

Abstract
Previously, electrophysiological studies in adult zebrafish have been limited to slice preparations or to eye cup preparations and electrorentinogram recordings. This paper describes how an adult zebrafish can be immobilized, intubated, and used for in vivo electrophysiological experiments, allowing recording of neural activity. Immobilization of the adult requires a mechanism to deliver dissolved oxygen to the gills in lieu of buccal and opercular movement. With our technique, animals are immobilized and perfused with habitat water to fulfill this requirement. A craniotomy is performed under tricaine methanesulfonate (MS-222; tricaine) anesthesia to provide access to the brain. The primary electrode is then positioned within the craniotomy window to record extracellular brain activity. Through the use of a multitube perfusion system, a variety of pharmacological compounds can be administered to the adult fish and any alterations in the neural activity can be observed. The methodology not only allows for observations to be made regarding changes in neurological activity, but it also allows for comparisons to be made between larval and adult zebrafish. This gives researchers the ability to identify the alterations in neurological activity due to the introduction of various compounds at different life stages.
Keywords: Neuroscience, Issue 81, Zebrafish, adult, Electrophysiology, in vivo, craniotomy, perfusion, neural activity
Introduction
In this article, a protocol is described for obtaining in vivo recordings of neural activity in adult zebrafish. Extracellular recording methods are used, providing voltage measurements of electrical activity within a small region of neural tissue. This method of investigation involves monitoring a large number of cells in a behaving animal1. Previously, slice recordings have been performed in both adults and larvae, as have eye cup preparations and electroretinogram recordings. These experiments have largely been performed to detail physiological responses of various sensory systems2-5. Until recently, intact brain preparations have only been available for performing electrophysiology with zebrafish larva3,6,7, where respiration and oxygen diffusion can occur through the skin. Our preparation allows the native neurological activity of an adult zebrafish to be measured while the animal remains fully conscious and aware of its surroundings.
Zebrafish (Danio rerio) currently play a fundamental role as a model for genetic, toxicological, pharmacological, and physiopathological studies3. Zebrafish have gained visibility within the field of neuroscience because they share extensive homology with mammals at the genetic, neural and endocrine levels8. Over the past decade, standard neuroanatomic and immunohistochemical techniques have been used to determine the detailed characteristic organization of the zebrafish nervous system9-12 and of the distribution of different neurotransmitters3,8,13. More recently, researchers have shifted their focus to functional studies14,15, many of which center on behavioral processes16-19 and electrophysiological characteristics of sensory systems2,13,20. A small number of these studies have concentrated on the electrical activity of specific areas of the adult zebrafish brain21-23, but were not carried out using an in vivo approach.
This protocol can be adapted for electrophysiological studies of both spontaneous and evoked activity within the zebrafish nervous system to describe the patterns of activity in specific brain regions. The use of this technique allows comparisons to be made between the neurological activity of young larval stages and adults. Further, our protocol permits comparisons between genetic or pharmacological alterations. Together with other approaches, such as genetic engineering or pharmacological tests, this method offers a new possibility for the functional analysis of neuronal communication and plasticity in the intact adult animal as well as for potential applications, such as studying late onset epilepsy or neurodegenerative processes.
Protocol
All experimental procedures were conducted in strict accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and followed protocol #A2011 09-003, which was reviewed, approved, and overseen by the University of Georgia Institutional Animal Care and Use Committee.
1. Equipment Setup
- Perfusion system for craniotomy Immobilization of the adult necessitates an intubation system to deliver dissolved oxygen to the fish. A variety of systems can be utilized, but a simple gravity system consisting of a 60 ml syringe is used. Elevate the pressure head to a height that consistently yields a flow rate of ~1 ml/min. This syringe can be placed in series with the syringes that are then used for the electrophysiological perfusion system.
- Obtain a single 60 ml Luer lock syringe tube and remove the plunger.
- Suspend the syringe approximately 8 in above the base of a ring-stand using a clamp.
- Connect a one-way stopcock to the end of the syringe. To the opposite end of the stopcock, connect a piece of tubing, 2 mm in diameter, which is long enough to extend to the intubation base.
- Perfusion system for electrophysiology For electrophysiological experiments, it is often necessary to introduce a variety of compounds during the course of an experiment. A variety of systems are available, but a gravity system consisting of 60 ml syringes in series can be used. This setup allows for simple, serial introduction of solutions to be carried out by opening the corresponding stopcock.
- Obtain two or more 60 ml Luer lock syringes and remove the plungers from each. One syringe is required to administer habitat water to the fish while each subsequent tube can be used to administer a variety of pharmacological compounds to the fish.
- Connect each syringe to a 3-way stopcock with a Luer connection.
- Use ⅛ in outer diameter (O.D.) tubing to connect the syringes in series with a male Luer lock on one end.
- Secure the device such that the pressure head is elevated to a height of 27 in above the base, or a height that consistently yields a flow rate of ~1 ml/min.
- Intubation cannula The intubation cannula facilitates the introduction of fluid to the fish. This setup provides both flexibility and firmness to best position the cannula within the animal's mouth.
- Using any general pair of scissors (e.g. Fiskars) or a razor blade, remove a 1.5 cm section from the wide end of a P-200 pipette tip.
- Insert a 6 cm x 1 mm piece of tubing into a reducing valve with a female Luer lock cap and silicone ferrule, i.e. a Tuohy Borst adapter, and insert this cannula into the modified pipette tip. Hold the pipette tip in position with the Luer lock using a short portion of ⅛ in diameter tubing.
 Figure 1
Figure 1
- Intubation base This dish is used to hold the immobilized animal in a stable, upright position and to facilitate the removal of the fluid entering the animal through intubation. If this is not established correctly, pooling can occur, leading to intrusive vibrational signals in the electrophysiological recording.
- Obtain the base dish of a 100 mm x 15 mm plastic Petri dish.
- Using a soldering tool, melt a hole, 6 mm in diameter and 7.5 mm below the lip of the rim, into the side of the dish. Using the same tool, melt a hole, 10 mm in diameter and 11 mm below the lip of the dish ~74° counter-clockwise from the smaller hole. This hole will serve as the drainage hole.
- Insert a tube,1 cm in diameter, into the hole with the end lifted such that the end cannot be blocked. Then insert a P-200 Pipetteman tip into the small hole; this will serve as a dummy cannula.
- With the Petri dish slightly elevated on the side of the larger hole, pour melted canning wax into the dish such that the cannula hole is just covered. The region will serve as a stop, allowing the cannula to be inserted ~3 mm into the mouth of the fish. This set up should be allowed to solidify.
- Using a flame-heated metal edge, carve a channel to hold the fish so that the channel begins from the tip of the cannula and extends approximately 1-2 in.
- Caution must be taken to ensure that a continuous downward angle from the cannula area to the drainage port is formed.
 Figure 2
Figure 2
- Perfusion setup
- Before beginning, flush the perfusion set-ups with habitat water, ensuring that all air bubbles are removed.
- Add ~30 ml of 0.016% (630 μM) tricaine solution to the craniotomy perfusion setup and allow ~2 ml to run through tubing to ensure that tricaine will be delivered to the animal upon perfusion initiation. See step 2.2 for dilution.
- To the main perfusion setup, add ~50 ml of habitat water to the first tube. Add any desired experimental compounds to each of the remaining tubes.
- Place the cannula setup into the small hole of the intubation base.
- Twist a 1 in wide strip of 42 cm Kimwipe into a tight spiral and insert it into the drainage tube such that the Kimwipe extends on both ends. This tissue will serve as a wick to remove the perfused liquid and to prevent fluid buildup within the intubation base, which could interfere with electrophysiological recording.
- Insert one end of the tubing into the large hole of the Petri dish, allowing the lower end to extend into an inert dish that is large enough to collect all perfusion waste. This dish will serve as the outflow reservoir for the setup. Position the tissue so that one end will lie near the animal's abdomen region and the lower extension can drip into the dish.
- Place this completed intubation base near the dissecting microscope. Connect the Luer lock of the cannula to the tricaine perfusion tubing.
- Capillary needle and electrodes
- Obtain a primary electrode consisting of 2.5 in of 0.010 in silver wire and a secondary electrode which should be made up of 15 in of the same wire to ground the setup. For the secondary electrode, use a 15 in section of 0.010 in silver wire with a soldered tip, which allows it to fit in back of a head-stage.
- Electroplate the tips of both the primary and secondary silver wires with chloride ions.
- Using a micropipette puller, pull a thin-walled, borosilicate capillary needle with a resistance of <15 mΩ.
- Fill the capillary needle with 2-3 μl of 2 M potassium chloride, or enough to partially cover the chloride-coated tip of the silver wire.
- Insert the primary electrode into the capillary head and mount it in the electrode holder.
- Mount the head-stage into a micromanipulator and then mount the secondary electrode into a second micromanipulator. With the setup, first position each electrode and then fit the soldered tip of the secondary electrode into the back of the head-stage.
2. Preparation of Solutions, Perfusion System, and Electrophysiological Recording Equipment
Obtain 1 L of habitat water from aquarium of fish.
- 0.016% T]tricaine methanesulfonate (tricaine) (50 ml of 630 μM)24.
- Thaw an aliquot of 0.4% Tris-buffered tricaine, pH 7.2.
- Add 2.1 ml of 0.4% Tris-buffered tricaine to 47.9 ml of habitat water and mix.
Thaw stock concentration of desired water-soluble experimental compound (e.g. 300 mM pentylenetetrazol, a common chemoconvulsant).
Thaw out an aliquot of 1 μg/μl pancuronium bromide in Ringer's solution.
- Fill both anesthetizing and experimental intubation systems with habitat water and drain at least enough to remove all bubbles from tubes. This must be done because bubbles obstruct fluid flow, leading to asphyxiation of the animal.
- Completely drain the perfusion tubes that will hold drugs without allowing air into the connecting tubing.
- Fill the anesthetizing tube with 0.016% (630 μM) tricaine solution and place any experimental compound(s) in additional tube(s) on experimental perfusion system.
- The first tube of experimental perfusion should contain only habitat water.
Moisten the small pore sponge with habitat water and place in an empty 60 mm x 15 mm Petri dish. This dish will be used to hold the fish while the anesthetic is injected.
3. Craniotomy
Add enough of the 630 μM tricaine solution to a 15 x 60 mm petri dish to fill it about three-quarters of the way. Weigh the filled dish. This will be used to immobilize the animal so that further anesthesia can be injected intraperitoneally.
- Immerse the animal into the dish containing tricaine and weigh the dish again.
- Subtract the difference between the weights from steps 1 and 2 to obtain the weight of the fish.
- Allow the animal to remain in the tricaine solution until calm and most movement has ceased.
- Using a pair of broad forceps, transfer the fish to the premoistened sponge and position it laterally. Transferring the fish by the tail works well for this process.
- Place the sponge and fish under the dissecting microscope.
With a Nanofil syringe containing a 34 G needle, measure out enough pancuronium bromide such that there is 1 μg/g of fish weight.
Using a popsicle stick along the dorsal side of the fish to hold it steady, administer the pancuronium bromide intraperitoneally using the Nanofil syringe.
Using the fine forceps, grab the fish by the lower jaw and quickly transfer the animal to the intubation base.
- Position the animal so that it is dorsal-up on the wax form and such that the 1 mm cannula can be inserted into the mouth. Use the fine forceps to maneuver the fish and open the mouth around the cannula. Due to the makeup of the intubation tray, a stop was engineered into the base, allowing the cannula to be inserted 3 mm into the mouth of the fish.
- Once in position, turn on the gravity-fed perfusion tube containing tricaine.
- Cut a 3 cm2 section of Kimqipe tissue and wet it with habitat water.
- Place the tissue over the animal to prevent desiccation. The Kimwipe section can also be positioned to aid in holding the animal dorsal-up.
- Under the dissection microscope, use vanna spring scissors to remove ~2 mm2 section of the cranium covering the optic tectum. This area looks like a dark, bony plate that sits behind the eye.
 Figure 3
Figure 3
- Insert one blade of the vanna scissors at the edge of the plate and push with enough force to penetrate the bone, without piercing the brain. Close the scissors to snip the bone.
- Remove the piece of bone.
- If any blood is present, remove using the edge of a Kimwipe. Bleeding generally stops shortly after carrying out the craniotomy.
4. Electrophysiology
- Shut off the intubation stop-cock and quickly disconnect the Luer lock connecting the intubation base to the tricaine drip.
- Move the intact intubation setup to the electrophysiology microscope and connect to the perfusion system.
- Turn on the habitat water and perfuse at a rate of 1 ml/min for ~1 hr, in order to wash out the tricaine.
Using a micromanipulator under visual control, position the secondary electrode so that the electrode tip can be inserted into the animal's nostril or into the dip behind the upper jaw.
- Insert the primary electrode needle into the craniotomy opening. Insert the needle into the tissue, such that the tip is positioned fairly superficial within the optic tectum.
- If the electrode is too deep, the electrical signal may be small.
- Collect and analyze the electrical difference recorded between the primary and secondary reference electrodes. This process will allow for extracellular recordings to be obtained.
- Collect the data in Gap-free mode at a 5 kHz sample rate, with a low pass filter of 0.1 kHz and a high pass filter of 1 Hz.
- Do not begin recording electrophysiological activity until habitat water has perfused for a minimum of 45 min.
- Record a baseline of native activity for at least 15 min prior to the addition of experimental drugs. Begin perfusion with experimental drugs of choice and record for the desired amount of time. In healthy preparations, it is possible to record the neurological activity for 2-3 hr.
5. Clean Up
- At the end of the experiment, remove the electrodes from the cranium and nostril and remove the intubation setup.
- Euthanize the animal by drug overdose using tricaine in accordance with the accepted practices, as outlined by the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals (2013).
- Zebrafish to be euthanized should be placed in a tricaine solution at 200-500 mg/L. Fish are to be left in the solution for a minimum of 10 min after cessation of rhythmic opercular activity, after which they are subjected to a physical means of euthanasia (cervical transection) to ensure death.
- Disconnect the Luer lock connecting the intubation base to the perfusion setup. Collect any remaining liquid in the perfusion system for appropriate disposal.
- Run DI water through all tubes and collect for appropriate disposal, if hazardous.
- To sanitize the perfusion system, run 70% ethanol through all tubes and collect for appropriate disposal.
- Leave the stopcocks for each tube in the open position to facilitate air drying.
- Soak the electrodes in 70% ethanol and allow to air dry.
Dispose of the capillary needle used for the primary electrode in a sharps container.
- Dismantle the intubation setup and rinse all pieces with water.
- Clean each piece with 70% ethanol and allow to air dry.
Representative Results
This protocol has been used to measure the neural activity of adult zebrafish in vivo. These electrophysiological recordings are consistently and reproducibly obtained. Figure 5 shows a representative example of native and induced alterations of the neural activity of an adult zebrafish when pentylenetetrazol (PTZ), a common chemoconvulsant6,7,25,26, is introduced in the intubation setup.
The native neurological activity of the adult zebrafish was monitored for each experiment (Figure 5A). It was consistently observed that this activity is spontaneous and small in amplitude during the duration of the recording. Following the introduction of 15 mM PTZ to the system, spontaneous epileptiform-like discharges began to develop about 5 min. following introduction. This activity was initially brief, small amplitude and occurred frequently, similar to what was observed within larval zebrafish6,7. With continued exposure to the PTZ, a consistent pattern of large amplitude discharges developed that were followed by smaller, more frequent bursts of activity and a subsequent quiet period (Figure 5B).
This technique has been used successfully to introduce other chemoconvulsants through the intubation system. With these compounds, changes in native neurological activity were also effectively evoked. Pentylenetetrazol was used here as an example due it ability to robustly alter the native neural activity of the zebrafish in a stereotypical manner.
Discussion
This protocol has been used to measure the neural activity of adult zebrafish in vivo. With practice, neural activity can be observed consistently, though the characteristics (amplitude and shape of events) of the recorded activity can vary between individual fish. Utilization of the extracellular recording technique can explain this observation. The method provides simultaneous monitoring of a large number of cells within a region1, so variations in the positioning of the primary electrode can play a role in the observed activity. Depth of the primary electrode also changes the region being measured. If the tip of the electrode is positioned too deep within a region, the observed signal will be smaller than expected and alterations in activity will be harder to observe; if this occurs, simply pull back the primary electrode so that the tip is sitting more superficially. Take note that this is characteristic of the optic tectum and has not been looked at in depth in other regions of the brain. However, this procedure could be carried out within a different region of the brain. If the observed neural activity decreases to baseline levels during the experiment and it is uncertain if the fish is still alive, remove the electrodes from the cranium and check to see if there is still blood flow throughout the animal. This can be checked by observing the large vessels in the lip or nasal area of the fish. Blood flow can be easily seen in these regions. If the electrodes are removed to check for blood flow in the middle of a recording, when repositioning the electrodes, they will be in a slightly different location than they were initially, negating the ability to directly compare this portion of the recording to the original baseline.
Before beginning this procedure, it must be ensured that the intubation tubes as well as the perfusion tubes for both the craniotomy and the electrophysiology setups are free of any bubbles. If bubbles do collect, they can be run out of the system by opening the stopcock and allowing some of the habitat water to run through. If this does not eliminate the problem, a small syringe can be attached to the end of the tube, where the cannula would be placed, and light suction applied to remove any remaining air within the system. For stubborn bubbles, habitat water can be forced through the tubing. Also, make sure that the perfusion tubes are primed and dripping before intubating the fish. If a flow rate of ~1 ml/min is not obtained, the height of the pressure head can be altered in order to achieve this. This will ensure that the fish has access to sufficient water flow. When the electrophysiology perfusion setup is setup with numerous tubes in series, it was determined that it took about ~1 min for the new solution to be introduced to the animal, though resulting changes in the recorded neurological activity were dependent upon the compound being introduced. It is advised that this flow rate be measured prior to carrying out any experiments. Previous work has shown that tricaine can attenuate neural activity, requiring it to be removed from the system in order to obtain accurate recordings27,28. Through previous work, it was determined that a period of 45-60 min of intubation with habitat water is required to wash out the tricaine and for neural activity to reach native levels. Similarly, washout of pentylenetetrazole was also carried out, and it was determined that a period of 20 min was required to return neural activity to baseline levels. Therefore, experimenters should complete trials to determine washout times for each compound of interest being introduced into the system.
This technique requires some practice. When carrying out the craniotomy, extra care must be taken to ensure that a small region of the bony plate that covers the head can be punctured and removed without damaging the brain. This can be done by introducing the vanna scissors at an acute angle with respect to the head. The bony area covering the optic tectum has fusions near the center that are weak and serve as good entry points for the scissors. Once a small break has been made within the plate, use scissors to trim a small area of the bone, and remove using a pair of fine forceps. Younger fish often have softer, more pliable craniums, so fish that are 1 year of age or less are suggested for use while learning this technique. Occasionally, blood will begin to collect around the region of the craniotomy, but this can be removed by dabbing a Kimwipe gently around the area. With this procedure, the craniotomy is very small, being about 2 mm2 in area. Due to this small size, desiccation of the cranial region has not occurred. However, if the craniotomy is larger in size, it is possible to apply agar or some other material to prevent moisture loss from the brain if this does become a problem.
A point of interest is the use of the pancuronium bromide. This chemical is stored in aliquots of 10 μl in order to prevent repetitive freezing and thawing. The suggested volume to be used in an experiment, being 1 μl per gram of weight, is usually sufficient to immobilize an adult fish. On occasion, however, this volume is not adequate. When this occurs and paralysis is not obtained, more pancuronium bromide can be administered intraperitoneally to the fish, as long as recording has not begun. When carrying out the injection, do not try to inject through the tough side scales; inject the fish in the softer abdomen region, near the vent. If the fish begins to move once the primary electrode has been positioned, there is a good chance that the needle has been broken off within the craniotomy, and the procedure must be repeated using a new animal. Through this work, it has been determined that pancuronium bromide can reduce the recorded neural activity in larval zebrafish (≤50% of baseline amplitude) and it is assumed that the same holds true for adults. However, in recent experience, the neural activity in an adult is robust enough that any dampening effects attributed to pancuronium bromide do not appear to adversely affect the ability to collect and analyze data. In contrast, confounds associated with motion can be significant. For this procedure, the benefits of complete immobility of the animal is of value beyond this limitation and the neural changes that are introduced are robust enough to persist beyond any dampening effects. Also, any motion from the animal can displace the electrode, be registered by the electrophysiology recording equipment and possibly endanger the animal.
The limitations of this technique primarily lie in the fact that extracellular recording is the only possible way of recording neurological activity within the intact adult zebrafish. When positioning the primary electrode, the needle must be inserted into the craniotomy, preventing one from seeing exactly where the electrode is being placed. Though, with practice, it is possible to position the electrode within the same region and at the same depth consistently.
In vivo electrophysiology has largely been limited to zebrafish larvae. This is due to the ability to easily immobilize the small fish and the fact that they do not require intubation. Therefore, electrophysiological studies have focused on the larval stages and little has been done regarding the electrophysiological activity within the adult brain. This has prevented any comparisons between juvenile and adult neurological activity to be made. The use of the multitube intubation system permits for easy introduction of a variety of compounds to the fish. This also allows for the effects of different chemicals or drugs to be studied within both the larval and adult systems.
Figure 1. Intubation Cannula. Remove a 1.5 cm section from the wide end of a yellow P-200 Pipetteman tip (A). Insert a 6 cm x 1 mm piece of tubing (arrow) into a Luer Lock reducing connector (B) and insert this cannula into the modified pipette tip. Hold the pipette tip in position with the Luer lock using a short portion of ⅛ in tubing (C).
Figure 2. Finished intubation base setup. Once the Gulf Wax has cooled, place the cannula setup into the smaller hole of the intubation base (A). Twist a 1 in wide strip of 42 cm Kimwipe into a tight spiral and insert it into the drainage tube such that the Kimwipe extends on both ends. Insert one end of the drainage tubing into the large hole of the Petri dish (B), allowing the lower end to extend into a 30 mm x 100 mm crystallizing dish (not in view). Position the tissue so that one end will lie near the animal's stomach area (outlined with black-dashed line) and the lower extension can drip into the dish.
Figure 3. Craniotomy. Under the dissection microscope, use vanna spring scissors to remove ~2 mm2 section of the cranium covering the optic tectum. This area looks like a dark, trapezoidal bony plate that sits behind the eye. The dotted circle demarcates where the craniotomy is positioned. The arrowhead shows there the needle of the primary electrode is positioned within the hole of the craniotomy. The primary electrode consists of a 2.5 in section of 0.010 in silver wire that has been electroplated with chloride ions and inserted into a borosilicate capillary needle. The capillary needle is filled with 2-3 μl of 2 M potassium chloride, or enough to partially cover the chloride-coated tip of the silver wire. The secondary electrode consists of a 15 in section of the same wire used for the primary electrode except that a tip is soldered at one end, allowing it to be inserted into the back of the head-stage. The end of the secondary wire that is to be positioned touching the fish must also be electroplated with chloride ions. The arrow shows where the secondary electrode has been positioned within the right nostril of the adult fish.
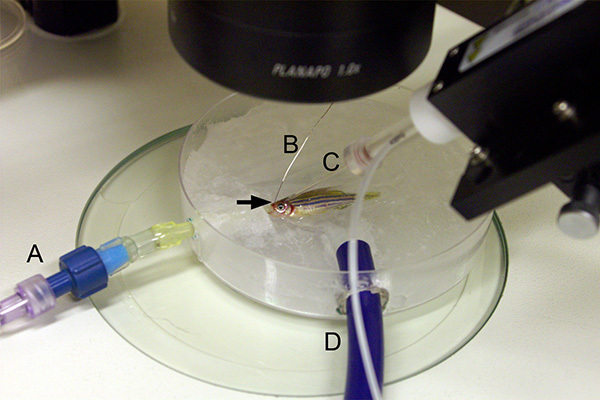 Figure 4. Setup for collection of electrophysiological activity. The intact intubation setup is moved to the electrophysiology microscope and the cannula is connected to the perfusion system (A). The system water must be turned on, allowing perfusion to start immediately. Using the micromanipulator, position the secondary electrode (B) so that the electrode tip is inserted into the animal's nostril or into the dip behind the upper jaw (arrow). Insert the primary electrode needle (C) into the craniotomy opening. Insert the needle such that it is positioned fairly superficial within the optic tectum. The large drainage tube (D) extends from the intubation dish, out of the field of view, and the other end empties into the collection dish.
Figure 4. Setup for collection of electrophysiological activity. The intact intubation setup is moved to the electrophysiology microscope and the cannula is connected to the perfusion system (A). The system water must be turned on, allowing perfusion to start immediately. Using the micromanipulator, position the secondary electrode (B) so that the electrode tip is inserted into the animal's nostril or into the dip behind the upper jaw (arrow). Insert the primary electrode needle (C) into the craniotomy opening. Insert the needle such that it is positioned fairly superficial within the optic tectum. The large drainage tube (D) extends from the intubation dish, out of the field of view, and the other end empties into the collection dish.
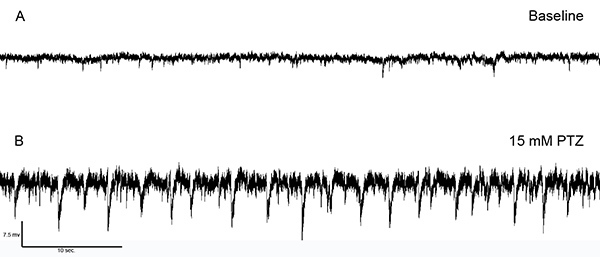 Figure 5. Electrophysiological recordings within the optic tectum. (A) It was consistently observed that the native activity within the optic tectum of the adult zebrafish is stereotypically spontaneous, showing small amplitude (2-10 mV) activity throughout the duration of the recording. (B) Following the introduction of 15 mM PTZ to the system, spontaneous epileptiform-like discharges began to develop about 5 min following introduction. This activity was initially brief, small amplitude. With continued exposure to the PTZ, a consistent pattern of large amplitude (up to 15 mV) discharges developed that were followed by smaller, more frequent bursts of activity.
Figure 5. Electrophysiological recordings within the optic tectum. (A) It was consistently observed that the native activity within the optic tectum of the adult zebrafish is stereotypically spontaneous, showing small amplitude (2-10 mV) activity throughout the duration of the recording. (B) Following the introduction of 15 mM PTZ to the system, spontaneous epileptiform-like discharges began to develop about 5 min following introduction. This activity was initially brief, small amplitude. With continued exposure to the PTZ, a consistent pattern of large amplitude (up to 15 mV) discharges developed that were followed by smaller, more frequent bursts of activity.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
This work was supported by NIH/NINDS Grant R01NS070159 (to T.M.D, J.D.L and A.T.S).
References
- Henze DA, et al. Intracellular features predicted by extracellular recordings in the hippocampus in vivo. J. Neurophysiol. 2000;84:390–400. doi: 10.1152/jn.2000.84.1.390. [DOI] [PubMed] [Google Scholar]
- Gabriel JP, et al. Locomotor pattern in the adult zebrafish spinal cord in vitro. J.Neurophysiol. 2008;99:37–48. doi: 10.1152/jn.00785.2007. [DOI] [PubMed] [Google Scholar]
- Vargas R, Johannesdottir IT, Sigurgeirsson B, Thornorsteinsson H, Karlsson KA. The zebrafish brain in research and teaching: a simple in vivo and in vitro model for the study of spontaneous neural activity. Adv Physiol Educ. 2011;35:188–196. doi: 10.1152/advan.00099.2010. [DOI] [PubMed] [Google Scholar]
- Makhankov YV, Rinner O, Neuhauss SC. An inexpensive device for non-invasive electroretinography in small aquatic vertebrates. J. Neurosci. Methods. 2004;135:205–210. doi: 10.1016/j.jneumeth.2003.12.015. [DOI] [PubMed] [Google Scholar]
- Brockerhoff SE, et al. A behavioral screen for isolating zebrafish mutants with visual system defects. Proc. Natl. Acad. Sci. U.S.A. 1995;92:10545–10549. doi: 10.1073/pnas.92.23.10545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baraban SC, Taylor MR, Castro PA, Baier H. Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression. Neuroscience. 2005;131:759–768. doi: 10.1016/j.neuroscience.2004.11.031. [DOI] [PubMed] [Google Scholar]
- Baraban SC, et al. A large-scale mutagenesis screen to identify seizure-resistant zebrafish. Epilepsia. 2007;48:1151–1157. doi: 10.1111/j.1528-1167.2007.01075.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maximino C. Serotonin and anxiety: Neuroanatomical, pharmacological and functional aspects. New York: Springer; 2012. [Google Scholar]
- Bally-Cuif L, Vernier P. Perry SF, Ekker M, Farrell AP, Brauner CJ, editors. Fish Physiology: Zebrafish. 2010.
- Kaslin J, Nystedt JM, Ostergard M, Peitsaro N, Panula P. The orexin/hypocretin system in zebrafish is connected to the aminergic and cholinergic systems. J. Neurosci. 2004;24:2678–2689. doi: 10.1523/JNEUROSCI.4908-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLean DL, Fetcho JR. Ontogeny and innervation patterns of dopaminergic, noradrenergic, and serotonergic neurons in larval zebrafish. J. Comp. Neurol. 2004;480:38–56. doi: 10.1002/cne.20280. [DOI] [PubMed] [Google Scholar]
- Mueller T, Vernier P, Wullimann MF. The adult central nervous cholinergic system of a neurogenetic model animal, the zebrafish Danio rerio. Brain Res. 2004;1011:156–169. doi: 10.1016/j.brainres.2004.02.073. [DOI] [PubMed] [Google Scholar]
- Higashijima S, Schaefer M, Fetcho JR. Neurotransmitter properties of spinal interneurons in embryonic and larval zebrafish. J. Comp. Neurol. 1002;480:19–37. doi: 10.1002/cne.20279. [DOI] [PubMed] [Google Scholar]
- Tao L, Lauderdale JD, Sornborger AT. Mapping Functional Connectivity between Neuronal Ensembles with Larval Zebrafish Transgenic for a Ratiometric Calcium Indicator. Front Neural Circuits. 2011;5:2. doi: 10.3389/fncir.2011.00002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fan X, et al. New statistical methods enhance imaging of cameleon fluorescence resonance energy transfer in cultured zebrafish spinal neurons. J Biomed Opt. 2007;12:034017. doi: 10.1117/1.2745263. [DOI] [PubMed] [Google Scholar]
- Burgess HA, Granato M. Sensorimotor gating in larval zebrafish. J. Neurosci. 2007;27:4984–4994. doi: 10.1523/JNEUROSCI.0615-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgess HA, Schoch H, Granato M. Distinct retinal pathways drive spatial orientation behaviors in zebrafish navigation. Curr. Biol. 2010;20:381–386. doi: 10.1016/j.cub.2010.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller KP, Neuhauss SC. Behavioral neurobiology: how larval fish orient towards the light. Curr. Biol. 2010;20:159–161. doi: 10.1016/j.cub.2009.12.028. [DOI] [PubMed] [Google Scholar]
- Haug MF, Biehlmaier O, Mueller KP, Neuhauss SC. Visual acuity in larval zebrafish: behavior and histology. Front. Zool. 2010;7:8. doi: 10.1186/1742-9994-7-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fetcho JR, Higashijima S, McLean DL. Zebrafish and motor control over the last decade. Brain Res.Rev. 2008;57:86–93. doi: 10.1016/j.brainresrev.2007.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connaughton VP, Nelson R, Bender AM. Electrophysiological evidence of GABAA and GABAC receptors on zebrafish retinal bipolar cells. Vis. Neurosci. 2008;25:139–153. doi: 10.1017/S0952523808080322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim YJ, Nam RH, Yoo YM, Lee CJ. Identification and functional evidence of GABAergic neurons in parts of the brain of adult zebrafish (Danio rerio) Neurosci. Lett. 2004;355:29–32. doi: 10.1016/j.neulet.2003.10.024. [DOI] [PubMed] [Google Scholar]
- Sato Y, Miyasaka N, Yoshihara Y. Hierarchical regulation of odorant receptor gene choice and subsequent axonal projection of olfactory sensory neurons in zebrafish. J. Neurosci. 2007;27:1606–1615. doi: 10.1523/JNEUROSCI.4218-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Westerfield M. The zebrafish book: a guide for the laboratory use of zebrafish (Brachydanio rerio) 1993.
- Lazarova M, Samanin R. Potentiation by yohimbine of pentylenetetrazol-induced seizures in rats: role of alpha 2 adrenergic receptors. Pharmacol. Res. Commun. 1983;15:419–425. doi: 10.1016/s0031-6989(83)80051-4. [DOI] [PubMed] [Google Scholar]
- Loscher W, Honack D, Fassbender CP, Nolting B. The role of technical, biological and pharmacological factors in the laboratory evaluation of anticonvulsant drugs. III. Pentylenetetrazole seizure models. Epilepsy res. 1991;8:171–189. doi: 10.1016/0920-1211(91)90062-k. [DOI] [PubMed] [Google Scholar]
- DeMicco A, Cooper KR, Richardson JR, White LA. Developmental neurotoxicity of pyrethroid insecticides in zebrafish embryos. Toxicol Sci. 2010;113:177–186. doi: 10.1093/toxsci/kfp258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnolds DE, et al. Physiological effects of tricaine on the supramedullary/dorsal neurons of the cunner, Tautogolabrus adspersus. Biol. Bull. 2002;203:188–189. doi: 10.2307/1543388. [DOI] [PubMed] [Google Scholar]