

Abstract
The RpoN-RpoS regulatory pathway plays a central role in governing adaptive changes by B. burgdorferi when the pathogen shuttles between its tick vector and mammalian hosts. In general, transcriptional activation of bacterial RpoN (σ54)-dependent genes requires an enhancer binding protein. B. burgdorferi encodes the putative enhancer binding protein Rrp2. Previous studies have revealed that the expression of σ54-dependent rpoS was abolished in an rrp2 point mutant. However, direct evidence linking the production of Rrp2 in B. burgdorferi and the expression of rpoS has been lacking, primarily due to the inability to inactivate rrp2 via deletion or insertion mutagenesis. Herein we introduced a regulatable (IPTG-inducible) rrp2 expression shuttle plasmid into B. burgdorferi, and found that the controlled up-regulation of Rrp2 resulted in the induction of σ54-dependent rpoS expression. Moreover, we created an rrp2 conditional lethal mutant in virulent B. burgdorferi. By exploiting this conditional mutant, we were able to experimentally manipulate the temporal level of Rrp2 expression in B. burgdorferi, and examine its direct impact on activation of the RpoN-RpoS regulatory pathway. Our data revealed that the synthesis of RpoS was coincident with the IPTG-induced Rrp2 levels in B. burgdorferi. In addition, the synthesis of OspC, a lipoprotein required by B. burgdorferi to establish mammalian infection, was rescued in the rrp2 point mutant when RpoS production was restored, suggesting that Rrp2 influences ospC expression indirectly via its control over RpoS. These data demonstrate that Rrp2 is required for the synthesis of RpoS, presumably via its action as an enhancer binding protein for the activation of RpoN and subsequent transcription of rpoS in B. burgdorferi.
Introduction
Borrelia burgdorferi, the causative agent of Lyme disease, is sustained in nature via a complex life cycle involving an arthropod tick vector (Ixodes scapularis) and mammals [1], [2]. During its transit between these two markedly different host and tick milieus, B. burgdorferi undergoes significant adaptive changes. In B. burgdorferi, host adaptation is achieved by dramatic changes in gene expression in response to various tick or host stimuli [3]–[9]. Among a number of potential regulators that have been postulated to be present in B. burgdorferi [10]–[45], a novel genetic regulatory pathway, the RpoN-RpoS pathway (or the σ54-σS cascade) [18], plays a central role in modulating B. burgdorferi host adaptive responses and virulence expression. In this pathway, one alternative sigma factor (σ54, RpoN) controls the expression of another alternative sigma factor (σS, RpoS) through binding to a canonical −24/−12 promoter sequence [12], [26], [40]. Once RpoS is produced, it functions as a master regulator to modulate the expression of a number of virulence-associated outer membrane lipoproteins such as outer surface lipoproteins (Osp) C and A, and decorin binding proteins (Dbp) B and A [4], [8], [12]–[14], [18], [29], [32], [36], [40], [46]–[57].
Transcriptional activation of σ54-dependent genes in bacteria requires a bacterial enhancer binding protein (bEBP), which is an AAA+ activator ATPase [58]–[63]. Sequence analyses have indicated that Rrp2 (BB0763) is composed of three structural domains, including an N-terminal regulatory domain (R), a central AAA+ ATPase core domain (C), and a C-terminal DNA binding domain (D), suggesting that Rrp2 may function as a bEBP to activate σ54–dependent rpoS transcription in B. burgdorferi [15], [45]. Previously, by exploiting a variant carrying a point mutation G239C in the C domain of Rrp2, we [10], [29], [45] reported that the rrp2 point mutant was incapable of expressing rpoS and virulence-associated factors such as OspC and DbpA, suggesting that, as expected, Rrp2 is essential for activation of the RpoN-RpoS regulatory pathway. Despite this important finding, there remain many unanswered questions concerning the roles of Rrp2 in B. burgdorferi gene regulation. In particular, the finding that expression of rpoS was abolished in the rrp2 point mutant has been hitherto the only evidence to support the role of Rrp2 in the activation of the RpoN-RpoS pathway. A direct link between Rrp2 protein levels produced in B. burgdorferi and the expression of rpoS has been lacking, primarily due to the inability to inactivate rrp2 via deletion or insertion mutagenesis. The G239C point mutation in rrp2 presumably abolishes the putative ATPase activity required for σ54–dependent rpoS transcriptional activation. However, it also remains possible that the G239C point mutation causes a change in Rrp2’s overall conformation, thereby preventing rpoS transcription in the rrp2 point mutant. In addition, although the expression of ospC was found to be lost in the rrp2 point mutant, how Rrp2 ultimately controls the expression of this key virulence-associated lipoprotein remains unknown. Rrp2 may indirectly modulate ospC expression via its control over RpoS. Alternatively, given that ospC was found to be constitutively expressed in B. burgdorferi when ospAB and rpoS were inactivated [64], Rrp2 may control ospC expression through another RpoS-independent factor(s). It is also possible that Rrp2 may directly modulate ospC expression by binding to its promoter. To address these questions, we employed an artificial gene expression system [65], [66] to experimentally control the protein levels of Rrp2 synthesized in B. burgdorferi, and examined its impact on rpoS expression. Such a strategy affects only the levels of Rrp2 produced in B. burgdorferi, and does not alter the overall structure of the protein. Our data show that the expression level of Rrp2 correlates closely with the expression of rpoS, indicating that Rrp2 activates the expression of σ54–dependent rpoS which, in turn, modulates ospC expression in B. burgdorferi.
Materials and Methods
Bacterial Strains and Culture Conditions
All strains and plasmids used in this study are described in Table 1. Low-passage infectious wild-type B. burgdorferi strain 297 [67], and the rrp2 point mutant OY01 [29], were routinely cultured at 37°C and 5% CO2 in either BSK-II medium [68] or BSK-H medium (Sigma) supplemented with 6% rabbit serum (Pel-Freeze). When appropriate, supplements were added to media at the following concentrations: erythromycin, 60 ng/ml; kanamycin, 150 µg/ml; streptomycin, 100 µg/ml. Spirochetes were enumerated by dark-field microscopy. E. coli strains were cultured in Lysogeny Broth (LB) supplemented with appropriate antibiotics at the following concentrations: ampicillin, 100 µg/ml; kanamycin, 50 µg/ml; spectinomycin, 100 µg/ml.
Table 1. Strains and plasmids used in this study.
| Strains or plasmids | Description | Source |
| B. burgdorferi | ||
| 297 | Infectious B. burgdorferi, human spinal fluid isolate | [67] |
| OY01 | 297, rrp2(G239C) point mutant | [29] |
| AH206 | 297, rpoS::ermC | [18] |
| OY159 | 297 transformed with pRrp2 | This study |
| OY160 | OY01 transformed with pRrp2 | This study |
| OY173 | OY01 transformed with pRrp2-FLAG | This study |
| OY179 | rrp2 conditional lethal mutant, OY173 transformed with pOY202 | This study |
| E. coli Top10F′ | F′[lacI q Tn10(Tetr)] mcrA Δ(mrr-hsdRMS-mcrBC) φ80lacZΔM15ΔlacX74 recA1 araΔ139 Δ (ara-leu)7697 galU galK rpsL(Strr) endA1 nupG | Invitrogen |
| Plasmids | ||
| pGEM-Teasy | TA cloning vector; Ampr | Promega |
| pJSB275 | Shuttle vector, Spec/Strepr | [69] |
| pRpoS | IPTG-inducible rpoS expression construct | [30] |
| pOY100 | PCR product of 86F/87R cloned into pGEM-Teasy, ampr | This study |
| pJD55 | Shuttle vector, Spec/Strepr | [46] |
| pOY202 | PflgB-kan cloned into pOY100, Amp/Kanr | This study |
| pRrp2 | PCR product of 303F/303R cloned into pJSB275, Spec/Strepr | This study |
| pRrp2-FLAG | PCR product of 303F/263R cloned into pJSB275, Spec/Strepr | This study |
Generation of the IPTG-inducible rrp2 Expression Construct
To experimentally control rrp2 expression in B. burgdorferi, one rrp2 expression construct pRrp2 was generated by using the lac-based gene inducible expression system [65], [66]. Briefly, rrp2 was amplified from B. burgdorferi using primers ZM303F and 303R (Table S1), and then cloned into pJSB275 [69] at the NdeI site. In pRrp2, rrp2 transcription is directly controlled by the IPTG-inducible T5 promoter (a.k.a. the promoter PpQE30) from plasmid pQE30 (Qiagen).
Generation of the rrp2 Conditional Lethal Mutant OY179
To create an rrp2 conditional mutant, we first generated one shuttle plasmid pRrp2-FLAG in which rrp2 expression was placed under the control of the IPTG-inducible promoter PpQE30. For creating pRrp2-FLAG, a DNA fragment encoding Rrp2-FLAG was amplified by using ZM303F and ZM263R (Table S1), and cloned into pJSB275 [69] at the NdeI site. This strategy adds a FLAG-tag (DYKDDDDK) to the C-terminus of Rrp2, thereby facilitating the detection of Rrp2 in B. burgdorferi. The plasmid pRrp2-FLAG was then electroporated into the rrp2 point mutant OY01, yielding the streptomycin-resistant strain OY173. B. burgdorferi transformation was performed as previously described [29], [70].
To inactivate rrp2 in B. burgdorferi through homologous recombination, a suicide plasmid pOY202 was created. Briefly, the left arm for creating pOY202 was PCR-amplified using primers ZM86F and ZM86R, whereas the right arm was amplified using ZM87F and ZM87R (Table S1). After digestion with AscI, these two fragments were fused together. By using this ligated DNA as the template, PCR was employed to amplify a fragment comprising the upstream and downstream regions of rrp2 by using primers ZM86F and ZM87R. The obtained fragment was cloned into pGEM-Teasy vector (Promega), yielding pOY100. The PflgB-Kan cassette, excised from pJD55 [46] using AscI, was cloned into pOY100 at the AscI site. In the resulting construct pOY202, the PflgB-Kan cassette was inserted in rrp2 in the opposite direction as transcription of csrA. All constructs were confirmed using PCR amplification, restriction digestion, and sequence analysis. The plasmid pOY202 was then transformed into B. burgdorferi strain OY173. Transformants were isolated in the presence of kanamycin and streptomycin, along with 0.05 mM of IPTG to allow the production of Rrp2-FLAG during selection.
RNA Isolation and qRT-PCR
RNA isolation and qRT-PCR were performed as previously described [29]–[31], [71]. Briefly, when total RNA was isolated from B. burgdorferi by using Trizol (Invitrogen), RNase-free DNase I (GenHunter Corporation) was used to digest genomic DNA. After RNA was further purified using RNeasy Mini Kit (Qiagen), 1 µg of RNA was used to synthesize cDNA using the SuperScript III Platinum Two-step qRT-PCR kit according to the manufacturer’s protocol (Invitrogen). qRT-PCR was employed to examine gene expression, using the relative quantification method (ΔΔCT) as described [29]–[31], [71]. Gene expression fold change was presented as mean ± SE values from three independent experiments. Statistical analyses of the data were performed using the Student’s t test.
SDS-PAGE and Semi-quantitative Immunoblot Analyses
A volume of whole cell lysate equivalent to 4×107 spirochetes was loaded per lane onto a 12.5% acrylamide gel. Resolved proteins were either stained with Coomassie brilliant blue or transferred to nitrocellulose membrane for immunoblot analysis as previously described [29]–[31], [71]. Rrp2, FLAG, RpoS, OspC, and DbpA were detected using anti-Rrp2 monoclonal antibody 5B8-100-A1, anti-FLAG M2 monoclonal antibody (Sigma), anti-RpoS monoclonal antibody 6A7-101, anti-OspC monoclonal antibody 1B2-105A, or anti-DbpA monoclonal antibody 6B3, respectively. Immunoblots were developed colorimetrically using 4-chloro-1-napthol as the substrate or by chemiluminescence using the ECL Plus Western Blotting Detection system (Amersham Biosciences). Images were documented by using a Fujifilm LAS-3000 Imager (Fujifilm), and semi-quantitative analyses were performed by using the MultiGauge V3.0 software (Fujifilm).
Results and Discussion
Complementation of the rrp2 Point Mutation by using an IPTG-inducible rrp2 Expression Construct
Previously, we introduced a G239C mutation into rrp2 and created an rrp2 point mutant OY01 (rrp2[G239C]) in B. burgdorferi [29]. The expression of rpoS and ospC is abolished in OY01. To confirm that the loss of rpoS and ospC expression in the rrp2 point mutant was due solely to the mutation of rrp2, a trans-complementation approach was employed. To this end, an IPTG-inducible plasmid pRrp2 (Fig. 1A) was created by placing the expression of rrp2 under the control of the IPTG-inducible PpQE30 promoter. By adjusting the amount of the inducer (IPTG) added to the medium, the expression of rrp2 on pRrp2 could be controlled. pRrp2 was then introduced into the rrp2 point mutant OY01, yielding the streptomycin-resistant strain OY160.
Figure 1. Trans-complementation of an rrp2 point mutant using a shuttle plasmid.
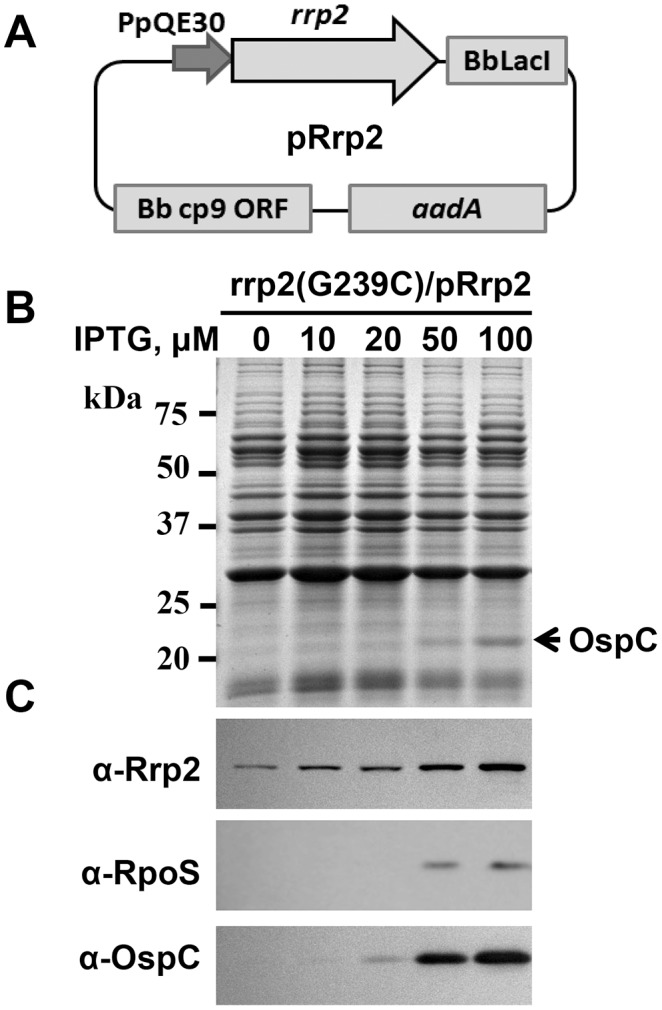
(A) Construction of an IPTG-inducible rrp2 expression shuttle plasmid. To create pRrp2, rrp2 was amplified from B. burgdorferi and cloned into pJSB275. The plasmid pRrp2 was then introduced into strain OY01 (rrp2[G239C]), yielding OY160. SDS-PAGE (B) and immunoblot (C) were performed to analyze gene expression in OY160. Bacteria were grown at 37°C in BSK-II medium with various concentrations of IPTG. When bacterial growth reached ∼108 cells per ml, spirochetes were harvested. Approximately 4×107 spirochetes were loaded onto each lane of a 12.5% SDS-PAGE gel. In (B), approximate molecular masses are indicated at the left in kDa; concentrations of IPTG are indicated above the image; and the arrow indicates OspC. Specific antibodies, denoted as α- used in the immunoblot (C), are indicated on the left.
To induce the expression of wild-type Rrp2, OY160 was grown in BSK-II medium containing varying concentrations of IPTG. In this experiment and subsequently, bacterial growth and morphology were not affected when spirochetes were grown in media with indicated levels of IPTG. Cells were collected when bacterial growth reached early stationary phase (∼108 cells/ml). As shown in Fig. 1C, when 10-, 20-, 50-, or 100-µM of IPTG was added into the media, synthesis of Rrp2 was enhanced in a dose-dependent manner in B. burgdorferi. Of note, when IPTG was not added into the medium, a band was also detected in immunoblot by using the Rrp2 antibody; this band represents the existing mutated protein Rrp2(G239C). The induction of Rrp2 also resulted in the synthesis of RpoS and OspC in OY160. As shown in Fig. 1B and Fig. 1C, when bacteria were grown in BSK-II containing 50- or 100-µM of IPTG, RpoS and OspC were readily detected in OY160, and the increased protein levels correlated well with the increased levels of Rrp2 in OY160. These data suggest that Rrp2 expressed from pRrp2 is capable of complementing the rrp2(G239C) point mutation, thereby activating the RpoN-RpoS pathway in B. burgdorferi.
Up-regulation of Rrp2 in B. Burgdorferi Induces the Expression of RpoS and OspC
The approach of gene overexpression has proven to be a highly valuable tool for examining gene functions, particularly for genes that cannot be inactivated [72]. This strategy was successfully employed previously to study the role of Rrp2 in B. burgdorferi, where it was reported that overexpression of Rrp2 in B. burgdorferi led to the induction of OspC [69]. However, it remained unknown whether RpoS synthesis was influenced by the overexpression of Rrp2 in B. burgdorferi. In this current study, the IPTG-inducible rrp2 expression construct pRrp2 was introduced into B. burgdorferi wild-type strain 297, yielding the merodiploid strain OY159. Gene expression in these spirochetes was measured through SDS-PAGE, semi-quantitative immunoblot, and quantitative RT-PCR analyses. In this experiment, two strategies employing shorter or longer induction times were used to induce Rrp2 in B. burgdorferi. For the longer induction time, bacteria were continuously grown in BSK-II containing various amounts of IPTG for about 7 days and harvested when growth reached the early stationary phase (∼108 cells/ml). As shown in Fig. 2B, synthesis of Rrp2 in OY159 was enhanced when bacteria were grown in media containing IPTG (compared with spirochetes cultivated in media without IPTG). Moreover, synthesis of RpoS and OspC was also found to be enhanced when IPTG was added into the medium (Fig. 2A, 2B). To exclude the possibility that gene induction might be an indirect effect resulting from prolonged exposure to IPTG, a shorter (9 h) induction period was also examined. When bacterial growth in BSK-II reached mid-log phase (∼107 cells/ml), varying amounts of IPTG were added into the media. After 9 h of induction, spirochetes were collected and gene expression was examined. As shown in Fig. 2C, immunoblot analyses showed that the syntheses of both Rrp2 and RpoS were induced by IPTG in a dose-dependent manner. Gene expression was also measured by using real-time quantitative RT-PCR (qRT-PCR) analyses. When gene transcription in spirochetes grown in BSK-II with 20-, 100-, 200-, or 500-µM of IPTG was compared with gene expression in bacteria grown without IPTG, transcription of rrp2 was induced at 1.3-, 8.8-, 12.4-, or 18.9-fold, respectively; accordingly, rpoS transcription was induced at 0.9-, 5.6-, 9.4-, or 17.1-fold, respectively. These combined data strongly suggest that Rrp2 is responsible for the induction of RpoS in B. burgdorferi.
Figure 2. Up-regulation of Rrp2 in B. burgdorferi induces the expression of rpoS.

Gene expression in OY159 was analyzed by SDS-PAGE (A), immunoblot (B, C), and qRT-PCR analyses (D). In (A) and (B), spirochetes grown in BSK-II media containing varying concentrations of IPTG were harvested when bacterial growth reached early stationary phase (∼108 cells per ml). In (C) and (D), spirochetes were grown in BSK-II medium. When bacterial growth reached mid-log phase (∼107 cells per ml), various amounts of IPTG were added into culture. Cells were collected at 9 h post-induction. In (A) and (C), concentrations of IPTG are indicated above the image. The arrow indicates OspC in (A). Specific antibodies, denoted as α- used in the immunoblot (B, C), are indicated on the left. In (D), data were collected from three independent experiments, and the bars represent the mean measurements ± standard deviation. The mean values between induced groups (100-, 200-, or 500 µM IPTG) and the uninduced group (0 µM IPTG) were compared using the Student’s t test and are significantly different (p<0.05). For data normalization, the B. burgdorferi flaB gene was used as an internal control.
Generation of an rrp2 Conditional Mutant in B. burgdorferi
Given that many attempts to fully inactivate rrp2 in B. burgdorferi have failed, and that rrp2 thus seems to be essential for B. burgdorferi in vitro growth [69], we generated a rrp2 conditional mutant in B. burgdorferi using a similar approach as described previously [69], [73]. In this conditional lethal mutant, the wild-type chromosomal copy of rrp2 is disrupted; rrp2 is expressed from an IPTG-inducible shuttle plasmid. As a prelude to this approach, we first created another IPTG-inducible rrp2 expression construct pRrp2-FLAG (Fig. 3A). From this shuttle plasmid, the expression of rrp2 in B. burgdorferi is tightly controlled by IPTG added into the medium. To assist in the detection of Rrp2, a DNA fragment encoding the FLAG tag was fused to the 3′ of rrp2. Because adding a FLAG tag to Rrp2 could affect its general function, this plasmid pRrp2-FLAG was initially introduced into the rrp2 point mutant OY01 (rrp2[G239C]) to create the strain OY173. OY173 was then used to test whether Rrp2-FLAG was functional in activating σ54-dependent rpoS expression. OY173 bacteria were grown continuously in media with varying concentrations of IPTG, and collected when growth reached the early stationary phase. As shown in Fig. 3C, when IPTG was not added into the medium, Rrp2-FLAG was not detected. However, when 20-, 50-, 100-, or 200-µM of IPTG was added into the media, the synthesis of Rrp2-FLAG was enhanced in a dose-dependent manner. The production of Rrp2-FLAG also resulted in the synthesis of RpoS and OspC in OY173. As shown in Fig. 3B and 3C, when bacteria were grown in BSK-II containing 50-, 100-, or 200-µM of IPTG, RpoS and OspC were readily detected in OY173, and the protein levels correlated closely with the increased levels of Rrp2-FLAG produced in B. burgdorferi. A 9-hr induction experiment was also carried out to examine gene induction in OY173. As shown in Fig. 3D, when bacteria at mid-log phase were exposed to 20-, 100-, 200-, or 500-µM of IPTG for 9 h, the synthesis of Rrp2-FLAG was readily detected in OY173. RpoS was also efficiently expressed in this strain (Fig. 3D). Moreover, qRT-PCR analyses revealed that exposing bacteria to IPTG for 9 h resulted in the concomitant induction of rrp2 and rpoS (Fig. 3E). Taken together, these data suggested that Rrp2-FLAG expressed from pRrp2-FLAG was capable of functioning like native Rrp2 to activate the expression of σ54-dependent rpoS.
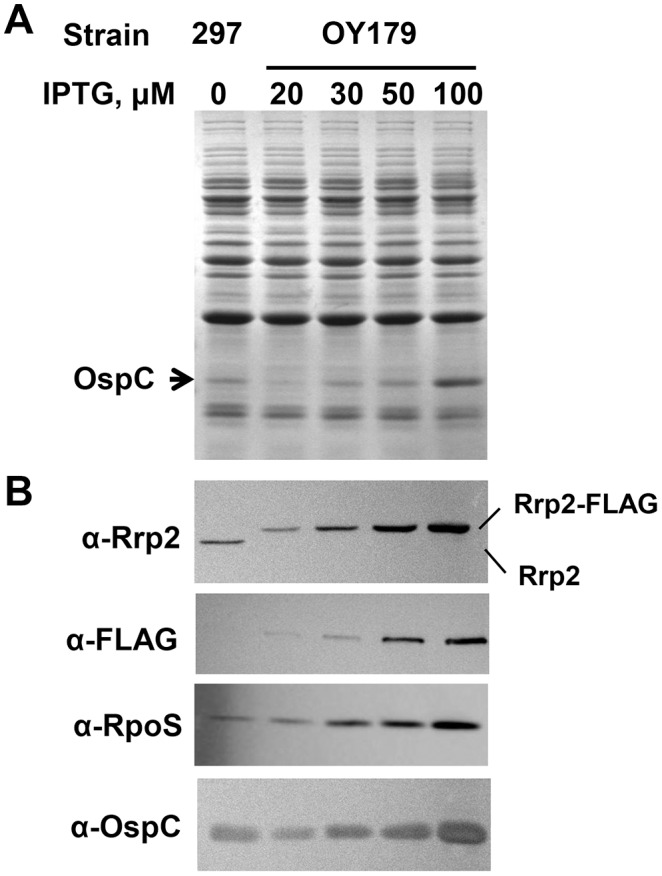
Figure 3. Gene expression in B. burgdorferi strain OY173.

(A) Construction of an IPTG-inducible rrp2-FLAG expression shuttle plasmid. The plasmid pRrp2-FLAG pRrp2 was introduced into strain OY01 (rrp2[G239C]), yielding OY173. SDS-PAGE (B), immunoblot (C, D), and qRT-PCR analyses (E) were performed to analyze gene expression. In (B) and (C), spirochetes grown in BSK-II medium containing varying concentrations of IPTG were harvested when bacterial growth reached early stationary phase (∼108 cells per ml). In (D) and (E), spirochetes were grown in BSK-II medium. When bacterial growth reached mid-log phase (∼107 cells per ml), various amounts of IPTG were added into culture. Cells were collected at 9 h post-induction. In (B) and (D), concentrations of IPTG are indicated above the image. The arrow indicates OspC in (B). Specific antibodies, denoted as α- used in the immunoblot (C, D), are indicated on the left. In (E), the bars represent the mean measurements ± standard deviation. The mean values between induced groups (100-, 200-, or 500 µM IPTG) and the uninduced group (0 µM IPTG) were compared using the Student’s t test and are significantly different (p<0.05). For data normalization, the B. burgdorferi flaB gene was used as an internal control.
Next, to fully inactivate rrp2, a suicide vector pOY202 that would target rrp2 was first introduced into strain 297, but no transformants could be recovered. We then introduced pOY202 into strain OY173 (containing pRrp2-FLAG), and the desired transformants were selected by using streptomycin and kanamycin, together with IPTG (IPTG was used to induce rrp2 expression from pRrp2-FLAG). Through allelic exchange, the chromosomal copy of rrp2 was replaced by the PflgB-kan cassette, yielding strain OY179 (Fig. 4A). The inactivation of chromosomal rrp2 was confirmed by PCR analyses. As shown in Fig. 4B, by using primers ZM235F and ZM235R, a 941-bp fragment spanning bb0764 and rrp2 was successfully amplified from WT strain 297 (lane 1), but not from the conditional mutant OY179 (lanes M1, M2, and M3). Also, PCR amplification revealed that strain OY179 contained the kan gene (conferring kanamycin resistance) and the aadA gene (conferring streptomycin resistance) (Fig. 4B).
Figure 4. Generation of an rrp2 conditional lethal mutant in B. burgdorferi.

(A) Schematic representation of the bb0764–bb0761 genes in the B. burgdorferi chromosome and the insertion of PflgB-kan cassette into rrp2 by homologous recombination. Arrows indicate the approximate positions of the oligonucleotide primers used for subsequent analyses. (B) Analyses of the wild-type 297 and the rrp2 conditional lethal mutant OY179 by PCR. The specific primer pairs are indicated on the right. Lanes WT, 297; lanes M1, M2, and M3, three clones of OY179.
The growth of the rrp2 conditional mutant OY179 was assessed by cultivating the bacteria in BSK-II medium containing various concentrations of IPTG. OY179 grew in BSK-II with 20-, 30-, 50-, or 100-µM IPTG, but failed to grow in BSK-II without IPTG (Fig. S1). Consistent with previous observations [69], we also conclude that rrp2 is essential for B. burgdorferi growth in vitro.
Expression of rrp2 Correlates with the Expression of RpoS and OspC in the rrp2 Conditional Mutant
To assess whether the protein level of Rrp2 correlated with the expression of rpoS, gene expression in OY179 was measured via SDS-PAGE and semi-quantitative immunoblot analyses. To this end, OY179 was grown in BSK-II with varying concentrations of IPTG and cultures were harvested when cell growth reached early stationary phase. For bacteria grown under these conditions, no obvious differences were observed when spirochete morphology and motility were examined using dark-field microscopy. As shown in Fig. 5B, immunoblot analyses revealed that OY179 produced Rrp2-FLAG but not native Rrp2. Moreover, the levels of Rrp2-FLAG were dependent on the concentrations of the inducer IPTG. Specifically, when 20-µM IPTG was added into the medium, the level of Rrp2-FLAG produced in OY179 was ∼3-fold lower than the level of Rrp2 observed in WT strain 297. When bacteria were cultivated in BSK-II containing 30-µM IPTG, Rrp2-FLAG was produced at a level commensurate with the level of Rrp2 in WT strain 297. When 50- or 100-µM IPTG was added into the medium, Rrp2-FLAG produced in OY179 was 3.5-fold or 10.3-fold higher, respectively, than the level of Rrp2 in strain 297. In addition, there was a close correlation between the levels of Rrp2-FLAG and the levels of RpoS and OspC (Fig. 5A and 5B). Taken together, the increased synthesis of RpoS was coincident with the IPTG-inducible production of Rrp2-FLAG, supporting that Rrp2 activates the RpoN-RpoS pathway in B. burgdorferi.
Figure 5. Gene expression in the rrp2 conditional lethal mutant OY179.
SDS-PAGE (A) and semi-quantitative immunoblot (B) analyses were performed to analyze gene expression. Bacteria were grown at 37°C in BSK-II medium with various concentrations of IPTG. When bacterial growth reached ∼108 cells per ml, spirochetes were harvested. Approximately 4×107 spirochetes were loaded onto each lane of a 12.5% SDS-PAGE gel. Concentrations of IPTG are indicated above the image, and the arrow in (A) indicates OspC. Specific antibodies, denoted as α- used in the immunoblot (B), are indicated on the left.
Rrp2 Indirectly Controls the Expression of OspC and DbpA via RpoS
Although the expression of ospC and dbpA was found to be lost in the rrp2 point mutant [10], [29], [45], how Rrp2 ultimately controls the expression of these key lipoproteins has remained somewhat unclear. Given that (1) the expression of ospC and dbpA are directly regulated by RpoS through RpoS-specific promoters [74]–[76], and (2) rpoS transcription is abolished in the rrp2 point mutant [10], [29], [45], we have hypothesized that Rrp2 likely regulates the expression of rpoS which, in turn, influences ospC and dbpA expression. To further test this hypothesis, we generated an IPTG-inducible rpoS expression shuttle construct pRpoS (i.e., pOY110) [30], in which rpoS expression is controlled solely by the IPTG-inducible PpQE30 promoter. This construct was introduced into the rrp2 point mutant OY01, to investigate whether the IPTG-induced RpoS could restore ospC and dbpA expression in this mutant. As shown in Fig. 6A and 6B, when RpoS was induced from pRpoS by IPTG, production of OspC and DbpA was consequently rescued. As aforementioned, when the rrp2 point mutation was complemented by the IPTG-inducible rrp2 expression construct pRrp2, expression of ospC was restored in the complemented strain OY160 (Fig. 1B and 1C). Consistent with previous findings [69], however, when pRrp2 was introduced into a B. burgdorferi rpoS mutant AH206 (ΔrpoS), RpoS and OspC were not produced in this strain, despite the fact that Rrp2 synthesis was induced by IPTG (Fig. S2). These combined data suggest that controlled induction of RpoS can overcome the Rrp2 deficiency, which constitutes compelling evidence that Rrp2 indirectly controls ospC and dbpA expression via RpoS.
Figure 6. Rrp2 controls expression of OspC and DbpA via RpoS.
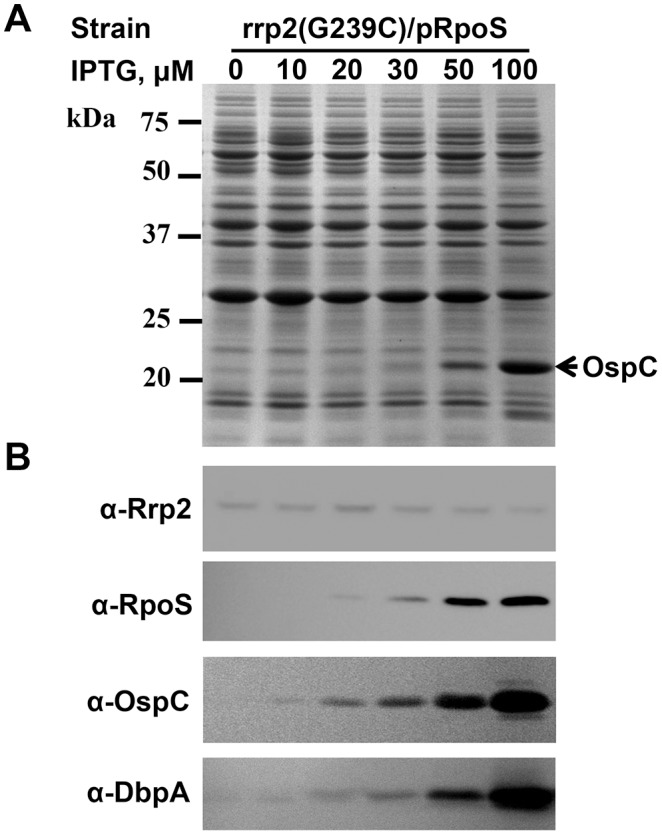
(A, B) the rrp2 point mutant rrp2(G239C) harboring the IPTG-inducible rpoS construct (pRpoS) was grown at 37°C with various concentrations of IPTG and gene expression was analyzed by SDS-PAGE (A) and immunoblot (B). The arrow in (A) indicates OspC. Specific antibodies, denoted as α-, used in the immunoblot (B) are indicated on the left.
Conclusions
This is the first study, to our knowledge, to show that the level of Rrp2 produced in B. burgdorferi directly correlates with RpoS levels. Moreover, our study provides further validation for the application of an IPTG-inducible expression system [65], [66] for assessing B. burgdorferi gene regulation. By using this system, an rrp2 conditional lethal mutant was generated. Given the inability to inactivate rrp2 via conventional deletion or insertion mutagenesis, and the fact that both σ54 and RpoS can be readily inactivated in B. burgdorferi [14], [18], [29], [48], [49], Rrp2 likely also controls the expression of genes independent of σ54 or RpoS control, which in turn suggests that Rrp2 may function as a unique bEBP (other characterized bEBPs activate only σ54–dependent promoters). Our new conditional mutant thus provides an innovative way not only to define the direct control of Rrp2 over the central RpoN-RpoS pathway, but also to interrogate the overall regulatory role of Rrp2 in B. burgdorferi gene regulation. For example, by profiling gene expression in this new conditional mutant, it may be possible to identify novel Rrp2-controlled, σ54- or RpoS-independent genes essential for B. burgdorferi growth or survival. Such studies will likely uncover new virulence-associated genes important for spirochetal pathogenesis, but also will provide the first definitive evidence that Rrp2 acts as an atypical bEBP to orchestrate virulence expression in B. burgdorferi. Although Rrp2 was presumed to be activated through phosphorylation [10], [29], [45], it still remains unanswered whether or how phosphorylated Rrp2 dynamically controls rpoS expression. Future work, such as pulse-chase analyses of the rrp2 point mutant OY01 trans-complemented with rrp2 variants, may help address these questions.
Supporting Information
Growth of the rrp2 conditional mutant OY179 in vitro . B. burgdorferi was inoculated into BSK-II medium with various concentrations of IPTG at 1000 spirochetes/ml. Spirochetes were enumerated using darkfield microscopy. Values are the means from three independent experiments. Error bars indicate standard deviations (n = 3).
(TIF)
Overexpression of Rrp2 does not restore expression of OspC and DbpA in the RpoS mutant. The rpoS point mutant AH206 harboring the IPTG-inducible rrp2 construct (pRrp2) was grown at 37°C with various concentrations of IPTG and gene expression was analyzed by SDS-PAGE (A) and immunoblot (B). RpoS and OspC were not detected in this strain, which is consistent with previous findings [69]. Specific antibodies, denoted as α-, used in the immunoblot (B) are indicated on the left.
(TIF)
Oligonucleotide primers used in this study.
(DOCX)
Funding Statement
This work was funded by grant NIAID-NIH AI-059062. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Burgdorfer W, Barbour AG, Hayes SF, Benach JL, Grunwaldt E, et al. (1982) Lyme disease-a tick-borne spirochetosis? Science 216: 1317–1319. [DOI] [PubMed] [Google Scholar]
- 2. Steere AC (1993) Current understanding of Lyme disease. Hosp Pract (Off Ed) 28: 37–44. [DOI] [PubMed] [Google Scholar]
- 3. Crother TR, Champion CI, Whitelegge JP, Aguilera R, Wu XY, et al. (2004) Temporal analysis of the antigenic composition of Borrelia burgdorferi during infection in rabbit skin. Infect Immun 72: 5063–5072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. de Silva AM, Telford SR 3rd, Brunet LR, Barthold SW, Fikrig E (1996) Borrelia burgdorferi OspA is an arthropod-specific transmission-blocking Lyme disease vaccine. J Exp Med 183: 271–275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Liang FT, Nelson FK, Fikrig E (2002) Molecular adaptation of Borrelia burgdorferi in the murine host. J Exp Med 196: 275–280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Montgomery RR, Malawista SE, Feen KJ, Bockenstedt LK (1996) Direct demonstration of antigenic substitution of Borrelia burgdorferi ex vivo: exploration of the paradox of the early immune response to outer surface proteins A and C in Lyme disease. J Exp Med 183: 261–269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Ohnishi J, Piesman J, de Silva AM (2001) Antigenic and genetic heterogeneity of Borrelia burgdorferi populations transmitted by ticks. Proc Natl Acad Sci U S A 98: 670–675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Schwan TG, Piesman J (2000) Temporal changes in outer surface proteins A and C of the lyme disease-associated spirochete, Borrelia burgdorferi, during the chain of infection in ticks and mice. J Clin Microbiol 38: 382–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Schwan TG, Piesman J, Golde WT, Dolan MC, Rosa PA (1995) Induction of an outer surface protein on Borrelia burgdorferi during tick feeding. Proc Natl Acad Sci U S A 92: 2909–2913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Boardman BK, He M, Ouyang Z, Xu H, Pang X, et al. (2008) Essential role of the response regulator Rrp2 in the infectious cycle of Borrelia burgdorferi . Infect Immun 76: 3844–3853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Boylan JA, Posey JE, Gherardini FC (2003) Borrelia oxidative stress response regulator, BosR: a distinctive Zn-dependent transcriptional activator. Proc Natl Acad Sci U S A 100: 11684–11689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Burtnick MN, Downey JS, Brett PJ, Boylan JA, Frye JG, et al. (2007) Insights into the complex regulation of rpoS in Borrelia burgdorferi . Mol Microbiol 65: 277–293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Dunham-Ems SM, Caimano MJ, Eggers CH, Radolf JD (2012) Borrelia burgdorferi requires the alternative sigma factor RpoS for dissemination within the vector during tick-to-mammal transmission. PLoS Pathog 8: e1002532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Fisher MA, Grimm D, Henion AK, Elias AF, Stewart PE, et al. (2005) Borrelia burgdorferi σ54 is required for mammalian infection and vector transmission but not for tick colonization. Proc Natl Acad Sci U S A 102: 5162–5167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Fraser CM, Casjens S, Huang WM, Sutton GG, Clayton R, et al. (1997) Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi . Nature 390: 580–586. [DOI] [PubMed] [Google Scholar]
- 16. Freedman JC, Rogers EA, Kostick JL, Zhang H, Iyer R, et al. (2010) Identification and molecular characterization of a cyclic-di-GMP effector protein, PlzA (BB0733): additional evidence for the existence of a functional cyclic-di-GMP regulatory network in the Lyme disease spirochete, Borrelia burgdorferi . FEMS Immunol Med Microbiol 58: 285–294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. He M, Ouyang Z, Troxell B, Xu H, Moh A, et al. (2011) Cyclic di-GMP is essential for the survival of the Lyme disease spirochete in ticks. PLoS Pathog 7: e1002133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hubner A, Yang X, Nolen DM, Popova TG, Cabello FC, et al. (2001) Expression of Borrelia burgdorferi OspC and DbpA is controlled by a RpoN-RpoS regulatory pathway. Proc Natl Acad Sci U S A 98: 12724–12729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Hyde JA, Shaw DK, Smith Iii R, Trzeciakowski JP, Skare JT (2009) The BosR regulatory protein of Borrelia burgdorferi interfaces with the RpoS regulatory pathway and modulates both the oxidative stress response and pathogenic properties of the Lyme disease spirochete. Mol Microbiol 74: 1344–1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Jutras BL, Chenail AM, Carroll DW, Miller MC, Zhu H, et al. (2013) Bpur, the Lyme disease spirochete’s PUR domain protein: identification as a transcriptional modulator and characterization of nucleic acid interactions. J Biol Chem 288: 26220–26234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Jutras BL, Verma A, Adams CA, Brissette CA, Burns LH, et al. (2012) BpaB and EbfC DNA-binding proteins regulate production of the Lyme disease spirochete’s infection-associated Erp surface proteins. J Bacteriol 194: 778–786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Karna SL, Sanjuan E, Esteve-Gassent MD, Miller CL, Maruskova M, et al. (2011) CsrA modulates levels of lipoproteins and key regulators of gene expression critical for pathogenic mechanisms of Borrelia burgdorferi . Infect Immun 79: 732–744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Katona LI, Tokarz R, Kuhlow CJ, Benach J, Benach JL (2004) The fur homologue in Borrelia burgdorferi . J Bacteriol 186: 6443–6456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Kostick JL, Szkotnicki LT, Rogers EA, Bocci P, Raffaelli N, et al. (2011) The diguanylate cyclase, Rrp1, regulates critical steps in the enzootic cycle of the Lyme disease spirochetes. Mol Microbiol 81: 219–231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Lybecker MC, Abel CA, Feig AL, Samuels DS (2010) Identification and function of the RNA chaperone Hfq in the Lyme disease spirochete Borrelia burgdorferi . Mol Microbiol 78: 622–635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Lybecker MC, Samuels DS (2007) Temperature-induced regulation of RpoS by a small RNA in Borrelia burgdorferi . Mol Microbiol 64: 1075–1089. [DOI] [PubMed] [Google Scholar]
- 27. Medrano MS, Policastro PF, Schwan TG, Coburn J (2010) Interaction of Borrelia burgdorferi Hbb with the p66 promoter. Nucleic Acids Res 38: 414–427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Miller CL, Karna SL, Seshu J (2013) Borrelia host adaptation Regulator (BadR) regulates rpoS to modulate host adaptation and virulence factors in Borrelia burgdorferi . Mol Microbiol 88: 105–124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Ouyang Z, Blevins JS, Norgard MV (2008) Transcriptional interplay among the regulators Rrp2, RpoN and RpoS in Borrelia burgdorferi . Microbiology 154: 2641–2658. [DOI] [PubMed] [Google Scholar]
- 30. Ouyang Z, Deka RK, Norgard MV (2011) BosR (BB0647) controls the RpoN-RpoS regulatory pathway and virulence expression in Borrelia burgdorferi by a novel DNA-binding mechanism. PLoS Pathog 7: e1001272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Ouyang Z, Kumar M, Kariu T, Haq S, Goldberg M, et al. (2009) BosR (BB0647) governs virulence expression in Borrelia burgdorferi . Mol Microbiol 74: 1331–1343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Radolf JD, Caimano MJ, Stevenson B, Hu LT (2012) Of ticks, mice and men: understanding the dual-host lifestyle of Lyme disease spirochaetes. Nat Rev Microbiol 10: 87–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Rogers EA, Terekhova D, Zhang HM, Hovis KM, Schwartz I, et al. (2009) Rrp1, a cyclic-di-GMP-producing response regulator, is an important regulator of Borrelia burgdorferi core cellular functions. Mol Microbiol 71: 1551–1573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Salman-Dilgimen A, Hardy PO, Dresser AR, Chaconas G (2011) HrpA, a DEAH-box RNA helicase, is involved in global gene regulation in the Lyme disease spirochete. PLoS One 6: e22168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Samuels DS, Radolf JD (2009) Who is the BosR around here anyway? Mol Microbiol 74: 1295–1299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Samuels DS (2011) Gene regulation in Borrelia burgdorferi . Annu Rev Microbiol 65: 479–499. [DOI] [PubMed] [Google Scholar]
- 37.Samuels DS, Radolf JD (2010) Borrelia: molecular biology, host interaction and pathogenesis. Wymondham: Caister Academic. xii, 547 p. [Google Scholar]
- 38. Sanjuan E, Esteve-Gassent MD, Maruskova M, Seshu J (2009) Overexpression of CsrA (BB0184) alters the morphology and antigen profiles of Borrelia burgdorferi . Infect Immun 77: 5149–5162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Scheckelhoff MR, Telford SR, Wesley M, Hu LT (2007) Borrelia burgdorferi intercepts host hormonal signals to regulate expression of outer surface protein A. Proc Natl Acad Sci U S A. 104: 7247–7252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Smith AH, Blevins JS, Bachlani GN, Yang XF, Norgard MV (2007) Evidence that RpoS (σS) in Borrelia burgdorferi is controlled directly by RpoN (σ54/σN). J Bacteriol 189: 2139–2144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Sultan SZ, Pitzer JE, Miller MR, Motaleb MA (2010) Analysis of a Borrelia burgdorferi phosphodiesterase demonstrates a role for cyclic-di-guanosine monophosphate in motility and virulence. Mol Microbiol 77: 128–142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Sze CW, Li C (2011) Inactivation of bb0184, which encodes carbon storage regulator A, represses the infectivity of Borrelia burgdorferi . Infect Immun 79: 1270–1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Sze CW, Smith A, Choi YH, Yang X, Pal U, et al. (2013) Study of the response regulator Rrp1 reveals its regulatory role in chitobiose utilization and virulence of Borrelia burgdorferi . Infect Immun 81: 1775–1787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Xu Q, Shi Y, Dadhwal P, Liang FT (2012) RpoS regulates essential virulence factors remaining to be identified in Borrelia burgdorferi . PLoS One 7: e53212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Yang XF, Alani SM, Norgard MV (2003) The response regulator Rrp2 is essential for the expression of major membrane lipoproteins in Borrelia burgdorferi . Proc Natl Acad Sci U S A 100: 11001–11006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Blevins JS, Hagman KE, Norgard MV (2008) Assessment of decorin-binding protein A to the infectivity of Borrelia burgdorferi in the murine models of needle and tick infection. BMC Microbiol 8: 82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Caimano MJ, Eggers CH, Gonzalez CA, Radolf JD (2005) Alternate sigma factor RpoS is required for the in vivo-specific repression of Borrelia burgdorferi plasmid lp54-borne ospA and lp6.6 genes. J Bacteriol 187: 7845–7852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Caimano MJ, Eggers CH, Hazlett KR, Radolf JD (2004) RpoS is not central to the general stress response in Borrelia burgdorferi but does control expression of one or more essential virulence determinants. Infect Immun 72: 6433–6445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Caimano MJ, Iyer R, Eggers CH, Gonzalez C, Morton EA, et al. (2007) Analysis of the RpoS regulon in Borrelia burgdorferi in response to mammalian host signals provides insight into RpoS function during the enzootic cycle. Mol Microbiol 65: 1193–1217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Grimm D, Tilly K, Byram R, Stewart PE, Krum JG, et al. (2004) Outer-surface protein C of the Lyme disease spirochete: a protein induced in ticks for infection of mammals. Proc Natl Acad Sci U S A 101: 3142–3147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Ouyang Z, Narasimhan S, Neelakanta G, Kumar M, Pal U, et al. (2012) Activation of the RpoN-RpoS regulatory pathway during the enzootic life cycle of Borrelia burgdorferi . BMC Microbiol 12: 44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Pal U, de Silva AM, Montgomery RR, Fish D, Anguita J, et al. (2000) Attachment of Borrelia burgdorferi within Ixodes scapularis mediated by outer surface protein A. J Clin Invest. 106: 561–569. [Google Scholar]
- 53. Pal U, Yang X, Chen M, Bockenstedt LK, Anderson JF, et al. (2004) OspC facilitates Borrelia burgdorferi invasion of Ixodes scapularis salivary glands. J Clin Invest 113: 220–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Shi Y, Xu Q, McShan K, Liang FT (2008) Both decorin-binding proteins A and B are critical for overall virulence of Borrelia burgdorferi . Infect Immun 76: 1239–1246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Tilly K, Krum JG, Bestor A, Jewett MW, Grimm D, et al. (2006) Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection. Infect Immun 74: 3554–3564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Weening EH, Parveen N, Trzeciakowski JP, Leong JM, Hook M, et al. (2008) Borrelia burgdorferi lacking DbpBA exhibits an early survival defect during experimental infection. Infect Immun 76: 5694–5705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Yang XF, Pal U, Alani SM, Fikrig E, Norgard MV (2004) Essential role for OspA/B in the life cycle of the Lyme disease spirochete. J Exp Med 199: 641–648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Bush M, Dixon R (2012) The role of bacterial enhancer binding proteins as specialized activators of σ54-dependent transcription. Microbiol Mol Biol Rev 76: 497–529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Morett E, Segovia L (1993) The σ54 bacterial enhancer-binding protein family: mechanism of action and phylogenetic relationship of their functional domains. J Bacteriol 175: 6067–6074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Rappas M, Bose D, Zhang X (2007) Bacterial enhancer-binding proteins: unlocking σ54-dependent gene transcription. Curr Opin Struct Biol 17: 110–116. [DOI] [PubMed] [Google Scholar]
- 61. Wigneshweraraj S, Bose D, Burrows PC, Joly N, Schumacher J, et al. (2008) Modus operandi of the bacterial RNA polymerase containing the σ54 promoter-specificity factor. Mol Microbiol 68: 538–546. [DOI] [PubMed] [Google Scholar]
- 62. Ghosh T, Bose D, Zhang X (2010) Mechanisms for activating bacterial RNA polymerase. FEMS Microbiol Rev 34: 611–627. [DOI] [PubMed] [Google Scholar]
- 63. Studholme DJ, Dixon R (2003) Domain architectures of σ54-dependent transcriptional activators. J Bacteriol 185: 1757–1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. He M, Oman T, Xu H, Blevins J, Norgard MV, et al. (2008) Abrogation of ospAB constitutively activates the Rrp2-RpoN-RpoS pathway (σN-σS cascade) in Borrelia burgdorferi . Mol Microbiol 70: 1453–1464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Blevins JS, Revel AT, Smith AH, Bachlani GN, Norgard MV (2007) Adaptation of a luciferase gene reporter and lac expression system to Borrelia burgdorferi . Appl Environ Microbiol 73: 1501–1513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Gilbert MA, Morton EA, Bundle SF, Samuels DS (2007) Artificial regulation of ospC expression in Borrelia burgdorferi . Mol Microbiol 63: 1259–1273. [DOI] [PubMed] [Google Scholar]
- 67. Hughes CA, Kodner CB, Johnson RC (1992) DNA analysis of Borrelia burgdorferi NCH-1, the first northcentral U.S. human Lyme disease isolate. J Clin Microbiol 30: 698–703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Pollack RJ, Telford SR 3rd, Spielman A (1993) Standardization of medium for culturing Lyme disease spirochetes. J Clin Microbiol 31: 1251–1255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Groshong AM, Gibbons NE, Yang XF, Blevins JS (2012) Rrp2, a prokaryotic enhancer-like binding protein, is essential for viability of Borrelia burgdorferi . J Bacteriol 194: 3336–3342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Samuels DS (1995) Electrotransformation of the spirochete Borrelia burgdorferi . Methods Mol Biol 47: 253–259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Ouyang Z, He M, Oman T, Yang XF, Norgard MV (2009) A manganese transporter, BB0219 (BmtA), is required for virulence by the Lyme disease spirochete, Borrelia burgdorferi . Proc Natl Acad Sci U S A 106: 3449–3454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Prelich G (2012) Gene overexpression: uses, mechanisms, and interpretation. Genetics 190: 841–854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Lenhart TR, Akins DR (2010) Borrelia burgdorferi locus BB0795 encodes a BamA orthologue required for growth and efficient localization of outer membrane proteins. Mol Microbiol 75: 692–709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Alverson J, Bundle SF, Sohaskey CD, Lybecker MC, Samuels DS (2003) Transcriptional regulation of the ospAB and ospC promoters from Borrelia burgdorferi . Mol Microbiol 48: 1665–1677. [DOI] [PubMed] [Google Scholar]
- 75. Ouyang Z, Haq S, Norgard MV (2010) Analysis of the dbpBA upstream regulatory region controlled by RpoS in Borrelia burgdorferi . J Bacteriol 192: 1965–1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Yang XF, Lybecker MC, Pal U, Alani SM, Blevins J, et al. (2005) Analysis of the ospC regulatory element controlled by the RpoN-RpoS regulatory pathway in Borrelia burgdorferi . J Bacteriol 187: 4822–4829. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Growth of the rrp2 conditional mutant OY179 in vitro . B. burgdorferi was inoculated into BSK-II medium with various concentrations of IPTG at 1000 spirochetes/ml. Spirochetes were enumerated using darkfield microscopy. Values are the means from three independent experiments. Error bars indicate standard deviations (n = 3).
(TIF)
Overexpression of Rrp2 does not restore expression of OspC and DbpA in the RpoS mutant. The rpoS point mutant AH206 harboring the IPTG-inducible rrp2 construct (pRrp2) was grown at 37°C with various concentrations of IPTG and gene expression was analyzed by SDS-PAGE (A) and immunoblot (B). RpoS and OspC were not detected in this strain, which is consistent with previous findings [69]. Specific antibodies, denoted as α-, used in the immunoblot (B) are indicated on the left.
(TIF)
Oligonucleotide primers used in this study.
(DOCX)