

ABSTRACT
African green monkeys (AGMs) are naturally infected with simian immunodeficiency virus (SIV) at high prevalence levels and do not progress to AIDS. Sexual transmission is the main transmission route in AGM, while mother-to-infant transmission (MTIT) is negligible. We investigated SIV transmission in wild AGMs to assess whether or not high SIV prevalence is due to differences in mucosal permissivity to SIV (i.e., whether the genetic bottleneck of viral transmission reported in humans and macaques is also observed in AGMs in the wild). We tested 121 sabaeus AGMs (Chlorocebus sabaeus) from the Gambia and found that 53 were SIV infected (44%). By combining serology and viral load quantitation, we identified 4 acutely infected AGMs, in which we assessed the diversity of the quasispecies by single-genome amplification (SGA) and documented that a single virus variant established the infections. We thus show that natural SIV transmission in the wild is associated with a genetic bottleneck similar to that described for mucosal human immunodeficiency virus (HIV) transmission in humans. Flow cytometry assessment of the immune cell populations did not identify major differences between infected and uninfected AGM. The expression of the SIV coreceptor CCR5 on CD4+ T cells dramatically increased in adults, being higher in infected than in uninfected infant and juvenile AGMs. Thus, the limited SIV MTIT in natural hosts appears to be due to low target cell availability in newborns and infants, which supports HIV MTIT prevention strategies aimed at limiting the target cells at mucosal sites. Combined, (i) the extremely high prevalence in sexually active AGMs, (ii) the very efficient SIV transmission in the wild, and (iii) the existence of a fraction of multiparous females that remain uninfected in spite of massive exposure to SIV identify wild AGMs as an acceptable model of exposed, uninfected individuals.
IMPORTANCE We report an extensive analysis of the natural history of SIVagm infection in its sabaeus monkey host, the African green monkey species endemic to West Africa. Virtually no study has investigated the natural history of SIV infection in the wild. The novelty of our approach is that we report for the first time that SIV infection has no discernible impact on the major immune cell populations in natural hosts, thus confirming the nonpathogenic nature of SIV infection in the wild. We also focused on the correlates of SIV transmission, and we report, also for the first time, that SIV transmission in the wild is characterized by a major genetic bottleneck, similar to that described for HIV-1 transmission in humans. Finally, we report here that the restriction of target cell availability is a major correlate of the lack of SIV transmission to the offspring in natural hosts of SIVs.
INTRODUCTION
Numerous African nonhuman primate (NHP) species have been shown to be naturally infected with species-specific simian immunodeficiency viruses (SIVs) (1–3). SIVs are at the origin of the human immunodeficiency virus (HIV) pandemic, with HIV type 1 (HIV-1) and HIV-2 arising from cross-species transmission from chimpanzees/gorillas and sooty mangabeys, respectively (4–6). Yet, while HIV infection is highly pathogenic and progresses to generalized immunodeficiency and AIDS, SIVs are generally believed to be nonpathogenic in their natural hosts (7–10), with only a few cases of progression to AIDS being reported thus far in African NHP species (11). However, the nonpathogenicity of SIVs in their natural hosts was concluded from the study of only a few host species (African green monkeys [AGMs], sooty mangabeys, and mandrills), and the vast majority of these studies were performed on captive monkeys (12–19). Over the last 2 decades, these studies consistently reported that the control of disease progression in the natural hosts of SIVs is due neither to control of viral replication nor to SIV-specific adaptive immune responses, as viral loads (VLs) in AGMs, sooty mangabeys, and mandrills are similar to those observed during pathogenic HIV/SIV infections (12, 14, 20–23) and adaptive immune responses are not quantitatively or qualitatively different from those mounted during pathogenic infections (24–30). Instead, the lack of disease progression was shown to be the result of adaptation of these natural hosts to control of the deleterious indirect consequences of SIV infection, i.e., excessive levels of immune activation, T cell proliferation, and apoptosis (23, 31–33).
A paradox of SIV infection in natural hosts is that high levels of viral replication occur in the context of reduced availability of target cells at mucosal sites. For example, uninfected AGMs have intrinsically low levels of CD4+ T cells (17, 18), and furthermore, their helper cells downregulate CD4 expression as they enter the memory pool, thus protecting the central-memory CD4+ T cells from virus-mediated killing (34, 35).
In addition to these adaptations aimed at preserving the target cells from being infected, AGMs, similar to multiple African NHP species, were reported to express very low levels of the CCR5 coreceptor on CD4+ T cells, especially at the mucosal sites (36). Previous studies from our lab suggest that this adaptation may represent a determinant factor impacting mucosal transmission in natural hosts of SIVs, especially in very young animals (19, 37). Again, these results were obtained in experimental transmission studies, which may or may not accurately reproduce SIV transmission in the wild.
In contrast to the observed low levels of target cells, epidemiological studies in wild African NHPs identified very high levels of SIVagm prevalence in adult AGMs (38–42). Since SIVagm prevalence dramatically increases with age, it is generally accepted that sexual transmission is the main route of SIVagm spread (38–40). Aggression and fights for dominance involving biting and exposure to blood also contribute to SIV transmission (43).
We have recently documented in a substantial cohort of wild vervet monkeys (Chlorocebus pygerythrus) from South Africa that these very high rates of transmission may be fueled by the high levels of viral replication observed in naturally SIVagmVer-infected vervets (40), which are higher than in chronically HIV-1-infected patients (44). In the same study, we documented a high proportion of acutely infected AGMs, again suggesting very active SIV transmission in the wild (40).
As such, a second paradox of SIVagm infection in the wild can be defined as high levels of prevalence of SIV infection in the context of low availability of target cells at mucosal sites. This raises the question of whether or not SIV transmission in the wild is truly comparable to transmission in pathogenic HIV-1 infection of humans or SIV infection of macaques. Recent studies have documented that in the pathogenic infections mucosal transmission is characterized by an extreme genetic bottleneck (45–47). Thus, characterization of the genetic diversity of the HIV population during the early stages of infection, when the transmitted strains can be inferred after single-genome amplification (SGA), established that in most of the cases of heterosexual transmission infection is founded by a single strain from the donor viral population (45, 48). In the case of intrarectal homosexual intercourse, more strains are transmitted, while in the case of transmission through intravenous drug use, which bypasses the mucosal bottleneck, multiple strains can be identified (49, 50).
This bottleneck effect is currently explained by the low efficiency of virus penetration through mucosal layers and the potential selective pressure at the sites of transmission in either the donor or the recipient that could shape the transmitted viral strains (50). In addition to the route of transmission, cofactors, such as the presence of a sexually transmitted infection or the use of hormonal contraceptives, may affect the complexity of the transmitted virus population (49). Recent studies have shown that the transmitted founder viral strains have selective properties that confer a higher propensity for transmission (i.e., they are more resistant to interferons) (51). Low-dose mucosal-transmission studies in rhesus macaques were also reported to recapitulate mucosal HIV-1 transmission (46).
To date, it is not known whether or not a similar bottleneck of transmission occurs in the wild. Our previous experimental results suggested that this might indeed be the case (37). Therefore, here we characterized SIV transmission in wild AGMs to investigate if a genetic bottleneck of transmitted variants is observed in naturally infected AGMs in the wild or if the high prevalence is due to a higher mucosal susceptibility to SIV infection. Given the continuous spread of the HIV pandemic and the absence of an effective cure or vaccine for AIDS, such studies of natural SIV transmission in the wild may be of pivotal importance in developing new strategies to control HIV transmission and disease progression (52).
MATERIALS AND METHODS
Samples.
The study was carried out in wild sabaeus AGMs (Chlorocebus sabaeus) (n = 121) living in different areas in the Gambia: the Western division (Abuko Forest Park, n = 27; Bijilo Forest Park region [Bijilo I, n = 59, and Bijilo II, n = 8]) and the Central division (Janjanbureh town region [Janjanbureh I, n = 17, and Janjanbureh II, n = 4]; River Gambia National Park Region, n = 3; and Yorobeli Kunda, n = 3) (Fig. 1) (53). Animals ranged from infants (6 months) to old adults (>10 years), and the AGMs included in the study were stratified as infants, juveniles, and adults, based on age classes determined upon dental eruption patterns (54). Animals were individually trapped using established methods (55). Details on animal capture and blood collection through venous puncture are provided elsewhere (40, 53). Each monkey included in the study had a microchip implanted for further identification and prevention of duplicate sampling. The volume of blood collected from each animal varied based on the body weight (which was determined before the blood draw) and age and generally ranged between 8.5 and 25.5 ml. The volume of blood collected from infants was significantly lower (1 to 3 ml).
FIG 1.
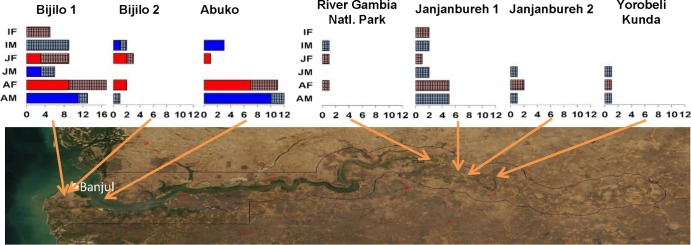
Satellite map of the Gambia indicating the geographical origin of the samples included in this study. Sabaeus monkey samples collected from the different locations are detailed. Monkeys for which both PBMCs and plasma samples were available are depicted as vivid colors (red, females; blue, males); sabaeus monkeys from which only plasma samples were available are depicted as light, patterned colors (light red, females; light blue, males). Sex and age group coverage is illustrated.
Upon sample collection, a detailed clinical assessment of the monkeys (including signs associated with immunodeficiency, i.e., lymphadenopathy, low weight, fever, or rashes) was performed. Plasma was separated through centrifugation (1,700 rpm), aliquoted, and stored at −80°C. A 500-μl aliquot of plasma from each monkey was available for this study. Peripheral blood mononuclear cells (PBMCs) were isolated from 56 animals (Fig. 1) as previously described (18, 56). Due to the low volume of blood collected from infant AGMs, PBMCs were isolated only from a small fraction of infants. All animal procedures employed in this study followed the regulations set forth by the Animal Welfare Act and were approved by the University of Wisconsin—Milwaukee (UWM) Institutional Animal Care and Use Committee (IACUC). Studies were carried out through an agreement with UWM.
Sample processing and testing strategy.
Upon arrival, plasma samples were thawed and separated into three aliquots. The first aliquot was used for RNA extraction and reverse transcription-PCR to assess the SIV prevalence levels. The same aliquot was used for quantification of SIVagm VLs by real-time PCR using primers and probes designed on the basis of SIVagm integrase sequences. The second aliquot was used for SIV enzyme-linked immunosorbent assay (ELISA) to establish the stage of SIV infection and infant exposure to SIVagm. Correlation between VLs and serology enabled identification of acutely SIV-infected AGMs (40). SGA was then performed on the RNA extracted from the first aliquot to assess the bottleneck of SIVagmSab transmission. The third aliquot was used to assess the natural history of SIVagm infection in the wild by testing surrogate markers of microbial translocation (sCD14), immune activation, inflammation (cytokine/chemokine levels and C-reactive protein [CRP]), and hypercoagulability (D dimer [DD]). These markers were previously shown in the SMART trial to predict progression to AIDS and death in HIV-infected patients (57–59).
PBMC samples were first tested for viability and then by flow cytometry to assess the major immune cell populations and levels of immune activation and proliferation.
Viral RNA extraction and cDNA synthesis.
From each plasma specimen, viral RNA was extracted using the QIAamp Viral RNA minikit (Qiagen, Germantown, MD). RNA was eluted and immediately subjected to cDNA synthesis. The RNA was first verified for its size in order to avoid any unspecific RNA smears or bands which could bias the accurate estimate of the RNA concentration. Reverse transcription of RNA to single-stranded cDNA was performed using SuperScript III reverse transcriptase according to the manufacturer's recommendations (Invitrogen, Carlsbad, CA). In brief, each cDNA reaction mixture included 1× reverse transcription buffer, 0.5 mM each deoxynucleoside triphosphate, 5 mM dithiothreitol, 2 U/ml RNaseOUT (RNase inhibitor), 10 U/ml of Super-Script III reverse transcriptase, and 0.25 μM antisense primer SIV-POL-OR (5′-ACB ACY GCN CCT TCH CCT TTC-3′) or SIV-ENV-B (5′-AGA GCT GTG ACG CGG GCA TTG AGG TT-3′). The mixture was incubated at 50°C for 60 min, followed by an increase in temperature to 55°C for an additional 60 min. The reaction mixture was then heat inactivated at 85°C for 5 min and treated with 2 U of RNase H at 37°C for 20 min. The newly synthesized cDNA was used immediately or frozen at −80°C.
Nested PCR and DNA gel extraction.
A 600-bp pol integrase fragment was amplified by nested PCR using outer primers POL-IS4 (5′-CCA GCN CAC AAA GGN ATA GGA GG-3′) and POL-OR and inner primers SIV-POL-IS4 and SIV-POL-Unipol2 (5′-CCC CTA TTC CTC CCC TTC TTT TAA AA-3′) (3, 60). A 400-bp gag fragment was amplified by nested PCR using outer primers GAG-UP-F1 (5′-ATT CAG TGC AGA AGT AGT GCC CAT-3′) and GAG-LOW-R1 (5′-CCA ATT CTT TAC TGC TGG GTC TGT-3′) and inner primers GAG-UP-F2 (5′-CAG AAG GAG CAA TCC CAT ATG ACA-3′) and GAG-LOW-R2 (5′-CTG AGA GCC TTG TAG AAT CTA TCC AC-3′) (61). Finally, a 900-bp env fragment encompassing the V3-V5 gp120 region and the 5′ end of gp41 envelope regions was amplified by nested PCR using outer primers ENV-A (5′-GAA GCT TGT GAT AAA ACA TAT TGG AT-3′) and ENV-B (5′-AGA GCT GTG ACG CGG GCA TTG AGG TT-3′) and inner primers ENV-C (5′-GTG CAT TGT ACA GGG TTA ATG AAT ACA ACA G-3′) and ENV-D (5′-TTC TTC TGC TGC AGTA TCC CAG CAA G-3′) (62). Nested PCR products were subjected to 1% agarose (Invitrogen, Carlsbad, CA) gel electrophoresis and extracted and purified using the QIAquick gel extraction kit (Qiagen, Germantown, MD).
SGA of SIVagm env genes.
To determine the number of transmitted founder viruses in newly infected animals, single-genome amplification of viral sequences was performed as described previously (37, 63).
Briefly, viral RNA was extracted from plasma samples with an EZ1 virus minikit (version 2.0; Qiagen, Valencia, CA) and reverse transcribed using primer SIVagmENVout-R (5′-GTACCTGGCCCATCAGTGTAATTCTGC-3′) and SuperScript III reverse transcriptase. The first-strand synthesis reaction mixture contained 1× reverse transcription buffer, 0.5 mM each deoxynucleoside triphosphate, 5 mM dithiothreitol, 2 U/μl of RNaseOUT reagent, 10 U/μl of SuperScript III reverse transcriptase, and 0.25 M antisense primer. Dilutions of this cDNA to determine the dilution at which no more than 30% of reactions yielded amplicons were performed to ensure that most positive reaction mixtures contained a single template molecule. A 909-bp fragment of env genes was amplified using the SIV-ENV-A/SIV-ENV-B outer primers and the SIV-ENV-C/SIV-ENV-D nested primers. PCR was performed using Platinum Taq High Fidelity polymerase (Invitrogen, Carlsbad, CA) in the presence of 1× PCR buffer, 2 mM MgSO4, 0.2 mM each deoxynucleoside triphosphate, 0.2 μM each primer, and 0.025 U/μl of polymerase in a 20-μl reaction mixture. PCR conditions were 94°C for 2 min, followed by 35 cycles of 94°C for 15 s, 55°C for 30 s, and 68°C for 1 min (first round) or 35 cycles with a 56°C annealing temperature (second round), followed by a final extension of 10 min at 68°C. Amplicons were inspected using 96-well E-gels (Invitrogen, Carlsbad, CA) and directly sequenced.
DNA sequencing.
SIV gene amplicons were directly sequenced by the University of Pittsburgh Genomics and Proteomics Core laboratories (GPCL) using the nested PCR primers. The individual sequence for each amplicon was edited and inspected using BioEdit 7.1.3.
Phylogenetic analyses.
The SIVagmSab sequences for fragments of gag, pol, and env genes were aligned using the MUSCLE sequence alignment software. For comparison, SIV/HIV reference sequence alignments corresponding to the same regions, with sequences published until 2012, were obtained from the Los Alamos National Laboratory (LANL) HIV Sequence Database (http://hiv-web.lanl.gov). Eighty-five to 88 sequences were retained from these reference alignments, including all major SIV and HIV lineages. Each reference alignment was manually aligned with the SIVagmSab sequence alignments using JalView (64). The SIVagmSab alignments corresponded to nucleotide positions 670 to 1023, 2308 to 2980, and 991 to 3175 of the gag, pol, and env reference alignments. Genetic diversity was measured, both within the Gambian SIVagmSab cluster and between other SIVagm clusters from the reference alignment, using MEGA v5.10 (65). Recombination was screened for by using Recco (66), SBP/GARD (67), and SplitsTree (68). Maximum likelihood molecular phylogenetic trees were computed separately for gag, pol, and env alignments, using RAxML v7.4.6 (69) using the general time reversible (GTR) substitution model and gamma distributed rate heterogeneity. Support values for bipartitions were assigned using 1,000 bootstrap replicates and RAxML. Phylogenetic trees were visualized via FigTree (http://tree.bio.ed.ac.uk).
For the SGA samples, maximum likelihood trees were constructed using RAxML, and sequence variation was visualized using the Highlighter tool provided by the LANL HIV Sequence Database (http://hiv-web.lanl.gov).
Viral load quantification.
We used the generated pol alignment to design specific primers and probe for VL quantification in wild C. sabaeus: SIV-pol-standard-F (5′-AGG AGG ATC ATG ACA AGT ACC ATG C-3′), SIV-pol-standard-R (5′-GCT TCA ACA GGA ACT TAG CTG TTT C-3′), and SIV-pol-standard-Probe (5′-/56-FAM/CCA GCA GTG/ZEN/GTG GCA AAG GAG A/3IABkF/-3′). A second set of primers and probe mapping the gag sequences were synthesized, as follows: SIV-gag-standard-F (5′-ATA GCA GGG ACC ACT AGC ACA AT-3′), SIV-gag-standard-R (5′-TCT TTG AAT GGT TCC TTG GGT CC-3′), and fluorogenic SIV-gag-standard-Probe (5′-/56-FAM/ATA GCA GGG/ZEN/ACC ACT AGC ACA ATA C/3IABkF/-3′). All primers and probes were synthesized by IDTDNA (Coralville, IA) and were used in a two-step real-time PCR assay. First, reverse transcription of RNA to single-stranded cDNA was performed using the SuperScript III First-Strand Synthesis SuperMix for qRT-PCR kit according to the manufacturer's recommendations (Invitrogen, Carlsbad, CA). Real-time PCR was performed in MicroAmp Optical 96-well plates (Applied Biosystems, Branchburg, NJ) by mixing TaqMan PCR Master Mix (Applied Biosystems, Branchburg, NJ) with 5 μl isolated RNA in a 50-μl final reaction volume. Real-time PCR conditions were as follows: 15 min at 95°C, followed by 45 cycles of 95°C for 15 s and 60°C for 1 min. Dilutions of all components were made using sterile RNase-free water. Data were collected and analyzed using the PE Applied Biosystems software. RNA copies/well were adjusted to copies per milliliter of plasma. Samples were tested in duplicate, and the numbers of RNA copies were determined by comparison with a standard curve obtained using known amounts of SIV-pol or SIV-gag RNA standards. The RNA standard was produced by cloning one of the gag or pol gene fragments into Vector pBluescript SK+ and linearizing it by SacI as a DNA template for RNA in vitro transcription. Standard RNA was produced using the MEGAscript kit (Applied Biosystems, Branchburgh, NJ) according to the manufacturer's instructions. Briefly, the transcription reaction system was assembled at room temperature and mixed thoroughly, followed by incubation at 37°C for 4 h. One microliter of Turbo DNase was added to the reaction mixture, followed by incubation at 37°C for 15 min to digest the DNA template. RNA was recovered by lithium chloride precipitation. The RNA was quantified at A260, aliquoted, and immediately stored at −80°C. The detection limit of the SIVagmSab quantification assay was 100 copies/ml.
Flow cytometry analysis of major immune populations.
Within hours of sample collection, PBMCs were isolated as described previously (56, 70, 71) and frozen in liquid nitrogen until used. Immunophenotyping of mononuclear cells isolated from the blood of wild C. sabaeus monkeys was performed using fluorescently conjugated monoclonal antibodies in multiparameter panels. Data were acquired on an LSR-II flow cytometer (Becton, Dickinson) and analyzed using FlowJo (Tree Star, Inc.). The following monoclonal antibodies (MAbs) were used for flow cytometry: anti-CD3-Pacific blue (clone SP34), anti-CD4-APC (clone L200), anti-CCR5-PE (clone 3A9), anti-HLA-DR-APC-Cy7 (clone L243), anti-CD28-PE (clone L293), anti-Ki-67-FITC (clone B56), anti-CD14 (clone M5E2), anti-CD20 (clone 2H7), anti-CD11c (SHCL-3), anti-CD103 (clone 2G5) (BD Bioscience), anti-CD95-CyChrome (clone DX2) (BD Pharmingen), and CD8αβ-PE-Texas Red (clone 2ST8.5H7) (Beckman Coulter). We have shown all MAbs to be cross-reactive for sabaeus monkeys in our previous work (17, 32, 72). For surface staining, mononuclear cells from blood were stained using monoclonal antibodies by incubation at 4°C for 30 min in fluorescence-activated cell sorter (FACS) buffer. Cells were then washed with FACS buffer and fixed with BD stabilizing fixative (BD Bioscience). For intracellular stains, cells were first stained for surface markers and washed with FACS buffer and then fixed and permeabilized with Cytofix/Cytoperm fixation/permeabilization solution (BD Biosciences) at room temperature for 20 min, washed with Cytofix/Cytoperm Buffer (BD Biosciences), incubated with Ki-67-FITC in Cytofix/Cytoperm Buffer at 4°C for 40 min, and finally washed with Cytofix/Cytoperm buffer prior to acquisition.
Serology.
Based on the env sequence data, a peptide mapping the immunodominant region of the Gp41 transmembrane glycoprotein was synthesized and used in a peptide ELISA as described previously (73). The inferred peptide sequence was TALEKYLEDQARLNIWGCAFRQVC, and this sequence was very well conserved between different SIVagmSab strains, including the previously reported reference strains SIVagmSab1 and SIVagmSab92018 (63). The Gp36 peptide was synthesized to a purity of at least 90% (Fisher Scientific, USA), and the assay was performed as previously reported (73). The cutoff for the reaction was arbitrarily set at 0.20, as per previous reports (73).
MT.
Microbial translocation (MT) was assessed using levels of soluble CD14 (sCD14) as a surrogate marker (74). CD14 is a transmembrane protein that also exists in soluble form (sCD14; both as a shed membrane form and an alternatively spliced form), as a part of the complex that presents endotoxin (lipopolysaccharide [LPS]) to Toll-like receptor 4 (TLR4) on monocytes. When monocytes are activated, ectodomain shedding results in increased sCD14 levels. sCD14 is therefore a surrogate for direct measurement of endotoxin or Gram-negative bacteria that translocate from the intestinal lumen to the general circulation as a result of the immunologic injury inflicted at the mucosal level by pathogenic HIV/SIV infection (74–77). sCD14 levels were measured using a quantitative sandwich enzyme immunoassay technique (Quantikine Human sCD14 immunoassay; R&D Systems, Minneapolis, MN). The detection limit of this kit is 200 ng/ml and can reach up to 5,000 ng/ml, with an interassay coefficient of variability of 7.19% to 10.9%.
Coagulation status.
Coagulation status was estimated by determining plasma levels of D dimer (DD). D dimer is a fibrin degradation product (FDP), a small protein fragment present in the blood after a blood clot is degraded by fibrinolysis. It is so named because it contains two cross-linked D fragments of the fibrinogen protein. DD increases during the activation states of coagulation, disseminated intravascular coagulation, and deep vein thrombosis. DD was reported to independently correlate with lentiviral disease progression and death in HIV-infected patients (57) and SIV-infected macaques (76). DD was measured using a Star automated coagulation analyzer (Diagnostica Stago) and an immunoturbidimetric assay (Liatest D-DI; Diagnostica Stago). Our previous studies optimized this assay for use in AGMs (76).
CRP testing.
C-reactive protein (CRP) is an acute-phase protein that rises in the plasma in response to inflammation. It was first identified as a substance in the serum of patients with acute inflammation that reacted with the C polysaccharide of Pneumococcus. It binds to the phosphocholine expressed on the surface of dead cells and some types of bacteria in order to activate the complement system via the C1Q complex. The SMART trial identified CRP as one of the biomarkers associated with death in HIV-infected patients (57). CRP was measured using a monkey CRP ELISA kit (Life Diagnostics).
Cytokine and chemokine testing.
Cytokine testing in plasma was done using a sandwich immunoassay-based protein array system, the Cytokine Monkey Magnetic 28-Plex Panel (Invitrogen, Camarillo, CA), as instructed by the manufacturer, and results were read by the Bio-Plex array reader (Bio-Rad Laboratories, Hercules, CA), which uses Luminex fluorescent-bead-based technology (Luminex Corporation, Austin, TX).
Statistical analyses.
Correlation analyses within the same group of animals for two different parameters were performed using Spearman's rank correlation test with α equal to 0.05. The Mann-Whitney U test (two-tailed; α < 0.05) was used to analyze the difference between the median percentage of specific immunological markers between SIV-infected and uninfected AGMs. These statistical analyses were performed using GraphPad Prism (version 4.0b) software (Graph-Pad, San Diego, CA).
Nucleotide sequence accession numbers.
Nucleotide sequences of the gag, pol, and env sequences from SIVagmSab-infected sabaeus AGMs from the Gambia were deposited in GenBank under accession numbers KJ467354 to KJ467498.
RESULTS
Clinical data.
At the time of sample collection, a thorough clinical assessment of all AGMs included in this study failed to document any clinical sign associated with SIV infection, including fever or weight loss. The cross-sectional nature of this study precluded a direct assessment of changes in monkey weight and thus a direct assessment of weight loss. Therefore, we performed comparative analyses of the body mass index (BMI) between SIV-positive and SIV-negative monkeys (40) and found that SIV status did not impact the body condition of wild sabaeus monkeys (data not shown).
Prevalence of SIVagmSab infection in wild sabaeus monkeys from the Gambia.
Blood samples were collected from 121 wild-trapped sabaeus monkeys over a 2-month period. Details on monkey capture and sample collection are presented elsewhere (40). The samples included represent 7 free-ranging populations located in different regions of the Gambia covering a significant proportion of sabaeus habitat in this region (Fig. 1). The two sexes were equally represented. Samples from infants (<1 year old) and juvenile AGMs were also included, even though these volumes were limited.
PCR using env, gag, and pol primers identified 53/121 (44%) of the tested sabaeus monkeys as being SIVagmSab infected. In general, the prevalence levels were relatively similar among the different locations (data not shown), but no virus strain was amplified from the samples collected from sabaeus monkeys from the River Gambia National Park or from Yorobeli Kunda locations, from where very few samples were collected (Fig. 1).
SIVagmSab infection was unevenly distributed in different age groups: only 1/29 (4%) of the tested infants was found to be infected with SIVagmSab, in contrast to 7/25 (28%) of the juveniles and 45/67 (67%) of the adults. Also, while no significant sex-related difference in prevalence was observed in infant and juvenile monkeys (with overall prevalence of 16% in males versus 14% in females) (Table 1), these differences were significant in adult monkeys (36% in males versus 90% in females, P = 0.0001) (Table 1). The range of SIVagmSab prevalence levels was similar to that of levels found in AGMs in previous studies (38–42, 78). As in our previous study on wild AGMs (40) and in contrast to findings from previous reports (79, 80), we identified SIVagmSab infection in an infant sabaeus monkey in the wild, thus confirming that maternal-to-infant transmission can occur in the wild at low levels of prevalence (less than 5%).
TABLE 1.
SIVagmSab infection prevalence in wild sabaeus AGMs (Chlorocebus sabaeus) from the Gambia
| Age group | Sex | Total no. of AGMs tested | No. of AGMs positive for SIV | Prevalence (%) | Overall prevalence (%) |
|---|---|---|---|---|---|
| Infant | M | 21 | 1 | 5 | 4 |
| F | 8 | 0 | 0 | ||
| Juvenile | M | 11 | 4 | 36 | 28 |
| F | 14 | 3 | 21 | ||
| Adult | M | 28 | 10 | 36 | 67 |
| F | 39 | 35 | 90 | ||
| Overall | M | 60 | 15 | 25 | 44 |
| F | 61 | 38 | 62 |
Phylogenetic analysis.
In order to characterize the SIVagmSab diversity in naturally infected AGMs in the wild in the Gambia, we amplified and sequenced the gag (∼400-bp), pol (∼600-bp), and env (∼1,800-bp) regions of the SIVagm viruses from plasma samples. These correspond to absolute nucleotide positions relative to SIVmac239 1575 to 1971 (capsid), 4454 to 5105 (integrase), and 7384 to 9190 (gp160), respectively. We obtained 47 pol, 47 env, and 51 gag fragments from a total of 121 plasma samples. These were aligned with available reference sequences.
The sampled SIVagmSab genetic sequences exhibited extensive genetic diversity, although less so than vervet monkeys, potentially due to the more-limited geographic range of sabaeus monkeys. The diversity (average nucleotide substitutions per site) for the cluster of SIVagmSab sequences from the Gambia was measured at 0.139 ± 0.013 nucleotide substitutions per site for gag, 0.156 ± 0.011 for pol, and 0.120 ± 0.006 for env. Comparative genetic diversity of SIVagmVer sequences from vervet monkeys for these regions was measured at 0.167 ± 0.016 for gag, 0.189 ± 0.014 for pol, and 0.194 ± 0.009 for env.
Phylogenetic analysis revealed a tendency for distinct viral clusters corresponding to geographic regions within the Gambia coupled with mixing between populations (Fig. 2). Two genetically divergent clusters exist within the Bijilo I location alone, with sporadic intermixing with strains from the Abuko Forest Park and Bijilo II locations. Recombination was tested for in the gag, pol, and env regions; the analysis indicated significant support for recombination in the env gene between the Gambian sequences (Fig. 3). Significant P values for recombination were detected by the SplitsTree phi test for recombination (P < 0.001), and both Recco and SBP/GARD indicated strong support (P < 0.001) for a breakpoint occurring around position 2130 in the env alignment (position ∼900 in the env fragment), further supporting extensive intermixing between SIVagmSab populations in the Gambia.
FIG 2.

Phylogenetic tree inferred from gag, env, and pol gene sequences. SIVagmSab sequences from the Gambia are shaded and colored according to the geographic region from which they were sampled. Bootstrap support values are shown at internal nodes, with low bootstrap values (<80%) and regions with densely populated nodes omitted for clarity. The boxed sequences represent samples for which SGA analysis was performed.
FIG 3.

Split network from env gene sequences generated using the NeighborNet algorithm as implemented in SplitsTree. The network indicates extensive recombination, and thus mixing, between SIVagmSab populations throughout the Gambia. The boxed sequences represent samples for which SGA analysis was performed.
As expected, the SIVagmSab samples from the Gambia were closest to the existing SIVagmSab strain in all regions (Table 2). In both the pol and env phylogenies, the SIVagmSab sequences cluster together and closest to SIVagm strains from the other AGM species. In the gag phylogeny, the SIVagmSab sequences cluster together but no longer with the other SIVagm strains (Fig. 2). This recombinant genome architecture of SIVagmSab has previously been noted (81). Differences in topologies between these studies may be explained by the relatively short gag fragment (∼400 bp), combined with high genetic sequence divergence and poor resolution of internal nodes in the SIV tree, making inferences of the SIV strains most closely related to SIVagmSab and other SIVagm strains difficult to resolve.
TABLE 2.
Number of nucleotide substitutions per site between Gambian SIVagmSab strains and groups of other SIVagm strains for gag, pol, and env genesa
| Gene | Avg no. of nucleotide substitutions ± SE |
|||
|---|---|---|---|---|
| SIVagmVer | SIVagmGrv | SIVagmTan | SIVagmSab | |
| gag | 0.389 ± 0.037 | 0.467 ± 0.045 | 0.430 ± 0.042 | 0.162 ± 0.016 |
| pol | 0.321 ± 0.022 | 0.334 ± 0.025 | 0.312 ± 0.023 | 0.206 ± 0.016 |
| env | 0.423 ± 0.018 | 0.408 ± 0.019 | 0.427 ± 0.022 | 0.235 ± 0.013 |
Staging of SIVagmSab infection in wild sabaeus monkeys from the Gambia.
We have previously reported that a combination of VL and serological testing can identify acutely infected NHPs in the wild (40). We employed the same algorithm to stage SIVagmsab infection in wild sabaeus monkeys from the Gambia. We defined consensus SIV gag and pol sequences based on the alignments generated here and used these consensus sequences to design primers and probes for real-time PCR quantification. The high degree of conservation of the integrase region allowed perfect coverage of SIVagmSab diversity. VL quantification in the 53 SIVagmSab-infected sabaeus monkeys in our group showed that plasma VLs ranged from 103 to 107 vRNA copies/ml (Fig. 4a), in close range to those determined in experimentally infected sabaeus monkeys (20). Interestingly, SIVagmSab VLs were significantly lower in sabaeus monkeys than in wild vervets from South Africa infected with SIVagmVer (40) (average geometric means, 4.24 ± 1.14 log SIVagmSab RNA copies versus 5.37 ± 0.72 log SIVagmVer RNA; P < 0.0001) (Fig. 4a), pointing to possible differences in the steady-state levels of viral replication between the different NHP species that are natural hosts of SIV. Similar to our previous study in vervets in South Africa (40), high VLs (>106 vRNA copies) were observed in 4 of 53 sabaeus monkeys (8%) (Fig. 4b). Interestingly, the highest viral loads, suggestive of acute SIVagmSab infection, occurred in 4 females (Fig. 4b). Of these, 1 was juvenile, while the remaining were young adult females. All of these animals were primiparous, which supports very active viral transmission in sexually active sabaeus, as well as very effective SIVagmSab transmission in the wild (likely after the first mating/sexual encounter).
FIG 4.

Staging of SIVagmSab infection in wild sabaeus monkeys from the Gambia. (a) Significantly lower plasma viral load levels in sabaeus monkeys from the Gambia than in vervets from South Africa; (b) comparative assessment of the viral load levels between female AGMs and male AGMs from the Gambia; (c) serological testing of anti-gp41 antibodies in wild sabaeus monkeys. Infant AGMs are illustrated as open circles and squares; juvenile AGMs are illustrated as gray circles and squares; adult AGMs are shown as black circles and squares. In panels b and c, samples collected from female AGMs are illustrated as circles (○); samples collected from male AGMs are shown as squares (□); the samples from acutely infected adult females (defined as having high viral loads and negative serologies) are illustrated as open circles with enhanced margins; the sample collected from an acutely infected juvenile female is illustrated as V; finally, two samples from juvenile monkeys with high viral loads but seropositive results (probably postacutely infected) are shown as half-empty symbols and are identified by arrows. Detection limit of the VL assays, 100 copies/ml. CO, cutoff for the serological assay (arbitrarily established at 0.2) (73). OD, optical density of the serum sample.
To confirm that the high VLs occurred in acutely infected sabaeus monkeys, we employed a gp41 peptide ELISA and assessed the levels of anti-SIVagmSab antibodies in all samples included in this study. As shown in Fig. 4c, while the majority of SIVagmSab-infected sabaeus monkeys harbored detectable levels of anti-gp41 antibodies, the AGMs presenting with high VLs were seronegative. These results strongly suggest that AGMs with high VLs were acutely infected (82). Conversely, two samples from juvenile AGMs that have higher VLs than the majority of the tested samples (albeit lower than those observed in the 4 acutely infected AGMs) had detectable levels of anti-Gp41 antibodies, which suggested a more advanced stage of SIVagm infection (Fig. 4b and c). We used the preseroconversion samples to infer the number of transmitted founder strains in these animals.
SIV transmission in natural hosts in the wild is characterized by a stringent genetic bottleneck.
In pathogenic HIV and SIV infections, a substantial population bottleneck occurs when the virus is transmitted through heterosexual contact, with only one virus being transmitted in 80% of cases (45, 83). As our epidemiological data suggested efficient SIVagm transmission in the wild, we next sought to assess whether the bottleneck also characterizes SIVagmSab transmission in AGMs or whether these natural hosts are more permissive to SIV infection, thus explaining the high rates of transmission observed in the context of reduced availability of mucosal target cells. We therefore enumerated the transmitted founder viruses in the wild SIV-infected sabaeus monkeys that we identified as being acutely infected by performing SGA analysis of env gene sequences.
Results are shown in Fig. 5. The number of transmitted variants was inferred from phylogenetic analyses and Highlighter plots as described previously (37, 45, 46). This analysis demonstrated that in each of the four acutely infected sabaeus monkeys there was a single founder strain. We confirmed these SGA results by comparing the strain diversity in acutely infected sabaeus monkeys with that observed in two chronically infected monkeys (Fig. 5, lower panels), in which we identified a significantly more divergent viral population. Our results demonstrate for the first time that a stringent strain selection, similar to that reported for HIV-1 (45, 47), occurs during the natural transmission of SIVs in natural hosts in the wild.
FIG 5.

Enumeration of transmitted founder viruses in SIV-infected African green monkeys in the wild. Single-genome amplification was used to generate SIVsab env sequences from the plasma of four AGMs diagnosed as acutely infected (VMT74182, VMT74497, VMT75142, and VMT75112) and two AGMs chronically infected (VMT74362 and VMT76297) (which were used as controls). Sequences were compared by Highlighter plot analysis as described previously (37), with the sequence length (in bp) indicated on the x axis and number of transmitted founder viruses (V) indicated on the y axis. Maximum likelihood phylogenetic trees were also generated from the nucleotide alignments (∼950 bp) and are presented. Tick marks indicate differences compared to the top sequence (red, T; green, A; blue, C; orange, G; gray, gap). The master sequences used for the Highlighter plots are boxed in Fig. 2 and 3.
SIVagmSab-infected sabaeus monkeys demonstrated a trend toward lower CD4+ T cell counts than uninfected animals in the wild. To assess the long-term immunological impact of SIVagmSab infection, we performed, for the first time in wild animals, a comprehensive immunophenotyping study on a significant number of infected and uninfected sabaeus monkeys. PBMCs were collected from 4 infants, 9 juveniles, and 43 adults. We assessed the impact of SIV infection on the major T cell populations and did not identify any significant change in the frequency of either CD4+ (Fig. 6a), CD8+ (Fig. 6b), or double negative (DN) CD4− CD8− T cells (Fig. 6c). There was, however, a trend toward lower CD4+ T cell counts in SIVagmSab-infected sabaeus monkeys than in uninfected ones (P = 0.0966) (Fig. 6a). Analysis of CD4+ T cell subsets showed that the overall lower frequency of the total CD4+ T cell population in SIVagmSab-infected monkeys is due to significantly lower frequencies of effector memory T cells (P = 0.0309) (Fig. 6d, e, and f). Note, however, that due to the differences in SIV prevalence between different age groups, the SIV-infected and uninfected groups are not age matched and that the bias of the SIV-infected group toward older ages might explain these differences. We therefore conducted more-detailed multiple regression analyses (using general linear models) for the CD4+ T cell subsets, taking into account simultaneously the age and gender of the animals. In this analysis, the combined impact of age and gender on CD4+ T cell counts resulted in the loss of significance for the difference in effector memory CD4+ T cells between SIVagmSab-infected and uninfected sabaeus monkeys. This is in agreement with previous studies reporting that the lower levels of CD4+ T cells observed in SIV-infected natural hosts of SIVs are age related rather than a consequence of SIV infection (12, 13, 34, 84).
FIG 6.

Cross-sectional analysis of the major immune cell populations and subsets in wild sabaeus monkeys from the Gambia. (a) CD4+ T cells; (b) CD8+ T cells; (c) double negative (CD4− CD8−) T cells; (d) naive (CD28− CD95+) CD4+ T cells; (e) central memory (CD28+ CD95+) CD4+ T cells; (f) Effector memory (CD28+ CD95−) CD4+ T cells; (g) B cells; (h) natural killer (NKG2b+) cells; (i) monocytes (CD14+); (j) plasmacytoid dendritic cells (Linneg HLA-DR+ CD123+); (k) myeloid dendritic cells (Linneg HLA-DR+ CD11c+). Infant AGMs are illustrated as open circles and squares; juvenile AGMs are illustrated as gray circles and squares; adult AGMs are shown as black circles and squares. Samples collected from female AGMs are illustrated as circles (○); samples collected from male AGMs are shown as squares (□); the samples from acutely infected adult females (defined as having high viral loads and negative serology) are illustrated as open circles with enhanced margins; the sample collected from a juvenile male AGM with high viral loads but seropositive (probably postacutely infected) is shown as a half-empty square. P values were calculated by the Mann-Whitney U test. The values on the y axes depict the proportion of the given immune cell population or subset.
We next compared the levels of other major immune cell populations between SIVagmSab-infected and uninfected sabaeus monkeys and found no significant differences between the two groups with regard to the levels of B cells (Fig. 6g), NK cells (Fig. 6h), monocytes (Fig. 6i), plasmacytoid dendritic cells (Fig. 6j), and myeloid dendritic cells (Fig. 6k). These results are in agreement with the view that, similar to what occurs in other natural hosts, SIV infection in AGMs does not result in a significant alteration of the immune homeostasis (7, 16, 52).
No significant increase in the levels of T cell activation and proliferation was seen in SIVagmSab-infected sabaeus monkeys in the wild. HIV infection is associated with an increase in the fraction of T cells expressing markers of activation and proliferation (85), the extent of which correlates directly with the level of viral replication and inversely with lower CD4+ T cell counts (86). Previous studies conducted in both AGMs and sooty mangabeys in captivity indicated that the levels of T cell activation and proliferation do not significantly increase during chronic SIV infection in natural hosts (16, 21, 23, 77, 87, 88). In this study, we investigated whether any difference in the percentage of activated and proliferating T cells can be observed between SIVagmSab-infected and uninfected sabaeus monkeys in the wild. We compared differences in T cell immune activation between SIV-uninfected and infected AGMs by assessing the expression of HLA-DR on CD4+ (Fig. 7a) and CD8+ T cells (Fig. 7b). We compared differences in T cell proliferation by assessing the expression of Ki-67 on CD4+ (Fig. 7c) and CD8+ T cells (Fig. 7d). As shown in Fig. 7, no discernible differences in the levels of CD4+ and CD8+ T cell activation and proliferation were observed between the SIVagmSab-infected and uninfected sabaeus monkeys. Of note, in SIVagmSab-infected sabaeus monkeys there was no correlation between the fractions of HLA-DR- or Ki-67-expressing T cells and the levels of viral replication (data not shown). Collectively, these data confirmed the results of studies conducted in captive NHP natural hosts of SIVs reporting that increased levels of CD4+ T cell activation and proliferation do not seem to be a direct reflection of higher viral antigenic load or CD4+ T cell counts (7, 8, 13, 89, 90).
FIG 7.

Cross-sectional assessment of the levels of T cell immune activation and proliferation in wild sabaeus monkeys from the Gambia. Immune activation was measured as the fraction of CD4+ (a) and CD8+ T cells (b) expressing HLA-DR. Cell proliferation was measured as the fraction of CD4+ and CD8+ T cells expressing Ki-67. Infant AGMs are illustrated as open circles and squares; juvenile AGMs are illustrated as gray circles and squares; adult AGMs are shown as black circles and squares. Samples collected from female AGMs are illustrated as circles (○); samples collected from male AGMs are shown as squares (□); the samples from acutely infected adult females (defined as having high viral loads and negative serologies) are illustrated as open circles with enhanced margins; the sample collected from a juvenile male AGM with high viral loads but seropositive (probably postacutely infected) is shown as a half-empty square. P values were calculated by the Mann-Whitney U test. The values on the y axes depict the fraction of activated or proliferating CD4+ or CD8+ T cells.
SIVagmSab transmission in the wild correlates with the availability of target cells.
We next sought to identify the immune correlates of the different transmission levels in wild sabaeus monkeys. We have previously reported that captive natural hosts express low levels of target cells (CD4+ T cells expressing the CCR5 chemokine receptor), especially at mucosal sites (36). We also reported that the efficacy of experimental mucosal (intrarectal and vaginal) (37) and breastfeeding (19) transmission in natural hosts of SIVs appears to be driven by target cell availability at mucosal sites (19, 37). We assessed CCR5 expression on circulating CD4+ T cells of the naturally SIV-infected and uninfected sabaeus monkeys in the wild (Fig. 8). FACS analysis showed that the fraction of CD4+ T cells expressing CCR5 was similar for SIV-infected and SIV-uninfected wild sabaeus monkeys (P = 0.9786). However, as the infants and juveniles from both groups expressed significantly lower levels of CCR5 than did adults (Fig. 8a) and the two groups were significantly biased by age, we next compared the levels of CD4+ T cells expressing CCR5 in SIV-infected and uninfected monkeys within similar age groups. While this analysis did not identify significant association between CCR5 expression on CD4+ T cells and risk of SIVagmSab infection in adult sabaeus monkeys, such an association could be established in the juvenile sabaeus group. SIV-infected juvenile sabaeus monkeys harbored higher levels of CCR5+ CD4+ T cells than uninfected juveniles (P = 0.0023). Finally, while the low prevalence of SIV infection in sabaeus infants precluded any statistical analysis, it should be noted that the SIV-infected infant exhibited the highest CCR5 expression on CD4+ T cells among this age group (Fig. 8a).
FIG 8.
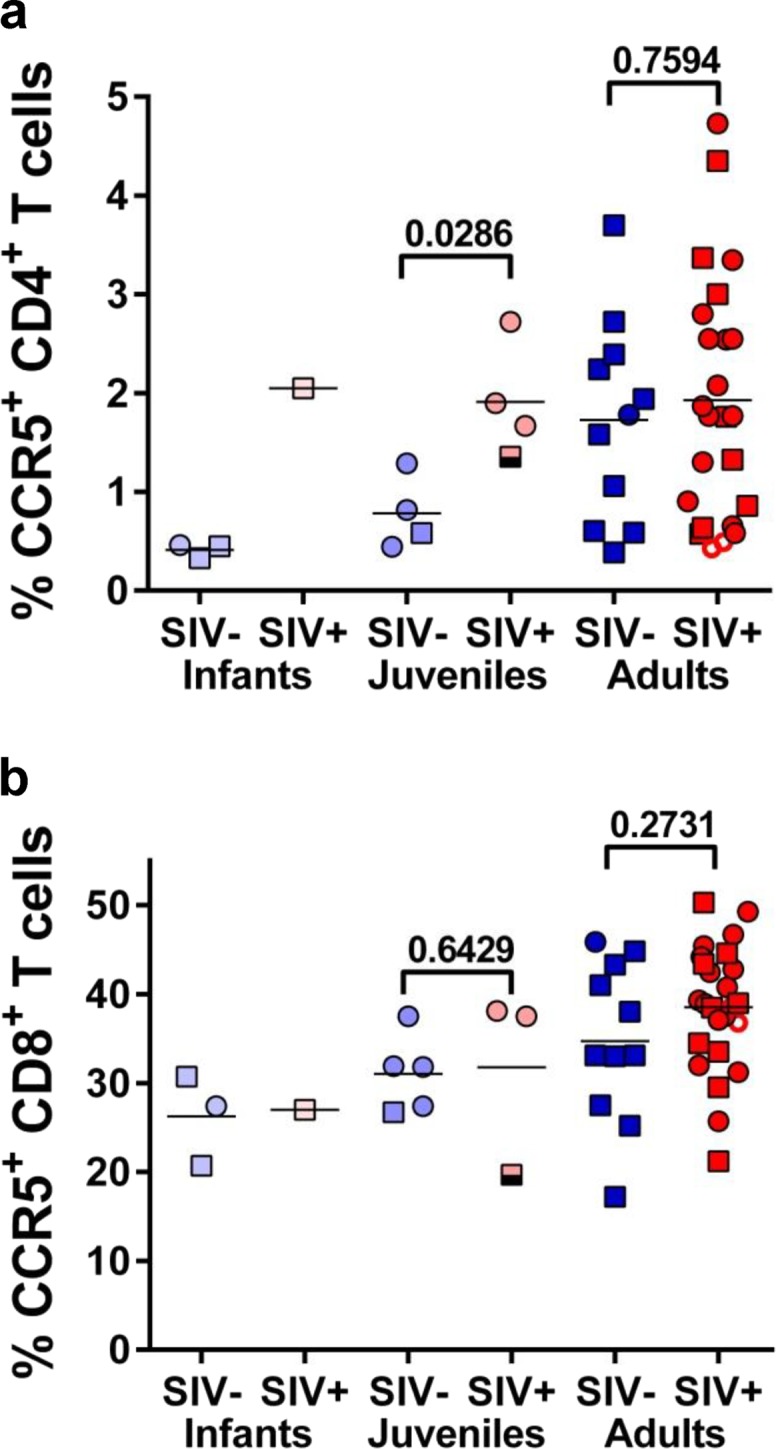
Cross-sectional assessment of the levels of T cells expressing the CCR5 coreceptor in wild sabaeus monkeys from The Gambia. CCR5 expression was assessed on both CD4+ T cells (a) and CD8+ T cells (b). Infant AGMs are illustrated as open circles and squares; juvenile AGMs are illustrated as light colored circles and squares; adult AGMs are shown as dark colored circles and squares. Samples collected from female AGMs are illustrated as circles (○); samples collected from male AGMs are shown as squares (□); samples from uninfected AGMs are illustrated in different tones of blue; samples collected from SIVsab-infected AGMs are illustrated in different tones of red; samples from acutely infected adult females (defined as having high viral loads and negative serologies) are illustrated as open circles with enhanced margins; a sample from a juvenile AGM male with high viral loads but seropositive (probably postacutely infected) is shown as a half-empty square. P values were calculated by the Mann-Whitney U test. The values on the y axes depict the fraction of CCR5-expressing CD4+ or CD8+ T cells.
The analysis of CCR5 expression on CD8+ T cells also identified significantly age-related differences (Fig. 8b), thus supporting an age-related maturation of CCR5 expression on T cells.
Taken together, these data confirm our previous observations in captive African nonhuman primate species, strongly supporting the paradigm that target cell availability determines susceptibility to infection in natural hosts of SIVs.
No significant difference in the biomarkers associated with disease progression and mortality between SIVagmSab-infected and uninfected sabaeus monkeys.
The SMART trial demonstrated that HIV-induced microbial translocation, inflammation, and hypercoagulation increase the risk of death among HIV-positive patients (57, 58). Furthermore, our previous studies showed that in SIV-infected NHPs increases in the levels of microbial translocation, immune activation, inflammation, and coagulation biomarkers are all strong predictors of disease progression (76). Therefore, in order to assess the pathogenic impact of SIV infection in wild animals, we investigated the markers associated with disease progression in the SMART trial as predictors of disease progression in SIVagmSab-infected sabaeus monkeys. Since the nature of this study precluded a prospective study, we compared the levels of biomarkers associated with death in the SMART trial between SIVagmSab-infected and uninfected monkeys. No significant difference was observed in the levels of sCD14 (Fig. 9a), interleukin-6 (IL-6) (Fig. 9b) and C-reactive protein (Fig. 9c) between SIVagmSab-infected and uninfected AGMs. Conversely, the levels of D dimer were significantly higher in SIV-infected than in SIV-uninfected AGMs (Fig. 9d). Since, in general, increases in the levels of D dimer are associated with those of sCD14 and IL-6, and because the groups were age biased (the infected monkeys included nearly all of the older monkeys, and the infants/juveniles were overrepresented in the uninfected group), we concluded that differences in the levels of D dimer may be age related rather than dependent on SIV infection status. Indeed, a correlation between age and levels of D dimer could be established, independent of SIV infection status (Fig. 10).
FIG 9.

Assessment of soluble biomarkers in wild AGMs from the Gambia. No significant increase in the markers associated with increased death in HIV-infected patients was observed in SIV-infected AGMs from the Gambia. (a) sCD14; (b) IL-6; (c) C-reactive protein. Significantly higher levels of D dimer were observed in SIV-infected AGMs, which were due to an age-related increase (d). Infant AGMs are illustrated as open circles and squares; juvenile AGMs are illustrated as gray circles and squares; adult AGMs are shown as black circles and squares. Samples collected from female AGMs are illustrated as circles (○); samples collected from male AGMs are shown as squares (□); the samples from acutely infected adult females (defined as having high viral loads and negative serologies) are illustrated as open circles with enhanced margins; the sample collected from an acutely infected juvenile female is illustrated as V; finally, two samples from juvenile monkeys with high viral loads but seropositive results (probably postacutely infected) are shown as half-empty symbols. P values were calculated by the Mann-Whitney U test.
FIG 10.
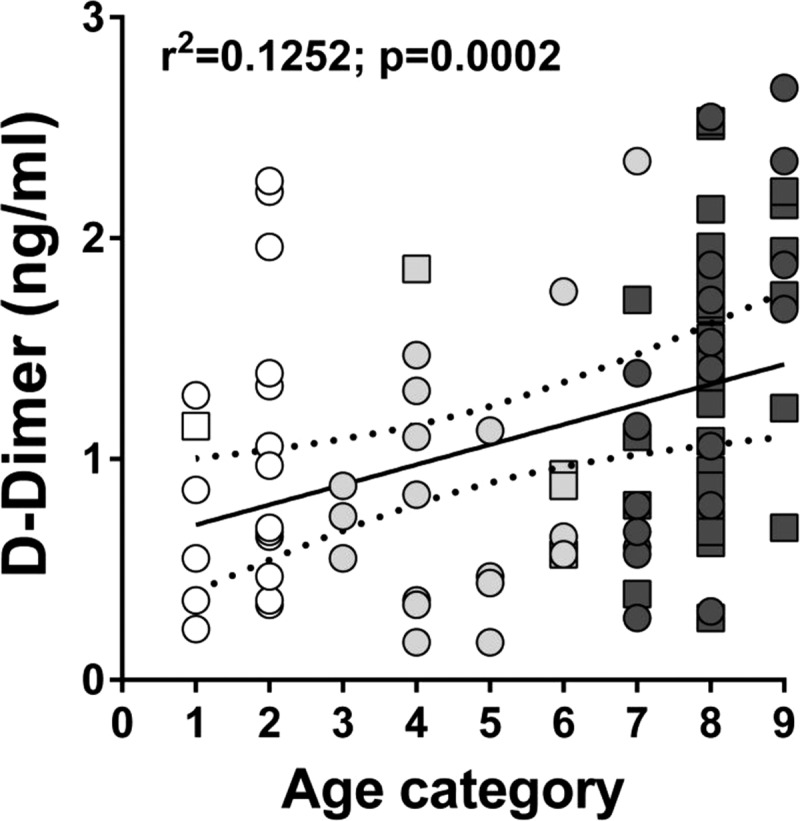
Age-related rather than SIV status-related increase in D dimer levels in wild AGMs from the Gambia. The linear regression line and 95% confidence intervals (dotted lines) are shown. The x axis represents the age category (established by dentition) of the animals; the y axis represents the levels of D dimer/ml of plasma. Infant AGMs are shown as open circles and squares; juvenile AGMs are illustrated as light gray circles and squares; adult AGMs are shown as dark gray circles and squares. Uninfected monkeys are illustrated as circles; infected monkeys are illustrated as squares.
DISCUSSION
In this study, we performed a large-scale analysis of the pathogenesis and transmission of SIVagmSab in its natural host, the sabaeus monkey, which is the AGM species endemic to West Africa. Using an algorithm that we recently optimized for the study of SIV pathogenesis in African NHP hosts in the wild (40), we characterized the natural history of SIVagmSab infection, with a special emphasis on aspects that may impact its natural transmission.
We have previously investigated the natural history of SIV in the wild in another AGM species that is endemic to southern Africa, the vervet monkey (40), and we outlined a strategy for a systematic investigation of the natural history of SIVs in wild natural hosts. Here, we performed a more detailed assessment of SIV pathogenicity in the wild by directly monitoring the impact of infection on immune cell populations and their degree of chronic immune activation. To this goal, PBMCs were isolated in the wild within hours from blood collection from a subset of AGMs, by employing protocols routinely utilized in the labs (17, 18, 32, 56). In spite of the challenging field conditions, when the PBMCs were thawed and stained for flow cytometry analyses, cell viability was >90%, which enabled us to perform immunophenotyping even on very rare cell populations, such as dendritic or natural killer cells. This analysis did not identify significant changes associated with SIVagmSab infection in any of the major immune cell populations. While these results are in agreement with previous reports showing that SIV infection does not have any major impact on immune cell populations in natural African NHP hosts (17, 18, 32, 88, 91–93), they should be interpreted with caution, because the SIV-infected and SIV-uninfected groups in our study were very unbalanced, with nearly all infants being SIV uninfected and most adult AGMs (and virtually all adult females) being SIV infected. As such, and considering that we and others have reported that the maturation of expression of immunophenotypic markers occurs in natural hosts of SIVs at adulthood (19, 34, 35, 90), the similar proportion of immunophenotypic expression may mask some discrete effects of SIV infection on immune cell populations. Furthermore, due to inherent limitation in the availability of clinical laboratories in the wild, we were not able to perform complete blood counts (CBCs) on the blood samples and therefore do not have absolute counts for these immune cell populations. Finally, while the major focus of the current pathogenesis studies is on tissues (such as lymph nodes and intestine) (94), we were not able to collect such tissues from wild animals, and thus our analyses are limited to the study of immune cell populations from circulation. Nevertheless, multiple studies in humans and Asian NHPs in which HIV/SIV infection invariably progresses to AIDS have pointed to a discernible impact of pathogenic HIV/SIV infections on multiple immune cell subsets (95, 96). Furthermore, in the few African NHP hosts that were reported to progress to AIDS (11), a significant impact of SIV infection on both CD4+ T cell counts and their levels of immune activation was documented to occur with progression to AIDS (97–99). Such an impact could not be identified in our study.
In addition to immunophenotypic correlates of disease progression, recent studies conducted in the SMART clinical trial identified biomarkers that are predictive of increased mortality and disease progression in HIV-1-infected patients (57, 58). Elevated levels of D dimer, IL-6, CRP, and sCD14 identified HIV-infected patients at high risk of death (57, 58). These results were recently confirmed in multiple NHP species (37). We therefore tested these biomarkers in our study group and compared those from SIV-infected to those from SIV-uninfected wild sabaeus monkeys. This analysis did not identify any differences in the levels of sCD14, IL-6, and CRP between SIV-infected and SIV-uninfected AGMs, suggesting that SIVagmSab infection of sabaeus monkeys is not associated with an increased risk of death in the wild. These results were in agreement with our previous results obtained in the wild in a different AGM species, the vervet monkeys (40). The only biomarker that was significantly increased in SIV-infected sabaeus monkeys was D dimer. However, increases in D dimer levels appeared to be dependent on age group rather than associated with SIV infection, as illustrated by a strong correlation between this biomarker and the age of the included monkeys, irrespective of their SIV infection status (Fig. 10). This is not surprising, as an age-related increase in coagulability that is reflected in higher levels of D dimer occurs in humans (100, 101). In our study, when levels of D dimer in infected and uninfected monkeys within the same age group were compared, no statistical significant differences could be identified (data not shown).
In addition to assessment of IL-6 levels between SIV-infected and SIV-uninfected sabaeus monkeys, we measured additional cytokines and chemokines in the two groups. The rationale is that chronic immune activation is one of the key factors driving the immune pathogenesis of AIDS. During pathogenic HIV/SIV infections, innate and adaptive antiviral immune responses contribute to chronic immune activation (85). In agreement with our previous reports (40), we could not identify significant differences in plasma levels of cytokines and chemokines, which strongly suggests that levels of immune activation and inflammation are normal in SIV-infected AGMs, thus supporting the benign outcome of SIV infection in sabaeus monkeys in the wild.
While these observations are of high significance and represent the first detailed assessment of major immune parameters in wild African NHPs, the novelty of our study relies on the fact that it is focused mainly on the characterization of natural SIV transmission in the wild. To date, the vast majority of studies in natural hosts of SIVs focused exclusively on understanding and identifying the correlates of the benign nature of SIV infection in these species (7, 8, 52). Such nearly exclusive focus on pathogenesis is not surprising given the desperate need to counterbalance the burden represented by the AIDS pandemic, especially in sub-Saharan Africa, and considering the paradox of natural SIV infections, which are characterized by a major disconnect between high levels of viral replication and lack of disease progression (7, 8, 20, 52). The extensive study of the correlates of lack of disease progression in natural hosts was a critical contributor to the paradigm shift in HIV pathogenesis studies from viral replication to chronic inflammation and immune activation as the critical drivers of disease progression (102).
The focus of this study on virus transmission in wild AGMs is justified by the second paradox of SIV infection in natural hosts: in these species, very efficient virus transmission to virtually the entire population of sexually active females occurs in the context of a massive restriction of target cells at mucosal sites (34–36, 71, 103), raising the question of whether or not the mucosal milieu is more permissive to SIV in these species than in humans or macaques. One may argue that this is indeed the case, given the fact that African NHP species coexisted with their species-specific viruses for many thousands of generations and that infection generally has no major deleterious consequences; in other words, there is no evolutionary pressure on virus transmission in the natural hosts of SIVs. Yet, there are several arguments against such an assumption. First, SIV infection retained its pathogenic potential in natural hosts, as demonstrated by the occurrence of a few cases of disease progression (11, 97–99, 104). Second, adaptations to avoid mother-to-infant transmissions clearly occurred in these species, resulting in low rates of transmission (19, 37, 40, 105). Third, the assumptions regarding the natural history of SIV infection in African NHPs relies only on studies performed in the three available models: AGMs, sooty mangabeys, and mandrills. In all these species, the levels of prevalence are high. Conversely, in several African NHP species (i.e., the great spotted nosed, mustached, or mona monkeys) (106, 107) for which no natural history data are yet available, the overall prevalence levels are low (1), raising the question of whether or not these low levels of prevalence are coincidental or due to increased pathogenicity of SIV infection in these species, which may limit it prevalence. This is not a trivial question, as viruses naturally infecting greater spotted nosed, mustached, and mona monkeys are the ancestors of HIV-1 (108). Altogether, these arguments demonstrate that the assumption that there is no evolutionary pressure on SIV transmission in natural hosts of SIVs is groundless.
In progressive, pathogenic hosts, HIV/SIV transmission is characterized by a massive genetic bottleneck, i.e., a substantial contraction of the genetic diversity of the quasispecies upon transmission and survival of only few lineages in the recipient. As a result, in the vast majority of transmissions, infection is initiated by a single genetic unit (50). It is considered that this genetic bottleneck is due to either a very low infectious dose at the time of transmission, a significant host barrier to new infections, or both (49, 50). The demonstration of the genetic bottleneck in natural hosts of SIVs upon natural transmission suggests that the same constraints of SIV transmission occur in the natural hosts of SIVs as in pathogenic infections. Since the levels of virus are generally higher in natural hosts than in HIV-1-infected patients, the low levels of target cells in the natural hosts might be responsible for the transmission of a very limited number of viral variants. Note, however, that to date, no study has investigated viral loads in vaginal secretions or sperm in natural hosts of SIVs.
Here, we assessed the genetic bottleneck of SIVagmSab transmission in wild sabaeus monkeys. Using the previously described algorithm for staging recently transmitted infections (i.e., seronegative monkeys with high viral loads [Fiebig stage II]) (82), we selected samples from acutely infected AGMs for the characterization of transmitted founder viruses and assessment of the number of virus variants that are naturally transmitted in the wild. The rationale for selecting acutely infected monkeys for inferring the transmitted founder variants is that, as previously reported, the virus diversifies in a relatively random fashion between transmission and peak viremia (Fiebig stage II), with no or little evidence of host-related selective pressures in this time frame (45, 47, 51, 109). The immune pressure exerted by the emerging immune responses results in strong strain selection and recombination (45, 47, 110), which prevent identification of the transmitted founder viruses at later times in infection. Since the analyzed fragment was relatively small, we confirmed the recent infection in the selected sabaeus monkeys by including in this analysis 2 chronically infected monkeys (i.e., seropositive monkeys with moderate levels of viral replication) as controls. The high genetic diversity identified in these chronic samples confirmed our assumption that the association between seronegativity and high VLs is indeed due to recent infection. We therefore assessed in these samples the number of transmitted founder variants and showed that SIV transmission in the wild is characterized by the same bottleneck as the one described in progressive HIV/SIV infections (49, 50, 111).
Our current study identified a rate of mother-to-infant transmission of less than 5% in sabaeus monkeys, which is in the range targeted by the World Health Organization as a “virtual elimination of HIV-1 maternal to infant transmission” (112) and very close to the levels recently reported for wild vervet monkeys (7%) (40) and captive sooty mangabeys (6.8%) (113). Such a low level of mother-to-infant transmission in natural hosts of SIVs suggests an active adaptive mechanism to prevent transmission to the offspring. As previously reported in our studies (19, 37), we identified this protective mechanism as relying on a delayed maturation of CCR5 expression on CD4+ T cells. Here, we were able to document in the wild and in the context of natural transmission that SIV infection occurred only in the infant and juveniles that expressed the highest levels of CCR5 on CD4+ T cells, in the range of those expressed by adult monkeys. Note, however, that SIVagmSab was reported to be able to use other coreceptors in vitro, such as CXCR4 and Bonzo (18). Yet, in our study the only correlate of SIV transmission was the expression of CCR5 on the CD4+ T cells, raising the question of the predictive value of the in vitro studies for in vivo coreceptor usage. Since it has been previously suggested that mutations in CCR5 could be associated with a resistance to SIV infection in AGM (114), future studies should focus on searching for mutations in CCR5 in the uninfected animals.
We have previously suggested that AGMs naturally infected with SIVs may represent a valuable model for the study of the correlates of resistance to HIV transmission in exposed seronegative (ESN) patients (40). For sabaeus monkeys in West Africa, we report prevalence levels in adult females that are even higher than those reported for AGMs in Southwest Africa. Yet, a small fraction of females remain uninfected, in spite of multiple lines of evidence supporting repeated exposure to SIV. These females are multiparous and, as such, have been exposed multiple times to the virus carried by all the dominant males. During mating season, there are multiple sexual encounters between females and the dominant male. In our study, all of the acute infections occurred in very young, primiparous females, suggesting that the susceptible females become infected very early during their sexual life, likely after the first mating season. This observation suggests that transmission processes in AGMs and humans share similar features. In humans, females are more susceptible than males during heterosexual transmission and the seroprevalence rates increase earlier in women already at the adolescent age (115). Women acquire HIV infection at least 5 to 7 years earlier than do men, and the prevalence of HIV infection is 3- to 7-fold higher among adolescent women than among adolescent men in sub-Saharan Africa (115).
Altogether, these observations strongly suggest that multiparous uninfected females are protected from infection. As such, our studies support the selection of exposed seronegative/exposed uninfected (ESN/EU) cases in wild populations of AGMs and call for studies to prospectively confirm resistance to infection in this subset of monkeys and identify the correlates of resistance to infection through a combination of genetic, immunologic, and virologic approaches. Defining the correlates of resistance to infection is instrumental in designing new therapeutic strategies aimed at preventing HIV infection.
The use of natural hosts in the wild to model HIV transmission and identify the correlates of effective control of SIV transmission would represent a major achievement in the field and contribute to reshaping current paradigms, as the study of correlates of lack of disease progression in natural hosts has shaped the current paradigms of HIV pathogenesis.
ACKNOWLEDGMENTS
We thank Beatrice Hahn, Vanessa Hirsch, Brandon Keele, and Preston Marx for helpful discussions. Samples used in this study were collected as a part of the Systems Biology Sample Repository. Sample collection was performed through the UCLA Systems Biology Sample Repository funded by NIH grants R01RR016300 and R01OD010980 to N.F. We thank the Department of Parks & Wildlife Management, Ministry of Forestry & the Environment, and Medical Research Council (MRC) The Gambia Unit for enabling our sample collection from free-ranging monkeys. We thank MRC The Gambia Unit for making laboratory, equipment, and cold-storage space available for our project and all the staff of the MRC that helped with organizing field sample collection and providing administrative support and transportation, in particular, Sanneh Mamkumba for administrative help, Ousman Secka for help with supplies and sample storage and shipment, and drivers Ousman Bah and Lamin Gibba. We gratefully acknowledge the expertise and assistance of Oliver (Pess) Morton, Ebou Jarjou, and Katherine Camfield during the field work as well as Ben Kigbu and Toye Adegboye for veterinary care.
Virology investigations were funded through NIH/NIAID/NCRR/NIDCR grants R01 RR025781 (C.A. and I.P.), P01 AI088564 (C.A.), and R56 DE023508 (C.A.). Coagulation and inflammation markers were tested through RO1 HL117715 (I.P.). F.F. is funded by a BBSRC studentship to D.L.R.
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Footnotes
Published ahead of print 12 March 2014
REFERENCES
- 1.Locatelli S, Peeters M. 2012. Cross-species transmission of simian retroviruses: how and why they could lead to the emergence of new diseases in the human population. AIDS 26:659–673. 10.1097/QAD.0b013e328350fb68 [DOI] [PubMed] [Google Scholar]
- 2.VandeWoude S, Apetrei C. 2006. Going wild: lessons from T-lymphotropic naturally occurring lentiviruses. Clin. Microbiol. Rev. 19:728–762. 10.1128/CMR.00009-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bibollet-Ruche F, Bailes E, Gao F, Pourrut X, Barlow KL, Clewley J, Mwenda JM, Langat DK, Chege GK, McClure HM, Mpoudi-Ngole E, Delaporte E, Peeters M, Shaw GM, Sharp PM, Hahn BH. 2004. A new simian immunodeficiency virus lineage (SIVdeb) infecting de Brazza's monkeys (Cercopithecus neglectus): evidence for a Cercopithecus monkey virus clade. J. Virol. 78:7748–7762. 10.1128/JVI.78.14.7748-7762.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hahn BH, Shaw GM, De Cock KM, Sharp PM. 2000. AIDS as a zoonosis: scientific and public health implications. Science 287:607–614. 10.1126/science.287.5453.607 [DOI] [PubMed] [Google Scholar]
- 5.Plantier JC, Leoz M, Dickerson JE, De Oliveira F, Cordonnier F, Lemee V, Damond F, Robertson DL, Simon F. 2009. A new human immunodeficiency virus derived from gorillas. Nat. Med. 15:871–872. 10.1038/nm.2016 [DOI] [PubMed] [Google Scholar]
- 6.Damond F, Worobey M, Campa P, Farfara I, Colin G, Matheron S, Brun-Vézinet F, Robertson DL, Simon F. 2004. Identification of a highly divergent HIV-2 and proposal for a change in HIV-2 classification. AIDS Res. Hum. Retroviruses 20:666–672. 10.1089/0889222041217392 [DOI] [PubMed] [Google Scholar]
- 7.Pandrea I, Apetrei C. 2010. Where the wild things are: pathogenesis of SIV infection in African nonhuman primate hosts. Curr. HIV/AIDS Rep. 7:28–36. 10.1007/s11904-009-0034-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pandrea I, Sodora DL, Silvestri G, Apetrei C. 2008. Into the wild: simian immunodeficiency virus (SIV) infection in natural hosts. Trends Immunol. 29:419–428. 10.1016/j.it.2008.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brenchley JM, Paiardini M. 2011. Immunodeficiency lentiviral infections in natural and non-natural hosts. Blood 118:847–854. 10.1182/blood-2010-12-325936 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Brenchley JM, Silvestri G, Douek DC. 2010. Nonprogressive and progressive primate immunodeficiency lentivirus infections. Immunity 32:737–742. 10.1016/j.immuni.2010.06.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pandrea I, Silvestri G, Apetrei C. 2009. AIDS in African nonhuman primate hosts of SIVs: a new paradigm of SIV infection. Curr. HIV Res. 6:57–72. 10.2174/157016209787048456 [DOI] [PubMed] [Google Scholar]
- 12.Apetrei C, Gautam R, Sumpter B, Carter AC, Gaufin T, Staprans SI, Else J, Barnes M, Cao R, Jr, Garg S, Milush JM, Sodora DL, Pandrea I, Silvestri G. 2007. Virus-subtype specific features of natural SIVsmm infection in sooty mangabeys. J. Virol. 81:7913–7923. 10.1128/JVI.00281-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Apetrei C, Sumpter B, Souquiere S, Chahroudi A, Makuwa M, Reed P, Ribeiro RM, Pandrea I, Roques P, Silvestri G. 2011. Immunovirological analyses of chronically simian immunodeficiency virus SIVmnd-1- and SIVmnd-2-infected mandrills (Mandrillus sphinx). J. Virol. 85:13077–13087. 10.1128/JVI.05693-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Goldstein S, Ourmanov I, Brown CR, Beer BE, Elkins WR, Plishka R, Buckler-White A, Hirsch VM. 2000. Wide range of viral load in healthy African green monkeys naturally infected with simian immunodeficiency virus. J. Virol. 74:11744–11753. 10.1128/JVI.74.24.11744-11753.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rey-Cuille MA, Berthier JL, Bomsel-Demontoy MC, Chaduc Y, Montagnier L, Hovanessian AG, Chakrabarti LA. 1998. Simian immunodeficiency virus replicates to high levels in sooty mangabeys without inducing disease. J. Virol. 72:3872–3886 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chakrabarti LA, Lewin SR, Zhang L, Gettie A, Luckay A, Martin LN, Skulsky E, Ho DD, Cheng-Mayer C, Marx PA. 2000. Normal T-cell turnover in sooty mangabeys harboring active simian immunodeficiency virus infection. J. Virol. 74:1209–1223. 10.1128/JVI.74.3.1209-1223.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Pandrea I, Apetrei C, Dufour J, Dillon N, Barbercheck J, Metzger M, Jacquelin B, Bohm R, Marx PA, Barre-Sinoussi F, Hirsch VM, Muller-Trutwin MC, Lackner AA, Veazey R. 2006. Simian immunodeficiency virus (SIV) SIVagm.sab infection of Caribbean African green monkeys: new model of the study of SIV pathogenesis in natural hosts J. Virol. 80:4858–4867. 10.1128/JVI.80.10.4858-4867.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pandrea I, Kornfeld C, Ploquin MJ-I, Apetrei C, Faye A, Rouquet P, Roques P, Simon F, Barré-Sinoussi F, Müller-Trutwin MC, Diop OM. 2005. Impact of viral factors on very early in vivo replication profiles in SIVagm-infected African green monkeys. J. Virol. 79:6249–6259. 10.1128/JVI.79.10.6249-6259.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pandrea I, Onanga R, Souquiere S, Mouinga-Ondéme A, Bourry O, Makuwa M, Rouquet P, Silvestri G, Simon F, Roques P, Apetrei C. 2008. Paucity of CD4+CCR5+ T-cells may prevent breastfeeding transmission of SIV in natural non-human primate hosts J. Virol. 82:5501–5509. 10.1128/JVI.02555-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pandrea I, Silvestri G, Onanga R, Veazey RS, Marx PA, Hirsch VM, Apetrei C. 2006. Simian immunodeficiency viruses replication dynamics in African non-human primate hosts: common patterns and species-specific differences. J. Med. Primatol. 35:194–201. 10.1111/j.1600-0684.2006.00168.x [DOI] [PubMed] [Google Scholar]
- 21.Pandrea I, Onanga R, Kornfeld C, Rouquet P, Bourry O, Clifford S, Telfer PT, Abernethy K, White LT, Ngari P, Muller-Trutwin M, Roques P, Marx PA, Simon F, Apetrei C. 2003. High levels of SIVmnd-1 replication in chronically infected Mandrillus sphinx. Virology 317:119–127. 10.1016/j.virol.2003.08.015 [DOI] [PubMed] [Google Scholar]
- 22.Broussard SR, Staprans SI, White R, Whitehead EM, Feinberg MB, Allan JS. 2001. Simian immunodeficiency virus replicates to high levels in naturally infected African green monkeys without inducing immunologic or neurologic disease. J. Virol. 75:2262–2275. 10.1128/JVI.75.5.2262-2275.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Silvestri G, Sodora DL, Koup RA, Paiardini M, O'Neil SP, McClure HM, Staprans SI, Feinberg MB. 2003. Nonpathogenic SIV infection of sooty mangabeys is characterized by limited bystander immunopathology despite chronic high-level viremia. Immunity 18:441–452. 10.1016/S1074-7613(03)00060-8 [DOI] [PubMed] [Google Scholar]
- 24.Gaufin T, Pattison M, Gautam R, Stoulig C, Dufour J, MacFarland J, Mandell D, Tatum C, Marx M, Ribeiro RM, Montefiori C, Apetrei C, Pandrea I. 2009. Effect of B cell depletion on viral replication and clinical outcome of SIV infection in a natural host. J. Virol. 83:10347–10357. 10.1128/JVI.00880-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gaufin T, Ribeiro RM, Gautam R, Dufour J, Mandell D, Apetrei C, Pandrea I. 2010. Experimental depletion of CD8+ cells in acutely SIVagm-infected African Green Monkeys results in increased viral replication. Retrovirology 7:42. 10.1186/1742-4690-7-42 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zahn RC, Rett MD, Korioth-Schmitz B, Sun Y, Buzby AP, Goldstein S, Brown CR, Byrum RA, Freeman GJ, Letvin NL, Hirsch VM, Schmitz JE. 2008. Simian immunodeficiency virus (SIV)-specific CD8+ T cell responses in chronically SIVagm-infected vervet African green monkeys. J. Virol. 82:11577–11588. 10.1128/JVI.01779-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zahn RC, Rett MD, Li M, Tang H, Korioth-Schmitz B, Balachandran H, White R, Pryputniewicz S, Letvin NL, Kaur A, Montefiori DC, Carville A, Hirsch VM, Allan JS, Schmitz JE. 2010. Suppression of adaptive immune responses during primary SIV infection of sabaeus African green monkeys delays partial containment of viremia but does not induce disease. Blood 115:3070–3078. 10.1182/blood-2009-10-245225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Schmitz JE, Zahn RC, Brown CR, Rett MD, Li M, Tang H, Pryputniewicz S, Byrum RA, Kaur A, Montefiori DC, Allan JS, Goldstein S, Hirsch VM. 2009. Inhibition of adaptive immune responses leads to a fatal clinical outcome in SIV-infected pigtailed macaques but not vervet African green monkeys. PLoS Pathog. 5:e1000691. 10.1371/journal.ppat.1000691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hirsch VM. 2004. What can natural infection of African monkeys with simian immunodeficiency virus tell us about the pathogenesis of AIDS? AIDS Rev. 6:40–53 [PubMed] [Google Scholar]
- 30.Gicheru MM, Otsyula M, Spearman P, Graham BS, Miller CJ, Robinson HL, Haigwood NL, Montefiori DC. 1999. Neutralizing antibody responses in African green monkeys naturally infected with simian immunodeficiency virus (SIVagm). J. Med. Primatol. 28:97–104. 10.1111/j.1600-0684.1999.tb00257.x [DOI] [PubMed] [Google Scholar]
- 31.Estaquier J, Idziorek T, de Bels F, Barre-Sinoussi F, Hurtrel B, Aubertin AM, Venet A, Mehtali M, Muchmore E, Michel P, Mouton Y, Girard M, Ameisen JC. 1994. Programmed cell death and AIDS: significance of T-cell apoptosis in pathogenic and nonpathogenic primate lentiviral infections. Proc. Natl. Acad. Sci. U. S. A. 91:9431–9435. 10.1073/pnas.91.20.9431 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Pandrea I, Gautam R, Ribeiro R, Brenchley JM, Butler IF, Pattison M, Rasmussen T, Marx PA, Silvestri G, Lackner AA, Perelson AS, Douek DC, Veazey RS, Apetrei C. 2007. Acute loss of intestinal CD4+ T cells is not predictive of SIV virulence. J. Immunol. 179:3035–3046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Cumont MC, Diop O, Vaslin B, Elbim C, Viollet L, Monceaux V, Lay S, Silvestri G, Le Grand R, Muller-Trutwin M, Hurtrel B, Estaquier J. 2008. Early divergence in lymphoid tissue apoptosis between pathogenic and nonpathogenic simian immunodeficiency virus infections of nonhuman primates. J. Virol. 82:1175–1184. 10.1128/JVI.00450-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Vinton C, Klatt NR, Harris LD, Briant JA, Sanders-Beer BE, Herbert R, Woodward R, Silvestri G, Pandrea I, Apetrei C, Hirsch VM, Brenchley JM. 2011. CD4-like immunological function by CD4− T cells in multiple natural hosts of simian immunodeficiency virus. J. Virol. 85:8702–8708. 10.1128/JVI.00332-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Beaumier CM, Harris LD, Goldstein S, Klatt NR, Whitted S, McGinty J, Apetrei C, Pandrea I, Hirsch VM, Brenchley JM. 2009. CD4 downregulation by memory CD4+ T cells in vivo renders African green monkeys resistant to progressive SIVagm infection. Nat. Med. 15:879–885. 10.1038/nm.1970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Pandrea I, Apetrei C, Gordon S, Barbercheck J, Dufour J, Bohm R, Sumpter B, Roques P, Marx PA, Hirsch VM, Kaur A, Lackner AA, Veazey RS, Silvestri G. 2007. Paucity of CD4+CCR5+ T cells is a typical feature of natural SIV hosts. Blood 109:1069–1076. 10.1182/blood-2006-05-024364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pandrea I, Parrish NF, Raehtz K, Gaufin T, Barbian HJ, Ma D, Kristoff J, Gautam R, Zhong F, Haret-Richter GS, Trichel A, Shaw GM, Hahn BH, Apetrei C. 2012. Mucosal simian immunodeficiency virus transmission in African green monkeys: susceptibility to infection is proportional to target cell availability at mucosal sites. J. Virol. 86:4158–4168. 10.1128/JVI.07141-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jolly CJ, Phillips-Conroy JE, Turner TR, Broussard S, Allan JS. 1996. SIVagm incidence over two decades in a natural population of Ethiopian grivet monkeys (Cercopithecus aethiops aethiops). J. Med. Primatol. 25:78–83. 10.1111/j.1600-0684.1996.tb00198.x [DOI] [PubMed] [Google Scholar]
- 39.Phillips-Conroy JE, Jolly CJ, Petros B, Allan JS, Desrosiers RC. 1994. Sexual transmission of SIVagm in wild grivet monkeys. J. Med. Primatol. 23:1–7. 10.1111/j.1600-0684.1994.tb00088.x [DOI] [PubMed] [Google Scholar]
- 40.Ma D, Jasinska A, Kristoff J, Grobler JP, Turner T, Jung Y, Schmitt C, Raehtz K, Feyertag F, Martinez Sosa N, Wijewardana V, Burke DS, Robertson DL, Tracy R, Pandrea I, Freimer N, Apetrei C, International Vervet Research Consortium 2013. SIVagm infection in wild African green monkeys from South Africa: epidemiology, natural history, and evolutionary considerations. PLoS Pathog. 9:e1003011. 10.1371/journal.ppat.1003011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Durand JP, Tuppin P, Maison P, Galat G, Galat-Luong A, Jeannel D, De The G. 1995. Increased risk for a second retroviral infection (SIV or STLV type I) for wild African green monkeys already infected by one retrovirus in Senegal (West Africa). AIDS Res. Hum. Retroviruses 11:985–988. 10.1089/aid.1995.11.985 [DOI] [PubMed] [Google Scholar]
- 42.Muller MC, Saksena NK, Nerrienet E, Chappey C, Herve VM, Durand JP, Legal-Campodonico P, Lang MC, Digoutte JP, Georges AJ. 1993. Simian immunodeficiency viruses from central and western Africa: evidence for a new species-specific lentivirus in tantalus monkeys. J. Virol. 67:1227–1235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nerrienet E, Amouretti X, Muller-Trutwin MC, Poaty-Mavoungou V, Bedjebaga I, Nguyen HT, Dubreuil G, Corbet S, Wickings EJ, Barre-Sinoussi F, Georges AJ, Georges-Courbot MC. 1998. Phylogenetic analysis of SIV and STLV type I in mandrills (Mandrillus sphinx): indications that intracolony transmissions are predominantly the result of male-to-male aggressive contacts. AIDS Res. Hum. Retroviruses 14:785–796. 10.1089/aid.1998.14.785 [DOI] [PubMed] [Google Scholar]
- 44.Mellors JW, Rinaldo CR, Jr, Gupta P, White RM, Todd JA, Kingsley LA. 1996. Prognosis in HIV-1 infection predicted by the quantity of virus in plasma. Science 272:1167–1170. 10.1126/science.272.5265.1167 [DOI] [PubMed] [Google Scholar]
- 45.Keele BF, Giorgi EE, Salazar-Gonzalez JF, Decker JM, Pham KT, Salazar MG, Sun C, Grayson T, Wang S, Li H, Wei X, Jiang C, Kirchherr JL, Gao F, Anderson JA, Ping LH, Swanstrom R, Tomaras GD, Blattner WA, Goepfert PA, Kilby JM, Saag MS, Delwart EL, Busch MP, Cohen MS, Montefiori DC, Haynes BF, Gaschen B, Athreya GS, Lee HY, Wood N, Seoighe C, Perelson AS, Bhattacharya T, Korber BT, Hahn BH, Shaw GM. 2008. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. U. S. A. 105:7552–7557. 10.1073/pnas.0802203105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Keele BF, Li H, Learn GH, Hraber P, Giorgi EE, Grayson T, Sun C, Chen Y, Yeh WW, Letvin NL, Nabel GJ, Haynes BF, Bhattacharya T, Perelson AS, Korber BT, Hahn BH, Shaw GM. 2009. Low dose rectal inoculation of rhesus macaques by SIVsmE660 or SIVmac251 recapitulates human mucosal infection by HIV-1. J. Exp. Med. 206:1117–1134. 10.1084/jem.20082831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Salazar-Gonzalez JF, Salazar MG, Keele BF, Learn GH, Giorgi EE, Li H, Decker JM, Wang S, Baalwa J, Kraus MH, Parrish NF, Shaw KS, Guffey MB, Bar KJ, Davis KL, Ochsenbauer-Jambor C, Kappes JC, Saag MS, Cohen MS, Mulenga J, Derdeyn CA, Allen S, Hunter E, Markowitz M, Hraber P, Perelson AS, Bhattacharya T, Haynes BF, Korber BT, Hahn BH, Shaw GM. 2009. Genetic identity, biological phenotype, and evolutionary pathways of transmitted/founder viruses in acute and early HIV-1 infection. J. Exp. Med. 206:1273–1289. 10.1084/jem.20090378 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Abrahams MR, Anderson JA, Giorgi EE, Seoighe C, Mlisana K, Ping LH, Athreya GS, Treurnicht FK, Keele BF, Wood N, Salazar-Gonzalez JF, Bhattacharya T, Chu H, Hoffman I, Galvin S, Mapanje C, Kazembe P, Thebus R, Fiscus S, Hide W, Cohen MS, Karim SA, Haynes BF, Shaw GM, Hahn BH, Korber BT, Swanstrom R, Williamson C, Team CAIS, Center for HIVAVIC 2009. Quantitating the multiplicity of infection with human immunodeficiency virus type 1 subtype C reveals a non-Poisson distribution of transmitted variants. J. Virol. 83:3556–3567. 10.1128/JVI.02132-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Keele BF, Derdeyn CA. 2009. Genetic and antigenic features of the transmitted virus. Curr. Opin. HIV AIDS 4:352–357. 10.1097/COH.0b013e32832d9fef [DOI] [PubMed] [Google Scholar]
- 50.Fennessey CM, Keele BF. 2013. Using nonhuman primates to model HIV transmission. Curr. Opin. HIV AIDS 8:280–287. 10.1097/COH.0b013e328361cfff [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Parrish NF, Gao F, Li H, Giorgi EE, Barbian HJ, Parrish EH, Zajic L, Iyer SS, Decker JM, Kumar A, Hora B, Berg A, Cai F, Hopper J, Denny TN, Ding H, Ochsenbauer C, Kappes JC, Galimidi RP, West AP, Jr, Bjorkman PJ, Wilen CB, Doms RW, O'Brien M, Bhardwaj N, Borrow P, Haynes BF, Muldoon M, Theiler JP, Korber B, Shaw GM, Hahn BH. 2013. Phenotypic properties of transmitted founder HIV-1. Proc. Natl. Acad. Sci. U. S. A. 110:6626–6633. 10.1073/pnas.1304288110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sodora DL, Allan J, Apetrei C, Brenchley JM, Douek DC, Else J, Estes J, Hahn BH, Hirsch VM, Kaur A, Kirchhoff F, Muller-Trutwin M, Pandrea I, Schmitz JE, Silvestri G. 2009. Towards an AIDS vaccine: lessons from natural SIV infections of African nonhuman primate hosts. Nat. Med. 15:861–865. 10.1038/nm.2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Jasinska AJ, Schmitt CA, Service SK, Cantor RM, Dewar K, Jentsch JD, Kaplan JR, Turner TR, Warren WC, Weinstock GM, Woods RP, Freimer NB. 2013. Systems biology of the vervet monkey. ILAR J. 54:122–143. 10.1093/ilar/ilt049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Turner TR, Anapol F, Jolly CJ. 1997. Growth, development, and sexual dimorphism in vervet monkeys (Cercopithecus aethiops) at four sites in Kenya. Am. J. Phys. Anthropol. 103:19–35 [DOI] [PubMed] [Google Scholar]
- 55.Grobler P, Turner T. 2010. A novel trap design for the capture and sedation of vervet monkeys (Chlorocebus aethiops). South Afr. J. Wildlife Res. 40:163–168. 10.3957/056.040.0208 [DOI] [Google Scholar]
- 56.Gautam R, Gaufin T, Butler I, Gautam A, Barnes M, Mandell D, Pattison M, Tatum C, Macfarland J, Monjure C, Marx PA, Pandrea I, Apetrei C. 2009. SIVrcm, a unique CCR2-tropic virus, selectively depletes memory CD4+ T cells in pigtailed macaques through rapid coreceptor expansion in vivo. J. Virol. 83:7894–7908. 10.1128/JVI.00444-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kuller LH, Tracy R, Belloso W, De Wit S, Drummond F, Lane HC, Ledergerber B, Lundgren J, Neuhaus J, Nixon D, Paton NI, Neaton JD. 2008. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 5:e203. 10.1371/journal.pmed.0050203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sandler NG, Wand H, Roque A, Law M, Nason MC, Nixon DE, Pedersen C, Ruxrungtham K, Lewin SR, Emery S, Neaton JD, Brenchley JM, Deeks SG, Sereti I, Douek DC, INSIGHT SMART Study Group 2011. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J. Infect. Dis. 203:780–790. 10.1093/infdis/jiq118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Deeks SG, Tracy R, Douek DC. 2013. Systemic effects of inflammation on health during chronic HIV infection. Immunity 39:633–645. 10.1016/j.immuni.2013.10.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Courgnaud V, Pourrut X, Bibollet-Ruche F, Mpoudi-Ngole E, Bourgeois A, Delaporte E, Peeters M. 2001. Characterization of a novel simian immunodeficiency virus from guereza colobus monkeys (Colobus guereza) in Cameroon: a new lineage in the nonhuman primate lentivirus family. J. Virol. 75:857–866. 10.1128/JVI.75.2.857-866.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Gao F, Yue L, Robertson DL, Hill SC, Hui H, Biggar RJ, Neequaye AE, Whelan TM, Ho DD, Shaw GM, et al. 1994. Genetic diversity of human immunodeficiency virus type 2: evidence for distinct sequence subtypes with differences in virus biology. J. Virol. 68:7433–7447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Jin MJ, Rogers J, Phillips-Conroy JE, Allan JS, Desrosiers RC, Shaw GM, Sharp PM, Hahn BH. 1994. Infection of a yellow baboon with simian immunodeficiency virus from African green monkeys: evidence for cross-species transmission in the wild. J. Virol. 68:8454–8460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gnanadurai CW, Pandrea I, Parrish NF, Kraus MH, Learn GH, Salazar MG, Gautam R, Apetrei C, Hahn BH, Kirchhoff F. 2010. Genetic identity and biological phenotype of a transmitted/founder virus representative of non-pathogenic simian immunodeficiency virus infection in African green monkeys. J. Virol. 84:12245–12254. 10.1128/JVI.01603-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Waterhouse AM, Procter JB, Martin DM, Clamp M, Barton GJ. 2009. Jalview Version 2—a multiple sequence alignment editor and analysis workbench. Bioinformatics 25:1189–1191. 10.1093/bioinformatics/btp033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28:2731–2739. 10.1093/molbev/msr121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Maydt J, Lengauer T. 2006. Recco: recombination analysis using cost optimization. Bioinformatics 22:1064–1071. 10.1093/bioinformatics/btl057 [DOI] [PubMed] [Google Scholar]
- 67.Kosakovsky Pond SL, Posada D, Gravenor MB, Woelk CH, Frost SD. 2006. Automated phylogenetic detection of recombination using a genetic algorithm. Mol. Biol. Evol. 23:1891–1901. 10.1093/molbev/msl051 [DOI] [PubMed] [Google Scholar]
- 68.Huson DH, Bryant D. 2006. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 23:254–267. 10.1093/molbev/msj030 [DOI] [PubMed] [Google Scholar]
- 69.Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22:2688–2690. 10.1093/bioinformatics/btl446 [DOI] [PubMed] [Google Scholar]
- 70.Pandrea I, Gaufin T, Gautam R, Kristoff J, Mandell D, Montefiori D, Keele BF, Ribeiro RM, Veazey RS, Apetrei C. 2011. Functional cure of SIVagm infection in rhesus macaques results in complete recovery of CD4+ T cells and is reverted by CD8+ cell depletion. PLoS Pathog. 7:e1002170. 10.1371/journal.ppat.1002170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Apetrei C, Gaufin T, Gautam R, Vinton C, Hirsch VM, Lewis M, Brenchley JM, Pandrea I. 2010. Pattern of SIVagm infection in patas monkeys suggests that host adaptation to SIV infection may result in resistance to infection and virus extinction. J. Infect. Dis. 202(Suppl 3):S371–S376. 10.1086/655970 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pandrea I, Ribeiro RM, Gautam R, Gaufin T, Pattison M, Barnes M, Monjure C, Stoulig C, Silvestri G, Miller M, Perelson AS, Apetrei C. 2008. Simian immunodeficiency virus SIVagm dynamics in African green monkeys. J. Virol. 82:3713–3724. 10.1128/JVI.02402-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Simon F, Souquiere S, Damond F, Kfutwah A, Makuwa M, Leroy E, Rouquet P, Berthier JL, Rigoulet J, Lecu A, Telfer PT, Pandrea I, Plantier JC, Barre-Sinoussi F, Roques P, Muller-Trutwin MC, Apetrei C. 2001. Synthetic peptide strategy for the detection of and discrimination among highly divergent primate lentiviruses. AIDS Res. Hum. Retroviruses 17:937–952. 10.1089/088922201750290050 [DOI] [PubMed] [Google Scholar]
- 74.Brenchley JM, Price DA, Schacker TW, Asher TE, Silvestri G, Rao S, Kazzaz Z, Lambotte O, Altmann D, Blazar BR, Rodriguez B, Teixeira-Johnson L, Landay A, Martin JN, Hecht FM, Picker LJ, Lederman M, Deeks SG, Douek DC. 2006. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 12:1365–1371. 10.1038/nm1511 [DOI] [PubMed] [Google Scholar]
- 75.Brenchley JM, Price DA, Douek DC. 2006. HIV disease: fallout from a mucosal catastrophe? Nat. Immunol. 7:235–239. 10.1038/ni1316 [DOI] [PubMed] [Google Scholar]
- 76.Pandrea I, Cornell E, Wilson C, Ribeiro RM, Ma D, Kristoff J, Xu C, Haret-Richter GS, Trichel A, Apetrei C, Landay A, Tracy R. 2012. Coagulation biomarkers predict disease progression in SIV-infected nonhuman primates. Blood 120:1357–1366. 10.1182/blood-2012-03-414706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Pandrea I, Gaufin T, Brenchley JM, Gautam R, Monjure C, Gautam A, Coleman C, Lackner AA, Ribeiro R, Douek DC, Apetrei C. 2008. Experimentally-induced immune activation in natural hosts of SIV induces significant increases in viral replication and CD4+ T cell depletion. J. Immunol. 181:6687–6691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Allan JS, Short M, Taylor ME, Su S, Hirsch VM, Johnson PR, Shaw GM, Hahn BH. 1991. Species-specific diversity among simian immunodeficiency viruses from African green monkeys. J. Virol. 65:2816–2828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Otsyula MG, Gettie A, Suleman M, Tarara R, Mohamed I, Marx P. 1995. Apparent lack of vertical transmission of simian immunodeficiency virus (SIV) in naturally infected African green monkeys, Cercopithecus aethiops. Ann. Trop. Med. Parasitol. 89:573–576 [DOI] [PubMed] [Google Scholar]
- 80.Fultz PN, Gordon TP, Anderson DC, McClure HM. 1990. Prevalence of natural infection with simian immunodeficiency virus and simian T-cell leukemia virus type I in a breeding colony of sooty mangabey monkeys. AIDS 4:619–625. 10.1097/00002030-199007000-00002 [DOI] [PubMed] [Google Scholar]
- 81.Jin MJ, Hui H, Robertson DL, Muller MC, Barre-Sinoussi F, Hirsch VM, Allan JS, Shaw GM, Sharp PM, Hahn BH. 1994. Mosaic genome structure of simian immunodeficiency virus from west African green monkeys. EMBO J. 13:2935–2947 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Fiebig EW, Wright DJ, Rawal BD, Garrett PE, Schumacher RT, Peddada L, Heldebrant C, Smith R, Conrad A, Kleinman SH, Busch MP. 2003. Dynamics of HIV viremia and antibody seroconversion in plasma donors: implications for diagnosis and staging of primary HIV infection. AIDS 17:1871–1879. 10.1097/00002030-200309050-00005 [DOI] [PubMed] [Google Scholar]
- 83.Liu J, Keele BF, Li H, Keating S, Norris PJ, Carville A, Mansfield KG, Tomaras GD, Haynes BF, Kolodkin-Gal D, Letvin NL, Hahn BH, Shaw GM, Barouch DH. 2010. Low-dose mucosal simian immunodeficiency virus infection restricts early replication kinetics and transmitted virus variants in rhesus monkeys. J. Virol. 84:10406–10412 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sumpter B, Dunham R, Gordon S, Engram J, Hennessy M, Kinter A, Paiardini M, Cervasi B, Klatt N, McClure H, Milush JM, Staprans S, Sodora DL, Silvestri G. 2007. Correlates of preserved CD4+ T cell homeostasis during natural, nonpathogenic simian immunodeficiency virus infection of sooty mangabeys: implications for AIDS pathogenesis. J. Immunol. 178:1680–1691 [DOI] [PubMed] [Google Scholar]
- 85.Sodora DL, Silvestri G. 2008. Immune activation and AIDS pathogenesis. AIDS 22:439–446. 10.1097/QAD.0b013e3282f2dbe7 [DOI] [PubMed] [Google Scholar]
- 86.Sachsenberg N, Perelson AS, Yerly S, Schockmel GA, Leduc D, Hirschel B, Perrin L. 1998. Turnover of CD4+ and CD8+ T lymphocytes in HIV-1 infection as measured by Ki-67 antigen. J. Exp. Med. 187:1295–1303. 10.1084/jem.187.8.1295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kornfeld C, Ploquin MJ, Pandrea I, Faye A, Onanga R, Apetrei C, Poaty-Mavoungou V, Rouquet P, Estaquier J, Mortara L, Desoutter JF, Butor C, Le Grand R, Roques P, Simon F, Barre-Sinoussi F, Diop OM, Muller-Trutwin MC. 2005. Antiinflammatory profiles during primary SIV infection in African green monkeys are associated with protection against AIDS. J. Clin. Invest. 115:1082–1091. 10.1172/JCI23006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Onanga R, Souquiere S, Makuwa M, Mouinga-Ondeme A, Simon F, Apetrei C, Roques P. 2006. Primary simian immunodeficiency virus SIVmnd-2 infection in mandrills (Mandrillus sphinx). J. Virol. 80:3303–3309. 10.1128/JVI.80.7.3301-3309.2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Silvestri G, Paiardini M, Pandrea I, Lederman MM, Sodora DL. 2007. Understanding the benign nature of SIV infection in natural hosts. J. Clin. Invest. 117:3148–3154. 10.1172/JCI33034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Chakrabarti LA, Lewin SR, Zhang L, Gettie A, Luckay A, Martin LN, Skulsky E, Ho DD, Cheng-Mayer C, Marx PA. 2000. Age-dependent changes in T cell homeostasis and SIV load in sooty mangabeys. J. Med. Primatol. 29:158–165. 10.1034/j.1600-0684.2000.290309.x [DOI] [PubMed] [Google Scholar]
- 91.Brenchley JM, Paiardini M, Knox KS, Asher AI, Cervasi B, Asher TE, Scheinberg P, Price DA, Hage CA, Kholi LM, Khoruts A, Frank I, Else J, Schacker T, Silvestri G, Douek DC. 2008. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood 112:2826–2835. 10.1182/blood-2008-05-159301 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Favre D, Lederer S, Kanwar B, Ma ZM, Proll S, Kasakow Z, Mold J, Swainson L, Barbour JD, Baskin CR, Palermo R, Pandrea I, Miller CJ, Katze MG, McCune JM. 2009. Critical loss of the balance between Th17 and T regulatory cell populations in pathogenic SIV infection. PLoS Pathog. 5:e1000295. 10.1371/journal.ppat.1000295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Onanga R, Kornfeld C, Pandrea I, Estaquier J, Souquiere S, Rouquet P, Mavoungou VP, Bourry O, M'Boup S, Barre-Sinoussi F, Simon F, Apetrei C, Roques P, Muller-Trutwin MC. 2002. High levels of viral replication contrast with only transient changes in CD4+ and CD8+ cell numbers during the early phase of experimental infection with simian immunodeficiency virus SIVmnd-1 in Mandrillus sphinx. J. Virol. 76:10256–10263. 10.1128/JVI.76.20.10256-10263.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Veazey RS, Lackner AA. 2004. Getting to the guts of HIV pathogenesis. J. Exp. Med. 200:697–700. 10.1084/jem.20041464 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Mellors JW, Munoz A, Giorgi JV, Margolick JB, Tassoni CJ, Gupta P, Kingsley LA, Todd JA, Saah AJ, Detels R, Phair JP, Rinaldo CR., Jr 1997. Plasma viral load and CD4+ lymphocytes as prognostic markers of HIV-1 infection. Ann. Intern. Med. 126:946–954. 10.7326/0003-4819-126-12-199706150-00003 [DOI] [PubMed] [Google Scholar]
- 96.Hirsch VM, Johnson PR. 1994. Pathogenic diversity of simian immunodeficiency viruses. Virus Res. 32:183–203. 10.1016/0168-1702(94)90041-8 [DOI] [PubMed] [Google Scholar]
- 97.Pandrea I, Onanga R, Rouquet P, Bourry O, Ngari P, Wickings EJ, Roques P, Apetrei C. 2001. Chronic SIV infection ultimately causes immunodeficiency in African non-human primates. AIDS 15:2461–2462. 10.1097/00002030-200112070-00019 [DOI] [PubMed] [Google Scholar]
- 98.Apetrei C, Gormus B, Pandrea I, Metzger M, ten Haaft P, Martin LN, Bohm R, Alvarez X, Koopman G, Murphey-Corb M, Veazey RS, Lackner AA, Baskin G, Heeney J, Marx PA. 2004. Direct inoculation of simian immunodeficiency virus from sooty mangabeys in black mangabeys (Lophocebus aterrimus): first evidence of AIDS in a heterologous African species and different pathologic outcomes of experimental infection. J. Virol. 78:11506–11518. 10.1128/JVI.78.21.11506-11518.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ling B, Apetrei C, Pandrea I, Veazey RS, Lackner AA, Gormus B, Marx PA. 2004. Classic AIDS in a sooty mangabey after an 18-year natural infection. J. Virol. 78:8902–8908. 10.1128/JVI.78.16.8902-8908.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Walston J, McBurnie MA, Newman A, Tracy RP, Kop WJ, Hirsch CH, Gottdiener J, Fried LP, Cardiovascular Health Study 2002. Frailty and activation of the inflammation and coagulation systems with and without clinical comorbidities: results from the Cardiovascular Health Study. Arch. Intern. Med. 162:2333–2341. 10.1001/archinte.162.20.2333 [DOI] [PubMed] [Google Scholar]
- 101.Cohen HJ, Harris T, Pieper CF. 2003. Coagulation and activation of inflammatory pathways in the development of functional decline and mortality in the elderly. Am. J. Med. 114:180–187. 10.1016/S0002-9343(02)01484-5 [DOI] [PubMed] [Google Scholar]
- 102.Douek DC, Roederer M, Koup RA. 2009. Emerging concepts in the immunopathogenesis of AIDS. Annu. Rev. Med. 60:471–484. 10.1146/annurev.med.60.041807.123549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Paiardini M, Cervasi B, Reyes-Aviles E, Micci L, Ortiz AM, Chahroudi A, Vinton C, Gordon SN, Bosinger SE, Francella N, Hallberg PL, Cramer E, Schlub T, Chan ML, Riddick NE, Collman RG, Apetrei C, Pandrea I, Else J, Munch J, Kirchhoff F, Davenport MP, Brenchley JM, Silvestri G. 2011. Low levels of SIV infection in sooty mangabey central memory CD4+ T cells are associated with limited CCR5 expression. Nat. Med. 17:830–836. 10.1038/nm.2395 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Traina-Dorge V, Blanchard J, Martin L, Murphey-Corb M. 1992. Immunodeficiency and lymphoproliferative disease in an African green monkey dually infected with SIV and STLV-I. AIDS Res. Hum. Retroviruses 8:97–100. 10.1089/aid.1992.8.97 [DOI] [PubMed] [Google Scholar]
- 105.Beer B, Denner J, Brown CR, Norley S, zur Megede J, Coulibaly C, Plesker R, Holzammer S, Baier M, Hirsch VM, Kurth R. 1998. Simian immunodeficiency virus of African green monkeys is apathogenic in the newborn natural host. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 18:210–220. 10.1097/00042560-199807010-00003 [DOI] [PubMed] [Google Scholar]
- 106.Courgnaud V, Abela B, Pourrut X, Mpoudi-Ngole E, Loul S, Delaporte E, Peeters M. 2003. Identification of a new simian immunodeficiency virus lineage with a vpu gene present among different cercopithecus monkeys (C. mona, C. cephus, and C. nictitans) from Cameroon. J. Virol. 77:12523–12534. 10.1128/JVI.77.23.12523-12534.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Courgnaud V, Salemi M, Pourrut X, Mpoudi-Ngole E, Abela B, Auzel P, Bibollet-Ruche F, Hahn B, Vandamme AM, Delaporte E, Peeters M. 2002. Characterization of a novel simian immunodeficiency virus with a vpu gene from greater spot-nosed monkeys (Cercopithecus nictitans) provides new insights into simian/human immunodeficiency virus phylogeny. J. Virol. 76:8298–8309. 10.1128/JVI.76.16.8298-8309.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Bailes E, Gao F, Bibollet-Ruche F, Courgnaud V, Peeters M, Marx PA, Hahn BH, Sharp PM. 2003. Hybrid origin of SIV in chimpanzees. Science 300:1713. 10.1126/science.1080657 [DOI] [PubMed] [Google Scholar]
- 109.Fenton-May AE, Dibben O, Emmerich T, Ding H, Pfafferott K, Aasa-Chapman MM, Pellegrino P, Williams I, Cohen MS, Gao F, Shaw GM, Hahn BH, Ochsenbauer C, Kappes JC, Borrow P. 2013. Relative resistance of HIV-1 founder viruses to control by interferon-alpha. Retrovirology 10:146. 10.1186/1742-4690-10-146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Goonetilleke N, Liu MK, Salazar-Gonzalez JF, Ferrari G, Giorgi E, Ganusov VV, Keele BF, Learn GH, Turnbull EL, Salazar MG, Weinhold KJ, Moore S, Letvin B CCCN, Haynes BF, Cohen MS, Hraber P, Bhattacharya T, Borrow P, Perelson AS, Hahn BH, Shaw GM, Korber BT, McMichael AJ. 2009. The first T cell response to transmitted/founder virus contributes to the control of acute viremia in HIV-1 infection. J. Exp. Med. 206:1253–1272. 10.1084/jem.20090365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Keele BF, Estes JD. 2011. Barriers to mucosal transmission of immunodeficiency viruses. Blood 118:839–846. 10.1182/blood-2010-12-325860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Ciaranello AL, Perez F, Keatinge J, Park JE, Engelsmann B, Maruva M, Walensky RP, Dabis F, Chu J, Rusibamayila A, Mushavi A, Freedberg KA. 2012. What will it take to eliminate pediatric HIV? Reaching WHO target rates of mother-to-child HIV transmission in Zimbabwe: a model-based analysis. PLoS Med. 9:e1001156. 10.1371/journal.pmed.1001156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Chahroudi A, Meeker T, Lawson B, Ratcliffe S, Else J, Silvestri G. 2011. Mother-to-infant transmission of simian immunodeficiency virus is rare in sooty mangabeys and is associated with low viremia. J. Virol. 85:5757–5763. 10.1128/JVI.02690-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Kuhmann SE, Madani N, Diop OM, Platt EJ, Morvan J, Muller-Trutwin MC, Barre-Sinoussi F, Kabat D. 2001. Frequent substitution polymorphisms in African green monkey CCR5 cluster at critical sites for infections by simian immunodeficiency virus SIVagm, implying ancient virus-host coevolution. J. Virol. 75:8449–8460. 10.1128/JVI.75.18.8449-8460.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Abdool Karim Q, Sibeko S, Baxter C. 2010. Preventing HIV infection in women: a global health imperative. Clin. Infect. Dis. 50(Suppl 3):S122–S129. 10.1086/651483 [DOI] [PMC free article] [PubMed] [Google Scholar]