

Abstract
Genetic polymorphisms in genes related to neurotransmitters or hormones affect personality or behavioral traits in many animal species including humans. In domestic animals, the allele frequency of such genes has been reported to be different among breeds and it may account for breed differences in behavior. In this study, we investigated breed differences in horses in the dopamine receptor D4 gene (DRD4), which has been reported to affect horse personality. We collected samples from seven horse breeds including those native to Japan and Korea, and compared the sequence of the DRD4 exon3 region among these breeds. We found that there were two types of polymorphisms (VNTR and SNPs) in the exon3 region, and some of them seemed to be breed-specific. In addition, we found that the allele frequency of G292A, reported to be associated with horse personality, differed greatly between native Japanese horses and Thoroughbred horses. The frequency of the A allele which is associated with low curiosity and high vigilance, was much lower in native Japanese horses (Hokkaido, 0.03; Taishu, 0.08) than in Thoroughbreds (0.62). This difference may account for breed differences in personality or behavioral traits. Further studies of the function of these polymorphisms and their effect on behavior are indicated.
Keywords: behavioral trait, breed difference, dopamine receptor D4 gene, native Japanese horse, SNP
Selective breeding by humans has resulted in various breeds of domestic animals. Great diversity between different breeds has been created not only in body size and coat color, but also in behavioral traits and personality. For example, dogs of different breeds differ greatly in social behavioral traits [9, 15]. Differences between breeds in personality, as measured by questionnaire surveys, are also reported in horses [11]. Breed differences in behaviors are especially important in companion animals such as dogs or horses. Scientific evaluation of behavioral differences between breeds should be useful for the development of methods of management or training better suited to each breed.
Variability of behavioral traits between breeds may be partly affected by genes. Recent studies have shown that genetic polymorphisms in genes related to neurotransmitters or hormones influence individual differences in personality [7]. The dopamine receptor D4 gene (DRD4) encodes one of the subtypes of dopamine receptor, and is one of the candidate genes underlying individual differences in personality in many species including humans. In human DRD4 there is a variable number of tandem repeats (VNTR), which is reported to be associated with a “novelty-seeking” trait [1, 2].
In dogs, breed differences in the allele frequency of DRD4 have been reported. For example, a VNTR in the canine DRD4 exon3 region is reported to be associated with aggressiveness and activity/impulsivity [5]. The allele frequency of this polymorphism is different among breeds [8], particularly between Asian breeds and European breeds [10].
In horses, DRD4 is located on chromosome 12 [13], and two types of polymorphisms have been found. They are VNTR consisting of 18 base pairs (6 amino acids) and some single nucleotide polymorphisms (SNPs) in the exon3 region [3]. One of these SNPs, G292A, was reported to be associated with horse personality scores measured by a questionnaire survey. The A allele in G292A is associated with low curiosity and high vigilance in Thoroughbred horses [12].
If there are breed differences in personality related genes such as DRD4, it may be possible to predict the behavioral traits of breeds based on the genotype. A previous study reported that one Kiso horse, a native Japanese horse breed, had shorter repeats in the VNTR region than Thoroughbred horses [3]. However, whether the allele frequency of this polymorphism differs between breeds has not yet been verified. Samples from more breeds are needed to validate the breed differences of DRD4 in horses.
In this study, we investigated breed differences in equine DRD4 using samples from more breeds than previously studied. We collected samples from two breeds of European origin (Thoroughbred and Selle Français), one South American breed (Criollo), three native Japanese breeds (Hokkaido, Taishu and Yonaguni) and one native Korean breed. We explored the polymorphisms in DRD4 and compared its allele frequency among breeds.
Materials and Methods
In total, we collected 70 horse samples including two European-origin breeds (Thoroughbred; n=4, Selle Français; n=1), one South American-origin breed (Criollo; n=5), three native Japanese breeds, and a native Korean breed. The Japanese samples were from Hokkaido (n=31), Taishu (Tsushima; n=20), and Yonaguni (n=1). The Korean samples were from Jeju in Korea (n=8). We excluded offspring or full-siblings from our samples when estimating the allele frequency of each breed.
We extracted genomic DNA from blood, buccal swabs, or hair using a QIAamp blood and tissue kit (QIAGEN, Valencia, CA, USA). For some hair samples, an InstaGeneTM Matrix (Bio-Rad Laboratories, Hercules, CA, USA) was used.
We amplified 504 base pairs of the exon3 region in equine DRD4 (GenBank: AB080626.1) by polymerase chain reaction (PCR). We used the primer pairs and PCR conditions described in a previous study [12]. PCR products were purified using a High Pure PCR Product Purification Kit (Roche, Basel, Switzerland), and their nucleotide sequences were determined by cycle sequencing using a Big Dye Terminator v3. 1 Cycle Sequencing Kit and an Applied Biosystems 3130xl Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). All determined sequences were checked visually using FinchTV software, version 1. 4. 0. Polymorphisms in the sequences (SNPs and VNTR) were detected by the alignment using MEGA software, version 5.
For SNPs found in more than one breed, the allele frequencies of each breed were statistically compared using Fisher’s exact test.
Results
In equine DRD4 exon3 region we found two types of polymorphisms, VNTR and SNPs. The sequence of the exon3 region and locations of polymorphisms we found are shown in Fig. 1. VNTR was found only in Criollo horses; these horses had a nine-repeat allele and a ten-repeat allele (Fig. 1). The ten-repeat allele was longer than previously reported alleles (nine and eight repeats), and its frequency was approximately 0.30. No length polymorphisms were found in the other breeds (all of them had the nine-repeat allele).
Fig. 1.

Sequences of the DRD4 exon3 region in four breeds (Criollo, Hokkaido, Taishu and Thoroughbred). Locations of SNPs are marked by squares. VNTR regions are marked by lines and bold numbers indicate the number of repeats. The native Korean breed had G292A and G445A polymorphisms. Novel sequences determined in this study (Criollo and Taishu) were deposited in the DDBJ database (Accession Nos. AB817803 and AB817804 ).
We also found six SNPs, three of which (C199T, C415T and G445A) have not previously been reported. Information about these SNPs is summarized in Table 1. The allele frequencies of each SNP in the five breeds (Criollo, Hokkaido, Korean, Taishu and Thoroughbred) are shown in Table 2.
Table 1. SNPs found in equine DRD4.
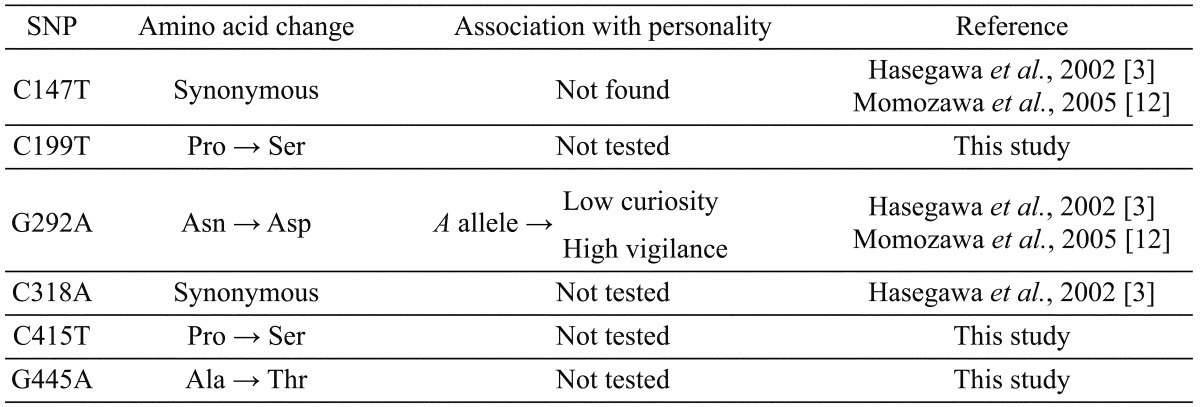
Table 2. The allele frequencies of SNPs in five breeds.
| Criollo | Hokkaido | Korean | Taishu | Thoroughbred | ||
|---|---|---|---|---|---|---|
| n | 5 | 31 | 8 | 20 | 4 | |
| VNTR | 9 | 0.70 | 1.00 | 1.00 | 1.00 | 1.00 |
| 10 | 0.30 | 0.00 | 0.00 | 0.00 | 0.00 | |
| He | 0.47 | 0.00 | 0.00 | 0.00 | 0.00 | |
| C147T | C | 1.00 | 0.97 | 1.00 | 1.00 | 0.88 |
| T | 0.00 | 0.03 | 0.00 | 0.00 | 0.12 | |
| He | 0.00 | 0.06 | 0.00 | 0.00 | 0.25 | |
| C199T | C | 0.70 | 1.00 | 1.00 | 1.00 | 1.00 |
| T | 0.30 | 0.00 | 0.00 | 0.00 | 0.00 | |
| He | 0.47 | 0.00 | 0.00 | 0.00 | 0.00 | |
| G292A | G | 0.70 | 0.97 | 0.62 | 0.92 | 0.38 |
| A | 0.30 | 0.03 | 0.38 | 0.08 | 0.62 | |
| He | 0.47 | 0.06 | 0.50 | 0.14 | 0.54 | |
| C318A | C | 1.00 | 1.00 | 1.00 | 0.72 | 1.00 |
| A | 0.00 | 0.00 | 0.00 | 0.28 | 0.00 | |
| He | 0.00 | 0.00 | 0.00 | 0.41 | 0.00 | |
| C415T | C | 1.00 | 1.00 | 1.00 | 0.65 | 1.00 |
| T | 0.00 | 0.00 | 0.00 | 0.35 | 0.00 | |
| He | 0.00 | 0.00 | 0.00 | 0.47 | 0.00 | |
| G445A | G | 0.90 | 0.81 | 0.88 | 0.90 | 1.00 |
| A | 0.10 | 0.19 | 0.12 | 0.10 | 0.00 | |
| He | 0.20 | 0.32 | 0.23 | 0.18 | 0.00 |
He: expected heterozygosity.
Among the SNPs we identified, C147T and C318A are synonymous SNPs, which cause no amino acid changes. We found the C147T polymorphism only in Hokkaido and Thoroughbred horses, and the C318A polymorphism only in Taishu horses. The nucleotide substitution reported at the C318A location in a previous study [3] was from cytosine to thymine, but the substitution we found in Taishu horses was from cytosine to adenine. In both cases, these substitutions cause no amino acid change.
The other four SNPs (C199T, G292A, C415T and G445A) are non-synonymous SNPs, which cause amino acid changes. C199T was unique to Criollo horses and C415T was unique to Taishu horses.
G292A is a non-synonymous SNP that causes an amino acid change from asparagine to aspartic acid, and it was linked to horse personality in a previous study [12]. The A allele of G292A is associated with low curiosity and high vigilance scores. We compared the allele frequency of G292A among five breeds (Hokkaido, Taishu, Korean, Criollo and Thoroughbred) and found a significant difference among breeds (Fisher’s exact test, P=4.4 × 10–6). In native Japanese breeds (Hokkaido and Taishu) the frequency of the A allele was very low (approximately 0.03 for Hokkaido, 0.08 for Taishu). In contrast, the frequency of the A allele was considerably higher (approximately 0.62) in Thoroughbreds. One Selle Français horse and one Yonaguni horse were heterozygous (their genotypes were A/G).
G445A is a novel SNP that causes an amino acid change from alanine to threonine. This polymorphism was found in four breeds (Hokkaido, Taishu, Korean and Criollo), but the allele frequencies did not differ significantly among these breeds (Fisher’s exact test, P=0.65).
The novel sequence data determined in this study was registered in the DDBJ nucleotide database with the accession numbers AB817803 (Criollo) and AB817804 (Taishu).
Discussion
In this study we found several novel polymorphisms in equine DRD4 exon3 region. Some of these seem to be breed-specific. Criollo horses had a unique non-synonymous SNP (C199T) and longer alleles (ten-repeat) in the VNTR region than other breeds. Taishu horses also had a unique non-synonymous SNP (C415T). These polymorphisms may change the amino acid sequence of the protein and modify its function. Functional analysis of these polymorphisms is now needed.
We also compared the allele frequencies of polymorphisms among different breeds. Although samples from unrelated individuals are needed to estimate the allele frequency more accurately, it is difficult to collect completely unrelated samples in the case of rare native breeds. So, we excluded offspring or full-siblings from our samples when estimating the allele frequency. We found that the allele frequency of G292A, which has been reported to be associated with horse personality, was different among breeds. In particular, native Japanese horses had quite different allele frequencies of G292A from Thoroughbred horses. Although our sample size of Thoroughbreds was relatively small, the A allele seems to be a major allele in these horses, because the frequency of this allele was also high in a previous study [12] (approximately 0.54). This allele was in the minority in all of the other breeds tested. This suggests that the A allele is a minor allele of horses in general, except for Thoroughbreds. It may be a byproduct of the special breeding history of Thoroughbred horses for racing performance. The low frequency of the A allele, linked with low curiosity and high vigilance, suggests that Japanese horses should be more curious and less vigilant than Thoroughbreds. In fact, a questionnaire-based behavioral evaluation of Kiso horses revealed these animals are curious and friendly with humans [14].
The differences of the A allele frequency of G292A may be used as a predictor of behavioral traits of horse breed. Also polymorphisms within each breed may be useful for management or training suited to each individual trait. However, the association between G292A and behavioral traits requires further confirmation using more objective behavioral tests. Behavioral tests used in canine studies showing the association between DRD4 and behavioral traits [4, 6] may be useful, following modification, for use with horses.
In conclusion, this study showed that there are differences in DRD4 polymorphisms among breeds of horses. These differences may be a result of different breeding histories or of geographical variation. They may affect the variance of behavioral traits or personality among the breeds. Further studies of both functional aspects of genes and behavioral variations are needed to validate the effect of these polymorphisms.
Acknowledgments
This study was supported financially by the Japan Society for the Promotion of Sciences (JSPS) with Grants-in-aid for Scientific Research (#21310150 to MI-M and #20220004 to KF) and the Asia and Africa Science Platform Program. This study was supported by the Cooperation Research Program of the Wildlife Research Center, Kyoto University. The authors would like to thank Dr. Shin’ichi Ito for cooperation with the collection of samples and suggestions useful to this research. We also would like to thank Ms. Tomoko Yabe (Ueno Zoo), Mr. Eiichiro Fujiwara (Tsurui Dosanko Farm), and the students of Shimabara Agricultural High School for their cooperation with the collection of samples.
References
- 1.Benjamin J., Li L., Patterson C., Greenberg B.D.1996. Population and familial association between the D4 dopamine receptor gene and measures of novelty seeking. Nat. Genet. 12: 81–84 [DOI] [PubMed] [Google Scholar]
- 2.Ebstein R.P., Novick O., Umansky R., Priel B.1996. Dopamine D4 receptor (D4DR) exon III polymorphism associated with the human personality trait of novelty seeking. Nat. Genet. 12: 78–80 [DOI] [PubMed] [Google Scholar]
- 3.Hasegawa T., Sato F., Ishida N.2002. Determination and variability of nucleotide sequences for D4 dopamine receptor genes (DRD4) in genus Equus. J. Equine Sci. 13: 57–62 [Google Scholar]
- 4.Hejjas K., Kubinyi E., Ronai Z., Szekely A., Vas J., Miklosi A., Sasvari-Szekely M., Kereszturi E.2009. Molecular and behavioral analysis of the intron 2 repeat polymorphism in the canine dopamine D4 receptor gene. Genes Brain Behav. 8: 330–336 [DOI] [PubMed] [Google Scholar]
- 5.Hejjas K., Vas J., Topal J., Szantai E., Ronai Z., Szekely A., Kubinyi E., Horvath Z., Sasvari-Szekely M., Miklosi A.2007. Association of polymorphisms in the dopamine D4 receptor gene and the activity-impulsivity endophenotype in dogs. Anim. Genet. 38: 629–633 [DOI] [PubMed] [Google Scholar]
- 6.Hori Y., Kishi H., Inoue-Murayama M., Fujita K.2013. Dopamine receptor D4 gene (DRD4) is associated with gazing toward humans in domestic dogs (Canis familiaris). Open J. Anim. Sci. 3: 54–58 [Google Scholar]
- 7.Inoue-Murayama M.2009. Genetic polymorphism as a background of animal behavior. Anim. Sci. J. 80: 113–120 [DOI] [PubMed] [Google Scholar]
- 8.Ito H., Nara H., Inoue-Murayama M., Shimada M.K., Koshimura A., Ueda Y., Kitagawa H., Takeuchi Y., Mori Y., Murayama Y., Morita M., Iwasaki T., Ota K., Tanabe Y., Ito S.2004. Allele frequency distribution of the canine dopamine receptor D4 gene exon III and I in 23 breeds. J. Vet. Med. Sci. 66: 815–820 [DOI] [PubMed] [Google Scholar]
- 9.Jakovcevic A., Elgier A.M., Mustaca A.E., Bentosela M.2010. Breed differences in dogs’ (Canis familiaris) gaze to the human face. Behav. Processes 84: 602–607 [DOI] [PubMed] [Google Scholar]
- 10.Koshimura A., Inoue-Murayama M., Ueda Y., Maejima M., Kitagawa H., Morita M., Iwasaki T., Murayama Y., Kim H.S., Ha J.H., Randi E., Kansaku N., Tanabe Y., Ota K., Ito S.2006. Genetic diversity in dog breeds of Asian origin and Asian native dogs. Rep. Soc. Res. Native Livest. 23: 189–207 [Google Scholar]
- 11.Lloyd A.S., Martin J.E., Bornett-Gauci H.L.I., Wilkinson R.G.2008. Horse personality: variation between breeds. Appl. Anim. Behav. Sci. 112: 369–383 [Google Scholar]
- 12.Momozawa Y., Takeuchi Y., Kusunose R., Kikusui T., Mori Y.2005. Association between equine temperament and polymorphisms in dopamine D4 receptor gene. Mamm. Genome 16: 538–544 [DOI] [PubMed] [Google Scholar]
- 13.Momozawa Y., Takeuchi Y., Tozaki T., Kikusui T., Hasegawa T., Raudsepp T., Chowdhary B.P., Kusunose R., Mori Y.2007. SNP detection and radiation hybrid mapping in horses of nine candidate genes for temperament. Anim. Genet. 38: 81–83 [DOI] [PubMed] [Google Scholar]
- 14.Mukoyama H., Endo M., Kuroda K., Furuta H., Yoshida T., Yoshimura I.2009. Evaluation as therapeutics horse of Kiso pony by behavioral related gene (DRD4 gene) polymorphism. DNA Polymorph 17: 36–39 [Google Scholar]
- 15.Passalacqua C., Marshall-Pescini S., Barnard S., Lakatos G., Valsecchi P., Prato Previde E.2011. Human-directed gazing behaviour in puppies and adult dogs, Canis lupus familiaris. Anim. Behav. 82: 1043–1050 [Google Scholar]