. Biochem Biophys Res Commun 136: 1185–1190, 1986 [DOI] [PubMed] [Google Scholar]
- 41.Lee JE, Jeon IS, Han NE, Song HJ, Kim EG, Choi JW, Song KD, Lee HK, Choi JK. Ubiquilin 1 interacts with Orai1 to regulate calcium mobilization. Mol Cells 35: 41–46, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lewis RS. The molecular choreography of a store-operated calcium channel. Nature 446: 284–287, 2007 [DOI] [PubMed] [Google Scholar]
- 43.Li WP, Tsiokas L, Sansom SC, Ma R. Epidermal growth factor activates store-operated Ca2+ channels through an IP3 independent pathway in human glomerular mesangial cells. J Biol Chem 279: 4570–4577, 2004 [DOI] [PubMed] [Google Scholar]
- 44.Liao Y, Erxleben C, Abramowitz J, Flockerzi V, Zhu MX, Armstrong DL, Birnbaumer L. Functional interactions among orai1, TRPCs, and STIM1 suggest a STIM-regulated heteromeric Orai/TRPC model for SOCE/Icrac channels. Proc Natl Acad Sci USA 105: 2895–2900, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Liou J, Kim ML, Heo WD, Jones JT, Myers JW, Ferrell JE, Meyer T. STIM is a Ca2+ store sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr Biol 15: 1235–1241, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Luan JJ, Li W, Han J, Zhang W, Gong W, Ma R. Renal protection of in vivo administration of tempol in streptozotocin-induced diabetic rats. J Pharmacol Sci 119: 167–176, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ma R, Pluznick JL, Sansom SC. Ion channels in mesangial cells: function, malfunction, or fiction. Physiology 20: 102–111, 2005 [DOI] [PubMed] [Google Scholar]
- 48.Ma R, Sansom SC. Epidermal growth factor activates store-operated calcium channels in human glomerular mesangial cells. J Am Soc Nephrol 12: 47–53, 2001 [DOI] [PubMed] [Google Scholar]
- 49.Ma R, Smith S, Child A, Carmines PK, Sansom SC. Store-operated Ca2+ channels in human glomerular mesangial cells. Am J Physiol Renal Physiol 278: F954–F961, 2000 [DOI] [PubMed] [Google Scholar]
- 50.Mancarella S, Potireddy S, Wang Y, Gao H, Gandhirajan RK, Autieri M, Scalia R, Cheng Z, Wang H, Madesh M, Houser SR, Gill DL. Targeted STIM deletion impairs calcium homeostasis, NFAT activation, and growth of smooth muscle. FASEB J 27: 893–906, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Menè P, Pascale C, Teti A, Bernardini SV, Cinotti GA, Pugliese F. Effects of advanced glycation end products on cytosolic Ca2+ signaling of cultured human mesangial cells. J Am Soc Nephrol 10: 1478–1486, 1999 [DOI] [PubMed] [Google Scholar]
- 52.Menè P, Pugliese G, Pricci F, DiMario U, Cinotti GA, Pugliese F. High glucose level inhibits capacitative Ca2+ influx in cultured rat mesangial cells by a protein kinase C-dependent mechanism. Diabetologia 40: 521–527, 1997 [DOI] [PubMed] [Google Scholar]
- 53.Menè P, Pugliese G, Pricci F, DiMario V, Cinotti GA, Pugliese F. High glucose inhibits cytosolic calcium signaling in cultured rat mesangial cells. Kidney Int 43: 585–591, 1993 [DOI] [PubMed] [Google Scholar]
- 54.Menè P, Simonson MS, Dunn MJ. Physiology of mesangial cell. Physiol Rev 69: 1347–1424, 1989 [DOI] [PubMed] [Google Scholar]
- 55.Menè P, Teti A, Pugliese F, Cinotti GA. Calcium release-activated calcium influx in cultured human mesangial cells. Kidney Int 46: 122–128, 1994 [DOI] [PubMed] [Google Scholar]
- 56.Mezzano S, Aros C, Droguett A, Burgos ME, Ardiles L, Flores C, Schneider H, Ruiz-Ortega M, Egido J. NF-κB activation and overexpression of regulated gene in human diabetic nephropathy. Nephrol Dial Transplant 19: 2505–2512, 2004 [DOI] [PubMed] [Google Scholar]
- 57.Misra RP. Isolation of glomeruli from mammalian kidneys by graded sieving. Am J Clin Pathol 58: 135–139, 1972 [DOI] [PubMed] [Google Scholar]
- 58.Nutt LK, O'Neil RG. Effect of elevated glucose on endothelin-induced store-operated and non-store-operated calcium influx in renal mesangial cells. J Am Soc Nephrol 11: 1225–1235, 2000 [DOI] [PubMed] [Google Scholar]
- 59.Parekh AB, Putney JW., Jr Store-operated calcium channels. Physiol Rev 85: 757–810, 2005 [DOI] [PubMed] [Google Scholar]
- 60.Peng H, Liu J, Chen R, Wang Y, Duan J, Li C, Li B, Jing Y, Chen X, Mao Q, Xu KF, Walker CL, Li J, Wang J, Zhang H. mTORC1 enhancement of STIM1-mediated store-operated Ca2+ entry constrains tuberous sclerosis complex-related tumor development. Oncogene 32: 4702–4711, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Prakriya M, Feske S, Gwack Y, Srikanth S, Rao A, Hogan PG. Orail1 is an essential pore subunit of the CRAC channel. Nature 443: 230–233, 2006 [DOI] [PubMed] [Google Scholar]
- 62.Prakriya M, Lewis RS. Separation and characterization of currents through store-operated CRAC channels and Mg2+-inhibited cation (MIC) channels. J Gen Physiol 119: 487–508, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Putney JW., Jr Recent breakthroughs in the molecular mechanism of capacitative calcium entry (with thoughts on how we got here). Cell Calcium 42: 103–110, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Putney JW., Jr A model for receptor-regulated calcium entry. Cell Calcium 7: 1–12, 1986 [DOI] [PubMed] [Google Scholar]
- 65.Reed MJ, Meszaros K, Entes LJ, Claypool MD, Pinkett JG, Reaven GM. A new rat model of type 2 diabetes: the fat-fed, streptozotocin-treated rat. Metabolism 49: 1390–1394, 2000 [DOI] [PubMed] [Google Scholar]
- 66.Rice LV, Bax HJ, Russell LJ, Barrett VJ, Walton SE, Deakin AM, Thomson SA, Lucas F, Solari R, Hous D, Begg M. Characterization of selective calcium-release activated calcium channel blockers in mast cells and T-cells from human, rat, mouse and guinea-pig preparations. Eur J Pharmacol 704: 49–57, 2013 [DOI] [PubMed] [Google Scholar]
- 67.Roos J, DiGregorio PJ, Yeromin AV, Ohlsen K, Lioudyno M, Zhang S, Safrina O, Kozak JA, Wagner SL, Cahalan MD, Velicelebi G, Stauderman KA. STIM1, an essential and conserved component of store-operated Ca2+ channel function. J Cell Biol 169: 435–445, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Schlöndorff D, Bana B. The mesangial cell revisited: no cell is an island. J Am Soc Nephrol 20: 1179–1187, 2009 [DOI] [PubMed] [Google Scholar]
- 69.Scindia YM, Deshmukh US, Bagavant H. Mesangial pathology in glomerular disease: targets for therapeutic intervention. Adv Drug Deliv Rev 62: 1337–1343, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Soboloff J, Spassova M, Xu W, He LP, Cuesta N, Gill DL. Role of endogenous TRPC6 channels in Ca2+ signal generation in A7r5 smooth muscle cells. J Biol Chem 280: 39786–39794, 2005 [DOI] [PubMed] [Google Scholar]
- 71.Song JH, Jung SY, Hong SB, Kim MJ, Suh CK. Effects of high glucose on basal intracellular calcium regulation in rat mesangial cell. Am J Nephrol 23: 343–352, 2003 [DOI] [PubMed] [Google Scholar]
- 72.Sours S, Du J, Chu S, Zhou JX, Ding M, Ma R. Expression of canonical transient receptor potential (TRPC) proteins in human glomerular mesangial cells. Am J Physiol Renal Physiol 290: F1507–F1515, 2006 [DOI] [PubMed] [Google Scholar]
- 73.Sours-Brothers S, Ding M, Graham S, Ma R. Interaction between TRPC1/TRPC4 assembly and STIM1 contributes to store-operated Ca2+ entry in mesangial cells. Exp Biol Med 234: 673–682, 2009 [DOI] [PubMed] [Google Scholar]
- 74.Stockand JD, Sansom SC. Glomerular mesangial cells: electrophysiology and regulation of contraction. Physiol Rev 78: 723–744, 1998 [DOI] [PubMed] [Google Scholar]
- 75.Sugano M, Yamato H, Hayashi T, Ochiai H, Kakuchi J, Goto S, Nishijima F, Lino N, Kazama JJ, Takeuchi T, Mokuda O, Ishikawa T, Okazaki R. High-fat diet in low-dose-streptozotocin-treated heminephrectomized rats induces all features of human type 2 diabetic nephropathy: a new rat model of diabetic nephropathy. Nutr Metab Cardiovasc Dis 16: 477–484, 2006 [DOI] [PubMed] [Google Scholar]
- 76.The Diabetes Control and Complications Trial Research Group. The effect of intensive treatment of diabetes on the development and progression of long-term complications in insulin-dependent diabetes mellitus. N Engl J Med 329: 977–986, 1993 [DOI] [PubMed] [Google Scholar]
- 77.Trebak M, Bird GS, Mckay RR, Putney JW., Jr Comparison of human TRPC3 channels in receptor-activated and store-operated modes Differential sensitivity to channel blockers suggests fundamental differences in channel composition. J Biol Chem 277: 21617–21623, 2002 [DOI] [PubMed] [Google Scholar]
- 78.Vig M, Peinelt C, Beck A, Koomoa DL, Rabah D, Koblan-Huberson M, Kraft S, Turner H, Fleig A, Penner R, Kinet JP. CRACM1 is a plasma membrane protein essential for store-operated Ca2+ entry. Science 312: 1220–1223, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Voelkers M, Salz M, Herzog N, Frank D, Dolatabadi N, Frey N, Gude N, Friedrich O, Koch WJ, Katus HA, Sussman MA, Most P. Orai1 and Stim1 regulate normal and hypertrophic growth in cardiomyocytes. J Mol Cell Cardiol 48: 1329–1334, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Wang Y, Deng X, Gill DL. Calcium signaling by STIM and Orai: intimate coupling details revealed. Sci Signal 3: pe42-1–pe42-4, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wang Y, Ding M, Chaudhari S, Ding Y, Yuan J, Stankowska D, He S, Krishnamorthy R, Cunningham JT, Ma R. Nuclear factor κB mediates suppression of canonical transient receptor potential 6 expression by reactive oxygen species and protein kinase C in kidney cells. J Biol Chem 288: 12852–12865, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Weigert C, Sauer U, Brodbeck K, Pfeiffer A, Häring HU, Schleicher ED. AP-1 proteins mediate hyperglycemia-induced activation of the human TGF-β1 promoter in mesangial cells. J Am Soc Nephrol 11: 2007–2016, 2000 [DOI] [PubMed] [Google Scholar]
- 83.Welsh DG, Morielli AD, Nelson MT, Brayden JE. Transient receptor potential channels regulate myogenic tone of resistance arteries. Circ Res 90: 248–250, 2002 [DOI] [PubMed] [Google Scholar]
- 84.Whiteside CI, Hurst RD, Stevanovic ZS. Calcium signaling and contractile response of diabetic glomerular mesangial cells. Kidney Int 48: S28–S33, 1995 [PubMed] [Google Scholar]
- 85.Williams RT, Manji SSM, Parker NJ, Hancock MS, van Stekelenburg L, Eid JP, Senior PV, Kazenwadel J, Shandala T, Saint R, Smith PJ. Identification and characterization of the STIM (stromal interaction molecule) gene family: coding for a novel class of transmembrane proteins. Biochem J 357: 673–685, 2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zbidi H, Lopez JJ, Amor NB, Bartegi A, Salido GM, Rosado JA. Enhanced expression of STIM1/Orai1 and TRPC3 in platelets from patients with type 2 diabetes mellitus. Blood Cells Mol Dis 43: 211–213, 2009 [DOI] [PubMed] [Google Scholar]
- 87.Zhang SL, Yeromin AV, Zhang XHF, Yu Y, Safrina O, Penna A, Roos J, Stauderman KA, Cahalan MD. Genome-wide RNAi screen of Ca2+ influx identifies genes that regulate Ca2+ release-activated Ca2+ channel activity. Proc Natl Acad Sci USA 103: 9357–9362, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ziyadeh FN, Sharma K. Role of transforming growth factor-β in diabetic glomerulosclerosis and renal hypertrophy. Kidney Int 48: S34–S36, 1995 [PubMed] [Google Scholar]