

Abstract
Protease activated receptors (PARs) have been recognized as a distinctive four-member family of seven transmembrane G protein-coupled receptors (GPCRs) that can be cleaved by certain serine proteases. In recent years, there has been considerable interest in the role of PARs in allergic inflammation, the fundamental pathologic changes of allergy, but the potential roles of PARs in allergy remain obscure. Since many of these proteases are produced and actively involved in the pathologic process of inflammation including exudation of plasma components, inflammatory cell infiltration, and tissue damage and repair, PARs appear to make important contribution to allergy. The aim of the present review is to summarize the expression of PARs in inflammatory and structural cells, the influence of agonists or antagonists of PARs on cell behavior, and the involvement of PARs in allergic disorders, which will help us to better understand the roles of serine proteases and PARs in allergy.
1. Introduction
Protease activated receptors (PARs), a four-member family of GPCRs, can be cleaved by certain serine proteases within the extracellular amino terminus and expose a tethered ligand domain, which binds to and activates the receptors to initiate multiple signaling cascades. Therefore, these PAR-activating proteases are named as agonists of PARs. Since many of these proteases are produced during inflammation, PARs make important contributions to inflammatory tissue responses including exudation of plasma components, inflammatory cell infiltration, and tissue damage and repair in inflammation [1]. The PAR-activating serine proteases may derive from the circulation (e.g., coagulation factors), inflammatory cells (e.g., mast cell and neutrophil proteases), and multiple other sources (e.g., epithelial cells, neurons, bacteria, and fungi). Compounds that mimic or interfere with the PAR-activating processes are attractive therapeutic candidates: selective agonists of PARs may facilitate healing, repair, and protection, whereas protease inhibitors and PAR antagonists can impede exacerbated inflammation and pain.
In recent years, there has been considerable interest in the role of PARs [2, 3] in allergic inflammation, the fundamental pathologic changes in allergy. Since serine proteases have long been discovered to be actively involved in the pathologic process of inflammation and large amount of information on PARs is accumulated over the last two decades, it is necessary to write a literature review on PARs in allergy, which will help us to better understand the roles of serine proteases as agonists or antagonists of PARs in allergy.
2. Classification and Molecular Structures of PARs
Since the landmark study from Shaun Coughlin's group in which an expression cloning screen was used to identify the first human thrombin receptor known as PAR-1 [4], four numbers of this receptor class were found both in human and murine and designated as PAR-1, -2, -3, and -4, respectively [5]. As the newly found members of the typical seven trans-transmembrane GPCRs' family, the expression of PARs is found on the surface of cells from a wide variety of tissues [6].
The structure, activation mechanism, and signaling of PARs have been reviewed extensively [1, 5]. In brief, encoding genes for human PAR-1, -2, and -3 are located on chromosome 5 (q13), and for human PAR-4 the encoding gene is on chromosome 19 (p12). Although the location of PAR genes differs, high degree of structural similarity of all four genes predicts the conserved overall structure and function of these receptors [7, 8]. In both mouse and human, all four PARs have two exons: the first encoding a signal peptide and the second encoding the entire functional receptor protein [7]. Human PAR-1 protein is composed of 425 residues with 7 hydrophobic domains of a typical GPCR. The deduced sequence of human PAR-1 contains a potential cleavage site for thrombin within the amino tail: LDPR41↓S42FLLRN (where ↓ denotes cleavage) [4]. PAR-2 protein consists of 395 residues with the typical characteristics of a GPCR and with about 30% of the amino acid identity of human PAR-1. The extracellular amino acid terminus of 46 residues of PAR-2 contains a putative trypsin cleavage site, SKGR34↓S35SLIGKV [9]. PAR-2 is the most functionally distinct receptor in the PAR family as it is the only PAR which is not cleaved by thrombin. PAR-2 is most effectively cleaved by trypsin [9], tryptase [10], coagulation factors VIIa and Xa [11], the membrane type serine protease 1 (MT-SP1) [12], chitinase [13], and TMPRSS2, a type II transmembrane-bound serine protease [14]. Sharing about 28% sequence homology with human PAR-1 and PAR-2, human PAR-3 is activated in a very similar fashion to human PAR-1 with a thrombin cleavage site within the extracellular amino terminus LPIK38↓T39FRGAP [15]. Notably, mouse PAR-3 does not signal upon thrombin cleavage but functions instead via a unique cofactoring mechanism to support the activation of PAR-4 [16]. Human PAR-4, about 33% homologous to the other human PARs, is a 385-amino-acid protein with a potential cleavage site for thrombin and trypsin in the extracellular amino terminal domain PAPR47↓G48 YPGQV [17].
The novel activation mechanism distinguishes PARs from all other GPCRs though they share basic structural features. The general mechanism by which proteases cleave and activate PARs is similar: proteases cleave at specific sites within the extracellular amino terminus of the receptors; this cleavage exposes a new amino terminus, a cryptic N-terminal domain that serves as a “tethered ligand” domain, which binds to conserved region in the second extracellular loop of the cleaved receptor, and thereby activates the cleaved receptor. Synthetic peptides corresponding to the sequence of the “tethered ligand” are capable of activating the receptor independently of N-terminal proteolysis, confirming the self-activation model and providing a useful experimental tool for the specific activation of PARs [18] (Figure 1). These peptides include PAR-1 agonists SFLLR-NH2 and TFLLRN-NH2; PAR-2 agonists SLIGKV-NH2 and transcinnamoyl- (tc-) LIGRLO-NH2; PAR-3 agonist TFRGAP-NH2; and PAR-4 agonist GYPGQV-NH2. Activation of PARs results in a multiple cellular signaling events including cell shape, secretion, integrin activation, metabolic responses, transcriptional responses, and cell motility. PARs are “single use” receptors meaning that proteolytic activation is irreversible and the cleaved receptors are degraded in lysosomes [19].
Figure 1.
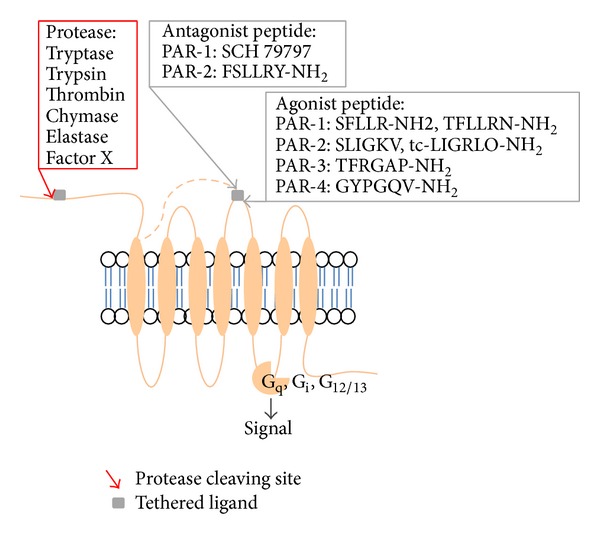
Activation mechanism of protease activated receptor (PAR). PARs are a group of four G protein-coupled receptors (GPCRs). Activation of PARs depends on the protease cleavage at the specific site of the extracellular N-terminal, upon which the exposed tethered ligand (gray square) binds to the second extracellular loop of PAR resulting in a series of cellular signaling events. The red arrow indicates cleavage site. The site for PAR-1 is at LDPR41↓S42FLLRN, PAR-2 is at SKGR34↓S35SLIGKV, PAR-3 is at LPIK38↓T39FRGAP, and PAR-4 is at PAPR47↓G48 YPGQV. PAR-2 antagonist peptide: FSLLRY-NH2; SCH 79797: a PAR-1 antagonist; the active peptides were PAR-1: SFLLR-NH2, TFLLRN-NH2; PAR-2: SLIGKV-NH2, transcinnamoyl- (tc-) LIGRLO-NH2; and PAR-3: TFRGAP-NH2 PAR-4, GYPGQV-NH2. S = Ser, F = Phe, L = Leu, R = Arg, T = Thr, I = Ile, G = Gly, K = Lys, V = Val, O = Pyl, A = Ala, P = Pro, Y = Tyr, Q = Gln, and V = Val.
3. Expression of PARs on Inflammatory Cells
Since inflammatory cells play a pivotal role in the pathogenesis of inflammation, what we should know first is the type and extent of PARs expression on inflammatory cells. As shown in Table 1, different PARs are expressed on mast cell, eosinophil, neutrophil, monocyte, macrophage, T cell, B cell, and dendritic cell (DC).
Table 1.
Expression of protease activated receptors (PARs) on inflammatory cells.
| Cell type | Expression of PARs | Regulator of PARs | ||||
|---|---|---|---|---|---|---|
| PAR-1 | PAR-2 | PAR-3 | PAR-4 | Upregulation | Downregulation | |
| MC | + [22, 27] |
+ [22, 23, 25, 27, 28] |
+ [22] |
+ [22, 28] |
RANTES for PAR-1 [32]; TNF-α for PAR-2 [23, 31] and PAR-4 [31]; IL-12 for PAR-4 [20]; GM-CSF for PAR-4 [30]; AC allergen rPer a1.01 for PAR-1,2,4 [22]. | IL-29 for PAR-1 [29]; IL-12 for PAR-2 [20]. |
|
| ||||||
| Eos | + [35, 36] |
+ [35–37] |
+ [36] |
− [36] |
na | na |
|
| ||||||
| Neu | + [38] |
+ [38, 39, 106] |
− [38] |
− [38] |
na | Simvastatin and pravastatin for PAR-2 [106]; GC for PAR-2 [39] |
|
| ||||||
| Mon | + [40–42, 46, 47] |
+ [43–47] |
+ [41, 46, 47] |
+ [47] |
IFN-γ for PAR-1 [42]; thrombosis for PAR-1; APS for PAR-2 [46] | na |
|
| ||||||
| Mac | + [41, 48, 50] |
+ [41, 43, 48–51] |
+ [41, 50] |
+ [50] |
LPS for PAR-1, -2, -3, and -4 [50]; dietary FA for PAR-2 [51]; MMIF for PAR-2 [77]; GM-CSF for PAR-1, -2, and -3 [41]; smoke for PAR-1 [48] | IL-4 for PAR-1, -2, and -3 [41] |
|
| ||||||
| TC | + [52, 162] |
+ [53, 162] |
na | + [162] |
HIV for PAR-1 [52] | na |
|
| ||||||
| BC | + [54] |
+ [163] |
na | na | na | na |
|
| ||||||
| DC | + [55, 56] |
+ [43, 57] |
+ [56] |
na | GC for PAR-2 [57] | LPS for PAR-1 and -3 [56] |
MC: mast cell; Eos: eosinophil; Neu: neutrophil; Mon: monocyte; Mac: macrophage; TC: T cell; BC: B cell; DC: dendritic cell; RANTES: regulated upon activation normal T cell expressed and secreted; TNF: tumor necrosis factor; GM-CSF: granulocyte-macrophage colony-stimulating factor; IL: interleukin; IFN: interferon; APS: antiphospholipid syndrome; MMIF: macrophage migration inhibitory factor; LPS: lipopolysaccharides; FA: fatty acids; HIV: human immunodeficiency virus; AC: American cockroach; GC: German cockroach; na: not available.
3.1. Expression of PARs on Mast Cells
It has been demonstrated that mast cells express all four PARs at both mRNA and protein levels. For example, PAR-1, -2, -3, and -4 are expressed on mouse mast cell line P815 cells [20–22] and MC/9 cells [20]. In addition, PAR-2 [23–25] and PAR-4 [24] are observed on the surface of human mast cell line HMC-1, and PAR-1 is found on murine bone marrow cultured mast cells [26]. The studies on human specimens show the presence of PAR-1 and PAR-2 in mast cells from various normal human tissues [27]; expression of PAR-2 and PAR-4 in mast cells both at protein and mRNA levels in the patients with postinfectious irritable bowel syndrome (PI-IBS) [28]; upregulated expression of PAR-2 in mast cells from ulcerative colitis tissues [25]; and the increased fraction of PAR-2-expressing mucosal mast cells in Crohn's specimens [23] (Table 1).
The expression of PARs appears to be regulated by cytokines. For example, interleukin- (IL-) 12 is able to downregulate PAR-2 expression [20], and IL-29 decreases PAR-1 expression [29] on P815 mast cells. On the other hand, granulocyte-macrophage colony-stimulating factor (GM-CSF) enhances expression of PAR-4 [30], tumor necrosis factor (TNF) [31] upregulates PAR-2 and PAR-4 expression, and RANTES increases expression of PAR-1 on P815 mast cells [32]. TNF also elevates PAR-2 expression on HMC-1 mast cells [23], and IL-12 upregulates PAR-4 expression on MC/9 mast cells [20]. Furthermore, tryptase-induced PAR-2 expression can be enhanced by IL-12 [20], TNF [31], RANTES [32], and IL-29 [29], and tryptase-induced PAR-3 and PAR-4 expression is upregulated by TNF on P815 mast cells [31]. It is also observed that trypsin-induced expression of PAR-1, -2, and -4 can be enhanced by RANTES [32] (Table 1).
3.2. Expression of PARs on Basophils
Little information on PAR expression on basophil is available. But a study showed lack of PAR expression on purified human basophils [33], and a report of the fact that trypsin rather than PAR-2 agonist induces histamine release from basophils [34] suggests that basophils may not express PARs.
3.3. Expression of PARs on Eosinophils
The expression of PAR-1 [35] and PAR-2 [35–37] has been observed on eosinophils. PAR-2 seems to be the major PAR receptor capable of modulating eosinophil functions [35]. It was reported that total numbers of eosinophils and the level of eosinophil expressing PAR-2 were significantly elevated in the nasal mucosa of seasonal allergic rhinitis (SAR) compared with the controls [37].
3.4. Expression of PARs on Neutrophils
Human peripheral blood neutrophils express PAR-1 and PAR-2, but not PAR-3 and PAR-4, proteins [38]. PAR-2 expression has also been observed on mouse pulmonary neutrophils [39].
3.5. Expression of PARs on Monocytes
The expression of PAR-1 [40–42], -2 [43–45], -3 [46], and -4 [47] on human monocytes is observed at both mRNA and protein levels. Interferon-gamma (IFN-γ) differentiated monocytes have increased expression of PAR-1 [42]. Monocyte surface PAR-2 expression is upregulated following static exposure to activated endothelial cell (EC) [44] (Table 1).
3.6. Expression of PARs on Macrophages
PAR-1, -2, and -3 expression has been showed in human macrophages [48, 49]. In Wistar rats, the expression of PAR-1, -2, -3, and -4 is revealed in airway macrophages (AMs) [50]. PAR-2 expression on human macrophages can be upregulated by dietary fatty acids (palmitic, stearic, and myristic) [51], but expression of PAR-1, -2, and -3 can be downregulated by IL-4 treatment [41]. Differentiation of human monocytes into macrophages by either macrophage colony-stimulating factor (M-CSF) or GM-CSF elicits enhanced expression of PAR-1, PAR-2, and PAR-3 [41]. There is a higher degree of PAR-1 protein staining in AMs from smokers compared with healthy controls (HC).
3.7. Expression of PARs on T Cells and B Cells
It has been showed that peripheral blood effector memory CD4(+) and CD8(+) T lymphocytes express PAR-1 [52] and that Jurkat T cells express PAR-2 [53]. In normal B cells, the expression of PAR-1 and PAR-3 is also reported [54].
3.8. Expression of PARs on DCs
It has been reported that monocyte-derived dendritic cells (MoDCs) do not express PARs [55, 56]. However, upon maturation with lipopolysaccharides (LPS), but not with TNF-α or CD40 ligand, DCs express PAR-1 and PAR-3 (not PAR-2 or PAR-4) [56]. IL-4 strongly downregulates PAR-1, -2, and -3 at both mRNA and protein levels in MoDCs [41]. Plasmacytoid DCs (pDCs) and myeloid DCs (mDCs) isolated from peripheral blood mononuclear cells (PBMC) express PAR-1 [55] and PAR-2 [57]. PAR-2 expression on mDCs is upregulated following German cockroach (GC) frass exposure [57].
4. Expression of PARs on Structural Cells
It has long been recognized that structural cells do have the ability to secrete proinflammatory mediators and cytokines, through which they actively participate in the pathogenesis of inflammation. More importantly, tissue remodeling processes in inflammation largely depend on structural cells and proteases. We therefore review the expression of PARs on epithelial cell, endothelial cell, fibroblast, smooth muscle cell (SMC), and keratinocyte in this section (Table 2).
Table 2.
Expression of protease activated receptors (PARs) on structural cells.
| Cell type | Expression of PARs | Regulator of PARs | ||||
|---|---|---|---|---|---|---|
| PAR-1 | PAR-2 | PAR-3 | PAR-4 | Upregulation | Downregulation | |
| EpC | + [50, 101] |
+ [50, 60, 101, 102] |
+ [50, 101] |
+ [50] |
NE for PAR-2; LPS for PAR-1, -2, -3, and -4 [50] | na |
|
| ||||||
| EnC | + [50, 62, 64–66, 100] |
+ [50, 61, 64] |
+ [50, 54, 66, 67] |
+ [50, 66] |
LPS for PAR-1, -2, -3, and -4 [50]; HCMV for PAR-1, -3, and -4 [66]; MMIF for PAR-1 and -2 [64]; TNF-α, IL-1α, and LPS for PAR-2 [63] | Phorbol ester for PAR-2 [63]; sheer stress for PAR-1 [62] |
|
| ||||||
| Fibro | + [68–71, 164] |
+ [53, 68–71, 164] |
+ [68, 70, 71] |
na | Malignancy for PAR-1 and -2 [164]; bFGF for PAR-2 [71] | PGE2 for PAR-2 [72] |
|
| ||||||
| SMC | + [73, 75] |
+ [74] |
+ [75] |
+ [75] |
na | PGI2/PGE2 for PAR-1, -3, and -4 [75] |
|
| ||||||
| Kerat | + [76, 82] |
+ [11, 76, 82, 103, 104] |
+ [76] |
na | na | na |
EpC: epithelial cell; EnC: endothelial cell; Fibro: fibroblast; SMC: smooth muscle cell; Kerat: keratinocyte; NE: neutrophil elastase; LPS: lipopolysaccharides; HCMV: human cytomegalovirus; MMIF: macrophage migration inhibitory factor; TNF: tumor necrosis factor; IL: interleukin; bFGF: basic fibroblast growth factor; PG: prostaglandin; na: not available.
4.1. Expression of PARs on Epithelial Cells
The A459 and BEAS-2B epithelial cell lines and primary human bronchial epithelial cells (HBECs) express PAR-1, -2, -3, and -4 as judged by RT-PCR and immunocytochemistry [58]. Both PAR-1 and PAR-2 are endogenously expressed in HBECs-16HBE14o-cells [59]. While PAR-1, -2, -3, and -4 expression is showed in alveolar epithelial cells of Wistar rats [50], PAR-2 appears to be a cellular receptor expressed prominently on epithelial cells of mice [60].
4.2. Expression of PARs on Endothelial Cells
Numerous studies have showed that PAR-2 is highly expressed on endothelial cells [50, 61, 62]. Stimulation with TNF-α, IL-1 α, and bacterial LPS elevates the expression of PAR-2 in a dose-dependent manner in cultured human umbilical vein endothelial cells (HUVECs) [63]. Macrophage migration inhibitory factor (MMIF) is found to enhance PAR-1 and PAR-2 mRNA expression in human endothelial cells [64], whereas phorbol ester treatment seems to decrease the expression of PAR-1 and PAR-2 [63]. In human endothelial cells, PAR-1 and β-arrestins form a preassembled complex [65]. Moreover, human cytomegalovirus (HCMV) induces expression of PAR-1 and PAR-3 but not PAR-4 proteins on HUVECs [66]. Since PAR-3 is postulated to represent a second thrombin receptor, its modest endothelial cell and platelet expression suggest that PAR-3 activation by alpha-thrombin is less relevant to physiological responses in these mature cells [67].
4.3. Expression of PARs on Fibroblasts
Human primary bronchial fibroblasts (HPBFs) express PAR-1, -2, and -3 but not PAR-4, and gingipains-induced secretion of hepatocyte growth factor (HGF) is significantly inhibited by RNA interference targeted at PAR-1 and PAR-2 [68, 69]. Similarly, both normal and fibrotic human lung fibroblasts express PAR-1, -2, and -3. There is no significant difference between normal and fibrotic fibroblasts in expression levels of PAR-1 and -3, whereas a fourfold higher expression level of PAR-2 is observed in fibrotic cells compared with normal cells [70]; in addition an important role of basic fibroblast growth factor (bFGF) in the regulation of functional PAR-2 expression in cultured RA synovial fibroblasts was reported [71]. It is reported that prostaglandin (PG) E2, via the prostanoid receptor EP2 and subsequent cAMP elevation, downregulates mRNA and protein levels of PAR-1 in human lung fibroblasts [72] and that PAR-2 upregulation by TNF-α may modulate myofibroblast proliferation [23].
4.4. Expression of PARs on SMCs
Human airway SMCs express PAR-1 and PAR-2 proteins [73, 74]. PAR-1, -3, and -4 are able to mediate thrombin-induced proliferation, migration, and matrix biosynthesis as well as generation of inflammatory and growth-promoting mediators in human vascular SMCs [75].
4.5. Keratinocytes
It is reported that TERT-2 cells constitutively express high levels of PAR-1 and PAR-2 and lower level of PAR-3 [76], whereas human keratinocytes express PAR-2 [11], which can be upregulated by MMIF [77].
5. Signal Transduction Pathways of PARs
As shown in Figure 2, depending on the PAR subtype and the phenotype of PAR-expressed cell, the PAR family is able to stimulate a variety of intracellular signaling pathways. Like other “GPCRs,” the PARs signal via a variety of G proteins: both PAR-1 [78] and PAR-2 [79] through Gα q, Gα i, Gα 12/13, and Gβ γ; PAR-4 via Gα q and Gα 12/13 [78]; and PAR-3 via PAR-1 signaling by receptor dimerization. PAR-1 heterodimerization with PAR-3 alters the PAR-1/Gα 13 binding conformation, enhancing Gα 13 signaling. Heterodimerization does not likely affect PAR-1/ Gα q selectivity [80]. In addition, PAR-2 is able to signal via a non-G-protein mechanism that involves the beta-arrestin-mediated signaling [81]. Particularly in HMC-1 cell line, it was found that curcumin inhibits PAR-2 and PAR-4 mediated human mast cell activation, not by inhibition of trypsin activity but by the blocking of extracellular signal-regulated kinase (ERK) pathway [24], and thrombin mediates PAR-1 mediated mast cell adhesion through the activation of G(i) proteins, phosphoinositol 3-kinase, protein kinase C, and mitogen-activated protein kinase (MAPK) pathways [26].
Figure 2.
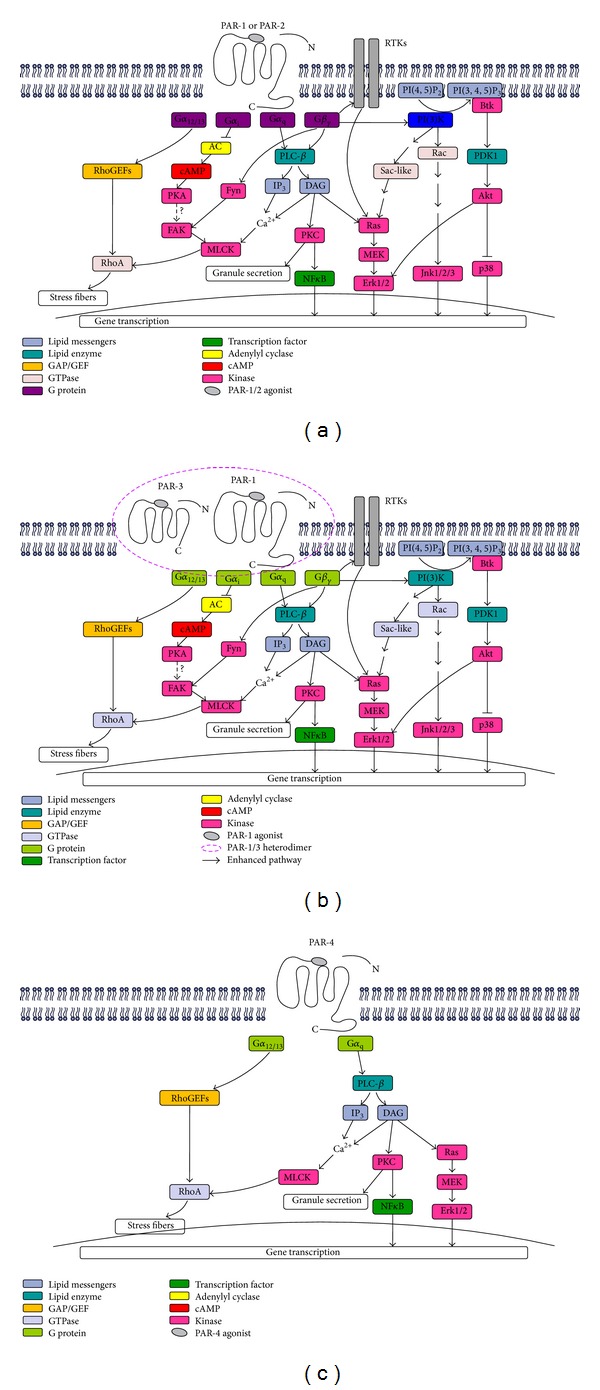
(a) Signal transduction pathways for PAR-1 and PAR-2. (b) Signal transduction pathways for PAR-1 and PAR-3. (c) Signal transduction pathways for PAR-4.
Proteases mediating PAR-1 and PAR-2 activation differentially signal via MAPK cascades. In addition, the production of chemokines induced by PAR-1 and PAR-2 activation is suppressed by PI3K/Akt, thus keeping the innate immune responses of human oral keratinocytes in balance [82]. The CUX1 homeodomain protein is a downstream effector of PAR-2. Treatment of epithelial and fibroblastic cells with trypsin or the PAR-2 agonist peptide (AP) causes a rapid increase in CUX1 DNA binding activity. The stimulation of CUX1 was specific to PAR-2 because no effect was observed with thrombin or the PAR-1 AP. These results suggest a model whereby activation of PAR-2 triggers a signaling cascade that culminates with the stimulation of p110 CUX1 DNA binding and the transcriptional activation of target genes [83]. Thrombin induces RPE cell proliferation by joint activation of PLC-dependent and atypical PKC isoforms and the Ras-independent downstream stimulation of the Raf/MEK/ERK1/2 MAPK cascade [84].
6. Actions of Agonists and Antagonists of PARs in Inflammation
Since PARs are receptors expressed on various types of cells, their actions must be triggered or inhibited by agonists and antagonists of PARs. Hence, we review actions of agonists and antagonists of PARs in inflammation in the current section.
6.1. Influence of Agonists and Antagonists of PARs on Inflammatory Cell Migration
A significant increase in PAR-2 expression is observed on cell surface of neutrophils from septic patients as compared with HC. PAR-2 agonists (serine proteases as well as synthetic peptides) upregulate cell adhesion molecule expression and cytokine production and reduce transendothelial migration of neutrophils [85]. On the other hand, basolateral, but not apical, PAR-1 and PAR-2 activation with selective agonists decreases transepithelial resistance (TER) and thereby facilitates neutrophil transepithelial migration [86]. It is observed that activation of PAR-1 by thrombin stimulates directed migration of human eosinophils and thereby affects eosinophils in tissue and allergic inflammation [87].
6.2. Influence of Agonists and Antagonists of PARs on Cell Proliferation and Repair
Thrombomodulin (TM) acts as a thrombin receptor that modulates the duration of pERK nuclear retention and HUVEC proliferation in response to thrombin [88]. It has been found that thrombin and a PAR-1 APs stimulate proliferation of HPBFs [68] and that thrombin and FoxO factors functionally interact through PI3K/Akt-dependent FoxO phosphorylation leading to vascular SMC proliferation [89]. Furthermore, activated protein C (APC) acts through both PAR-1 and PAR-2 to activate Akt and to increase keratinocyte proliferation [90]. In mechanically wounded 16HBE 14o(-) epithelial cell layers in culture, PAR-1 and PAR-2 APs stimulate the rate of repair and enhance the formation of a fibrin provisional matrix to support the repair process. Locally expressed serine proteases of the coagulation cascade activate PAR-1 and PAR-2 to enhance fibrin formation and bronchial epithelial repair [91].
6.3. Influence of Agonists and Antagonists of PARs on Mediator and Cytokine Release from Cells
It has been observed that thrombin, tryptase, elastase, and trypsin, as well as APs of PAR-1, -2, and -4, induce IL-8 [92] and MCP-1 [93] release from A549 cells, suggesting that the actions of thrombin and trypsin may be via PAR-1 and -4, and cell responses to tryptase, trypsin, and elastase may be through PAR-2. Thrombin, trypsin, tryptase, elastase, SFLLR-NH2, and GYPGQV-NH2 stimulate also IL-6 release from monocytes [94]. A rank order of potency of the APs corresponding to the nascent N-termini of PAR-1, -2, and -4 appears as PAR-2 > PAR-4 > PAR-1 for induction of the release of IL-6 and IL-8 from A549, BEAS-2B, and HBECs. The APs of PAR-1, -2, and -4 also cause the release of PGE2 from A549 and HBECs [58]. Moreover, it is noticeable that thrombin, trypsin, tryptase, SFLLR-NH2, and SLIGKV-NH2 are capable of eliciting IL-6 release from T cells [95] (Table 3).
Table 3.
(a) Influence of agonists of protease activated receptors (PARs) on mediator and cytokine release from cells. (b) Influence of antagonists of protease activated receptors (PARs) on mediator and cytokine release from cells.
(a)
| Agonist | Targeted cell | Response of cell |
|---|---|---|
| PAR-1 | ||
| Thrombin | PBMC [47]; A549 [92, 93]; Mon [94]; TC [95]; HPDF [96, 97]; CHAEC [98, 99]; EnC [54] | IL-1β [47], IL-6 [47, 94, 95], IL-8 [92, 96], MCP-1 [93], MMP-9 [97], MMP-12 [99], VCAM-1 [54], VEGF [98] |
| Trypsin | A549 [92, 93]; Mon [94]; TC [95] | Release of IL-8 [92]; MCP-1 [93]; IL-6 [94, 95] |
| SFLLR-NH2 | A549 [58, 92, 93]; Mon [94]; HBEC [58]; TC [95]; CHAEC [98, 99] | IL-8 [92]; MCP-1 [93]; IL-6 [94, 95]; PGE2 [58]; VEGF [98]; MMP-12 [99] |
|
| ||
| PAR-2 | ||
| Tryptase | A549 [92, 93]; Neu [38]; Mon [94]; TC [95]; Astr [111] | Release of IL-8 [38, 92]; MCP-1 [93]; IL-6 [94, 95, 111]; LF [38];TNF-α, ROS [111] |
| Trypsin | A549 [92, 93]; Mon [94]; HPBF [68]; Eos [35]; MC [107]; HUVEC [108] | Release of IL-8 [68, 92, 108]; MCP-1 [93]; IL-6 [94]; G-CSF, VCAM-1 [68]; sys-LT, ROS [35]; tryptase [107] |
| Elastase | A549 [92, 93]; AEC [101]; Mon [94] | Release of IL-8 [92]; MCP-1 [93]; IL-6 [94]; MUC5AC [101] |
| GCE | MH-S [49]; RAW264.7 [49] | TNF-α [49] |
| WCE | PHKC [104] | Release of IL-8, GM-CSF [104] |
| SLIGKV-NH2 | A549 [58, 92, 93]; HPBF [68]; HBEC [58]; TC [95]; HTEC [102]; Kerat [103]; Neu [38]; Eos [35, 36]; MC [107]; BEAS-2B, Calu-3 [109] | Release of IL-8 [38, 68, 92, 109]; MCP-1 [93]; PGE2 [58]; IL-6 [95, 109]; G-CSF [68]; VCAM-1 [68, 102]; GM-CSF [102, 109]; TNF-α [102]; MMP-1,10 [102]; TSLP [103]; LF [38]; sys-LT, ROS [35]; SOD [36]; His [107] |
| tc-LIGRLO-NH2 | Neu [38]; MC [107] | Release of IL-8, LF [38]; His [107] |
|
| ||
| PAR-3 | ||
| Thrombin | A549 [92, 93]; Mon [94]; EnC [54] | Release of IL-8 [92]; MCP-1 [93]; IL-6 [94]; ICAM-1, VCAM-1 [54] |
|
| ||
| PAR-4 | ||
| Thrombin | A549 [92, 93]; Mon [94] | Release of IL-8 [92]; MCP-1 [93]; IL-6 [94] |
| GYPGQV-NH2 | A549 [58, 92, 93]; Mon [94]; HBEC [58] | IL-8 [92]; MCP-1 [93]; IL-6 [94]; PGE2 [58] |
(b)
| Antagonist | Targeted cell | Response of cell |
|---|---|---|
| PAR-1 | ||
| SCH 79797 | HUVEC [165]; HDF [166]; MC [29] | Block thrombin induced CX3CL1 [165]; inhibit plasmin-induced IL-8, PGE [166]; inhibit trypsin and tryptase induced IL-4 release [29] |
| BMS 200261 | PL [114] | Inhibit PAR-1 activation in response to thrombin [114]; |
| RWJ-56110 [122, 167]; RWJ-58259 [123] | HSC [167]; HVC, PL [112, 113] | Reduced liver type I collagen [167], reduced liver calcium mobilization, and cellular functions (platelet aggregation and cell proliferation [112, 113]) |
|
| ||
| PAR-2 | ||
| FSLLRY-NH2 | HUVEC [108]; Astr [111]; MC [20, 29, 31, 32] | Inhibit trypsin-induced IL-8 [108]; alleviate IL-6 and TNF-α secretion [111]; inhibit trypsin and tryptase induced IL-4 release [20, 29, 31]; inhibit tryptase induced IL-13 release [32] |
| ENMD-1068 | Eos [168] | Abolish tryptase-induced eosinophil recruitment [168] |
| GB88 | HTEC [102]; HT29, A549, Panc-1, MKN1, MKN45, MDA-MB231, HUVEC [117]; Mac, MC [169] | Block PAR-2 agonist-induced increases in GM-CSF, IL-6, IL-8, and TNF-α [102]; ameliorate macrophage invasion and MC degranulation [169]; inhibit PAR-2 activated Ca(2+) release induced by trypsin or synthetic peptide and nonpeptide agonists [117] |
|
| ||
| PAR-4 | ||
| Pepducin P4pal-10 | Neu [115] | Diminish neutrophil migration mediated by neutrophil expressed PAR-4 [115] |
PBMC: peripheral blood mononuclear cells; A549: A549 epithelial cells; Mon: monocyte; TC: T cell; HPDF: human primary dermal fibroblast; CHAEC: cultured human airway epithelial cell; EnC: endothelial cell; HBEC: human bronchial epithelial cell; Neu: neutrophil; Astr: astrocytes; HPBF: human primary bronchial fibroblasts; Eos: eosinophil; MC: mast cell; HUVEC: human umbilical vein endothelial cell; AEC: airway epithelial cells; MH-S: alveolar macrophage cell lines; RAW264.7: peritoneal macrophage cell lines; PHKC: primary human keratinocyte; HTEC: kidney tubular epithelial cells; Kerat: keratinocyte; BEAS-2B: human airway epithelial cell line BEAS-2B; Calu-3: human airway epithelial cell line Calu-3; HDF: human dental pulp fibroblast-like cells; PL: platelet; HSC: hepatic stellate cell; HVC: human vascular cell; HT29: human colon adenocarcinoma cell line; Panc-1: pancreas epithelioid carcinoma line Panc-1; MKN1: human gastric carcinoma cell line MKN1; MKN45: human gastric carcinoma cell line MKN45; MDA-MB231: breast adenocarcinoma cell line MDA-MB231; Mac: macrophage; IL: interleukin; MCP-1: monocyte chemoattractant protein-1; MMP: matrix metalloproteinase; VCAM: vascular cell adhesion molecule; VEGF: vascular endothelial growth factor; PG: prostaglandin; LF: lactoferrin; TNF: tumor necrosis factor; ROS: reactive oxygen species; G-CSF: granulocyte colony-stimulating factor; cys-LT: cysteinyl leukotrienes; MUC5AC: mucin5AC; GM-CSF: granulocyte-macrophage colony-stimulating factor; TSLP: thymic stromal lymphopoietin; SOD: superoxide; His: histamine; ICAM: intercellular adhesion molecule.
6.3.1. Agonists of PAR-1
Thrombin is able to mediate induction of IL-1β and IL-6 cytokine production from PBMCs and PBMC cell proliferation in a PAR-1-dependent manner [47]. Thrombin can also induce IL-8 [96] and matrix metalloprotease- (MMP-) 9 release [97] from human primary dermal fibroblasts (HPDF) through activation of PAR-1. Similarly, thrombin and PAR-1 APs significantly stimulate vascular endothelial growth factor (VEGF) secretion from cultured human airway epithelial cells (HAEC) [98] and MMP-12 release from peritoneal macrophages [99]. A very recent study shows that MMP-1 causes activation of the nuclear factor-κB (NF-κB) pathway (p65/RelA) in endothelial cells, and this response is dependent upon activation of PAR-1 [100] (Table 3).
6.3.2. Agonists of PAR-2
In the alveolar macrophage cell lines (MH-S cells) and peritoneal macrophage cell lines (RAW264.7 cells), GC extract activates PAR-2 and thereby produces TNF-α. GC extract can also enhance TNF-α production by alveolar macrophages through the PAR-2 pathway [49]. The observation that trypsin and SLIGKV-NH2 are able to stimulate an increase in vascular cell adhesion molecule- (VCAM-) 1 expression and the release of IL-8 and granulocyte colony-stimulating factor (G-CSF) from bronchial fibroblasts suggests the importance of PAR-2 in promoting neutrophilic airway inflammation [68]. A finding demonstrates that neutrophil elastase can increase PAR-2 expression and MUC5AC mucin release [101] may also implicate the involvement of PAR-2 in airway inflammation. While treatment with SLIGKV-NH2 at the apical or basolateral cell surface of epithelial cells induces GM-CSF, ICAM-1, TNF-α, MMP-1, and MMP-10 secretion [102], PAR-2 agonist also provokes the production of thymic stromal lymphopoietin (TSLP) in the skin of mice [103]. Since these mediators and cytokines are important promoters of inflammation, PAR-2 should be a pivotal contributor of inflammation. Moreover, it is reported that mite-derived serine protease activity may contribute to the pathogenesis of atopic dermatitis (AD) by activating keratinocytes via PAR-2 activation [104]. Allergen-induced, PAR-2/epidermal growth factor receptor- (EGFR-) mediated signaling may also decrease epithelial resistance and promotes junction disassembly [105], thereby promoting epithelial inflammation.
It has been showed that tryptase, trypsin, SLIGKV-NH2, tc-LIGRLO-NH2, but not thrombin, elastase, and SFLLR-NH2 can induce IL-8 and lactoferrin secretion from peripheral blood neutrophils [38], implicating that the actions of tryptase and trypsin are likely via the activation of PAR-2. Studies on mice show that neutrophils from antiphospholipid (aPL) antibody-treated mice express PAR-2 and that stimulation of this receptor leads to neutrophil activation [106]. Antineutrophil cytoplasmic antibodies against proteinase 3 (PR3) activate human monocytic THP-1 cells in a PR3- and PAR-2-dependent manner [45].
Human eosinophils express PAR-1 and -2, and PAR-2 is the major PAR receptor that is capable of modulating eosinophil function. Trypsin and the PAR-2 APs are seen in triggering shape change, release of cysteinyl leukotrienes, and, most obviously, generation of reactive oxygen species in eosinophils [35]. A study demonstrates that a specific tethered peptide ligand for PAR-2 potently induces superoxide production and degranulation may support the finding above [36]. It is reported that SLIGKV-NH2 and tc-LIGRLO provoke histamine release from skin mast cells and that trypsin is able to induce a bell shape increase in tryptase release from tonsil mast cells [107]. In HUVEC, trypsin-induced IL-8 release is inhibited by the inhibitor peptide of PAR-2 Phe-Ser-Leu-Leu-Arg-Tyr-NH2 (FSLLRY-NH2), suggesting that the action of trypsin on HUVEC is most likely through activation of PAR-2 [108]. Very recently, it is reported that PAR-3 cooperates with PAR-1 to mediate the effect of thrombin on cytokine production and VCAM-1 expression in endothelial cells, but ICAM-1 expression in endothelial cells requires PAR-3 without PAR-1 [54]. It has been reported that aspartate proteases from Alternaria induce GM-CSF, IL-6, and IL-8 production and calcium response in airway epithelium through PAR-2 [109] and that Alternaria-derived aspartate proteases cleave PAR-2 to activate eosinophil degranulation [110], which may add another novel mechanism for activation of PARs in the development and exacerbation of airway allergic diseases (Table 3). We have recently found that tryptase can induce the release of IL-6 and TNF-α from astrocytes via PAR-2-MAPKs or PAR-2-PI3K/Akt signaling pathway, which reveals PAR-2 as a new target actively participating in the regulation of astrocytic functions [111].
6.3.3. Antagonists of PARs
Potent heterocycle-based peptide-mimetic antagonists of PAR-1, RWJ-56110 [112], and RWJ-58259 [113] are potent, selective PAR-1 antagonists, which bind to PAR-1, interfere with calcium mobilization and cellular functions (platelet aggregation and cell proliferation), and do not affect PAR-2, -3, or -4. Both PAR-1 and PAR-4 activation peptides were significantly inhibited by affinity-purified anti-PAR-1-IgY and anti-PAR-4-IgY and by the specific PAR-1 antagonist BMS 200261 [114]. A cell-penetrating pepducin antagonist of PAR-4 (P4pal-10) dose-dependently diminishes the severity of endotoxemia and preserves liver, kidney, and lung function of mice, suggesting that inhibition of PAR-4 signaling in neutrophils could be protective in systemic inflammation and DIC [115]. Using a fluorescence assay, a novel compound, GB88, is shown to antagonize PAR-2-induced intracellular Ca(2+) release in human monocyte-derived macrophages, being 1000 times more potent than a novel small molecule PAR-2 antagonist N1-3-methylbutyryl-N4-6-aminohexanoyl-piperazine (ENMD-1068). GB88 inhibits also the acute paw edema induced in Wistar rats by PAR-2 agonist 2-furoyl-LIGRLO-NH2 or mast cell β-tryptase, without inhibiting proteolytic activity of tryptase in vitro [116, 117].
7. Roles of PARs in Allergic Diseases
Accumulated evidence suggests that PARs are closely related to allergic inflammation, but the detailed relationship between PARs and allergic diseases remains obscure. We therefore review the known roles of PARs in allergic diseases in the current section (Table 4).
Table 4.
Roles of PARs in allergic diseases.
| Disease | PAR involved | Response of cell or molecule |
|---|---|---|
| Rhinitis | PAR-1 | VEGF secretion from CHAEC [98]; |
| PAR-2 | Higher secretion rate and numbers of responding glands [122]; fluid hypersecretion of airway mucosa [123]; suppression of Cx26 production in human NEC [124]; IL-6 and IL-8 production in NECs [125]; tachykinin-mediated neurogenic inflammation [126]; | |
|
| ||
| Asthma | PAR-1 | Bronchial inflammation is worsened [118]; expression of TGF-β1 to promote airway remodeling [119]; susceptible to AHR [120] |
| PAR-2 | Contractions of human airways [128]; bronchial SMC proliferation and migration [129]; increased anion secretion [130]; eosinophil infiltration, AHR, IgE levels to OVA sensitization [131] influx of eosinophils in BALF, protein leak in the bronchoalveolar space, increase BALF levels of the anaphylatoxins C3a and C5a [132]; increase IL-6 expression and induce the proliferation of asthmatic bronchial SMC [134]; augment TNF-α-induced MMP-9 expression [135]; generate Ca(2+) in HAECs [136, 138]; AHR, Th2, and Th17 cytokine release, serum IgE levels, and cellular infiltration [137]; CCL20 and GM-CSF production, increase recruitment and/or differentiation of mDC populations [139]; beta-adrenergic desensitization [140]; increase chemotactic activity for the HMC-1 mast cell line [141]; exaggerate cough [142]; chitinase-mediated [Ca(2+)] increase [13]; inhibit the development of airway eosinophilia and hyperresponsiveness in allergic mice through COX-2-mediated generation of the anti-inflammatory mediator PGE2 [143]; inhibit bronchoconstriction and airway hyperresponsiveness [144] | |
|
| ||
| Skin disorders | PAR-2 | Production of the TSLP and TNF-α [146]; modulation of the calcium ions in skin [147]; scratching behavior in mice [148, 149]; ear edema and infiltration of inflammatory cells [150] |
|
| ||
| Colitis | PAR-2 | Inflammatory process in the intestinal mucosa [25]; impaired epithelial barrier [151]; somatic and visceral hyperalgesia and allodynia [152]; relaxation in colonic smooth muscle [153]; cytoskeleton contraction with subsequent changes in tight junction permeability [154]; visceral hypersensitivity [155] |
| PAR-4 | Contraction of the longitudinal muscle of colon [156]; antinociceptive [158]; increase paracellular permeability and myeloperoxidase activity [159, 160]; colonic hyposensitivity [157, 161] | |
VEGF: vascular endothelial growth factor; CHAEC: cultured human airway epithelial cell; Cx: connexin; NEC: nasal epithelial cell; TGF: transforming growth factor; AHR: airway hyperresponsiveness; SMC: smooth muscle cell; BALF: bronchoalveolar lavage fluid; TNF: tumor necrosis factor; HAEC: human airway epithelial cells; GM-CSF: granulocyte-macrophage colony-stimulating factor; mDC: myeloid DC; TSLP: thymic stromal lymphopoietin; CCL20: chemokine C-C motif ligand 20; PGE2: prostaglandin E2.
7.1. Roles of PAR-1 in Allergic Diseases
7.1.1. In Allergic Rhinitis (AR)
It has been reported that thrombin is increased in nasal secretion of the patients with chronic rhinosinusitis and thrombin and PAR-1 APs stimulate VEGF secretion from cultured HAEC [98], suggesting that thrombin may play a role in nasal polyp formation by stimulating VEGF production from airway epithelial cells.
7.1.2. In Asthma
Heterozygous PAR-1 mice have less allergic inflammation but PAR-1 agonist worsens it. Allergic bronchial inflammation is worsened in mice that receive adoptive transfer of PAR-1 agonist-treated Th2 cells compared to controls. Low concentrations of thrombin suppress but high dose of it enhances maturation and secretion of cytokines in DCs [118]. The expression of PAR-1 is upregulated by thrombin that induces the expression of TGF-β1 to promote airway remodeling in OVA-allergic rats [119]. An integrated effect is also observed in one haplotype cluster consisting of both regions of the EGFR gene and the PAR-1 gene, suggesting the possibility that the integrated effect of functionally related EGFR and PAR-1 genes (haplotype cluster) is associated with susceptibility to airway hyperresponsiveness (AHR) [120].
7.2. Roles of PAR-2 in Allergic Diseases
7.2.1. In AR
The expression levels of PAR-2 mRNA and immunoreactivity for PAR-2 in the nasal mucosa of AR are significantly upregulated as compared with normal nasal mucosa [121], suggesting that PAR-2 is very likely to be involved in allergic nasal inflammation. Indeed, it is found that house-dust mite (HDM) induces a higher secretion rate and numbers of responding glands in the AR group of patients than in the control group. Since PAR-2 is highly expressed in nasal mucosa, PAR-2 activating peptide provokes similar responses in nasal mucosa, and both HDM and PAR-2 activating peptide induced responses are suppressed by ENMD-1068; the involvement of PAR-2 in AR is confirmed [122]. In addition, HDM-induced PAR-2 activation and fluid secretion in porcine airway mucosa suggest a role for PAR-2 in mucociliary clearance and fluid hypersecretion of airway mucosa [123]. It is found that suppression of connexin 26 in HDM-sensitized AR patients is related to a PAR-2 mediated pathway and may be involved in the initiation and maintenance of AR [124]. Stimulation of elevated IL-6 and IL-8 production in primarily cultured nasal epithelial cells (NECs) by a major allergen of HDM Der p1 may also contribute to AR. Der p1 induced IL-6 and IL-8 production which seems to be associated with the PAR/PI3 K/NFκB signaling pathway [125].
Furthermore, the abundant presence of PAR-2 in different cell types including eosinophils and epithelial cells in the nasal mucosa suggests that PAR-2 may contribute to the pathogenesis of seasonal AR [37]. Based on the colocalization of PAR-2 and tachykinins in trigeminal sensory neurons innervating the nasal mucosa, it is suggested that, following an activation of PAR-2 in tachykinergic neurons by trypsin and mast cell tryptase, there may be a triggering of tachykinin-mediated neurogenic inflammation in allergic or nonallergic rhinitis [126].
7.2.2. In Asthma
It is observed that, in normal and asthmatic subjects, epithelial staining intensity of PAR-1 and PAR-3 is greater than that of PAR-4. However, PAR-2 staining in asthmatic epithelium is increased in comparison with normal epithelium [127], which suggests that PAR-2 may be involved in the pathogenesis of asthma. In fact, activation of PAR-2 has been found to induce contraction of human airways and potentiate contraction to histamine and therefore may contribute to asthma [128]. Through a PAR-2-dependent mechanism, chitinase 3-like 1 protein promotes bronchial SMC proliferation and migration [129]. Basolateral PAR-2 activation in the mouse airways leads to increased anion secretion through apical calcium-activated chloride channels, which is more pronounced in allergic animals [130]. Compared with wild-type animals, eosinophil infiltration is inhibited by 73% in mice lacking PAR-2 and increased by 88% in mice overexpressing PAR-2. Similarly, airway hyperreactivity to inhaled methacholine is diminished by 38% in mice lacking PAR-2 and increased by 52% in mice overexpressing PAR-2. PAR-2 deletion also reduces IgE levels to OVA sensitization. These results indicate that PAR-2 contributes to the development of immunity and to allergic inflammation of the airway [131].
HDM exposure to wild-type (Wt) mice causes a profound influx of eosinophils in bronchoalveolar lavage fluid (BALF) and accumulation of eosinophils in lung tissue. Both of them are strongly reduced in PAR-2 KO mice. PAR-2 KO mice demonstrate also attenuated lung pathology and protein leak in the bronchoalveolar space, accompanied by lower BALF levels of the anaphylatoxins C3a and C5a [132]. Through its serine protease activity, HDM potentiates capsaicin-evoked Ca(2+) transient in mouse pulmonary sensory neurons via PAR-2-phosholipase C-protein kinase C intracellular transduction cascade [133]. HDM extract also upregulates calreticulin (CRT) protein, activates PAR-2, increases IL-6 expression, and induces the proliferation of asthmatic bronchial SMC [134].
GC fecal remnants contain active serine proteases which augment TNF-α-induced MMP-9 expression by a mechanism involving PAR-2, ERK, and AP-1 [135]. GC extract has a direct effect on HAEC, in particular generating [Ca(2+)](i) oscillations through activation of PAR-2 [136]. PAR-2-deficient mice have significantly reduced AHR, Th2, and Th17 cytokine release, serum IgE levels, and cellular infiltration compared to Wt mice when sensitizaed to GC frass [137]. Alternaria proteases act through PAR-2 to induce rapid increases in human airway epithelial [Ca(2+)](i) in vitro and cell recruitment in vivo, the critical early steps in the development of allergic asthma [138]. Through the activation of PAR-2, allergen-derived proteases are sufficient to induce CCL20 (chemokine C-C motif ligand 20) and GM-CSF production in the airways leads to increased recruitment and/or differentiation of mDC populations in the lungs, which likely plays an important role in the initiation of allergic airway responses [139].
A report that tryptase and SLIGKV-NH2 act on airway smooth muscle and lead to homologous beta-adrenergic desensitization via PAR-2 activation [140] suggests a relatively novel mechanism involved in asthma. Supernatants of human SMCs treated with the major mast cell product tryptase have increased chemotactic activity for the HMC-1 mast cell line. The effect depends on an intact catalytic site for tryptase and can be induced by a peptide agonist for PAR-2, indicating that the action of tryptase is via PAR-2 activation [141]. Moreover, PAR-2 activation, by sensitizing the transient receptor potential vanilloid 1 (TRPV1) in primary sensory neurons, may play a role in the exaggerated cough observed in certain airways inflammatory diseases such as asthma and chronic obstructive pulmonary disease [142]. Desensitization of PAR-2 by repetitive agonist stimulation or siRNA-mediated PAR-2 knockdown reveals that chitinase-mediated [Ca(2+)](i) increase is exclusively mediated by PAR-2 activation. Chitinase is found to cleave the cleavage site of PAR-2 and enhance IL-8 production, indicating that exogenous chitinase is a potent proteolytic activator of PAR-2 in human airway epithelial cells [13]. In contrast, SLIGRL-NH2 has been found to inhibit the development of airway eosinophilia and hyperresponsiveness in allergic mice through COX-2-mediated generation of the anti-inflammatory mediator PGE2. SLIGRL also displays bronchodilator activity in allergic mice. These studies support the concept that PAR-2 exerts predominantly bronchoprotective actions within allergic murine airways [143]. It is also found that PAR-2 AP pretreatment is able to inhibit airway hyperresponsiveness and bronchoconstriction and to modulate the immune response induced by allergic challenge in sensitized rabbits [144].
7.2.3. In Allergic Skin Disorders
It has been reported that certain proteases signal to cells by activating PARs in the skin. Recent studies have revealed aberrant expression and activation of serine proteases and PAR-2 in the lesional skin of AD patients. Upregulated proteases stimulate PAR-2 and lead to the production of cytokines and chemokines involved in inflammation and immune responses, itching sensation, and sustained epidermal barrier perturbation with easier allergen penetration. In addition, PAR-2 is an important sensor for exogenous danger molecules, such as exogenous proteases from various allergens, and plays an important role in AD pathogenesis [145]. Defective skin barrier facilitates allergen and microbe penetration and generates danger signals leading to PAR-2 activation, which triggers the production of the major pro-Th2 cytokine TSLP and TNF-α [146]. It is also found that allergens with protease activity can influence the epidermal permeability barrier homeostasis through PAR-2 activation and consequent modulation of the calcium ions in skin [147]. PAR-2 may be involved in passive cutaneous anaphylaxis-induced scratching behavior in ICR mice [148]. Intradermal injections of histamine and SLIGRL-NH2 induce scratching in naive mice, but protease-associated itch and allergy-associated itch are different from those of histamine-induced itch [149]. It is reported that PAR-2 may also play a crucial role in type IV allergic dermatitis [150].
7.2.4. In Allergic Colitis
Increased levels of serine proteases activating PAR-2 are found in the lumen and colonic tissue of inflammatory bowel diseases (IBD) patients. PAR-2 activity and proinflammatory cytokines impair epithelial barrier, facilitating the uptake of luminal aggressors that perpetuate inflammation in IBD [151]. Trypsin and tryptase expression and release are increased in colonic biopsies from irritable bowel syndrome (IBS) patients compared with control subjects. Biopsies from IBS patients release mediators that sensitize murine sensory PAR-2 expressing neurons in culture. Supernatants from colonic biopsies of IBS patients also cause somatic and visceral hyperalgesia and allodynia in mice. These pronociceptive effects are absent in PAR-2-deficient mice, indicating that proteases released in IBS can directly stimulate sensory neurons and generate hypersensitivity symptoms through the activation of PAR-2 [152]. PAR-2 mediated relaxation system in colonic smooth muscle is suppressed in experimental colitis rat model, which may contribute to motility disorders in IBD [153].
Activation of PAR-2 on epithelial cells may directly affect cytoskeleton contraction by triggering phosphorylation of myosin light chain with subsequent changes in tight junction permeability [154]. Dexamethasone treatment improves PAR-2 agonist-induced visceral hypersensitivity but does not prevent PAR-2 agonist-induced increase in colonic permeability in rats [155]. It has been showed that PAR-2 is upregulated on ileal mucosal mast cells in Crohn's ileitis, which may contribute to perpetuating the inflammatory process in the intestinal mucosa in Crohn's ileitis [23].
It is reported that GB88 is a potent antagonist of PAR-2 activation in colonocytes. Acute colonic inflammation induced in rats by SLIGRL-NH2 is inhibited by oral administration of GB88 with markedly reduced edema, mucin depletion, PAR-2 receptor internalization, and mastocytosis. Chronic trinitrobenzenesulfonic acid-induced colitis in rats is ameliorated by GB88, which reduces mortality and pathology (including colon obstruction, ulceration, wall thickness, and myeloperoxidase release) more effectively than the clinically used drug sulfasalazine, suggesting the therapeutic potential for PAR-2 antagonist in inflammatory diseases of colon [116].
7.3. Roles of PAR-4 in Allergic Diseases
7.3.1. In Allergic Colitis
PAR-4 is functionally expressed in rat colon and its activation induces contraction of the longitudinal muscle both through tetrodotoxin sensitive release of acetylcholine and release of tachykinins, probably from sensory nerves [156]. PAR-4 agonists modulate colonic nociceptive response and inhibit colonic hypersensitivity and primary afferent responses to pronociceptive mediators. Endogenous activation of PAR-4 also plays a major role in controlling visceral pain. These results identify PAR-4 as a previously unknown modulator of visceral nociception [157]. The PAR-4 activation is endogenously involved as a feedback loop to attenuate inflammatory colonic hyperalgesia to colorectal distension [158]. Ulcerative colitis (UC) fecal supernatants, cathepsin G (Cat-G), and PAR-4 agonist can increase both paracellular permeability and myeloperoxidase activity [159]. Increased colonic paracellular permeability that is triggered by UC fecal supernatants can be blocked by both specific Cat-G inhibitor (SCGI) (77%) and pepducin P4pal-10 (PAR-4 antagonist) (85%) [160]. UC supernatant also promotes colonic hyposensitivity to distension, an effect can be mimicked by PAR-4 AP or Cat-G. Blockage of PAR-4 or Cat-G inhibition results in colonic hypersensitivity [161].
8. Summary: Potential Roles of Agonists and Antagonists of PARs in Allergic Inflammation
The potential roles of agonists (Figure 3(a)) and antagonists of PARs ( Figure 3(b)) in allergic inflammation are summarized. Although PARs as a unique family of GPCRs are widely expressed on inflammatory cells, each member of the family appears to selectively be expressed on different cell types. For example, mast cells express PAR-1, -2, -3, and -4 under different conditions, but eosinophils and neutrophils seem to only express PAR-1 and PAR-2. Selective expression of subtypes of PARs has also been found in different structural cells such as epithelial cells, SMCs, and fibroblasts.
Figure 3.
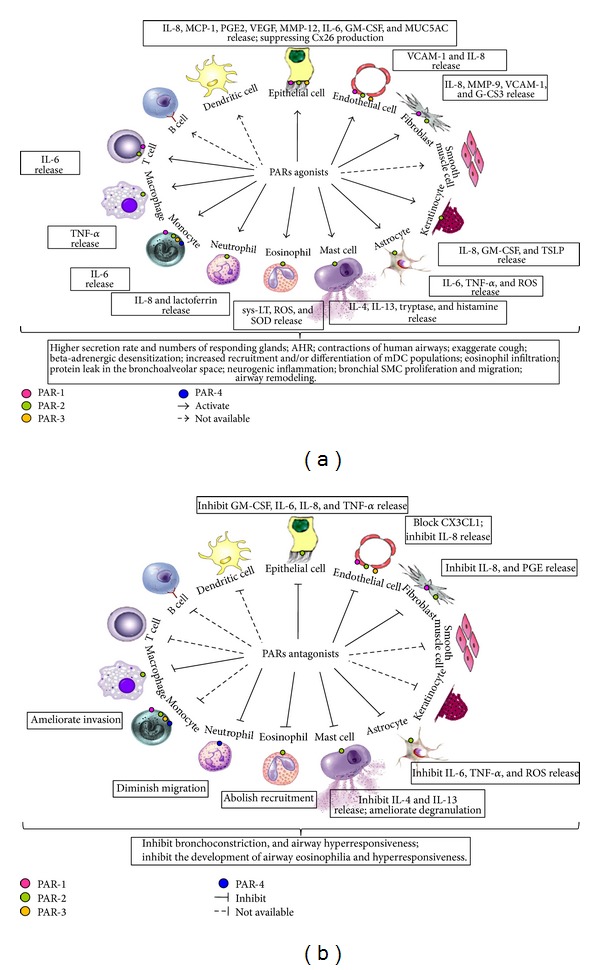
(a) Potential roles of agonists of PARs in allergic inflammation. (b) Potential roles of antagonists of PARs in allergic inflammation.
Agonists of PARs including proteases can modulate neutrophil transepithelial migration, provoke proliferation of HUVEC, HPBFs, vascular SMC, and keratinocyte, and induce murine wound healing, fibrin formation, and bronchial epithelial repair. They are also able to induce release of various types of cytokines and proinflammatory mediators from inflammatory cells. In addition, increased vascular leakage and enhanced bronchial smooth muscle contraction can be elicited by agonists of PARs.
On the other hand, selective PAR-1 antagonists interfere with platelet aggregation and cell proliferation, whereas an antagonist of PAR-4 diminishes the systemic inflammation and local neutrophilic inflammatory responses. A novel PAR-2 antagonist GB88 inhibits the acute paw edema induced by PAR-2 agonist or mast cell β-tryptase.
In AR, since PAR-2 is highly expressed in nasal mucosa, PAR-2 activating peptide provokes higher secretion rate and numbers of responding glands in nasal mucosa, and both HDM- and PAR-2 activating peptide induced responses are suppressed by ENMD-1068; the involvement of PAR-2 in AR is confirmed. In asthma, PAR-2 staining in asthmatic epithelium is increased and activation of PAR-2 has been found to induce contraction of human airways and potentiate contraction to histamine, and therefore may contribute to asthma. Studies that revealed aberrant expression and activation of serine proteases and PAR-2 in the lesional skin of AD patients, increased levels of PAR-2 in the lumen and colonic tissue of IBD patients, and upregulated PAR-2 on ileal mucosal mast cells in Crohn's ileitis implicate that PAR-2 is likely involved in the pathogenesis of these diseases. Moreover, inhibition of acute colonic inflammation by GB88 suggests the therapeutic potential for PAR-2 antagonist in inflammatory diseases.
In conclusion, increased expression and activation of PARs, particularly PAR-2, are closely associated with inflammatory conditions, suggesting that this relatively novel receptor family is likely to contribute to inflammatory process and subsequently facilitates allergic disorders. However, lack of clinically effective anti-PAR drugs and multihospital involved clinical investigation is not supportive for the statement that PARs play key roles in allergy as yet.
9. Future Work
In order to further evaluate the potential roles of PARs in allergy, the following work should be done: (1) investigating the reasons for selective expression of PARs on different cell types; (2) investigating influence of different types of allergens on expression and functions of PARs; (3) investigating further the effects of various inflammatory mediators on expression and functions of PARs, and vice versa; (4) investigating PARs in allergic conditions in a more and better designed clinical way; (5) developing clinically effective drugs for treatment of allergy.
Acknowledgment
This project was sponsored by the Grants from the Major State Basic Research Program of China (973 Program) (no. 2013CB530501); the National Natural Science Foundation of China (nos. 81241135, 81060250, 81030054, 81001428, and 81172836); State Scholarship Fund of China (201208460001); Jiangsu Province's Key Provincial Talents Program (RC201170); National “Twelfth Five-Year” Plan for Science & Technology Support Project (2014BAI07B02); the Innovation Team Project of Education Department of Liaoning Province (LT2013017); the Higher Education Climb Scholars Program of Liaoning Province, China (no. LJ2013222); and the Liaoning Province Translational Medicine Research Center for Allergy (no. LK2013041).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Authors' Contribution
Huiyun Zhang and Xiaoning Zeng equally contributed to this paper as the first authors.
References
- 1.Ossovskaya VS, Bunnett NW. Protease-activated receptors: contribution to physiology and disease. Physiological Reviews. 2004;84(2):579–621. doi: 10.1152/physrev.00028.2003. [DOI] [PubMed] [Google Scholar]
- 2.Landis RC. Protease activated receptors: clinical relevance to hemostasis and inflammation. Hematology/Oncology Clinics of North America. 2007;21(1):103–113. doi: 10.1016/j.hoc.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 3.Ma L, Dorling A. The roles of thrombin and protease-activated receptors in inflammation. Seminars in Immunopathology. 2012;34(1):63–72. doi: 10.1007/s00281-011-0281-9. [DOI] [PubMed] [Google Scholar]
- 4.Vu TK, Hung DT, Wheaton VI, Coughlin SR. Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation. Cell. 1991;64(6):1057–1068. doi: 10.1016/0092-8674(91)90261-v. [DOI] [PubMed] [Google Scholar]
- 5.Coughlin SR. Thrombin signalling and protease-activated receptors. Nature. 2000;407(6801):258–264. doi: 10.1038/35025229. [DOI] [PubMed] [Google Scholar]
- 6.Lee H, Hamilton JR. Physiology, pharmacology, and therapeutic potential of protease-activated receptors in vascular disease. Pharmacology and Therapeutics. 2012;134(2):246–259. doi: 10.1016/j.pharmthera.2012.01.007. [DOI] [PubMed] [Google Scholar]
- 7.Kahn ML, Hammes SR, Botka C, Coughlin SR. Gene and locus structure and chromosomal localization of the protease- activated receptor gene family. Journal of Biological Chemistry. 1998;273(36):23290–23296. doi: 10.1074/jbc.273.36.23290. [DOI] [PubMed] [Google Scholar]
- 8.Xu WF, Andersen H, Whitmore TE, et al. Cloning and characterization of human protease-activated receptor 4. Proceedings of the National Academy of Sciences of the United States of America. 1998;95(12):6642–6646. doi: 10.1073/pnas.95.12.6642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nystedt S, Emilsson K, Wahlestedt C, Sundelin J. Molecular cloning of a potential proteinase activated receptor. Proceedings of the National Academy of Sciences of the United States of America. 1994;91(20):9208–9212. doi: 10.1073/pnas.91.20.9208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Molino M, Barnathan ES, Numerof R, et al. Interactions of mast cell tryptase with thrombin receptors and PAR-2. Journal of Biological Chemistry. 1997;272(7):4043–4049. doi: 10.1074/jbc.272.7.4043. [DOI] [PubMed] [Google Scholar]
- 11.Camerer E, Huang W, Coughlin SR. Tissue factor- and factor X-dependent activation of protease-activated receptor 2 by factor VIIa. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(10):5255–5260. doi: 10.1073/pnas.97.10.5255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Takeuchi T, Harris JL, Huang W, Yan KW, Coughlin SR, Craik CS. Cellular localization of membrane-type serine protease 1 and identification of protease-activated receptor-2 and single-chain urokinase-type plasminogen activator as substrates. Journal of Biological Chemistry. 2000;275(34):26333–26342. doi: 10.1074/jbc.M002941200. [DOI] [PubMed] [Google Scholar]
- 13.Jeong HH, Jung YH, Park B, et al. Chitinase activates protease-activated receptor-2 in human airway epithelial cells. The American Journal of Respiratory Cell and Molecular Biology. 2008;39(5):530–535. doi: 10.1165/rcmb.2007-0410OC. [DOI] [PubMed] [Google Scholar]
- 14.Wilson S, Greer B, Hooper J, et al. The membrane-anchored serine protease, TMPRSS2, activates PAR-2 in prostate cancer cells. Biochemical Journal. 2005;388(3):967–972. doi: 10.1042/BJ20041066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ishihara H, Connolly AJ, Zeng D, et al. Protease-activated receptor 3 is a second thrombin receptor in humans. Nature. 1997;386(6624):502–506. doi: 10.1038/386502a0. [DOI] [PubMed] [Google Scholar]
- 16.Hakanishi-Matsui M, Zheng YW, Sulciner DJ, Welss EJ, Ludeman MJ, Coughlin SR. PAR3 is a cofactor for PAR4 activation by thrombin. Nature. 2000;404(6778):609–613. doi: 10.1038/35007085. [DOI] [PubMed] [Google Scholar]
- 17.Kahn ML, Zheng YW, Huang W, et al. A dual thrombin receptor system for platelet activation. Nature. 1998;394(6694):690–694. doi: 10.1038/29325. [DOI] [PubMed] [Google Scholar]
- 18.Faruqi TR, Weiss EJ, Shapiro MJ, Huang W, Coughlin SR. Structure-function analysis of protease-activated receptor 4 thetered Ligand peptides. Determinants of specificity and utility in assays of receptor function. Journal of Biological Chemistry. 2000;275(26):19728–19734. doi: 10.1074/jbc.M909960199. [DOI] [PubMed] [Google Scholar]
- 19.Cottrell GS, Coelho A-M, Bunnett NW. Protease-activated receptors: the role of cell-surface proteolysis in signalling. Essays in Biochemistry. 2002;38:169–183. doi: 10.1042/bse0380169. [DOI] [PubMed] [Google Scholar]
- 20.Zhang H, Yang X, Yang H, et al. Modulation of mast cell proteinase-activated receptor expression and IL-4 release by IL-12. Immunology and Cell Biology. 2007;85(7):558–566. doi: 10.1038/sj.icb.7100085. [DOI] [PubMed] [Google Scholar]
- 21.Qiao L, Zhang H, Wu S, He S. Downregulation of protease activated receptor expression and cytokine production in P815 cells by RNA interference. BMC Cell Biology. 2009;10, article 62 doi: 10.1186/1471-2121-10-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.He S, Zhang Z, Zhang H, et al. Analysis of properties and proinflammatory functions of cockroach allergens per a 1.01s. Scandinavian Journal of Immunology. 2011;74(3):288–295. doi: 10.1111/j.1365-3083.2011.02571.x. [DOI] [PubMed] [Google Scholar]
- 23.Christerson U, Keita ÅV, Söderholm JD, Gustafson-Svärd C. Potential role of protease-activated receptor-2-stimulated activation of cytosolic phospholipase A2 in intestinal myofibroblast proliferation: implications for stricture formation in Crohn's disease. Journal of Crohn's and Colitis. 2009;3(1):15–24. doi: 10.1016/j.crohns.2008.10.002. [DOI] [PubMed] [Google Scholar]
- 24.Baek OS, Kang OH, Choi YA, et al. Curcumin inhibits protease-activated receptor-2 and -4-mediated mast cell activation. Clinica Chimica Acta. 2003;338(1-2):135–141. doi: 10.1016/j.cccn.2003.08.015. [DOI] [PubMed] [Google Scholar]
- 25.Kim JA, Choi SC, Yun KJ, et al. Expression of protease-activated receptor 2 in ulcerative colitis. Inflammatory bowel diseases. 2003;9(4):224–229. doi: 10.1097/00054725-200307000-00002. [DOI] [PubMed] [Google Scholar]
- 26.Vliagoftis H. Thrombin induces mast cell adhesion to fibronectin: evidence for involvement of protease-activated receptor-1. Journal of Immunology. 2002;169(8):4551–4558. doi: 10.4049/jimmunol.169.8.4551. [DOI] [PubMed] [Google Scholar]
- 27.D’Andrea MR, Rogahn CJ, Andrade-Gordon P. Localization of protease-activated receptors -1 and -2 in human mast cells: indications for an amplified mast cell degranulation cascade. Biotechnic and Histochemistry. 2000;75(2):85–90. doi: 10.3109/10520290009064152. [DOI] [PubMed] [Google Scholar]
- 28.Han W, Wang Z, Lu X, Guo C. Protease activated receptor 4 status of mast cells in post infectious irritable bowel syndrome. Neurogastroenterology and Motility. 2012;24(2):113–e82. doi: 10.1111/j.1365-2982.2011.01841.x. [DOI] [PubMed] [Google Scholar]
- 29.Zhang H, Yang H, Ma W, Zhang Z, He S. Modulation of PAR expression and tryptic enzyme induced IL-4 production in mast cells by IL-29. Cytokine. 2013;61(2):469–477. doi: 10.1016/j.cyto.2012.10.032. [DOI] [PubMed] [Google Scholar]
- 30.Zhang H, Yang H, Zhang L, et al. Induction of IL-4 release and upregulated expression of protease activated receptors by GM-CSF in P815 cells. Cytokine. 2009;48(3):196–202. doi: 10.1016/j.cyto.2009.07.001. [DOI] [PubMed] [Google Scholar]
- 31.Zhang H, Yang H, He S. TNF increases expression of IL-4 and PARs in mast cells. Cellular Physiology and Biochemistry. 2010;26(3):327–336. doi: 10.1159/000320556. [DOI] [PubMed] [Google Scholar]
- 32.Zhang H, Yang H, Ma W, He S. Induction of IL-13 production and upregulated expression of protease activated receptor-1 by RANTES in a mast cell line. Cytokine. 2011;53(2):231–238. doi: 10.1016/j.cyto.2010.10.005. [DOI] [PubMed] [Google Scholar]
- 33.Falcone FH, Morroll S, Gibbs BF. Lack of protease activated receptor (PAR) expression in purified human basophils. Inflammation Research. 2005;54(supplement 1):S13–S14. doi: 10.1007/s00011-004-0405-y. [DOI] [PubMed] [Google Scholar]
- 34.Zhu W, He SH, Lin ZX, Fu YL. Histamine release properties of human basophils in response to various stimuli. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 2005;21(4):519–521. [PubMed] [Google Scholar]
- 35.Bolton SJ, McNulty CA, Thomas RJ, Hewitt CRA, Wardlaw AJ. Expression of and functional responses to protease-activated receptors on human eosinophils. Journal of Leukocyte Biology. 2003;74(1):60–68. doi: 10.1189/jlb.0702351. [DOI] [PubMed] [Google Scholar]
- 36.Miike S, McWilliam AS, Kita H. Trypsin induces activation and inflammatory mediator release from human eosinophils through protease-activated receptor-2. Journal of Immunology. 2001;167(11):6615–6622. doi: 10.4049/jimmunol.167.11.6615. [DOI] [PubMed] [Google Scholar]
- 37.Dinh QT, Cryer A, Trevisani M, et al. Gene and protein expression of protease-activated receptor 2 in structural and inflammatory cells in the nasal mucosa in seasonal allergic rhinitis. Clinical and Experimental Allergy. 2006;36(8):1039–1048. doi: 10.1111/j.1365-2222.2006.02537.x. [DOI] [PubMed] [Google Scholar]
- 38.Wang H, He S. Induction of lactoferrin and IL-8 release from human neutrophils by tryptic enzymes via proteinase activated receptor-2. Cell Biology International. 2006;30(9):688–697. doi: 10.1016/j.cellbi.2006.04.007. [DOI] [PubMed] [Google Scholar]
- 39.Lutfi R, Lewkowich IP, Zhou P, Ledford JR, Page K. The role of protease-activated receptor-2 on pulmonary neutrophils in the innate immune response to cockroach allergen. Journal of Inflammation. 2012;9(1):p. 32. doi: 10.1186/1476-9255-9-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Veiga Cde S, Carneiro-Lobo TC, Coelho CJBP, et al. Increased expression of protease-activated receptor 1 (PAR-1) in human leukemias. Blood Cells, Molecules, and Diseases. 2011;46(3):230–234. doi: 10.1016/j.bcmd.2010.12.005. [DOI] [PubMed] [Google Scholar]
- 41.Colognato R, Slupsky JR, Jendrach M, Burysek L, Syrovets T, Simmet T. Differential expression and regulation of protease-activated receptors in human peripheral monocytes and monocyte-derived antigen-presenting cells. Blood. 2003;102(7):2645–2652. doi: 10.1182/blood-2002-08-2497. [DOI] [PubMed] [Google Scholar]
- 42.Naldini A, Carney DH, Pucci A, Pasquali A, Carraro F. Thrombin regulates the expression of proangiogenic cytokines via proteolytic activation of protease-activated receptor-1. General Pharmacology. 2000;35(5):255–259. doi: 10.1016/s0306-3623(01)00113-6. [DOI] [PubMed] [Google Scholar]
- 43.Johansson U, Lawson C, Dabare M, et al. Human peripheral blood monocytes express protease receptor-2 and respond to receptor activation by production of IL-6, IL-8, and IL-1β . Journal of Leukocyte Biology. 2005;78(4):967–975. doi: 10.1189/jlb.0704422. [DOI] [PubMed] [Google Scholar]
- 44.Macey MG, Wolf SI, Wheeler-Jones CPD, Lawson C. Expression of blood coagulation factors on monocytes after exposure to TNF-treated endothelium in a novel whole blood model of arterial flow. Journal of Immunological Methods. 2009;350(1-2):133–141. doi: 10.1016/j.jim.2009.08.007. [DOI] [PubMed] [Google Scholar]
- 45.Uehara A, Iwashiro A, Sato T, Yokota S, Takada H. Antibodies to proteinase 3 prime human monocytic cells via protease-activated receptor-2 and NF-κB for Toll-like receptor- and NOD-dependent activation. Molecular Immunology. 2007;44(14):3552–3562. doi: 10.1016/j.molimm.2007.03.010. [DOI] [PubMed] [Google Scholar]
- 46.López-Pedrera C, Aguirre MÁ, Buendía P, et al. Differential expression of protease-activated receptors in monocytes from patients with primary antiphospholipid syndrome. Arthritis and Rheumatism. 2010;62(3):869–877. doi: 10.1002/art.27299. [DOI] [PubMed] [Google Scholar]
- 47.Nieuwenhuizen L, Falkenburg WJ, Schutgens RE, et al. Stimulation of naïve monocytes and PBMCs with coagulation proteases results in thrombin-mediated and PAR-1-dependent cytokine release and cell proliferation in PBMCs only. Scandinavian Journal of Immunology. 2013;77(5):339–349. doi: 10.1111/sji.12033. [DOI] [PubMed] [Google Scholar]
- 48.Roche N, Stirling RG, Lim S, et al. Effect of acute and chronic inflammatory stimuli on expression of protease-activated receptors 1 and 2 in alveolar macrophages. Journal of Allergy and Clinical Immunology. 2003;111(2):367–373. doi: 10.1067/mai.2003.6. [DOI] [PubMed] [Google Scholar]
- 49.Kim JY, Sohn JH, Choi JM, et al. Alveolar macrophages play a key role in cockroach-induced allergic inflammation via TNF-α pathway. PLoS ONE. 2012;7(10) doi: 10.1371/journal.pone.0047971.e47971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Jesmin S, Gando S, Zaedi S, Sakuraya F. Differential expression, time course and distribution of four PARs in rats with endotoxin-induced acute lung injury. Inflammation. 2006;30(1-2):14–27. doi: 10.1007/s10753-006-9017-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lim J, Iyer A, Liu L, et al. Diet-induced obesity, adipose inflammation, and metabolic dysfunction correlating with PAR2 expression are attenuated by PAR2 antagonism. The FASEB Journal. 2013;27(12):4757–4767. doi: 10.1096/fj.13-232702. [DOI] [PubMed] [Google Scholar]
- 52.Hurley A, Smith M, Karpova TJ, et al. Enhanced effector function of CD8+ T cells from healthy controls and HIV-infected patients occurs through thrombin activation ofprotease-activated receptor 1. The Journal of Infectious Diseases. 2013;207(4):638–650. doi: 10.1093/infdis/jis730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Belibasakis GN, Bostanci N, Reddi D. Regulation of protease-activated receptor-2 expression in gingival fibroblasts and Jurkat T cells by Porphyromonas gingivalis. Cell Biology International. 2010;34(3):287–292. doi: 10.1042/CBI20090290. [DOI] [PubMed] [Google Scholar]
- 54.Kalashnyk O, Petrova Y, Lykhmus O, et al. Expression, function and cooperating partners of protease-activatedreceptor type 3 in vascular endothelial cells and B lymphocytesstudied with specific monoclonal antibody. Molecular Immunology. 2013;54:319–326. doi: 10.1016/j.molimm.2012.12.021. [DOI] [PubMed] [Google Scholar]
- 55.Yanagita M, Kobayashi R, Kashiwagi Y, Shimabukuro Y, Murakami S. Thrombin regulates the function of human blood dendritic cells. Biochemical and Biophysical Research Communications. 2007;364(2):318–324. doi: 10.1016/j.bbrc.2007.10.002. [DOI] [PubMed] [Google Scholar]
- 56.Li X, Syrovets T, Paskas S, Laumonnier Y, Simmet T. Mature dendritic cells express functional thrombin receptors triggering chemotaxis and CCL18/pulmonary and activation-regulated chemokine induction. Journal of Immunology. 2008;181(2):1215–1223. doi: 10.4049/jimmunol.181.2.1215. [DOI] [PubMed] [Google Scholar]
- 57.Lewkowich IP, Day SB, Ledford JR, et al. Protease-activated receptor 2 activation of myeloid dendritic cells regulates allergic airway inflammation. Respiratory Research. 2011;12, article 122 doi: 10.1186/1465-9921-12-122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Asokananthan N, Graham PT, Fink J, et al. Activation of protease-activated receptor (PAR)-1, PAR-2, and PAR-4 stimulates IL-6, IL-8, and prostaglandin E2 release from human respiratory epithelial cells. Journal of Immunology. 2002;168(7):3577–3585. doi: 10.4049/jimmunol.168.7.3577. [DOI] [PubMed] [Google Scholar]
- 59.Page K, Strunk VS, Hershenson MB. Cockroach proteases increase IL-8 expression in human bronchial epithelial cells via activation of protease-activated receptor (PAR)-2 and extracellular-signal-regulated kinase. Journal of Allergy and Clinical Immunology. 2003;112(6):1112–1118. doi: 10.1016/j.jaci.2003.08.050. [DOI] [PubMed] [Google Scholar]
- 60.Shea-Donohue T, Notari L, Stiltz J, et al. Role of enteric nerves in immune-mediated changes in protease-activated receptor 2 effects on gut function. Neurogastroenterology and Motility. 2010;22(10):1138–e291. doi: 10.1111/j.1365-2982.2010.01557.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Pan SL, Tao KY, Guh JH, et al. The p38 mitogen-activated protein kinase pathway plays a critical role in PAR2-induced endothelial IL-8 production and leukocyte adhesion. Shock. 2008;30(5):496–502. doi: 10.1097/SHK.0b013e3181673233. [DOI] [PubMed] [Google Scholar]
- 62.Nguyen KT, Eskin SG, Patterson C, Runge MS, McIntire LV. Shear stress reduces protease activated receptor-1 expression in human endothelial cells. Annals of Biomedical Engineering. 2001;29(2):145–152. doi: 10.1114/1.1349700. [DOI] [PubMed] [Google Scholar]
- 63.Nystedt S, Ramakrishnan V, Sundelin J. The proteinase-activated receptor 2 is induced by inflammatory mediators in human endothelial cells: comparison with the thrombin receptor. Journal of Biological Chemistry. 1996;271(25):14910–14915. doi: 10.1074/jbc.271.25.14910. [DOI] [PubMed] [Google Scholar]
- 64.Shimizu T, Nishihira J, Watanabe H, et al. Macrophage migration inhibitory factor is induced by thrombin and factor Xa in endothelial cells. Journal of Biological Chemistry. 2004;279(14):13729–13737. doi: 10.1074/jbc.M400150200. [DOI] [PubMed] [Google Scholar]
- 65.Soh UJK, Trejo J. Activated protein C promotes protease-activated receptor-1 cytoprotective signaling through β-arrestin and dishevelled-2 scaffolds. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(50):E1372–E1380. doi: 10.1073/pnas.1112482108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Popović M, Paskaš S, Živković M, Burysek L, Laumonnier Y. Human cytomegalovirus increases HUVEC sensitivity to thrombin and modulates expression of thrombin receptors. Journal of Thrombosis and Thrombolysis. 2010;30(2):164–171. doi: 10.1007/s11239-010-0447-7. [DOI] [PubMed] [Google Scholar]
- 67.Schmidt VA, Nierman WC, Maglott DR, et al. The human proteinase-activated receptor-3 (PAR-3) gene. Identification within a PAR gene cluster and characterization in vascular endothelial cells and platelets. Journal of Biological Chemistry. 1998;273(24):15061–15068. doi: 10.1074/jbc.273.24.15061. [DOI] [PubMed] [Google Scholar]
- 68.Ramachandran R, Morice AH, Compton SJ. Proteinase-activated receptor2 agonists upregulate granulocyte colony-stimulating factor, IL-8, and VCAM-1 expression in human bronchial fibroblasts. American Journal of Respiratory Cell and Molecular Biology. 2006;35(1):133–141. doi: 10.1165/rcmb.2005-0362OC. [DOI] [PubMed] [Google Scholar]
- 69.Uehara A, Muramoto K, Imamura T, et al. Arginine-specific gingipains from Porphyromonas gingivalis stimulate production of hepatocyte growth factor (scatter factor) through protease-activated receptors in human gingival fibroblasts in culture. Journal of Immunology. 2005;175(9):6076–6084. doi: 10.4049/jimmunol.175.9.6076. [DOI] [PubMed] [Google Scholar]
- 70.Sokolova E, Grishina Z, Bühling F, Welte T, Reiser G. Protease-activated receptor-1 in human lung fibroblasts mediates a negative feedback downregulation via prostaglandin e2. The American Journal of Physiology—Lung Cellular and Molecular Physiology. 2005;288(5):L793–L802. doi: 10.1152/ajplung.00343.2004. [DOI] [PubMed] [Google Scholar]
- 71.Abe K, Aslam A, Walls AF, Sato T, Inoue H. Up-regulation of protease-activated receptor-2 by bFGF in cultured human synovial fibroblasts. Life Sciences. 2006;79(9):898–904. doi: 10.1016/j.lfs.2006.03.034. [DOI] [PubMed] [Google Scholar]
- 72.Sokolova E, Hartig R, Reiser G. Downregulation of protease-activated receptor-1 in human lung fibroblasts is specifically mediated by the prostaglandin E2 receptor EP2 through cAMP elevation and protein kinase A. FEBS Journal. 2008;275(14):3669–3679. doi: 10.1111/j.1742-4658.2008.06511.x. [DOI] [PubMed] [Google Scholar]
- 73.Tran T, Stewart AG. Protease-activated receptor (PAR)-independent growth and pro-inflammatory actions of thrombin on human cultured airway smooth muscle. British Journal of Pharmacology. 2003;138(5):865–875. doi: 10.1038/sj.bjp.0705106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Chambers LS, Black JL, Ge Q, et al. PAR-2 activation, PGE2, and COX-2 in human asthmatic and nonasthmatic airway smooth muscle cells. The American Journal of Physiology—Lung Cellular and Molecular Physiology. 2003;285(3):L619–L627. doi: 10.1152/ajplung.00416.2002. [DOI] [PubMed] [Google Scholar]
- 75.Schrör K, Bretschneider E, Fischer K, et al. Thrombin receptors in vascular smooth muscle cells—function and regulation by vasodilatory prostaglandins. Thrombosis and Haemostasis. 2010;103(5):884–890. doi: 10.1160/TH09-09-0627. [DOI] [PubMed] [Google Scholar]
- 76.Giacaman RA, Asrani AC, Ross KF, Herzberg MC. Cleavage of protease-activated receptors on an immortalized oral epithelial cell line by Porphyromonas gingivalis gingipains. Microbiology. 2009;155(10):3238–3246. doi: 10.1099/mic.0.029132-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Enomoto A, Yoshihisa Y, Yamakoshi T, et al. UV-B radiation induces macrophage migration inhibitory factor-mediated melanogenesis through activation of protease-activated receptor-2 and stem cell factor in keratinocytes. The American Journal of Pathology. 2011;178(2):679–687. doi: 10.1016/j.ajpath.2010.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Holinstat M, Voss B, Bilodeau ML, McLaughlin JN, Cleator J, Hamm HE. PAR4, but not PAR1, signals human platelet aggregation via Ca2+ mobilization and synergistic P2Y12 receptor activation. Journal of Biological Chemistry. 2006;281(36):26665–26674. doi: 10.1074/jbc.M602174200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hasdemir B, Murphy JE, Cottrell GS, Bunnett NW. Endosomal deubiquitinating enzymes control ubiquitination and down-regulation of protease-activated receptor 2. Journal of Biological Chemistry. 2009;284(41):28453–28466. doi: 10.1074/jbc.M109.025692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.McLaughlin JN, Patterson MM, Malik AB. Protease-activated receptor-3 (PAR3) regulates PAR1 signaling by receptor dimerization. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(13):5662–5667. doi: 10.1073/pnas.0700763104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.DeFea KA, Zalevsky J, Thoma MS, Dery O, Mullins RD, Bunnett NW. β-Arrestin-dependent endocytosis of proteinase-activated receptor 2 is required for intracellular targeting of activated ERK1/2. Journal of Cell Biology. 2000;148(6):1267–1281. doi: 10.1083/jcb.148.6.1267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Rohani MG, DiJulio DH, An JY, Hacker BM, Dale BA, Chung WO. PAR1- and PAR2-induced innate immune markers are negatively regulated by PI3K/Akt signaling pathway in oral keratinocytes. BMC Immunology. 2010;11, article 53 doi: 10.1186/1471-2172-11-53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Wilson BJ, Harada R, Leduy L, Hollenberg MD, Nepveu A. CUX1 transcription factor is a downstream effector of the proteinase-activated receptor 2 (PAR2) Journal of Biological Chemistry. 2009;284(1):36–45. doi: 10.1074/jbc.M803808200. [DOI] [PubMed] [Google Scholar]
- 84.Palma-Nicolas JP, López E, López-Colomé AM. PKC isoenzymes differentially modulate the effect of thrombin on MAPK-dependent RPE proliferation. Bioscience reports. 2008;28(6):307–317. doi: 10.1042/BSR20080083. [DOI] [PubMed] [Google Scholar]
- 85.Shpacovitch VM, Seeliger S, Huber-lang M, et al. Agonists of proteinase-activated receptor-2 affect transendothelial migration and apoptosis of human neutrophils. Experimental Dermatology. 2007;16(10):799–806. doi: 10.1111/j.1600-0625.2007.00605.x. [DOI] [PubMed] [Google Scholar]
- 86.Chin AC, Lee WY, Nusrat A, Vergnolle N, Parkos CA. Neutrophil-mediated activation of epithelial protease-activated receptors-1 and -2 regulates barrier function and transepithelial migration. Journal of Immunology. 2008;181(8):5702–5710. doi: 10.4049/jimmunol.181.8.5702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Feistritzer C, Mosheimer BA, Kaneider NC, Riewald M, Patsch JR, Wiedermann CJ. Thrombin affects eosinophil migration via protease-activated receptor-1. International Archives of Allergy and Immunology. 2004;135(1):12–16. doi: 10.1159/000080037. [DOI] [PubMed] [Google Scholar]
- 88.Olivot J-M, Estebanell E, Lafay M, Brohard B, Aiach M, Rendu F. Thrombomodulin prolongs thrombin-induced extracellular signal-regulated kinase phosphorylation and nuclear retention in endothelial cells. Circulation Research. 2001;88(7):681–687. doi: 10.1161/hh0701.088769. [DOI] [PubMed] [Google Scholar]
- 89.Mahajan SG, Fender AC, Meyer-Kirchrath J, et al. A novel function of FoxO transcription factors in thrombin-stimulated vascular smooth muscle cell proliferation. Thrombosis and Haemostasis. 2012;108(1):148–158. doi: 10.1160/TH11-11-0756. [DOI] [PubMed] [Google Scholar]
- 90.Julovi SM, Xue M, Dervish S, Sambrook PN, March L, Jackson CJ. Protease activated receptor-2 mediates activated protein cinduced cutaneous wound healing via inhibition of p38. American Journal of Pathology. 2011;179(5):2233–2242. doi: 10.1016/j.ajpath.2011.07.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Ewen D, Clarke SL, Smith JR, et al. The role of protease-activated receptors PAR-1 and PAR-2 in the repair of 16HBE 14o− epithelial cell monolayers in vitro. Clinical and Experimental Allergy. 2010;40(3):435–449. doi: 10.1111/j.1365-2222.2010.03453.x. [DOI] [PubMed] [Google Scholar]
- 92.Wang H, Zheng Y, He S. Induction of release and up-regulated gene expression of interleukin (IL)-8 in A549 cells by serine proteinases. BMC Cell Biology. 2006;7, article 22 doi: 10.1186/1471-2121-7-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Wang H, Yi T, Zheng Y, He S. Induction of monocyte chemoattractant protein-1 release from A549 cells by agonists of protease-activated receptor-1 and -2. European Journal of Cell Biology. 2007;86(4):233–242. doi: 10.1016/j.ejcb.2006.12.003. [DOI] [PubMed] [Google Scholar]
- 94.Li T, Wang H, He S. Induction of interleukin-6 release from monocytes by serine proteinases and its potential mechanisms. Scandinavian Journal of Immunology. 2006;64(1):10–16. doi: 10.1111/j.1365-3083.2006.01772.x. [DOI] [PubMed] [Google Scholar]
- 95.Li T, He S. Induction of IL-6 release from human T cells by PAR-1 and PAR-2 agonists. Immunology and Cell Biology. 2006;84(5):461–466. doi: 10.1111/j.1440-1711.2006.01456.x. [DOI] [PubMed] [Google Scholar]
- 96.Wang L, Luo J, Fu Y, He S. Induction of interleukin-8 secretion and activation of ERK1/2, p38 MAPK signaling pathways by thrombin in dermal fibroblasts. International Journal of Biochemistry and Cell Biology. 2006;38(9):1571–1583. doi: 10.1016/j.biocel.2006.03.016. [DOI] [PubMed] [Google Scholar]
- 97.Wang L, Luo J, He S. Induction of MMP-9 release from human dermal fibroblasts by thrombin: involvement of JAK/STAT3 signaling pathway in MMP-9 release. BMC Cell Biology. 2007;8, article 14 doi: 10.1186/1471-2121-8-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Shimizu S, Gabazza EC, Ogawa T, et al. Role of thrombin in chronic rhinosinusitis-associated tissue remodeling. The American Journal of Rhinology and Allergy. 2011;25(1):7–11. doi: 10.2500/ajra.2011.25.3535. [DOI] [PubMed] [Google Scholar]
- 99.Raza SL, Nehring LC, Shapiro SD, Cornelius LA. Proteinase-activated receptor-1 regulation of macrophage elastase (MMP-12) secretion by serine proteinases. Journal of Biological Chemistry. 2000;275(52):41243–41250. doi: 10.1074/jbc.M005788200. [DOI] [PubMed] [Google Scholar]
- 100.Mazor R, Alsaigh T, Shaked H, et al. Matrix metalloproteinase-1-mediated up-regulation of vascular endothelial growth factor-2 in endothelial cells. The Journal of Biological Chemistry. 2013;288(1):598–607. doi: 10.1074/jbc.M112.417451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Zhou J, Perelman JM, Kolosov VP, Zhou X. Neutrophil elastase induces MUC5AC secretion via protease-activatedreceptor 2. Molecular and Cellular Biochemistry. 2013;377(1-2):75–85. doi: 10.1007/s11010-013-1572-3. [DOI] [PubMed] [Google Scholar]
- 102.Vesey DA, Suen JY, Seow V, et al. PAR2-induced inflammatory responses in human kidney tubular epithelial cells. The American Journal of Physiology—Renal Physiology. 2013;304(6):F737–F750. doi: 10.1152/ajprenal.00540.2012. [DOI] [PubMed] [Google Scholar]
- 103.Moniaga CS, Jeong SK, Egawa G, et al. Protease activity enhances production of thymic stromal lymphopoietin and basophil accumulation in flaky tail mice. The American Journal of Pathology. 2013;182(3):841–851. doi: 10.1016/j.ajpath.2012.11.039. [DOI] [PubMed] [Google Scholar]
- 104.Kato T, Takai T, Fujimura T, et al. Mite serine protease activates protease-activated receptor-2 and induces cytokine release in human keratinocytes. Allergy. 2009;64(9):1366–1374. doi: 10.1111/j.1398-9995.2009.02023.x. [DOI] [PubMed] [Google Scholar]
- 105.Heijink IH, van Oosterhout A, Kapus A. Epidermal growth factor receptor signalling contributes to house dust mite-induced epithelial barrier dysfunction. European Respiratory Journal. 2010;36(5):1016–1026. doi: 10.1183/09031936.00125809. [DOI] [PubMed] [Google Scholar]
- 106.Girardi G. Pravastatin prevents miscarriages in antiphospholipid antibody-treated mice. Journal of reproductive immunology. 2009;82(2):126–131. doi: 10.1016/j.jri.2009.03.003. [DOI] [PubMed] [Google Scholar]
- 107.He SH, Xie H, Fu YL. Activation of human tonsil and skin mast cells by agonists of proteinase activated receptor-2. Acta Pharmacologica Sinica. 2005;26(5):568–574. doi: 10.1111/j.1745-7254.2005.00079.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Niu QX, Chen HQ, Chen ZY, Fu YL, Lin JL, He SH. Induction of inflammatory cytokine release from human umbilical vein endothelial cells by agonists of proteinase-activated receptor-2. Clinical and Experimental Pharmacology and Physiology. 2008;35(1):89–96. doi: 10.1111/j.1440-1681.2007.04755.x. [DOI] [PubMed] [Google Scholar]
- 109.Matsuwaki Y, Wada K, White T, Moriyama H, Kita H. Alternaria fungus induces the production of GM-CSF, interleukin-6 and interleukin-8 and calcium signaling in human airway epithelium through protease-activated receptor 2. International Archives of Allergy and Immunology. 2012;158(supplement 1):19–29. doi: 10.1159/000337756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Matsuwaki Y, Wada K, White TA, et al. Recognition of fungal protease activities induces cellular activation and eosinophil-derived neurotoxin release in human eosinophils. Journal of Immunology. 2009;183(10):6708–6716. doi: 10.4049/jimmunol.0901220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Zeng X, Zhang S, Xu L, Yang H, He S. Activation of protease-activated receptor 2-mediated signaling by mast cell tryptase modulates cytokine production in primary cultured astrocytes. Mediators of Inflammation. 2013;2013:10 pages. doi: 10.1155/2013/140812.140812 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Andrade-Gordon P, Maryanoff BE, Derian CK, et al. Design, synthesis, and biological characterization of a peptide-mimetic antagonist for a tethered-ligand receptor. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(22):12257–12262. doi: 10.1073/pnas.96.22.12257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Maryanoff BE, Zhang HC, Andrade-Gordon P, Derian CK. Discovery of potent peptide-mimetic antagonists for the human thrombin receptor, protease-activated receptor-1 (PAR-1) Current Medicinal Chemistry—Cardiovascular and Hematological Agents. 2003;1(1):13–36. doi: 10.2174/1568016033356724. [DOI] [PubMed] [Google Scholar]
- 114.Ofosu FA, Dewar L, Craven SJ, et al. Coordinate activation of human platelet protease-activated receptor-1 and -4 in response to subnanomolar α-thrombin. Journal of Biological Chemistry. 2008;283(40):26886–26893. doi: 10.1074/jbc.M802237200. [DOI] [PubMed] [Google Scholar]
- 115.Slofstra SH, Bijlsma MF, Groot AP, et al. Protease-activated receptor-4 inhibition protects from multiorgan failure in a murine model of systemic inflammation. Blood. 2007;110(9):3176–3182. doi: 10.1182/blood-2007-02-075440. [DOI] [PubMed] [Google Scholar]
- 116.Lohman R-J, Cotterell AJ, Suen J, et al. Antagonism of protease-activated receptor 2 protects against experimental colitis. Journal of Pharmacology and Experimental Therapeutics. 2012;340(2):256–265. doi: 10.1124/jpet.111.187062. [DOI] [PubMed] [Google Scholar]
- 117.Suen JY, Barry GD, Lohman RJ, et al. Modulating human proteinase activated receptor 2 with a novel antagonist (GB88) and agonist (GB110) British Journal of Pharmacology. 2012;165(5):1413–1423. doi: 10.1111/j.1476-5381.2011.01610.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Miyake Y, D'Alessandro-Gabazza CN, Takagi T, et al. Dose-dependent differential effects of thrombin in allergic bronchial asthma. Journal of Thrombosis and Haemostasis. 2013;11(10):1903–1015. doi: 10.1111/jth.12392. [DOI] [PubMed] [Google Scholar]
- 119.Zhu W, Bi M, Liu Y, et al. Thrombin promotes airway remodeling via protease-activatedreceptor-1 and transforming growth factor-β1 in ovalbumin-allergic rats. Inhalation Toxicology. 2013;25(10):577–586. doi: 10.3109/08958378.2013.813995. [DOI] [PubMed] [Google Scholar]
- 120.Yoshikawa T, Kanazawa H. Integrated effect of EGFR and PAR-1 signaling crosstalk on airway hyperresponsiveness. International Journal of Molecular Medicine. 2012;30(1):41–48. doi: 10.3892/ijmm.2012.981. [DOI] [PubMed] [Google Scholar]
- 121.Lee H-M, Hyo YK, Hee JK, et al. Up-regulation of protease-activated receptor 2 in allergic rhinitis. Annals of Otology, Rhinology and Laryngology. 2007;116(7):554–558. doi: 10.1177/000348940711600712. [DOI] [PubMed] [Google Scholar]
- 122.Cho HJ, Lee HJ, Kim SC, et al. Protease-activated receptor 2-dependent fluid secretion from airway submucosal glands by house dust mite extract. Journal of Allergy and Clinical Immunology. 2012;129(2):529–535. doi: 10.1016/j.jaci.2011.11.024. [DOI] [PubMed] [Google Scholar]
- 123.Cho HJ, Choi JY, Yang Y-M, et al. House dust mite extract activates apical Cl− channels through protease-activated receptor 2 in human airway epithelia. Journal of Cellular Biochemistry. 2010;109(6):1254–1263. doi: 10.1002/jcb.22511. [DOI] [PubMed] [Google Scholar]
- 124.Zheng J, Liu W, Fan Y, et al. Suppression of connexin 26 is related to protease-activated receptor 2-mediated pathway in patients with allergic rhinitis. The American Journal of Rhinology and Allergy. 2012;26(1):p. e5, p. e9. doi: 10.2500/ajra.2012.26.3740. [DOI] [PubMed] [Google Scholar]
- 125.Shi J, Luo Q, Chen F, Chen D, Xu G, Li H. Induction of IL-6 and IL-8 by house dust mite allergen Der p1 in cultured human nasal epithelial cells is associated with PAR/PI3K/NFkappaB signaling. ORL. 2010;72(5):256–265. doi: 10.1159/000312687. [DOI] [PubMed] [Google Scholar]
- 126.Dinh QT, Cryer A, Dinh S, et al. Protease-activated receptor 2 expression in trigeminal neurons innervating the rat nasal mucosa. Neuropeptides. 2005;39(5):461–466. doi: 10.1016/j.npep.2005.07.003. [DOI] [PubMed] [Google Scholar]
- 127.Knight DA, Lim S, Scaffidi AK, et al. Protease-activated receptors in human airways: Upregulation of PAR-2 in respiratory epithelium from patients with asthma. Journal of Allergy and Clinical Immunology. 2001;108(5):797–803. doi: 10.1067/mai.2001.119025. [DOI] [PubMed] [Google Scholar]
- 128.Chambers LS, Black JL, Poronnik P, Johnson PRA. Functional effects of protease-activated receptor-2 stimulation on human airway smooth muscle. The American Journal of Physiology—Lung Cellular and Molecular Physiology. 2001;281(6):L1369–L1378. doi: 10.1152/ajplung.2001.281.6.L1369. [DOI] [PubMed] [Google Scholar]
- 129.Bara I, Ozier A, Girodet PO, et al. Role of YKL-40 in bronchial smooth muscle remodeling in asthma. The American Journal of Respiratory and Critical Care Medicine. 2012;185(7):715–722. doi: 10.1164/rccm.201105-0915OC. [DOI] [PubMed] [Google Scholar]
- 130.Rievaj J, Davidson C, Nadeem A, Hollenberg M, Duszyk M, Vliagoftis H. Allergic sensitization enhances anion current responsiveness of murine trachea to PAR-2 activation. Pflugers Archiv European Journal of Physiology. 2012;463(3):497–509. doi: 10.1007/s00424-011-1064-9. [DOI] [PubMed] [Google Scholar]
- 131.Schmidlin F, Amadesi S, Dabbagh K, et al. Protease-activated receptor 2 mediates eosinophil infiltration and hyperreactivity in allergic inflammation of the airway. Journal of Immunology. 2002;169(9):5315–5321. doi: 10.4049/jimmunol.169.9.5315. [DOI] [PubMed] [Google Scholar]
- 132.de Boer JD, Van't Veer C, Stroo I, et al. Protease-activated receptor-2 deficient mice have reduced house dust mite-evoked allergic lung inflammation. Innate Immunity. 2013 doi: 10.1177/1753425913503387. [DOI] [PubMed] [Google Scholar]
- 133.Gu Q, Lee LY. House dust mite potentiates capsaicin-evoked Ca2+ transients in mouse pulmonary sensory neurons via activation of protease-activated receptor-2. Experimental Physiology. 2012;97(4):534–543. doi: 10.1113/expphysiol.2011.060764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Miglino N, Roth M, Tamm M, Borger P. House dust mite extract downregulates C/EBPα in asthmatic bronchial smooth muscle cells. European Respiratory Journal. 2011;38(1):50–58. doi: 10.1183/09031936.00068010. [DOI] [PubMed] [Google Scholar]
- 135.Page K, Hughes VS, Bennett GW, Wong HR. German cockroach proteases regulate matrix metalloproteinase-9 in human bronchial epithelial cells. Allergy. 2006;61(8):988–995. doi: 10.1111/j.1398-9995.2006.01103.x. [DOI] [PubMed] [Google Scholar]
- 136.Hong JH, Lee SI, Kim KE, et al. German cockroach extract activates protease-activated receptor 2 in human airway epithelial cells. Journal of Allergy and Clinical Immunology. 2004;113(2):315–319. doi: 10.1016/j.jaci.2003.11.026. [DOI] [PubMed] [Google Scholar]
- 137.Page K, Ledford JR, Zhou P, Dienger K, Wills-Karp M. Mucosal sensitization to German cockroach involves protease-activated receptor-2. Respiratory Research. 2010;11, article 62 doi: 10.1186/1465-9921-11-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Boitano S, Flynn AN, Sherwood CL, et al. Alternaria alternata serine proteases induce lung inflammation and airway epithelial cell activation via PAR2. The American Journal of Physiology—Lung Cellular and Molecular Physiology. 2011;300(4):L605–L614. doi: 10.1152/ajplung.00359.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Day SB, Ledford JR, Zhou P, Lewkowich IP, Page K. German cockroach proteases and protease-activated receptor-2 regulate chemokine production and dendritic cell recruitment. Journal of Innate Immunity. 2011;4(1):100–110. doi: 10.1159/000329132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Kobayashi M, Kume H, Oguma T, Makino Y, Ito Y, Shimokata K. Mast cell tryptase causes homologous desensitization of β-adrenoceptors by Ca2+ sensitization in tracheal smooth muscle. Clinical and Experimental Allergy. 2008;38(1):135–144. doi: 10.1111/j.1365-2222.2007.02879.x. [DOI] [PubMed] [Google Scholar]
- 141.Berger P, Girodet PO, Begueret H, et al. Tryptase-stimulated human airway smooth muscle cells induce cytokine synthesis and mast cell chemotaxis. The FASEB Journal. 2003;17(14):2139–2141. doi: 10.1096/fj.03-0041fje. [DOI] [PubMed] [Google Scholar]
- 142.Gatti R, Andre E, Amadesi S, et al. Protease-activated receptor-2 activation exaggerates TRPV1-mediated cough in guinea pigs. Journal of Applied Physiology. 2006;101(2):506–511. doi: 10.1152/japplphysiol.01558.2005. [DOI] [PubMed] [Google Scholar]
- 143.De Campo BA, Henry PJ. Stimulation of protease-activated receptor-2 inhibits airway eosinophilia, hyperresponsiveness and bronchoconstriction in a murine model of allergic inflammation. British Journal of Pharmacology. 2005;144(8):1100–1108. doi: 10.1038/sj.bjp.0706150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.D’Agostino B, Roviezzo F, De Palma R, et al. Activation of protease-activated receptor-2 reduces airways inflammation in experimental allergic asthma. Clinical and Experimental Allergy. 2007;37(10):1436–1443. doi: 10.1111/j.1365-2222.2007.02793.x. [DOI] [PubMed] [Google Scholar]
- 145.Lee SE, Jeong SK, Lee SH. Protease and Protease-activated Receptor-2 signaling in the pathogenesis of atopic dermatitis. Yonsei Medical Journal. 2010;51(6):808–822. doi: 10.3349/ymj.2010.51.6.808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hovnanian A. Netherton syndrome: skin inflammation and allergy by loss of protease inhibition. Cell & Tissue Research. 2013;351(2):289–300. doi: 10.1007/s00441-013-1558-1. [DOI] [PubMed] [Google Scholar]
- 147.Jeong SK, Kim HJ, Youm JK, et al. Mite and cockroach allergens activate protease-activated receptor 2 and delay epidermal permeability barrier recovery. Journal of Investigative Dermatology. 2008;128(8):1930–1939. doi: 10.1038/jid.2008.13. [DOI] [PubMed] [Google Scholar]
- 148.Zhu Y, Peng C, Xu JG, et al. Participation of proteinase-activated receptor-2 in passive cutaneous anaphylaxis-induced scratching behavior and the inhibitory effect of tacrolimus. Biological and Pharmaceutical Bulletin. 2009;32(7):1173–1176. doi: 10.1248/bpb.32.1173. [DOI] [PubMed] [Google Scholar]
- 149.Nakano T, Andoh T, Tayama M, Kosaka M, Lee JB, Kuraishi Y. Effects of topical application of tacrolimus on acute itch-associated responses in mice. Biological and Pharmaceutical Bulletin. 2008;31(4):752–754. doi: 10.1248/bpb.31.752. [DOI] [PubMed] [Google Scholar]
- 150.Kawagoe J, Takizawa T, Matsumoto J, et al. Effect of protease-activated receptor-2 deficiency on allergic dermatitis in the mouse ear. Japanese Journal of Pharmacology. 2002;88(1):77–88. doi: 10.1254/jjp.88.77. [DOI] [PubMed] [Google Scholar]
- 151.Moussa L, Bézirard V, Salvador-Cartier C, et al. A low dose of fermented soy germ alleviates gut barrier injury, hyperalgesia and faecal protease activity in a rat model of inflammatory bowel disease. PLoS ONE. 2012;7(11) doi: 10.1371/journal.pone.0049547.e49547 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Cenac N, Andrews CN, Holzhausen M, et al. Role for protease activity in visceral pain in irritable bowel syndrome. Journal of Clinical Investigation. 2007;117(3):636–647. doi: 10.1172/JCI29255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Sato K, Ninomiya H, Ohkura S, Ozaki H, Nasu T. Impairment of PAR-2-mediated relaxation system in colonic smooth muscle after intestinal inflammation. British Journal of Pharmacology. 2006;148(2):200–207. doi: 10.1038/sj.bjp.0706717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Bueno L, Fioramonti J. Protease-activated receptor 2 and gut permeability: a review. Neurogastroenterology and Motility. 2008;20(6):580–587. doi: 10.1111/j.1365-2982.2008.01139.x. [DOI] [PubMed] [Google Scholar]
- 155.Róka R, Ait-Belgnaoui A, Salvador-Cartier C, et al. Dexamethasone prevents visceral hyperalgesia but not colonic permeability increase induced by luminal protease-activated receptor-2 agonist in rats. Gut. 2007;56(8):1072–1078. doi: 10.1136/gut.2006.115352. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Mulè F, Pizzuti R, Capparelli A, Vergnolle N. Evidence for the presence of functional protease activated receptor 4 (PAR4) in the rat colon. Gut. 2004;53(2):229–234. doi: 10.1136/gut.2003.021899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Augé C, Balz-Hara D, Steinhoff M, Vergnolle N, Cenac NS. Protease-activated receptor-4 (PAR4): a role as inhibitor of visceral pain and hypersensitivity. Neurogastroenterology and Motility. 2009;21(11):1189–e107. doi: 10.1111/j.1365-2982.2009.01310.x. [DOI] [PubMed] [Google Scholar]
- 158.Annaházi A, Dabek M, Gecse K, et al. Proteinase-activated receptor-4 evoked colorectal analgesia in mice: an endogenously activated feed-back loop in visceral inflammatory pain. Neurogastroenterology and Motility. 2012;24(1):76–85. doi: 10.1111/j.1365-2982.2011.01805.x. [DOI] [PubMed] [Google Scholar]
- 159.Dabek M, Ferrier L, Annahazi A, et al. Intracolonic infusion of fecal supernatants from ulcerative colitis patients triggers altered permeability and inflammation in mice: role of cathepsin G and protease-activated receptor-4. Inflammatory Bowel Diseases. 2011;17(6):1409–1414. doi: 10.1002/ibd.21454. [DOI] [PubMed] [Google Scholar]
- 160.Dabek M, Ferrier L, Roka R, et al. Luminal cathepsin G and protease-activated§ receptor 4: a duet involved in alterations of the colonic epithelial barrier in ulcerative colitis. The American Journal of Pathology. 2009;175(1):207–214. doi: 10.2353/ajpath.2009.080986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Annaházi A, Gecse K, Dabek M, et al. Fecal proteases from diarrheic-IBS and ulcerative colitis patients exert opposite effect on visceral sensitivity in mice. Pain. 2009;144(1-2):209–217. doi: 10.1016/j.pain.2009.04.017. [DOI] [PubMed] [Google Scholar]
- 162.Yun LW, Decarlo AA, Hunter N. Blockade of protease-activated receptors on T cells correlates with altered proteolysis of CD27 by gingipains of Porphyromonas gingivalis. Clinical and Experimental Immunology. 2007;150(2):217–229. doi: 10.1111/j.1365-2249.2007.03488.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Jiang B, Grage-Griebenow E, Csernok E, et al. The role of proteinase 3 (PR3) and the protease-activated receptor-2 (PAR-2) pathway in dendritic cell (DC) maturation of human-DC-like monocytes and murine DC. Clinical and Experimental Rheumatology. 2010;28(1) supplement 57:S56–S61. [PubMed] [Google Scholar]
- 164.D’Andrea MR, Derian CK, Santulli RJ, Andrade-Gordon P. Differential expression of protease-activated receptors-1 and -2 in stromal fibroblasts of normal, benign, and malignant human tissues. The American Journal of Pathology. 2001;158(6):2031–2041. doi: 10.1016/S0002-9440(10)64675-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Popovic M, Laumonnier Y, Burysek L, Syrovets T, Simmet T. Thrombin-induced expression of endothelial CX3CL1 potentiates monocyte CCL2 production and transendothelial migration. Journal of Leukocyte Biology. 2008;84(1):215–223. doi: 10.1189/jlb.0907652. [DOI] [PubMed] [Google Scholar]
- 166.Kamio N, Hashizume H, Nakao S, Matsushima K, Sugiya H. Plasmin is involved in inflammation via protease-activated receptor-1 activation in human dental pulp. Biochemical Pharmacology. 2008;75(10):1974–1980. doi: 10.1016/j.bcp.2008.02.018. [DOI] [PubMed] [Google Scholar]
- 167.Fiorucci S, Antonelli E, Distrutti E, et al. PAR1 antagonism protects against experimental liver fibrosis. Role of proteinase receptors in stellate cell activation. Hepatology. 2004;39(2):365–375. doi: 10.1002/hep.20054. [DOI] [PubMed] [Google Scholar]
- 168.Matos NA, Silva JF, Matsui TC, et al. Mast cell tryptase induces eosinophil recruitment in the pleural cavity of mice via proteinase-activated receptor 2. Inflammation. 2013;36(6):1260–1267. doi: 10.1007/s10753-013-9664-5. [DOI] [PubMed] [Google Scholar]
- 169.Lohman RJ, Cotterell AJ, Barry GD, et al. An antagonist of human protease activated receptor-2 attenuates PAR2 signaling, macrophageactivation, mast cell degranulation, and collagen-induced arthritis in rats. The FASEB Journal. 2012;26(7):2877–2887. doi: 10.1096/fj.11-201004. [DOI] [PubMed] [Google Scholar]