

Abstract
Background:
Os odontoideum is a well identified anomaly of the craniovertebral junction. Since its initial description, there has been a continuous debate regarding the nature of its etiology: Whether congenital or traumatic. We sought to compare the gene expression profiles in patients with congenital os odontoideum, those with traumatic os odontoideum and controls.
Methods:
We have evaluated a pair of identical twins both with os odontoideum. We identified two additional patients with and four subjects without os odontoideum. We analyzed the gene expression profiles in these patients using a custom TaqMan microarray and quantitative reverse transcriptase polymerase chain reaction (qRT-PCR). The relative gene expression profiles in the two identical twins, the two nontwin patients with os odontoideum and the controls were assessed.
Results:
A total of 213 genes with significantly different expression between the twin os odontoideum patients and the subjects without os odontoideum were detected. CACNG6, PHEX, CACNAD3, IL2, FAS, TUFT1, KIT, TGFBR2, and IGF2 were expressed at levels greater than 100-fold more in the twins. There were six genes with significantly different expression profiles in the twins as compared with the nontwin os odontoideum patients: CMK4, ATF1, PLCG1, TAB1, E2F3, and ATF4. There were no statistically significant differences in gene expression in the four patients with os odontoideum and the subjects without. Trends, however, were noted in MMP8, KIT, HIF1A, CREB3, PWHAZ, TGFBR1, NFKB2, FGFR1, IPO8, STAT1, COL1A1, and BMP3.
Conclusions:
Os odontoideum has multiple etiologies, both traumatic and congenital and perhaps some represent a combination of the two. This work has identified a number of genes that show increased expression in a pair of twins with congenital os odontoideum and also demonstrates trends in gene expression profiles between a larger group of os odontoideum patients and non-os patients. A number of these genes are related to bone morphogenesis and maintenance.
Keywords: Anomaly, cervical spine, craniovertebral junction, identical twins, os odontoideum
INTRODUCTION
Os odontoideum is a well-defined anatomic anomaly consisting of a smooth ossicle of bone separated from a shortened odontoid process.[1] The incidence and prevalence of os odontoideum is unknown. This entity was first described in 1886 by Giacomini, which has been followed by numerous publications debating its etiology. Two theories—congenital and traumatic—dominate the discussion. A recent review by Arvin et al.[3] provides an excellent discussion of the regional embryology and anatomy as related to the pathogenesis of os odontoideum. The congenital hypothesis is founded on the principle that the os odontoideum is a segmental defect, which represents a failed fusion of the odontoid and C2 vertebral body.[8] Conversely, the traumatic hypothesis considers the os odontoideum an acquired pathology resulting from avascular necrosis following an odontoid fracture.[11] The congenital hypothesis is supported by reports of identical twins both with os odontoideum[18] and two separate communications of families with an autosomal dominant inheritance pattern of os odontoideum.[26,43] Support of the traumatic hypothesis comes from the observation of a pair of identical twins one with os and one without[40] and from 13 patients reported in the literature who had radiographic documentation of a normal odontoid and subsequently acquired os odontoideum after a trauma.[11,12,13,31,35,37,42,48] In this manuscript we describe the genetic features of two identical male twins both with os odontoideum, their family members, other patients with traumatic os odontoideum, and control patients without an os odontoideum.
MATERIALS AND METHODS
Patients and study design
IRB approval was obtained from Rush University Medical Center and informed consent was obtained from participants.
Two 20-year-old identical male twins were evaluated in the neurosurgery clinic at Rush University Medical Center. Neither twin had a significant history of trauma and both were active collegiate water-polo players. The clinical presentation was related to neck pain in one of the twins. Imaging revealed an os odontoideum. Flexion-extension imaging revealed an orthotopic os odontoideum with gross instability. Given this finding his brother underwent similar evaluation with comparable results. Preoperative cervical spine films are shown in Figures 1 and 2.
Figure 1.
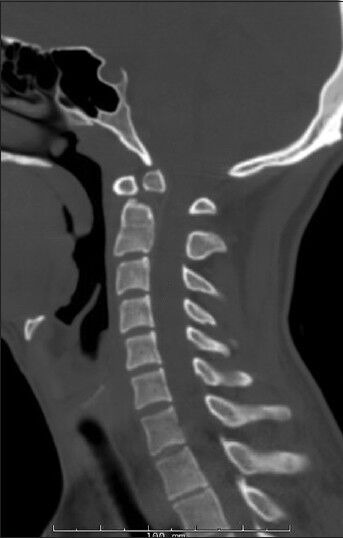
Preoperative sagittal cervical spine CT scan of Twin-1
Figure 2.
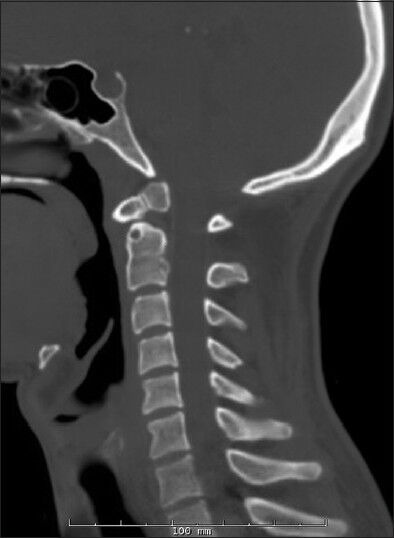
Preoperative sagittal cervical spine CT scan of Twin-2
Two additional, unrelated patients with an os odontoideum were subsequently treated. The first was a 49-year-old female who had a prior history of occipitocervical fusion at an outside hospital. She initially presented with hardware failure, pseudoarthrosis, and a wound infection. She did have a remote history of significant head trauma. Imaging revealed an os odontoideum, a C2-3 Klippel–Feil anomaly and a kyphotic deformity [Figure 3]. She ultimately underwent revision of her occipitocervical fusion. The second nontwin os patient was a 53-year-old female with neck pain and myelopathy. Imaging studies revealed an unstable os odontoideum with cord compression at C1 [Figure 4]. She was treated with a C1-laminectomy and an instrumented atlanto-axial fusion.
Figure 3.
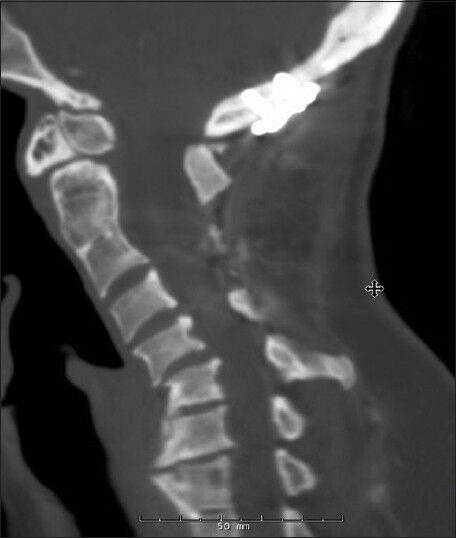
Preoperative sagittal cervical spine CT scan of non-twin Os-1
Figure 4.
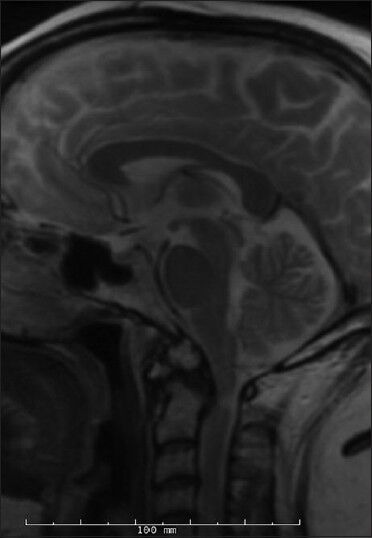
Preoperative sagittal T2-weighted cervical spine MRI of non-twin Os-2
Four subjects with prior cervical spine radiography that clearly demonstrated normal craniovertebral junction (CVJ) anatomy—absence of os odontoideum or other congenital anomaly—were identified to serve as controls. One of these subjects was the biological mother of the identical twins with os odontoideum. This control group included a 52-year-old female, an 87-year-old female, a 49-year-old male, and a 59-year-old female.
Comparisons were utilized in a case-control model and included relative gene expression levels between the identical twins with os odontoideum and the controls without os odontoideum; between the identical twins and the unrelated patients with os odontoideum; and finally, between all patients with os odontoideum and all controls without os odontoideum.
Sample collection and RNA preparation
Blood samples were collected via venipuncture from all patients into 5 ml collection tubes containing EDTA and immediately placed on ice. Total RNA from the blood cell pellet of each sample and isolated mRNA for gene expression profiling using the miRVana RNA isolation kit (Ambion, Grand Island, NY).
mRNA qRT-PCR array assays and data analysis
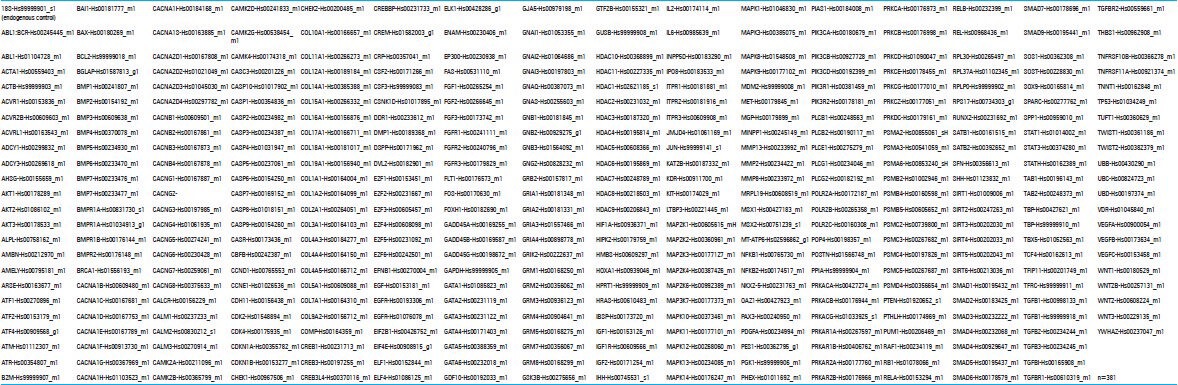
A total of 1 μg of RNA of each sample was used for gene expression profiling. Gene expression profiling was performed on the HT7900 real-time PCR system (Applied Biosystems, Foster City, CA) using custom TaqMan array cards (Applied Biosystems, Supplement 1) on all RNA samples. Reverse transcription was performed using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems) according to manufacturer's protocol. Amplification using the custom Taqman array cards was performed using the Taqman Gene Expression Master mix (Applied Biosystem) according to manufacturer's protocol on a 7900HTReal-time PCR system at the miRNA and Gene Expression Core, Rush University Medical Center. All card amplification reactions were successful. Amplification data were acquired using the SDS2.3/RQ Manager (Applied Biosystems) and DataAssist 3.01 software (Applied Biosystems). The relative expression level for each mRNA was represented as cycle threshold (Ct). mRNAs with Ct < 40 were accepted as “detected". Eukaryotic 18s rRNA (18S-Hs99999901_s1) was identified as the most stably expressed endogenous control was used as the normalization control. Normalized expression level was calculated as Delta CT (DCt) or ∆Ct = Ct (test RNA) -Ct (18s RNA). Differential expression between patients was calculated as DDCt or ∆(∆Ct) = Ave DCt (test) - Ave DCt (control). Fold change or relative quantification (RQ) was calculated as RQ = 2-∆(∆Ct). The Benjamini-Hochberg method[5] was applied to minimize the false discovery rate. An adjusted P < 0.05 was set as the criterion for significance of differential expression. FiRe Macro[15] was used in Microsoft Excel (Microsoft Corporation, Redmond, WA) to generate heatmaps of the normalized RNA expression levels.
Supplementary Table 1.
Custom TaqMan Array design
Functional analysis
To better understand the functions of the differentially expressed mRNAs, we performed a NCBI Gene search[28] using the test RNA as a key word to compile known functions of this mRNA. We report the NCBI Gene Summary of selected genes in Tables 1, 3, and 4.
Table 2.
Complete results of os vs. non-os relative gene expression

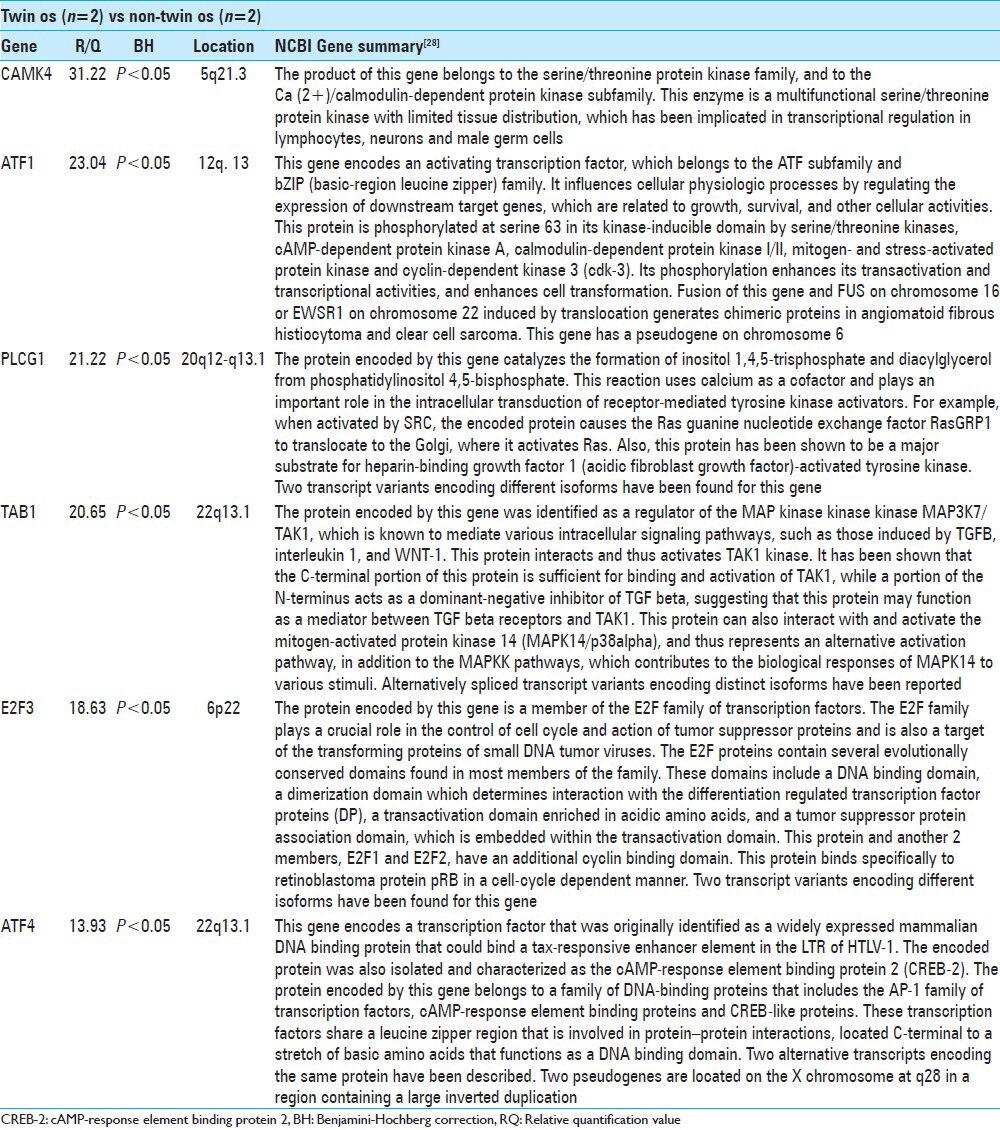
Table 3.
Twins vs non-twin os
RESULTS
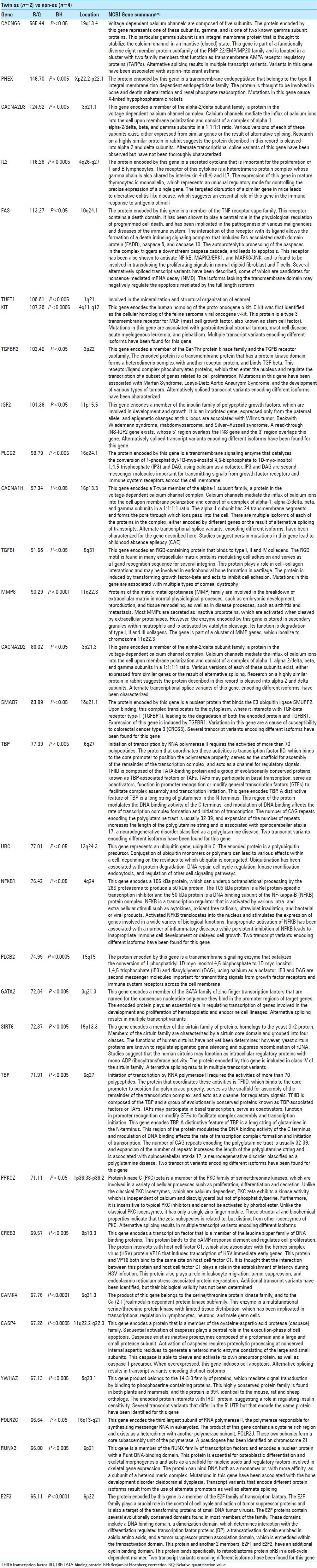
A comparison between the identical twins with os odontoideum (Twins) to the four control subjects (non-os) revealed 213 statistically significant differences in gene expression. The top 30 genes and a brief summary[23,28] of their function are provided in Table 1. Nine genes were expressed in the Twins at levels greater-than 100 times that of the non-os group. CACNG6 showed a 565-fold increase in expression in the Twins (P < 0.05). This mRNA is derived from a gene located on chromosome 19p13.4 and encodes the gamma-6 subunit of the voltage dependant calcium channel. PHEX expression was 447-fold higher in the Twins (P < 0.005). The PHEX mRNA encodes a transmembrane zinc-dependent endopeptidase involved in bone and dentin mineralization and renal phosphate reabsorption. CACNA2D3 showed a 125-fold increase in the Twins (P < 0.005). This gene is located on chromosome 3p21.2 and encodes the alpha-2/delta-3 subunit in the voltage-dependent calcium channel. Interleukin 2 (IL2) was increased 116-fold in the Twins (P < 0.0005); this gene is located on 4q26-q27 and encodes an inflammatory cytokine that is important for T- and B-cell proliferation. FAS was increased 113-fold in the Twins (P < 0.05); this gene is located on chromosome 10q24.1 and encodes a transmembrane protein important in mediating the extrinsic pathway of apoptosis. It is also involved in transducing proliferative signals from normal fibroblasts and T-cells. TUFT1 was increased 109-fold (P < 0.005) in the Twins; this gene is located on chromosome 1q21 and is involved in the mineralization and structural organization of enamel. KIT was increased 107-fold in the Twins (P < 0.0005); this gene is located on 4q11-q12 and encodes a transmembrane receptor for mast cell growth factor. It is a proto-oncogene and mutations are associated with malignancies (e.g. gastrointestinal stromal tumor and acute myelogenous leukemia). TGFBR2 was increased 102-fold in the twins (P < 0.05). This gene is located on chromosome 3p22 and encodes a transmembrane Ser/Thr protein kinase related to cell proliferation. Mutations have been associated with Marfan syndrome, Loeys–Dietz Aortic Aneurysm Syndrome and various malignancies. IGF2 was increased 101-fold in the Twins (P < 0.05); this gene is located on 11p15.5 and encodes a member of the insulin-like growth factors that is a major fetal growth factor. Complete results of all 213 genes with significant differences in expression between the Twins and non-os groups are shown in Table 2.
Table 1.
Top 30 gene expression differences between Twins and non-os
Comparing the Twins with the two unrelated patients with os odontoideum (non-Twin os) revealed six genes with statistically significant differences in expression. These genes and a brief summary of their functions are presented in Table 3. CAMK4 was increased 31-fold in the Twins as compared with the non-Twin os patients (P < 0.05). This gene is located on chromosome 5q21.3 and encodes a calcium-calmodulin dependent Ser/Thr kinase. ATF1 was increased 23-fold in the Twins (P < 0.05); this gene is located on chromosome 12q13 and encodes an activating transcription factor and fusion products have been implicated in angiomatoid fibrous histiocytoma and clear cell sarcoma. Expression of PLCG1 was increased 21-fold in the Twins (P < 0.05). This gene encodes gamma-1 phospholipase-C and is located on chromosome 20q12-q13.1. TAB1 was increased 21-fold (P < 0.05); this gene is located on chromosome 22q13.1 and encodes a protein involved in the regulation of the MAP kinase pathway. E2F3 was increased 19-fold (P < 0.05). This gene is located on chromosome 6p22 and encodes a protein regulator of the cell cycle. ATF4 was found to be elevated 14-fold in the Twins (P < 0.05); this gene is located on chromosome 22q13.1 and encodes a transcription factor that modulates DNA transcription in response to cAMP.
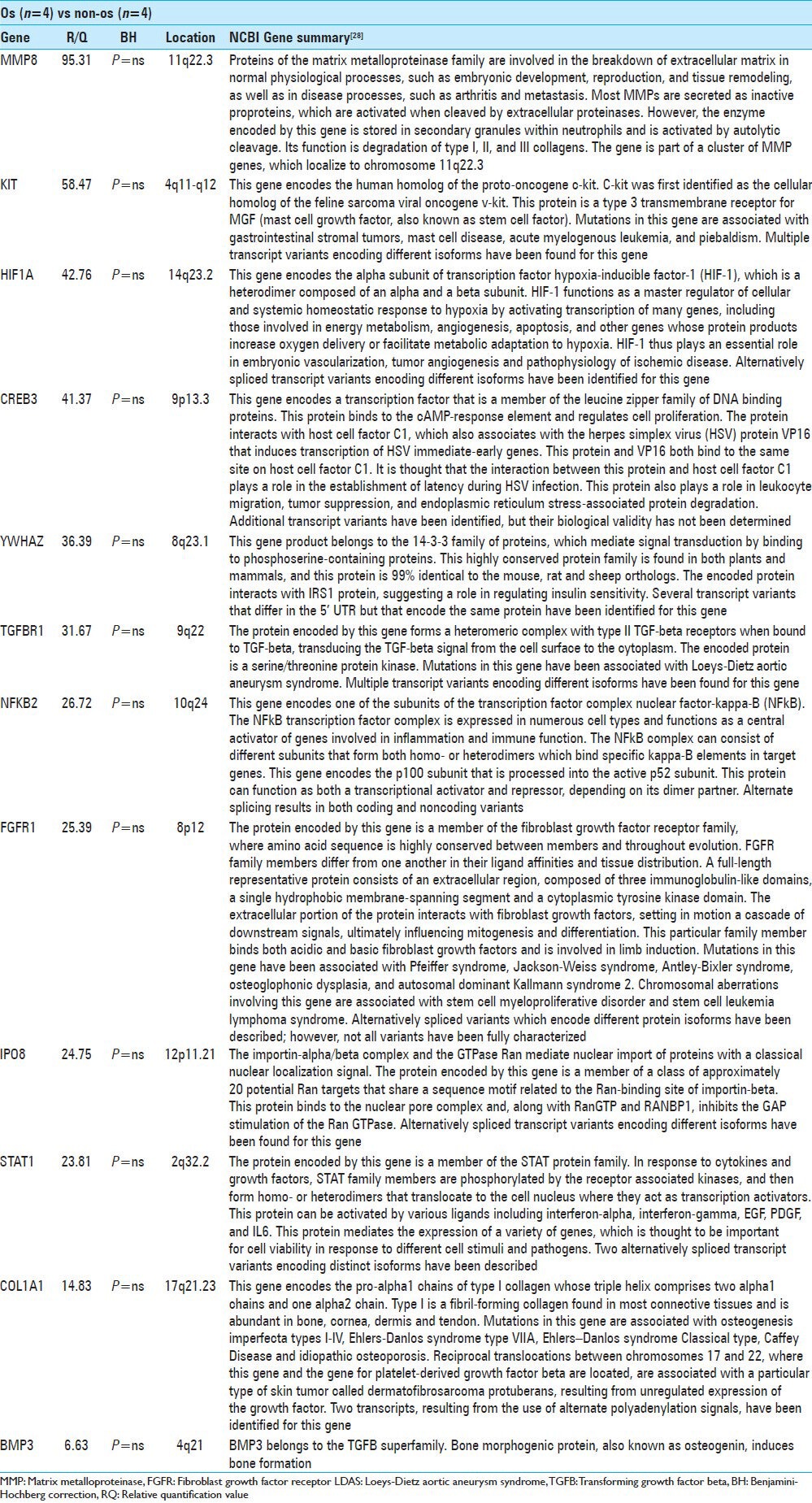
Comparing both the Twins and non-Twin os (Os) to the non-os group revealed no statistically significant differences in gene expression. We report the 10 genes with the greatest fold-change and an unadjusted P < 0.1 [Table 4]. MMP8 was increased 95-fold in Os patients (P = ns); this gene is located on chromosome 11q22.3 and encodes a matrix metalloproteinase that is stored in neutrophil granules and functions in the degradation of collagen types I, II and III. KIT was elevated 58-fold in the Os patients (P = ns). HIF1A was elevated 43-fold in the Os group. This gene is located on chromosome 14q23.2 and encodes the alpha subunit of hypoxia-inducible factor-1, a transcription factor, which functions in cellular homeostasis in response to hypoxia. CREB3 was elevated 41-fold in the Os group; this gene is located on chromosome 9p13.3 and encodes a transcription factor that regulates cell proliferation. CREB3 is involved in establishing latent herpes simplex virus infections and also plays a role in leukocyte migration, tumor suppression, and endoplasmic reticulum stress-associated protein degradation. YWHAZ was elevated 36-fold in the os patients (P = ns); this gene is located on chromosome 8q23.1 and belongs to the 14-3-3 family of proteins. It is involved in signal transduction and has been shown to interact with the insulin receptor-substrate-1 protein. TGFBR1 was elevated 32-fold in os patients (P = ns); it is located on chromosome 9q22 and encodes a Ser/Thr protein kinase involved in signal transduction. NFKB2 was elevated 27-fold in the os group. The gene is located on chromosome 10q24 and encodes the beta-subunit of the NF-kB transcription factor complex, which serves to activate inflammatory responses. FGFR1 was elevated 25-fold (P = ns) in the os patients. This gene is located on 8p12 and encodes a tyrosine-kinase protein that binds both basic and acidic fibroblast growth factor and mediates cell differentiation and mitogenesis. Mutations in this gene are associated with Pfeiffer syndrome, Jackson–Weiss syndrome, Antley–Bixler syndrome, osteoglophonic dysphagia, and autosomal dominant Kallman syndrome 2. IPO8 was increased 25-fold; this gene is located on chromosome 12p11.21 and encodes a protein that modulates Ran GTPase activity. STAT1 was increased 24-fold; this gene is located on chromosome 2q32.2 and encodes a protein that serves as a transcription activator in response to cytokine and growth factor signaling. In addition to the 10 genes reported above, COL1A1 was found to be elevated 14.8-fold (P = ns) and BMP3 was elevated 6.6-fold (P = ns) in the Os group. The COL1A1 gene is located on chromosome 17q21.23 and encodes the pro-alpha1 chain of type-1 collagen, which is found in most connective tissues and is abundant in bone. Mutations in COL1A1 are associated with osteogenesis imperfect types I-IV, Ehlers–Danlos syndrome type VIIA, Ehlers–Danlos syndrome Classical type, Caffey disease and idiopathic osteoporosis. BMP3 (osteogenin) is a gene that is located on chromosome 4q21 and encodes a member of the TGF-B superfamily that induces bone formation.
Table 4.
Top 10 RQ values in os vs non-os with uncorrected P<0.1
DISCUSSION
The identification of a pair of identical twins without a history of trauma both harboring os odontoidea of nearly identical morphology [Figures 1 and 2] lends further support to the concept of a congenital etiology for this disorder at least for some individuals. While the traumatic hypothesis has been widely supported, it is clear from this and other studies[7,18,19,33,42] that the development an os odontoideum may result from a different process. Some cases may be clearly traumatic in origin and as such represent odontoid fracture nonunions, others have a congenital etiology and there may be a few that represent a combination of the two.
Numerous case reports and case series attest to the rarity of this anomaly and neither the overall incidence nor the prevalence is well documented. The closest (albeit flawed) estimate is provided by Sankar et al.,[33] who noted a 3.1% prevalence of os odontoideum following a review of all abnormal radiographs at the Children's Hospital of Philadelphia. In this series of 16 patients, only 3 individuals were identified as having a clinical history compatible with a traumatic etiology, while 6 harbored associated cervical spine anomalies. If one accepts the existence of both a traumatic and congenital etiology,[33] the question of whether these two separate etiologies result in distinct clinical disorders, especially with regards to ligamentous competency and overall spinal stability must be addressed as this may impact the natural history and accordingly treatment recommendations.
This study identified a litany of significant differences in gene expression (213/380 genes in total) between the Twins with the non-os group. Notably, these genes have biological functionality related to bone formation and maintenance. PHEX, a gene involved in bone mineralization, was elevated 447-fold in the Twins. TUFT1, another gene related to mineralization (of enamel), was increased 109-fold. TFGBI, a gene that mediates cell–collagen interactions and is thought to be involved in endochondral bone formation, was elevated 92-fold. MMP8, a neutrophil MMP that breaks down collagen types I, II, and III, was elevated 90-fold in the Twins. RUNX2, encodes a transcription factor that induces osteoblastic differentiation and is critical in skeletal morphogenesis, was elevated 66-fold in the Twins. Though none of the expression profiles comparing the os patients to the non-os subjects held statistical significance, we found trends in genes similarly involved in bone development and maintenance. MMP8 again was elevated 95-fold; FGFR1 was elevated 25-fold; COL1A1 was elevated 15-fold and BMP3 was elevated 7-fold. Further investigation of these genes may aid in further understanding of the cellular and molecular pathogenesis of os odontoideum.
This study is compromised by the small sample size, which may, in part, be responsible for the lack of statistical significance in the trends in gene expression profile that we found between the os and non-os groups. We opted to include the gene expression profile from the Twins’ mother in order to control for potential hereditary gene expression patterns unrelated to os odontoideum. The comparison group does not contain age-matched cases, adding a potential confounding variable. There is also the limitation of data selection: There were over 200 genes with significant differences in expression profiles between the Twins and the non-os controls. The strategy employed to overcome this potential problem was to focus on those genes with the greatest magnitude of change. Though this approach allows for a concise and informative presentation of the results, it may have excluded some relevant gene profiles.
CONCLUSION
Os odontoideum has multiple etiologies, both congenital and traumatic and perhaps some cases represent a combination of the two. Further definition of each type and examination of their relative prevalence will be informative. Moreover, investigation of the relevance of this distinction as to the clinical evaluation, natural history, and treatment is appropriate. We have identified a number of genes that show increased expression in a pair of twins with congenital os odontoideum and also demonstrated trends in gene expression profiles between a larger group of os odontoideum patients and non-os patients. A number of these genes are related to bone morphogenesis and maintenance. Further investigations of the molecular biology of these genes may confer a greater understanding of this anomaly.
Footnotes
Available FREE in open access from: http://www.surgicalneurologyint.com/text.asp?2014/5/1/37/129259
Contributor Information
David Straus, Email: David_Straus@rush.edu.
Shunbin Xu, Email: Shunbin_Xu@rush.edu.
Vincent C. Traynelis, Email: Vincent_Traynelis@rush.edu.
REFERENCES
- 1.Al Kaissi A, Chehida FB, Ghachem MB, Grill F, Klaushofer K. Atlanto-axial segmentation defects and os odontoideum in two male siblings with opsismodysplasia. Skeletal Radiol. 2009;38:293–6. doi: 10.1007/s00256-008-0623-4. [DOI] [PubMed] [Google Scholar]
- 2.Arvin B, Fournier-Gosselin MP, Fehlings MG. Os odontoideum: Etiology and surgical management. Neurosurgery. 2010;66(3 Suppl):22–31. doi: 10.1227/01.NEU.0000366113.15248.07. [DOI] [PubMed] [Google Scholar]
- 3.Asagiri M, Takayanagi H. The molecular understanding of osteoclast differentiation. Bone. 2007;40:251–64. doi: 10.1016/j.bone.2006.09.023. [DOI] [PubMed] [Google Scholar]
- 4.Benjamini Y, Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J R Stat Soc B. 1995;57:289–300. [Google Scholar]
- 5.Brazma A, Hingamp P, Quackenbush J, Sherlock G, Spellman P, Stoeckert C, et al. Minimum information about a microarray experiment (MIAME)-toward standards for microarray data. Nat Genet. 2001;29:365–72. doi: 10.1038/ng1201-365. [DOI] [PubMed] [Google Scholar]
- 6.Clements WD, Mezue W, Mathew B. Os odontoideum--congenital or acquired?--that's not the question. Injury. 1995;26:640–2. doi: 10.1016/0020-1383(95)00123-q. [DOI] [PubMed] [Google Scholar]
- 7.Currarino G. Segmentation defect in the midodontoid process and its possible relationship to the congenital type of os odontoideum. Pediatr Radiol. 2002;32:34–40. doi: 10.1007/s00247-001-0579-1. [DOI] [PubMed] [Google Scholar]
- 8.Ellies DL, Krumlauf R. Bone formation: The nuclear matrix reloaded. Cell. 2006;125:840–2. doi: 10.1016/j.cell.2006.05.022. [DOI] [PubMed] [Google Scholar]
- 9.Fagan AB, Askin GN, Earwaker JW. The jigsaw sign. A reliable indicator of congenital aetiology in os odontoideum. Eur Spine J. 2004;13:295–300. doi: 10.1007/s00586-004-0732-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fielding JW, Griffin PP. Os odontoideum: An acquired lesion. J Bone Joint Surg Am. 1974;56:187–90. [PubMed] [Google Scholar]
- 11.Fielding JW, Hensinger RN, Hawkins RJ. Os Odontoideum. J Bone Joint Surg Am. 1980;62:376–83. [PubMed] [Google Scholar]
- 12.Freiberger RH, Wilson PD, jr, Nicholas JA. Acquired Absence of the Odontoid Process A case report. J Bone Joint Surg Am. 1965;47:1231–6. [PubMed] [Google Scholar]
- 13.Gadgil M, Lian W, Gadgil C, Kapur V, Hu WS. An analysis of the use of genomic DNA as a universal reference in two channel DNA microarrays. BMC Genomics. 2005;6:66. doi: 10.1186/1471-2164-6-66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Garcion C, Baltensperger R, Fournier T, Pasquier J, Schnetzer MA, Gabriel JP, et al. FiRe and microarrays: A fast answer to burning questions. Trends Plant Sci. 2006;11:320–2. doi: 10.1016/j.tplants.2006.05.009. [DOI] [PubMed] [Google Scholar]
- 15.Hadley MN. Os odontoideum. Neurosurgery. 2002;50:S148–55. doi: 10.1097/00006123-200203001-00023. [DOI] [PubMed] [Google Scholar]
- 16.Hawkins RJ, Fielding JW, Thompson WJ. Os odontoideum: Congenital or acquired. A case report. J Bone Joint Surg Am. 1976;58:413–4. [PubMed] [Google Scholar]
- 17.Karsenty G, Kronenberg HM, Settembre C. Genetic control of bone formation. Annu Rev Cell Dev Biol. 2009;25:629–48. doi: 10.1146/annurev.cellbio.042308.113308. [DOI] [PubMed] [Google Scholar]
- 18.Kirlew KA, Hathout GM, Reiter SD, Gold RH. Os odontoideum in identical twins: Perspectives on etiology. Skeletal radiology. 1993;22:525–7. doi: 10.1007/BF00209102. [DOI] [PubMed] [Google Scholar]
- 19.Klimo P, Jr, Coon V, Brockmeyer D. Incidental os odontoideum: Current management strategies. Neurosurg Focus. 2011;31:E10. doi: 10.3171/2011.9.FOCUS11227. [DOI] [PubMed] [Google Scholar]
- 20.Klimo P, Jr, Kan P, Rao G, Apfelbaum R, Brockmeyer D. Os odontoideum: Presentation, diagnosis, and treatment in a series of 78 patients. J Neurosurg Spine. 2008;9:332–42. doi: 10.3171/SPI.2008.9.10.332. [DOI] [PubMed] [Google Scholar]
- 21.Kovacs B, Lumayag S, Cowan C, Xu S. MicroRNAs in early diabetic retinopathy in streptozotocin-induced diabetic rats. Invest Ophthalmol Vis Sci. 2011;52:4402–9. doi: 10.1167/iovs.10-6879. [DOI] [PubMed] [Google Scholar]
- 22.Lian JB, Stein GS, Javed A, van Wijnen AJ, Stein JL, Montecino M, et al. Networks and hubs for the transcriptional control of osteoblastogenesis. Rev Endocr Metab Disord. 2006;7:1–16. doi: 10.1007/s11154-006-9001-5. [DOI] [PubMed] [Google Scholar]
- 23.Mao Z, Shay B, Hekmati M, Fermon E, Taylor A, Dafni L, et al. The human tuftelin gene: cloning and characterization. Gene. 2001;279:181–96. doi: 10.1016/s0378-1119(01)00749-1. [DOI] [PubMed] [Google Scholar]
- 24.McCarthy JG, Stelnicki EJ, Mehrara BJ, Longaker MT. Distraction osteogenesis of the craniofacial skeleton. Plast Reconstr Surg. 2001;107:1812–27. doi: 10.1097/00006534-200106000-00029. [DOI] [PubMed] [Google Scholar]
- 25.Menezes AH. Pathogenesis, dynamics, and management of os odontoideum. Neurosurg Focus. 1999;6:e2. doi: 10.3171/foc.1999.6.6.5. [DOI] [PubMed] [Google Scholar]
- 26.Morgan MK, Onofrio BM, Bender CE. Familial os odontoideum. Case report. J Neurosurg. 1989;70:636–9. doi: 10.3171/jns.1989.70.4.0636. [DOI] [PubMed] [Google Scholar]
- 27.Morriss-Kay GM, Wilkie AO. Growth of the normal skull vault and its alteration in craniosynostosis: Insights from human genetics and experimental studies. J Anat. 2005;207:637–53. doi: 10.1111/j.1469-7580.2005.00475.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.NCBI. NCBI Gene. [Last accessed on 2012 Oct 12]. Available from: http://www.ncbi.nlm.nih.gov/gene .
- 29.Peeters JK, Van der Spek PJ. Growing applications and advancements in microarray technology and analysis tools. Cell Biochem Biophys. 2005;43:149–66. doi: 10.1385/CBB:43:1:149. [DOI] [PubMed] [Google Scholar]
- 30.Provot S, Schipani E. Molecular mechanisms of endochondral bone development. Biochem Biophys Res Commun. 2005;328:658–65. doi: 10.1016/j.bbrc.2004.11.068. [DOI] [PubMed] [Google Scholar]
- 31.Ricciardi JE, Kaufer H, Louis DS. Acquired os odontoideum following acute ligament injury. Report of a case. J Bone Joint Surg Am. 1976;58:410–2. [PubMed] [Google Scholar]
- 32.Rieu I, Powers SJ. Real-time quantitative RT-PCR: Design, calculations, and statistics. Plant Cell. 2009;21:1031–3. doi: 10.1105/tpc.109.066001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sankar WN, Wills BPD, Dormans JP, Drummond DS. Os odontoideum revisited: The case for a multifactorial etiology. Spine (Phila Pa 1976) 2006;31:979–84. doi: 10.1097/01.brs.0000214935.70868.1c. [DOI] [PubMed] [Google Scholar]
- 34.Schmitt JM, Hwang K, Winn SR, Hollinger JO. Bone morphogenetic proteins: An update on basic biology and clinical relevance. J Orthop Res. 1999;17:269–78. doi: 10.1002/jor.1100170217. [DOI] [PubMed] [Google Scholar]
- 35.Schuler TC, Kurz L, Thompson DE, Zemenick G, Hensinger RN, Herkowitz HN. Natural history of os odontoideum. J Pediatr Orthop. 1991;11:222–5. doi: 10.1097/01241398-199103000-00017. [DOI] [PubMed] [Google Scholar]
- 36.Stains JP, Civitelli R. Cell-to-cell interactions in bone. Biochem Biophys Res Commun. 2005;328:721–7. doi: 10.1016/j.bbrc.2004.11.078. [DOI] [PubMed] [Google Scholar]
- 37.Stillwell WT, Fielding JW. Acquired os odontoideum. A case report. Clin Orthop Relat Res. 1978;135:71–3. [PubMed] [Google Scholar]
- 38.Teitelbaum SL, Ross FP. Genetic regulation of osteoclast development and function. Nat Rev Genet. 2003;4:638–49. doi: 10.1038/nrg1122. [DOI] [PubMed] [Google Scholar]
- 39.Tsiridis E, Giannoudis PV. Transcriptomics and proteomics: Advancing the understanding of genetic basis of fracture healing. Injury. 2006;37(Suppl 1):S13–9. doi: 10.1016/j.injury.2006.02.036. [DOI] [PubMed] [Google Scholar]
- 40.Verska JM, Anderson PA. Os odontoideum. A case report of one identical twin. Spine. 1997;22:706–9. doi: 10.1097/00007632-199703150-00026. [DOI] [PubMed] [Google Scholar]
- 41.Wan M, Cao X. BMP signaling in skeletal development. Biochem Biophys Res Commun. 2005;328:651–7. doi: 10.1016/j.bbrc.2004.11.067. [DOI] [PubMed] [Google Scholar]
- 42.Wang S, Wang C. Acquired os odontoideum: A case report and literature review. Childs Nerv Syst. 2012;28:315–9. doi: 10.1007/s00381-011-1642-3. [DOI] [PubMed] [Google Scholar]
- 43.Wang S, Wang C. Familial dystopic os odontoideum: A report of three cases. J Bone Joint Surg Am. 2011;93:e44. doi: 10.2106/JBJS.J.01018. [DOI] [PubMed] [Google Scholar]
- 44.Weston AD, Ozolins TR, Brown NA. Thoracic skeletal defects and cardiac malformations: A common epigenetic link? Birth Defects Res C Embryo Today. 2006;78:354–70. doi: 10.1002/bdrc.20084. [DOI] [PubMed] [Google Scholar]
- 45.Yamamoto T, Kurosawa K, Masuno M, Okuzumi S, Kondo S, Miyama S, et al. Congenital anomaly of cervical vertebrae is a major complication of Rubinstein-Taybi syndrome. Am J Med Genet A. 2005;135:130–3. doi: 10.1002/ajmg.a.30708. [DOI] [PubMed] [Google Scholar]
- 46.Yuan JS, Reed A, Chen F, Stewart CN., Jr Statistical analysis of real-time PCR data. BMC Bioinformatics. 2006;7:85. doi: 10.1186/1471-2105-7-85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zaidi M, Brain SD, Tippins JR, Di Marzo V, Moonga BS, Chambers TJ, et al. Structure-activity relationship of human calcitonin-gene-related peptide. Biochem J. 1990;269:775–80. doi: 10.1042/bj2690775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zygourakis CC, Cahill KS, Proctor MR. Delayed development of os odontoideum after traumatic cervical injury: Support for a vascular etiology. J Neurosurg Pediatr. 2011;7:201–4. doi: 10.3171/2010.11.PEDS10289. [DOI] [PubMed] [Google Scholar]