

Abstract
Oxidative stress plays an important role in the development of various human diseases. Aqueous chamomile extract is used as herbal medicine, in the form of tea, demonstrated to possess antiinflammatory and antioxidant properties. We demonstrate the cytoprotective effects of chamomile on hydrogen peroxide (H2O2)-induced cellular damage in macrophage RAW 264.7 cells. Pretreatment of cells with chamomile markedly attenuated H2O2-induced cell viability loss in a dose-dependent manner. The mechanisms by which chamomile-protected macrophages from oxidative stress was through the induction of several antioxidant enzymes including NAD (P)H:quinone oxidoreductase, superoxide dismutase, and catalase and increase nuclear accumulation of the transcription factor Nrf2 and its binding to antioxidant response elements. Furthermore, chamomile dose-dependently reduced H2O2-mediated increase in the intracellular levels of reactive oxygen species. Our results, for the first time, demonstrate that chamomile has protective effects against oxidative stress and might be beneficial to provide defense against cellular damage.
Keywords: chamomile, macrophages, phase II enzymes, oxidative stress, antioxidant defense
INTRODUCTION
Oxidative stress-induced cellular damage caused by reactive oxygen species (ROS) has been implicated in the aging process and remains a major factor in the development of various human diseases including diabetes, liver diseases, neurodegenerative disorders, cardiovascular diseases, and cancer (Cooke et al., 1997; Farinati et al., 1998). Dietary antioxidants have been considered as an attractive strategy to prevent or attenuate the progression of diseases caused by oxidative stress. Chamomile (Matricaria chamomilla) has been used for centuries in many human cultures to treat various inflammatory conditions such as eczema, ulcers, gout, neuralgia, and rheumatic pains (Rombi, 1993; Tyler, 1993; Bhaskaran et al., 2010). Recent studies have demonstrated its hypocholesteroemic, antiparasitic, antiaging, and anticancer properties, supporting its longstanding traditional use for treating various human ailments (Lee and Shibamoto, 2002; Babenko and Shakhova, 2006; Srivastava and Gupta, 2009). Dried flowers of chamomile are used in the preparation of tea, which is consumed at a rate of more than a million cups per day (Speisky et al., 2006). The beneficial effects of chamomile are related to the presence of several flavonoid constituents and the core structure consists of either flavone (apigenin and luteolin) or flavonol derivatives (quercetin and patuletin). These occur in various forms such as aglycoglycoside, monoglycoside, and diglycoside and/or acyl derivatives. This plant also contains high levels of polyphenolic compounds such as coumarins and hydroxycoumarins. Other principal components are essential oils such as terpenoids, α-bisabolol and its oxides, azulenes including chalmuzene, and acetylene derivatives (McKay and Blumberg, 2006). Chamomile is one of the richest sources of dietary antioxidants. The coumarins and flavonoids are soluble in hot water and, ingested as tea, provide suppressive effects on diabetic complications and oxidative damage to skins, membranes, proteins, and DNA (Paya et al., 1992; Kaneko et al., 2007). However, the mechanisms of antioxidant effects of chamomile have not been elucidated.
Antioxidants exert their effects via directly scavenging ROS and/or indirectly enhancing the endogenous antioxidant defense system (Jackson et al., 2002). Several genes encoding antioxidant phase II enzymes are coordinately induced upon exposure to electrophiles and ROS (Itoh et al., 2004). Antioxidant response element (ARE), a cis-acting regulatory element present in the promoter region of the genes encoding antioxidant proteins such as heme oxygenase-1 (HO-1), NAD(P)H:quinone oxidoreductase (NQO1), and glutathione S-transferase are involved in the detoxification and elimination of reactive oxidants and electrophilic agents through conjugative reactions and by enhancing cellular antioxidant capacity (Ishii et al., 2000; Jang et al., 2009). Central to the antioxidant phase II gene response is the Nrf2–Keap1 transcription system. Under basal conditions, Nrf2 is sequestered in the cytoplasm as an inactive complex with the repressor Kelch-like ECH-associated protein 1 (Keap1). The release of Nrf2 from its repressor is most likely to be achieved by alterations in Keap1 structure. Keap1 contains several reactive cysteine residues that function as sensors of cellular redox changes results in disruption of Nrf2–Keap1 complex that facilitates the translocation of Nrf2 into the nucleus, where it induces the transcription of phase II detoxifying genes by binding to the antioxidant responsive element ARE sequences (Motohashi and Yamamoto, 2004; Jang et al., 2009). Because of the potential role of Nrf2–Keap1 pathway in inducing antioxidant proteins and protecting the cells against oxidative damage, there is mounting interest to determine whether chamomile could activate this pathway. In this study, we used RAW 264.7 macrophages to investigate whether chamomile activates Nrf2 signaling and increases the antioxidant phase II enzyme proteins, protecting the cells from oxidative stress.
MATERIALS AND METHODS
Materials
Commercially available dry chamomile (M. chamomilla) flower of Egyptian origin was purchased from Bec’s Tea Nirvana, Cleveland, OH, USA. The voucher specimen was recorded and stored in our laboratory. Cell culture medium, Dulbecco’s modified essential medium (Hyclone, Fisher Scientific; PA, USA), fetal bovine serum, penicillin–streptomycin cocktail, and phosphate buffer saline were purchased from Cellgro Mediatech, Inc. (Herndon, VA, USA). Hydrogen peroxide (H2O2) and apigenin 7-O-glucoside (>95% pure) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies for anti-Nrf2 (sc-722), anti-Keap1 (sc-33569), anti-superoxide dismutase (SOD) (sc-11407), anti-catalase (CAT) (sc-69762), anti-NQO1 (sc-25591), and anti-β-actin (sc-47778) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA), and anti-histone 3 (05-928) was purchased from Upstate Cell Signaling solutions (Billerica, MA, USA). Secondary antibodies for mouse and rabbit (horseradish peroxidase conjugates) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Enhanced chemiluminescence kit (ECL) for chemiluminescence was purchased from GE Healthcare Biosciences (Piscataway, NJ, USA). All reagents used in the experiments were of analytical reagent grade or HPLC grade where applicable.
Preparation of extracts
Dry chamomile flowers were weighed and crushed to powder with a marble pestle and mortar, and a 5% w/v suspension was prepared in a flask by adding hot boiled water. The flask was then placed on a shaker for 4 h, and the temperature was maintained at 37 °C. After shaking, the flask was brought to room temperature and the suspension was filtered through a series of Whatman filters and finally passed through 0.22-micron filter (Millipore, Billerica, MA, USA). The filtered aqueous extract was freeze-dried and stored at −20 °C until use. For cell culture studies, the dried material from aqueous extract was weighed and dissolved in culture medium to achieve desired concentration.
HPLC analysis
Aqueous chamomile extract was analyzed on an Agilent 1200 HPLC system (Agilent Technologies, Santa Clara, CA, USA) with the use of a C-18 column. The mobile phase consisted of acetonitrile and water as the isocratic solvent (30:70, v/v) maintained at a flow rate of 1 mL/min with an injection volume of 5 μL and a run time of 8 min. Data were collected at 335 nm (λmax for the majority of the apigenin glucosides).
Mass spectrometric analysis
Electrospray ionization tandem mass spectrometry was used to identify apigenin and its derivatives in HPLC fractions. In brief, HPLC fractions were dissolved in 50% methanol and introduced onto a Quattro Ultima triple-quadruple mass spectrometer (Micromass, Inc., Beverly, MA, USA) at a rate of 50 μL/min and analyzed using electrospray ionization in both negative and positive ion modes. Apigenin and its derivatives were identified using both full and product scans. The capillary and cone voltages were set at 3.5 kV and 50 V, respectively. The desolvation and cone temperatures were set at 250 and 120 °C, respectively. The nitrogen gas flow rates for desolvation and cone were 600 and 80 L/h, respectively. Collision-induced dissociation was achieved using argon gas.
Cell culture
Murine RAW 264.7 macrophages were obtained from the American Type Culture Collection (Manassas, VA, USA) and grown in appropriate culture conditions. Cells were cultured in Dulbecco’s modified essential medium with 10% heat-inactivated fetal bovine serum, 100 μg/mL penicillin–streptomycin (Invitrogen, Carlsbad, CA, USA), and maintained in an incubator with a humidified atmosphere of 95% air and 5% CO2 at 37 °C as previously described (Bhaskaran et al., 2010).
Cell viability assay
The effect of chamomile on the viability of cells was determined by 3-[4, 5-dimethylthiazol-2-yl]-2, 5-diphenyl tetrazoliumbromide (MTT) assay. Briefly, the cells were plated at 1 × 104 cells per well in 200 μL of complete culture medium containing 5–80 μg/mL concentrations of freeze-dried aqueous extract in 96-well micro titer plates. The concentration of DMSO remained within maximum permissible concentration of 0.1% in both control and treated samples. Each concentration of chamomile was repeated in 10 wells. After incubation for desired times at 37 °C in a humidified incubator, cell viability was determined. A 50-μL MTT (5 mg/mL in phosphate buffered saline stock, diluted to working strength 1 mg/mL with media) was added to each well and incubated for 2 h after which the plate was centrifuged at 600 g for 5 min at 4 °C. The MTT solution was removed from the wells by aspiration. After careful removal of the medium, 0.1 mL of buffered DMSO was added to each well, and plates were shaken. The absorbance was recorded on a microplate reader at the wavelength of 540 nm. The effect of chamomile on growth inhibition was assessed as percent cell viability where vehicle-treated cells were taken as 100% viable. The doses of chamomile used for other experiments ranged from 5 to 40 μg/mL.
Intracellular reactive oxygen species assay
Oxidative damage of cells was performed using hydrogen peroxide. Oxidative stress of the cells was determined using the fluorescent probe 2′7′-dichlorofluorescein diacetate (DCF-DA). Briefly, RAW 264.7 cells (1×105/plate) were suspended in a 96-well microplate treated with a various concentrations of chamomile extract, with and without H2O2 (50 μM) and further incubated according to the specified time course at 37 °C. At the end of the oxidation treatment, cells were incubated with 10 μM of fluorescent probe DCF-DA for 20min at 37 °C. The fluorescence intensity was measured using FluoStar Omega Spectrophotometer (BMG Labtech; NC, USA) at excitation and emission wavelengths of 485 and 560 nm, respectively.
Measurement of NQO1 activity in RAW 264.7 cells
The NQO1 activity in RAW 264.7 was determined using dichloroindophenol (DCIP) as the two-electron acceptor, as previously described (Zhu et al., 2007). Briefly, the reaction mixture containing 50 mM Tris–HCl, pH 7.5, 0.08% Triton X-100, 0.25 mM NADPH, 80 μM 2,6-dichloroindophenol (DCIP) in the presence or absence of 60 μM dicumarol, a potent inhibitor of NQO1, and the two-electron reduction of DCIP was monitored at 600 nm, 25 °C for 3 min. The dicumarol inhibition of cellular NQO1 activity was calculated using the extinction coefficient of 21.0 mM−1 cm−1 and expressed as nanomoles of DCIP reduced per minute per milligram of cellular protein.
Measurement of superoxide dismutase activity in RAW 264.7 cells
Total SOD activity in RAW 264.7 was determined by Superoxide dismutase assay kit (Cayman Chemical Cat# 70600; MI, USA). The enzyme activity was assessed using the manufacturer’s protocol. The sample total SOD activity was calculated using a concurrently run SOD (Sigma-Aldrich) standard curve and expressed as units per milligram of cellular protein.
Measurement of catalase activity in RAW 264.7 cells
The CAT activity in RAW 264.7 cells was measured according to the Catalase assay kit (Cayman Chemical Cat# 707002; MI, USA). The enzyme activity was assessed using the manufacturer’s protocol. The cellular CAT activity was expressed as micromoles of H2O2 consumed per minute per milligram of cellular protein.
Western blot analysis
Macrophages, grown in 6-well plates to confluence, were incubated with or without H2O2 in the absence or presence of aqueous chamomile extract. Cells were washed with ice-cold PBS and stored at −70° until further analysis. Frozen plates were put on ice, and cells were lysed in 1% Triton X-100, 0.15 M NaCl, and 10 mM Tris–HCl pH 7.4 for 30 min. Lysates were homogenized through a 22 G needle and centrifuged at 10,000 g for 10 min at 4 °C. The supernatants were collected, and protein was measured by the method according to Bradford assay (Bradford, 1976). Cell lysates, containing equal amounts of protein, were boiled in SDS sample buffer for 5 min before running on a 10% SDS–polyacrylamide gel. Proteins were transferred to polyvinylidene fluoride membranes (Invitrogen, Carlsbad, CA, USA). Membranes were blocked with 5% fat-free dry milk in TBS-T pH 8.0 (Tris-buffered saline [50mM Tris, pH 8.0, 150 mM NaCl] with 0.1% Tween 20) and then incubated with primary antibodies at appropriate dilutions overnight at 4°C. After washing, the membrane was incubated with secondary antibody IgG:horseradish peroxidase conjugate and the enhanced chemiluminescence system (ECL™, Amersham Pharmacia Biotech; NJ, USA). Signal intensities were evaluated by densitometric analysis (Kodak Digital Science™ Image Station 2000R Life Science Products; NY, USA).
Electrophoretic mobility shift assay
Electrophoretic mobility shift assay (EMSA) for Nrf2 was performed in the nuclear fraction of RAW 264.7 macrophages incubated for 16 h with or without various concentrations of chamomile and H2O2 (50 μM) using Lightshift™ Chemiluminiscent EMSA kit (Pierce Biotechnology, Rockford, IL, USA) following manufacturer’s protocol as previously described (Shukla et al., 2004). The DNA probes used were Nrf2 oligonucleotides as forward (5′-CAC GAG CTG CCG GCG CTG TCC ACA TC-3′) and reverse (5′-GAT GTG GAC AGC GCC GGC AGC TCG TG-3′).
Statistical analysis
The enzyme estimations and cell viability assay were performed in triplicate. All experiments were repeated at least twice. Results are expressed as mean values ±SD. Statistical comparisons were made by ANOVA followed by a Dunnett’s multiple comparison test. p-values <0.05 were considered significant.
RESULTS
First, we analyzed for constituents present in the aqueous chamomile extract. Reports demonstrate that aqueous standardized extract of chamomile flowers contains various flavonoids including apigenin, quercetin, and patuletin as glucosides and various acetylated derivatives (Svehliková et al., 2004). We performed an HPLC scan at wavelengths ranging from 200 to 590 nm to determine various constituents present in the chamomile extract. Altogether, a total of 10 peaks were recorded in aqueous chamomile extract during the scanning process, most of which correspond to water soluble flavonoids (data not shown). The identified two major peaks with retention times of 1.15 min (27.7%) and 1.52 min (63.3%) and other minor peaks that together constitute 9% of the total flavonoids. MS–MS confirmation of these two peaks in the aqueous chamomile extract preparation corresponded to apigenin 7-O-glucoside (63.3%) and apigenin 7-O-neohespridoside (27.7%), respectively (Supplemental Figure 1).
Next, we determined the cell viability after exposure of RAW 264.7 macrophages with H2O2 and aqueous chamomile extract. H2O2 is a stable agent and causes oxidative stress and DNA damage resulting in decreased cell viability and induction of apoptosis in many different cell types (Halliwell and Aruoma, 1991; Wijeratne et al., 2005; Yoshikawa et al., 2006). Exposure to H2O2 for 6 h caused a concentration-dependent decrease in cell viability, which ranges from 82% at 50 μM to 8% at the maximum dose of 800 μM, respectively (Fig. 1A). Next, we determined the effect of chamomile on cell viability. Exposure of cells to chamomile for 24 h caused a modest decrease in cell viability, which ranges from 0.1% to 3.5% between the doses 5–40 μg/mL; 10.1% at the highest dose of 80 μg/mL (Fig. 1B). Therefore, for further studies, we elected to work on the 5–40 μg/mL, which causes minimum inhibition of cell viability.
Figure 1.
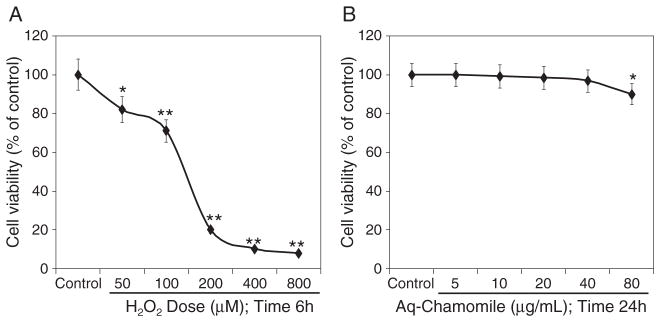
Cell viability assay. (A) Concentration-dependent effect of H2O2 on cell viability for 6 h as determined by MTT assay; (B) cell viability assay by MTT after treatment with chamomile at 0, 5, 10, 20, 40, and 80 μg/mL for 24 h in RAW 264.7 macrophage cells. Data shown are representative of three independent experiments. Mean ±SD; *p <0.05 and **p <0.001 represent significant differences as compared with the control group. The details are described in the Materials and methods section.
In the next set of experiments, we sought to determine the protective effect of chamomile in reversing the decrease in cell viability caused by H2O2. As shown in Figure 2A, RAW 264.7 macrophages were exposed to 5–40 μg/mL concentration of aqueous chamomile extract for 16 h prior to 50–200 μM doses of H2O2 exposure. Pretreatment of cells with 5–40 μg/mL chamomile caused a significant increase in cell viability with 19–30% increase at 50μM H2O2; 33–53% at 100 μM H2O2 and 157–375% at 200μM H2O2, compared with the H2O2-treated cells. We also measured the generation of ROS after H2O2 exposure to RAW 264.7 macrophages (Fig. 2B). Exposure of cells with H2O2 caused a significant increase in ROS generation as measured by the addition of DCF-DA in the culture medium, which converts to highly fluorescent dichlorofluorescein in the presence of intracellular ROS. Pretreatment of cells with 5–40 μg/mL doses of chamomile caused a decrease in ROS generation with 89–46% decrease at 50 μM H2O2; 16–51% at 100 μM H2O2 and 33–30% at 200 μM H2O2, compared with the H2O2-treated cells (Fig. 2B). The effect of chamomile was also confirmed by the morphological observations. There was a significant injury in RAW 264.7 macrophages after treatment with 50 μM H2O2 for 6 h, including the disappearance of cellular process, shrinkage of the cell and the nucleus as well as condensation of nuclear chromatin, nuclear blebbing consistent with apoptosis. Chamomile pretreatment with 20 μg/mL for 16h to these cells markedly reduced H2O2-mediated cellular damage (Fig. 2C).
Figure 2.

Effect of chamomile on cell viability and ROS generation with H2O2 in RAW 264.7 cells. (A) Cell viability assay with MTT on RAW 264.7 cells treated with chamomile at concentrations 0–40 μg/mL for 16 h and further challenged with H2O2 at 50, 100, and 200 μM for 6 h; (B) ROS assay with DCF-DA on RAW 264.7 cells treated with chamomile at concentrations 0–40 μg/mL for 16 h and further challenged with H2O2 at 50, 100, and 200 μM for 6 h; (C) photographs of RAW 264.7 macrophages (subpanel a–d) showing cells after treatment with 50 μM H2O2 for 6 h, 20 μg/mL chamomile for 16 h alone and chamomile with H2O2. The bars represent mean ± SD of at least three independent experiments each performed in triplicate, *p <0.05 and **p <0.001 represent significant differences as compared with the chamomile and H2O2 treated group. ¶p <0.05 and #p <0.001 represent significant differences as compared with the H2O2 model group. The details are described in the Materials and methods section.
To verify the effect of chamomile on the induction of phase II genes, we selected the typical phase 2 enzymes: NQO1, SOD, and CAT, which catalyze the detoxification of electrophiles. RAW 264.7 macrophages incubated with chamomile modified the activity of antioxidant enzymes compared with untreated controls. Exposure of cells with 10–40 μg/mL chamomile resulted in 1.74-fold to 1.96-fold increase in NQO1, 1.22-fold to 1.45-fold increases in SOD and 1.4-fold to 1.8-fold increases in CAT, respectively. Furthermore, treatment of cells with 50 μM H2O2 prior to chamomile treatment resulted in further increase in NQO1 (1.88-fold to 4.24-fold) and CAT (2.44-fold to 3.12-fold) activity; whereas a decrease in SOD activity was observed (1.1-fold to 0.98-fold) after treatment with H2O2 (Fig. 3A). In agreement with the results of the enzyme activity determinations, chamomile-alone treatment resulted in dose-dependent increase in the protein expression of NQO1, SOD, and CAT, compared with the untreated controls. Similar effects were noted as of enzyme activity after treatment of cells with chamomile followed by H2O2 treatment (Fig. 3B).
Figure 3.

Effect of chamomile on antioxidants enzyme activity and their respective protein expression. (A) NQO1, SOD, and CAT enzymes activity after treating the RAW 264.7 macrophages with chamomile at 0, 10, 20, and 40 μg/mL with and without H2O2. The bars represent mean ±SD of at least three independent experiments each performed in triplicate, *p <0.05 and **p <0.001 represent significant differences as compared with the chamomile and H2O2treated group. #p <0.001 represent significant differences as compared with the H2O2 model group. (B) Protein expression of the phase 2 antioxidants when RAW 264.7 macrophages treated with chamomile at 0, 10, 20, and 40 μg/mL for 16 h without H2O2 and chamomile for 16 h with H2O2 for 6 h. The numeric value below each band represents intensity performed by densitometric analysis normalized to the loading control, β-actin. The details are described in the Materials and methods section.
Because Nrf2 is a major transcription factor involved in the cellular protection against oxidative stress through ARE-mediated induction of phase 2 antioxidant enzymes, we therefore measured the levels of Nrf2. Under resting condition, Nrf2 is sequestered in the cytosol by Keap1, and chamomile treatment to RAW 264.7 macrophages resulted in significant increase of Nrf2 and Keap1 in the cytosol. Treatment of cells with H2O2 after chamomile pretreatment resulted in further increase in Keap1 in the cytosol, whereas Nrf2 levels tend to decrease in the cytosolic fraction (Fig. 4A). Chamomile treatment significantly enhanced the nuclear accumulation of Nrf2 in a dose-dependent manner and obtained the highest induction at 20–40 μg/mL doses of chamomile. Treatment with H2O2 further increased the nuclear accumulation of Nrf2 (Fig. 4B). These results suggest that chamomile stimulate Nrf2-mediated ARE activation by enhancing the nuclear accumulation of Nrf2 protein.
Figure 4.

Effect of chamomile on Nrf2/Keap1 expression in RAW 264.7 macrophages. (A) Western blot for cytosolic Nrf2 and Keap1 protein expression, and (B) nuclear expression of Nrf2 in RAW 264.7 macrophages stimulated with chamomile 0, 10, 20, and 40 μg/mL for 16h without H2O2 and chamomile for 16h with H2O2 for 6hrs as indicated. (C) EMSA assay. EMSA was performed to determine the effect of chamomile on the nuclear translocation of Nrf2 dimers and their binding to the DNA. Controls: #1 Biotin-EBNA control DNA, #2 Biotin-EBNA control DNA+EBNA extract, #3 Biotin-EBNA control DNA+EBNA extract+20-fold molar excess of unlabeled EBNA DNA. The details of the treatment have been described in the Materials and methods section.
To further confirm Nrf2-mediated ARE gene regulation, an EMSA was performed using an oligonucleotide containing a consensus of Nrf2 binding sequence in the nuclear fraction prepared after H2O2 challenge and chamomile pretreatment. A strong Nrf2 band was observed in response to chamomile treatment, which was further increased with H2O2 treatment (Fig. 4C). These results demonstrate that chamomile-activated ARE response elements through Nrf2 activation and subsequent binding, which might account for the increase in antioxidant enzyme offering protection against H2O2-mediated cellular damage.
DISCUSSION
Studies have shown that oxidative stress is a major cause of cellular damage and has been implicated in several human diseases including cancer (Valko et al., 2007). ROS such as H2O2, superoxide anion ), and hydroxyl radical (OH−) readily damage biomolecules, which leads to cellular mutation or cell death via apoptosis or necrosis. Therefore, removal of excess ROS or suppression of their generation by antioxidants may be effective in preventing oxidative damage or cell death. Considerable efforts have been made by researchers to search for natural agents with antioxidant properties. Chamomile has been used to treat various inflammatory and immune-regulated disorders and might possess antioxidant potential as well. We explored the molecular mechanism underlying antioxidant potential of chamomile to protect against H2O2-mediated cellular damage using murine RAW 264.7 macrophages. For the first time, we demonstrate that chamomile increases phase II enzymes through Nrf2 activation.
Macrophages are widely distributed cells found in mammalian tissues and are well known for their role in both innate and adaptive immunity (Yan and Hansson, 2007). Macrophages also play important role in other physiological processes, and its dysfunction has been implicated in many pathophysiological process including inflammatory disorders, neurodegeneration, atherosclerosis, and cancer (Shibata and Glass, 2009). Macrophages are critical targets of oxidative stress and demonstrate rapid response through loss of viability culminating in apoptotic and necrotic cell death. In this study, H2O2 has been used as an inducer of oxidative stress in in vitro model (Satoh et al., 1996). Exposure of H2O2 to cultured cells result in alterations in intracellular redox status and generation of hydroxyl and peroxyl radicals, which cause damage to membranes and biomolecules resulting in dose-dependent loss of cell viability (Zhang et al., 2007). However, pretreatment with various concentrations (5–40 μg/mL) of chamomile greatly reduced ROS generation and loss of cell viability, which was further confirmed by morphological observations. These results indicated that chamomile did significantly protect RAW 264.7 macrophages from H2O2-induced cytotoxicity.
It is well known that antioxidant defense enzymes play pivotal roles in preventing cellular damage. Our results demonstrate that chamomile caused increase in the enzyme activity and protein expression of NQO1, SOD, and CAT in RAW 264.7 macrophages alone and also after H2O2 exposure, although without any effect observed on SOD levels. Interestingly, the effects of chamomile on SOD expression seem to be selective after H2O2 exposure. Because in our studies we have determined the levels of Cu/Zn-SOD and not Mn-SOD, it is possible that chamomile might increase the levels of Mn-SOD and not Cu/Zn-SOD after H2O2 exposure. However, additional studies are needed to explore the mechanism(s) of SOD regulation by chamomile.
A number of studies demonstrate that Nrf2 is a critical transcription factor in regulating the expression of a variety of cytoprotective genes encoding for antioxidants and phase II enzymes. Under basal conditions, Nrf2 is sequestered in the cytoplasm by actin-binding protein Keap1 and rapidly undergoes ubiquitin-mediated proteasomal degradation assisted by Keap1 as an adaptor for Cul3-based E3 ligase (Zhang, 2006). Keap1 possess abundant-free cysteine residue (25 cysteine), which makes it an ideal redox-sensing partner. Exposure of cells to chemopreventive agents and/or oxidative stress result in phosphorylation (by upstream kinases) and/or redox modification of Nrf2/Keap1 complex leading to dissociation of Keap1 and subsequent nuclear translocation of Nrf2 (Huang et al., 2000; Nguyen et al., 2003; Giudice and Montella, 2006). In the nucleus, Nrf2 forms a heterodimer with small Maf proteins and activates the transcription of antioxidant genes via ARE. In the present study, chamomile caused dissociation of Keap1 and increased nuclear accumulation of Nrf2 and its binding to ARE presumably serves to enhance the expression of various phase II antioxidant enzymes. Whether chamomile results in posttranslational modification of these proteins by various upstream kinases remains to be elucidated.
Aqueous chamomile extract contains a mixture of various polyphenols and essential oils, which has been used as folk medicine for centuries (Srivastava and Gupta, 2007). These include therapeutically active compound class, namely, sesquiterpenes, flavonoids, coumarins, and polyacetylenes. Some major bioactive phenolic compounds present in chamomile extract are herniarin and umbelliferone (coumarin), chlorogenic acid and caffeic acid (phenylpropanoids), apigenin, apigenin-7-O-glucoside, luteolin and luteolin-7-O-glucoside (flavones), quercetin and rutin (flavonols), and naringenin (flavanone). It is well established that phenolic compounds are potent inhibitors of oxidative damage and H2O2 detoxification due to the availability of phenolic hydrogen (Rice-Evans et al., 1996). Chamomile extract is a naturally occurring mixture of polyphenols, which contributes to its potent antioxidant activity. The dose used in cell culture study is physiologically attainable in humans. Chamomile use in the form of tea might be enough to induce the expression of phase II antioxidant enzymes, which would likely contribute to the beneficial effects of this readily available dietary agent. In summary, our studies demonstrate that chamomile elicits antioxidant potential through induction of phase II enzymes through Nrf2–Keap1 signaling pathway as a means to provide protection against cellular damage.
Supplementary Material
Acknowledgments
This research work was supported by grants from the United States Public Health Services RO1 AT002709 and RO1 CA108512 to SG.
Abbreviations
- ARE
antioxidant response element
- CAT
catalase
- DCF-DA
dichlorofluorescein diacetate
- EMSA
electrophoretic mobility shift assay
- HPLC
high performance liquid chromatography
- Keap1
Kelch-like ECH-associated protein 1
- NADPH
nicotinamide adenine dinucleotide phosphate-oxidase
- NQO1
NAD(P)H dehydrogenase (quinone 1)
- H2O2
hydrogen peroxide
- ROS
reactive oxygen species
- Nrf2
nuclear factor (erythroid-derived 2)-like 2
- SOD
superoxide dismutase
Footnotes
Conflict of Interest
There is no conflict of interest in the study.
Supporting information may be found in the online version of this article (Supplementary Material)
References
- Babenko NA, Shakhova EG. Effects of chamomilla recutita flavonoids on age-related liver sphingolipid turnover in rats. Exp Gerontol. 2006;41:32–39. doi: 10.1016/j.exger.2005.08.008. [DOI] [PubMed] [Google Scholar]
- Bhaskaran N, Shukla S, Srivastava JK, Gupta S. Chamomile: an anti-inflammatory agent inhibits inducible nitric oxide synthase expression by blocking RelA/p65 activity. Int J Mol Med. 2010;26:935–940. doi: 10.3892/ijmm_00000545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–54. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
- Cooke MS, Mistry N, Wood C, Herbert KE, Lunec J. Immunogenicity of DNA damaged by reactive oxygen species implications for anti-DNA antibodies in lupus. Free Radic Biol Med. 1997;22:151–159. doi: 10.1016/s0891-5849(96)00283-3. [DOI] [PubMed] [Google Scholar]
- Farinati F, Cardin R, Degan P, et al. Oxidative DNA damage accumulation in gastric carcinogenesis. Gut. 1998;42:351–356. doi: 10.1136/gut.42.3.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giudice A, Montella M. Activation of the Nrf2–ARE signaling pathway: a promising strategy in cancer prevention. Bioessays. 2006;28:169–181. doi: 10.1002/bies.20359. [DOI] [PubMed] [Google Scholar]
- Halliwell B, Aruoma OI. DNA damage by oxygen-derived species. Its mechanism and measurement in mammalian systems. FEBS Lett. 1991;281:9–19. doi: 10.1016/0014-5793(91)80347-6. [DOI] [PubMed] [Google Scholar]
- Huang HC, Nguyen T, Pickett CB. Regulation of the antioxidant response element by protein kinase C-mediated phosphorylation of NFE2- related factor 2. Proc Natl Acad Sci USA. 2000;97:12475–12480. doi: 10.1073/pnas.220418997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishii T, Itoh K, Takahashi S, et al. Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. J Biol Chem. 2000;275:16023–16029. doi: 10.1074/jbc.275.21.16023. [DOI] [PubMed] [Google Scholar]
- Itoh K, Tong KI, Yamamoto M. Molecular mechanism activating Nrf2–Keap1 pathway in regulation of adaptive response to electrophiles. Free Radic Biol Med. 2004;36:1208–1213. doi: 10.1016/j.freeradbiomed.2004.02.075. [DOI] [PubMed] [Google Scholar]
- Jackson MJ, Papa S, Bolaños J, et al. Antioxidants, reactive oxygen and nitrogen species, gene induction and mitochondrial function. Mol Aspects Med. 2002;23:209–285. doi: 10.1016/s0098-2997(02)00018-3. [DOI] [PubMed] [Google Scholar]
- Jang JS, Piao S, Cha YN, Kim C. Taurine chloramine activates Nrf2, Increases HO-1 expression and protects cells from death caused by hydrogen peroxide. J Clin Biochem Nutr. 2009;45:37–43. doi: 10.3164/jcbn.08-262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaneko T, Tahara S, Takabayashi F. Inhibitory effect of natural coumarin compounds, esculetin and esculin, on oxidative DNA damage and formation of aberrant crypt foci and tumors induced by 1,2-dimethylhydrazine in rat colons. Biol Pharm Bull. 2007;30:2052–2057. doi: 10.1248/bpb.30.2052. [DOI] [PubMed] [Google Scholar]
- Lee KG, Shibamoto T. Determination of antioxidant potential of volatile extracts isolated from various herbs and spices. J Agr Food Chem. 2002;50:4947–4952. doi: 10.1021/jf0255681. [DOI] [PubMed] [Google Scholar]
- McKay DL, Blumberg JB. A review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.) Phytother Res. 2006;20:519–530. doi: 10.1002/ptr.1900. [DOI] [PubMed] [Google Scholar]
- Motohashi H, Yamamoto M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol Med. 2004;10:549–556. doi: 10.1016/j.molmed.2004.09.003. [DOI] [PubMed] [Google Scholar]
- Nguyen T, Sherrat PJ, Pickett CB. Regulatory mechanisms controlling gene expression mediated by the antioxidant response element. Annu Rev Pharmacol Toxicol. 2003;43:233–260. doi: 10.1146/annurev.pharmtox.43.100901.140229. [DOI] [PubMed] [Google Scholar]
- Paya M, Halliwell B, Hoult JR. Interactions of a series of coumarins with reactive oxygen species. Scavenging of superoxide, hypochlorous acid and hydroxyl radicals. Biochem Pharmacol. 1992;44:205–214. doi: 10.1016/0006-2952(92)90002-z. [DOI] [PubMed] [Google Scholar]
- Rice-Evans CA, Miller NJ, Paganga G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med. 1996;20:933–56. doi: 10.1016/0891-5849(95)02227-9. [DOI] [PubMed] [Google Scholar]
- Rombi M. Cento Piante Medicinali. Nuovo Insttuto d’Arti Grafiche; Bergamo, Italy: 1993. pp. 63–65. [Google Scholar]
- Satoh T, Sakai N, Enokido Y, Uchiyama Y, Hatanaka H. Free radical-independent protection by nerve growth factor and Bcl-2 of PC12 cells from hydrogen peroxide-triggered apoptosis. J Biochem. 1996;120:540–546. doi: 10.1093/oxfordjournals.jbchem.a021447. [DOI] [PubMed] [Google Scholar]
- Shibata N, Glass CK. Regulation of macrophage function in inflammation and atherosclerosis. J Lipid Res. 2009;50:277–281. doi: 10.1194/jlr.R800063-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shukla S, MacLennan GT, Fu P, et al. Nuclear factor-κB/p65 (Rel A) is constitutively activated in human prostate adenocarcinoma and correlates with disease progression. Neoplasia. 2004;6:390–400. doi: 10.1593/neo.04112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Speisky H, Rocco C, Carrasco C, Lissi EA, Lopez-Alarcon C. Antioxidant screening of medicinal herbal teas. Phytother Res. 2006;20:462–467. doi: 10.1002/ptr.1878. [DOI] [PubMed] [Google Scholar]
- Srivastava JK, Gupta S. Antiproliferative and apoptotic effects of chamomile extract in various human cancer cells. J Agric Food Chem. 2007;55:9470–9478. doi: 10.1021/jf071953k. [DOI] [PubMed] [Google Scholar]
- Srivastava JK, Gupta S. Health promoting benefits of chamomile in the elderly population. In: Watson RR, editor. Complementary and Alternative Therapies in the Aging Population. Elsevier Inc., Academic Press; San Diego, CA, USA: 2009. [Google Scholar]
- Svehliková V, Bennett RN, Mellon FA, et al. Isolation, identification and stability of acylated derivatives of apigenin 7-O-glucoside from chamomile (Chamomilla recutita [L.] Rauschert) Phytochemistry. 2004;65:2323–2332. doi: 10.1016/j.phytochem.2004.07.011. [DOI] [PubMed] [Google Scholar]
- Tyler VE. In: The Honest Herbal. 3. George F, editor. Stickley Co: Philadelphia; 1993. [Google Scholar]
- Valko M, Leibfritz D, Moncola J, Cronin MT, Mazura M, Telser J. Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol. 2007;39:44–84. doi: 10.1016/j.biocel.2006.07.001. [DOI] [PubMed] [Google Scholar]
- Wijeratne SSK, Cuppett SL, Schlegel V. Hydrogen peroxide induced oxidative stress damage and antioxidant enzyme response in Caco-2 human colon cells. J Agric Food Chem. 2005;53:8768–8774. doi: 10.1021/jf0512003. [DOI] [PubMed] [Google Scholar]
- Yan ZQ, Hansson GK. Innate immunity, macrophage activation, and atherosclerosis. Immunol Rev. 2007;219:187–203. doi: 10.1111/j.1600-065X.2007.00554.x. [DOI] [PubMed] [Google Scholar]
- Yoshikawa A, Saito Y, Maruyama K. Lignan compounds and 4,4′-dihydroxybiphenyl protect C2C12 cells against damage from oxidative stress. Biochem Biophys Res Commun. 2006;344:394–399. doi: 10.1016/j.bbrc.2006.03.107. [DOI] [PubMed] [Google Scholar]
- Zhang DD. Mechanistic studies of the Nrf2-Keap1 signaling pathway. Drug Metab Rev. 2006;38:769–789. doi: 10.1080/03602530600971974. [DOI] [PubMed] [Google Scholar]
- Zhang L, Yu H, Sun Y, et al. Protective effects of salidroside on hydrogen peroxide-induced apoptosis in SH-SY5Y human neuroblastoma cells. Eur J Pharmacol. 2007;564:18–25. doi: 10.1016/j.ejphar.2007.01.089. [DOI] [PubMed] [Google Scholar]
- Zhu H, Jia Z, Mahaney JE, Ross D, Misra HP, Trush MA, Li Y. The highly expressed and inducible endogenous NAD(P)H:quinine oxido-reductase1 in cardiovascular cells acts as a potential superoxide scavenger. Cardiovasc Toxicol. 2007;7:202–211. doi: 10.1007/s12012-007-9001-z. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.