

Abstract
Upon allergen challenge, DC subsets are recruited to target sites under the influence of chemotactic agents; however, details pertinent to their trafficking remain largely unknown. We investigated the kinetic profiles of blood and skin-infiltrating DC subsets in twelve atopic subjects receiving six weekly intradermal allergen and diluent injections. The role of activin-A, a cytokine induced in allergic and tissue repair processes, on the chemotactic profiles of DC subsets was also examined. Plasmacytoid (pDCs) and conventional DCs (cDCs) were evaluated at various time-points in the blood and skin. In situ activin-A expression was assessed in the skin and its effects on chemokine receptor expression of isolated cDCs were investigated. Blood pDCs were reduced 1 h after challenge, while cDCs decreased gradually within 24 h. Skin cDCs increased significantly 24 h after the first challenge, inversely correlating with blood cDCs. Activin-A in the skin increased 24 h after the first allergen challenge and correlated with infiltrating cDCs. Activin-A increased the CCR10/CCR4 expression ratio in cultured human cDCs. DC subsets demonstrate distinct kinetic profiles in the blood and skin especially during acute allergic inflammation, pointing to disparate roles depending on each phase of the inflammatory response. The effects of activin-A on modulating the chemotactic profile of cDCs suggest it may be a plausible therapeutic target for allergic diseases.
1. Introduction
Allergic diseases, such as asthma and atopic dermatitis, are on the rise in western societies and pose a significant burden for patients and health care systems. Allergic inflammation relates to excessive T helper type 2 (TH2) cell-mediated responses against innocuous environmental allergens. Dendritic cells (DCs) are known sentinels of the immune system entrusted with the chief task of antigen recognition, presentation, and T cell activation. Different DC subsets with diverse functionalities have been described, depending on the level of expression of specific surface markers, their activation status, and anatomic location. There are two major human DC subsets: myeloid or conventional DCs (cDCs), generally considered immunogenic [1, 2], and plasmacytoid DCs (pDCs), which can exhibit suppressive functions on allergen-driven TH2 cell-mediated responses [3]. Pertinent to the skin, cDCs constitute the major resident DC population in normal human dermis and are characterized by CD1c expression (also known as blood dendritic cell antigen-1 (BDCA)-1) [4, 5]. Plasmacytoid DCs are present in the skin and can be readily identified by expression of CD303, also known as BDCA-2 [6, 7].
Increased numbers of DC subsets have been previously reported in the blood, nasal, and/or lung mucosa of subjects with atopy, allergic rhinitis, and/or asthma [8–11], suggesting that specific DC subgroups are induced in response to allergic inflammation. Nevertheless, few clinical studies have addressed DC kinetics in allergic responses, and these involve mostly allergen inhalation or segmental bronchial allergen challenge [12–17]. Understanding of the trafficking of human DCs upon allergen challenge in vivo is essential for controlling the balance between immunity and tolerance.
DC migration is guided by the rapid upregulation of adhesion molecules and chemokine receptors in response to chemokine and cytokine gradients generated by tissue-resident and infiltrating immune cells. Activin-A is a pleiotropic cytokine-member of the TGF-β superfamily of proteins. It is produced by inflammatory and structural tissue cells and acts as an important regulator of allergic inflammation [18–20] and skin repair processes [21]. In fact, recent studies have revealed potent anti-inflammatory effects of activin-A in vivo during allergen-induced cutaneous sensitization [22]. Activin-A is produced by different DC subsets, promotes DC differentiation, and affects the ability of mature DCs to take up antigens [23]. In addition, activin-A is involved in the differentiation and migration of human Langerhans dendritic cells, mostly through the regulation of chemokines and chemokine receptor networks [24, 25]. Still, the effects of activin-A on shaping the chemotactic responses of human DC subsets during exposure to allergen in vitro remain elusive.
In the present study, we hypothesized that circulating cDCs and pDCs exhibit different kinetic profiles in vivo upon repeated skin allergen challenges, reflecting their distinct roles during acute and established chronic inflammation. To explore this, we used a well-established human in vivo model of repeated skin allergen challenges. Furthermore, we examined activin-A expression in the inflamed skin and explored possible correlations with DC subset infiltration following allergen challenge. Finally, the effects of activin-A on the chemokine receptor profile of allergen stimulated human CD1c+ DCs were also investigated.
2. Methods
2.1. Study Population and Design
Healthy nonsmoking volunteers between the ages of 18 and 55 were initially screened and subjected to skin prick testing (SPT) to a panel of 18 common aeroallergens (HAL Allergy, Benelux). Total blood counts, serum biochemistry, total IgE levels measurements, and spirometry were performed. A total of twelve subjects with strong positive SPT reaction to Dermatophagoides pteronyssinus, the European house dust mite, were enrolled in the study. Subject characteristics are summarized in Table 1. All subjects signed an informed consent form before enrolment. The study was approved by the “Sotiria” Hospital Research Ethics Committee and the Greek National Organization for Medicines and was conducted according to the Declaration of Helsinki principles.
Table 1.
Subject characteristics and summary of results for peripheral and tissue DC subset recruitment.
| Subject 1 | Subject 2 | Subject 3 | Subject 4 | Subject 5 | Subject 6 | Subject 7 | Subject 8 | Subject 9 | Subject 10 | Subject 11 | Subject 12 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gender | M | F | F | M | F | F | M | M | F | F | M | F | ||||||||||||
| Age | 30 | 53 | 23 | 30 | 31 | 28 | 40 | 31 | 31 | 27 | 26 | 28 | ||||||||||||
| IgE | 830 | 1100 | 720 | 54 | 106 | 71 | 19 | 19 | 44 | 235 | 630 | 120 | ||||||||||||
|
| ||||||||||||||||||||||||
| Peripheral dendritic cells (% of total WBC) | ||||||||||||||||||||||||
| cDCs baseline | 0,3986 | 0,209 | 0,2445 | 0,1706 | 0,2122 | 0,2147 | 0,1283 | 0,2223 | 0,2102 | 0,1334 | 0,178 | 0,175 | ||||||||||||
| cDCs 1 h | 0,3602 | 0,2454 | 0,2016 | 0,1544 | 0,1804 | 0,1845 | 0,1421 | 0,1887 | 0,1778 | 0,174 | 0,154 | 0,1444 | ||||||||||||
| cDCs 24 h | 0,3731 | 0,1571 | 0,2424 | 0,1089 | 0,2424 | 0,1908 | 0,1296 | 0,1777 | 0,1132 | 0,1278 | 0,1057 | 0,1011 | ||||||||||||
| cDCs 5 w | 0,2706 | 0,2307 | 0,2923 | 0,1262 | 0,174 | 0,2711 | 0,1527 | 0,1368 | 0,1593 | 0,1124 | 0,1162 | 0,1282 | ||||||||||||
| pDCs baseline | 0,1284 | 0,1071 | 0,2468 | 0,1213 | 0,225 | 0,2554 | 0,1139 | 0,1798 | 0,0553 | 0,1413 | 0,2369 | 0,244 | ||||||||||||
| pDCs 1 h | 0,1102 | 0,105 | 0,1895 | 0,0828 | 0,1708 | 0,1681 | 0,1175 | 0,1105 | 0,0581 | 0,0524 | 0,167 | 0,104 | ||||||||||||
| pDCs 24 h | 0,0859 | 0,1313 | 0,2099 | 0,0732 | 0,144 | 0,1982 | 0,119 | 0,1503 | 0,0647 | 0,1526 | 0,212 | 0,2059 | ||||||||||||
| pDCs 5 w | 0,0214 | 0,0013 | 0,2119 | 0,066 | 0,3738 | 0,1559 | 0,1715 | 0,078 | 0,1308 | 0,0907 | 0,2113 | 0,198 | ||||||||||||
|
| ||||||||||||||||||||||||
| Tissue dendritic cells (cells per mm2) | ||||||||||||||||||||||||
| ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | ALL | DIL | |
|
| ||||||||||||||||||||||||
| cDCs 24 h | 5 | 3 | 24 | 8 | 15 | 7 | 34 | 12 | 12 | 5 | 21 | 13 | 58 | 25 | 24 | 19 | 17 | 5 | 22 | 5 | 23 | 9 | 30 | 8 |
| cDCs 5 w | 14 | 6 | 16 | 8 | 20 | 13 | 10 | 13 | 23 | 10 | 30 | 8 | 25 | 15 | 15 | 48 | 29 | 17 | 25 | 5 | 27 | 6 | 14 | 6 |
| pDCs 24 h | 12 | 10 | 6 | 8 | 8 | 5 | 12 | 15 | 4 | 4 | 24 | 14 | 5 | 2 | 5 | 3 | 3 | 2 | 18 | 10 | 7 | 3 | 12 | 10 |
| pDCs 5 w | 10 | 8 | 12 | 5 | 4 | 25 | 8 | 6 | 9 | 6 | 11 | 5 | 14 | 10 | 10 | 16 | 6 | 3 | 4 | 1 | 5 | 4 | 5 | 8 |
All participants were free of atopic symptoms for at least one month before and during the study. Participants had no history of infection and had not received any treatment with oral/inhaled corticosteroids, antihistamines, anti-IgE, or antileukotrienes for one month before and during the study. Each subject received a total of six weekly intradermal injections of allergen, to which they were sensitive, and diluent on the extensor aspect of the left and right forearms, respectively. Skin biopsies were obtained 24 h after the first challenge and 24 h after the last challenge at both allergen and diluent sites. Whole blood was taken before the first challenge, 1 h after the first challenge, and just before each biopsy acquisition. The study design is depicted in Figure 1.
Figure 1.
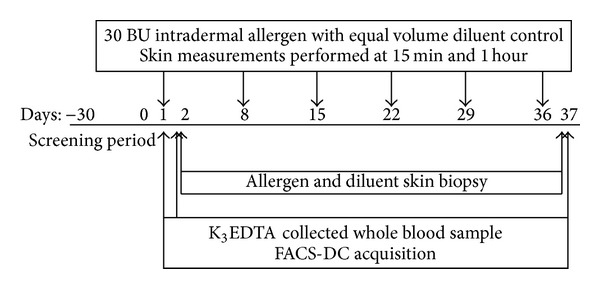
Study design. Flow diagram showing the time-points when the skin challenges with allergen and diluent were performed and when the samples were taken. The dose of Dermatophagoides pteronyssinus administered was 30 BU at the allergen site with equal volume of diluent at the opposing site every week (solid arrows). A screening period of 2 weeks was introduced to clinically verify that subjects did not exhibit seasonal allergic symptoms or an upper/lower respiratory infection.
2.2. In Vivo Allergen Challenges
A 21-gauge needle was used to deliver 100 μL (30 BU) of allergen aqueous solution (Dermatophagoides pteronyssinus, Allergopharma Joachim Ganzer KG, Germany) intradermally at the same site on the extensor aspect of the left forearm. The same diluent volume (0.9% sodium chloride) was administered in the same manner on the extensor aspect of the right forearm. In this way, each patient served as his/her own control. The challenge tests were performed at the same time each day. Measurements of skin reactions were performed at 15 min, 1 h, and 6 h, as previously described [26], by a single investigator throughout the study.
2.3. Preparation of Skin Biopsies
Skin specimens were obtained from the centre of both allergen and diluent sites in each subject using a 4 mm punch biopsy tool (Stiefel Laboratories). Local anaesthesia was induced by injecting subcutaneously 0.5 mL of 2% lidocaine hydrochloride. Tissue samples from the same arm after the last challenge were taken 0.8 cm apart. After appropriate orientation and handling, specimens were cut in half and one of the resulting tissue samples was embedded in OCT medium and snap-frozen in isopentane (BDH Chemicals) precooled in liquid nitrogen, while the other was fixed in formalin overnight and embedded in paraffin. Frozen specimens were stored at 80°C.
2.4. Immunohistochemistry
Paraffin sections 4-5 mm thick were deparaffinised in xylol, rehydrated in graded alcohol series, and immunostained for activin-A, as previously described [19]. In brief, the alkaline phosphatase-antialkaline phosphatase method was used, and specific antibody binding was visualized using Vectastain ABC-AP kits and the Fast-Red chromogen (rabbit anti-goat IgG, AK-5005 Vector Laboratories). All incubations were performed at room temperature. Washes were performed in PBS. Normal rabbit serum (10%) was used to reduce nonspecific binding. An affinity-purified polyclonal goat antibody against human activin-A and a goat IgG control antibody (which served as negative control) were used as primary antibodies (AF338 and AB-108-C, resp., R&D Systems).
2.5. Immunofluorescence Staining
To determine the numbers of cDCs and pDCs in the skin, we applied an immunostaining technique to serial cryosections (6 μm). Slides were warmed up at room temperature (RT) for 30 min, fixed in ice cold (4°C) acetone for 5 min, air dried for 45 min, and washed in PBS. Sections were then blocked with normal goat serum (S-1000, Vector Laboratories) and incubated overnight at 4°C with unconjugated mouse anti-human CD1c (BDCA-1) for the determination of cDCs or mouse anti-human CD303 (BDCA-2) for pDCs (130-090-695 and 130-090-690, resp., Miltenyi Biotec). A secondary goat anti-mouse antibody conjugated with AlexaFluor568 or goat anti-mouse conjugated with AlexaFluor488 (A21134 and A21121, resp., Molecular Probes) was applied for 30 min at RT. Negative controls were obtained by substitution of the primary antibody with the same concentration of the corresponding IgG isotype control (MAB003 and MAB002, resp., R&D Systems). Slides were rinsed and then counterstained with Hoechst (H3569, Molecular Probes).
2.6. Confocal Laser Microscopy
Skin sections processed with immunofluorescence staining were examined using a confocal laser microscope. Fluorescent images of the tissue sections were acquired, using a Leica DMI6000 inverted microscope with DIC optics (Leica TCS SP5). Hoechst was excited by the 405 nm laser diode and the fluorescence was collected using a long-pass (LP) 420 emission filter; the green (AF488) antibody was excited by the 488 nm argon ion laser line and the fluorescence was collected using a band-pass (BP) 505–530 emission filter; the red (AF568) antibody was excited by the 568 nm green helium-neon laser line and the fluorescence was collected using an LP560 emission filter. Tissue sections were visualised using the confocal microscope at 1,024 × 1,024-pixel resolution through a HCX PL APO CS 20.0 × 0.70 DRY UV with eight-times averaging in sequential scanning (multitrack) mode with the pinhole set to obtain an optical section of approximately 1 mm in all channels.
2.7. Quantification of DCs and Activin-A Positive Cells
Images of tissue sections were recorded using a computerized image analysis system (AxioVision, Carl Zeiss). Cells stained for activin-A were counted in the epidermis and dermis by using an AxioScopeA1 light microscope (Carl Zeiss). Results were normalized to the area of the epithelium and to the length of the basement membrane and expressed as number of cells per mm2. Quantitative measurements of BDCA-1 and BDCA-2 positive cells in skin tissue specimens were performed as previously described [5, 7]. DCs that showed positive staining were counted to a dermis depth of 500 μm. Stained DCs were regarded positive when showing DC morphology and were expressed as number of cells per mm2 of dermis area. Activin-A, BDCA-1, and BDCA-2 staining was performed in serial tissue sections, which were coded and examined in a blinded manner at the end of the study by two investigators. The intra- and interobserver variations were calculated to be less than 4% and 8%, respectively.
2.8. CD1c+ DC Isolation and In-Vitro Stimulation
Peripheral blood mononuclear cells (PBMCs) were obtained by Ficoll gradient centrifugation and CD1c+ DCs were isolated using the CD1c+ (BDCA-1+) human dendritic cell isolation kit, (Miltenyi Biotec 130-090-506). Briefly, in a first step, PBMCs were magnetically labeled with CD19 microbeads. Using this approach, CD19+ cells were depleted by separation over a MACS column which was placed in the magnetic field of a MACS separator. In a second step, CD1c+ DCs, in the B cell-depleted flow-through fraction, were magnetically labeled with CD1c-biotin and antibiotin microbeads. Upon separation, the labeled CD1c+ DCs were retained within the column and eluted after removing the column from the magnetic field. This two-step procedure allows the isolation of purified CD1c+ DCs (with a purity >80%). A total of 5 × 104 CD1c+ DCs were placed in 96 flat-bottom plates and cultured for 24 h with (a) medium, (b) 100 ng/mL LPS, or (c) 1 μg/mL Dermatophagoides pteronyssinus in the presence or absence of 50 ng/mL recombinant activin-A (R&D). LPS is a TLR4 ligand extensively used for the activation of DCs and served as a positive control [27]. Following 24 h of culture, cells were stained with fluorochrome-conjugated antibodies against human CCR4 (CD194), CCR6 (CD196), CCR9 (CD199), CXCR3 (CD183), and CCR10 (all from Biolegend) and analyzed by flow cytometry.
2.9. Flow-Cytometric Analysis
Peripheral blood was drawn and processed for flow-cytometric analysis within 30 min of collection. The identification of peripheral DC subsets was based on two different three-color assays (IOTest PN-A07405 “myeloid subset” and PN-A07412 “plasmacytoid subset,” Immunotech, Marseille, France) and was performed as previously described [28]. Whole blood (100 μL) was mixed with 20 μL of each monoclonal antibody or appropriate labelled isotype control cocktail, vortexed for 1 sec, and incubated for 30 min at RT, protected from light. The samples were lysed and fixed (VersaLyse PN-IM3648 and fixative solution PN-IM3515, Beckman Coulter, Krefeld, Germany), washed, resuspended in PBS, and kept on ice until flow cytometric analysis. At least 300000 events were acquired on a dual-laser Beckman-Coulter FC500. Further analysis of the flow-cytometric raw data was performed with the FLOWJO software (Tree Star Inc., USA). The gating strategy used for the analysis of DC subsets is provided in Figure 2.
Figure 2.

Early- and late-phase reactions. Mean diameter of skin reactions after each allergen challenge with Dermatophagoides pteronyssinus measured at 15 min and 1 h (a), as well as 6 h (b), is presented in mm. No statistical significant differences were observed between challenges or at different time-points. Values are expressed as mm ± SEM.
2.10. Statistical Analysis
Data are expressed as median with interquartile range (first and third quartiles) unless specified otherwise. Normality was assessed using D' Agostino—Pearson omnibus normality tests. Data differed from normal distribution and were, thus, analyzed with non-parametric statistics. Statistical analysis of skin reactions measured at 15 min, 1 h and 6 h after each challenge was performed by two-way ANOVA mixed model, followed by Bonferroni post hoc analysis. DC kinetics in peripheral blood as well as the analysis of the number of tissue DC subsets at different time-points were performed by repeated measures nonparametric one-way ANOVA (Friedman test), followed by Dunn's post hoc analysis for all pairs. The correlation between DC subsets in skin tissue and peripheral blood at different time-points, as well as between activin-A tissue expression and DC subsets in the skin, was performed by nonparametric Spearman correlation tests. Two-group analysis of the effects of activin-A's on chemokine receptor expression by allergen-stimulated CD1c+ DCs was performed by the Mann-Whitney U test. A P value of ≤0.05 was considered statistically significant. Data storage and analysis were performed with statistical analysis software (GraphPad Prism v5, GraphPad Inc., CA, USA).
3. Results
3.1. Early- and Late-Phase Allergen-Induced Cutaneous Reactions
The mean diameter of skin reactions after each allergen challenge measured at 15 min, 1 h, and 6 h (Figure 2). Statistical analysis showed no diameter differences between challenges or at different time-points, minimizing the possibility of induction of tolerance during the in vivo protocol. No indurations were observed at the diluent site. No diluent and/or allergen-related adverse events were observed in any of the subjects.
3.2. DC Subsets Exhibit Different Kinetic Patterns in the Peripheral Blood Following Repeated Allergen Skin Challenges In Vivo
Circulating cDCs and pDCs were assessed before allergen challenge, 1 h and 24 h after the first allergen challenge, and 24 h after the sixth challenge (Figures 3(a) and 3(b), resp.). Interestingly, our data revealed significant differences for both cDC and pDC numbers in the peripheral blood of atopics at distinct time-points following in vivo allergen skin challenge, compared to baseline (P < 0.05, resp.). Post hoc analysis revealed that changes in the pDC population were very rapid, as they were significantly decreased 1 h after the first allergen challenge and returned to baseline after 24 h (Figure 3(c)). On the other hand, cDCs started to decrease by the first hour after challenge and continued to decrease, reaching significantly lower levels at 24 h, with a later-on trend to return to baseline after 5 weeks (Figure 3(d)).
Figure 3.
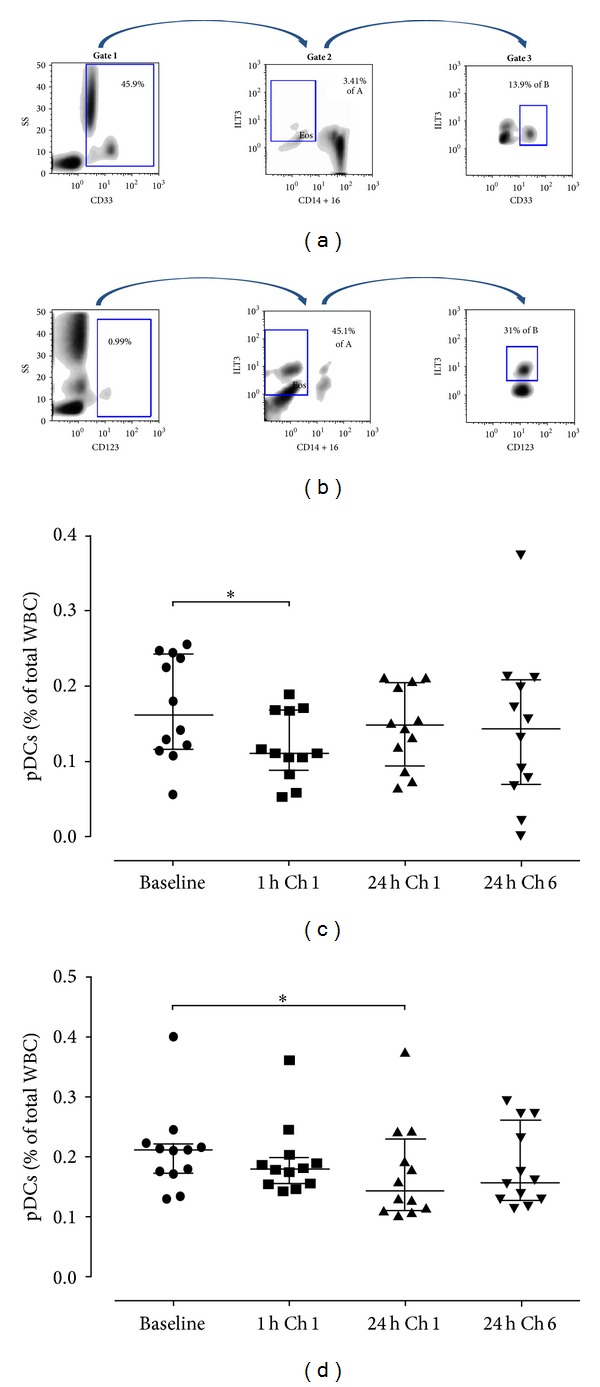
DC subsets exhibit different kinetic patterns in the peripheral blood following repeated allergen skin challenges in vivo. Gating strategies utilized to identify cDC and pDC subsets in the peripheral blood by flow cytometry. (a) For cDC identification, a 3-step analysis was performed. Initially, CD33pos cells were selected (Gate 1) to differentiate between mature lymphoid cells or lymphoid precursors (CD33neg) from other cells of myeloid origin that include cDCs. Next, all CD(14+16)dim to neg/ILT3pos cells are selected in Gate 2 to exclude monocytes, macrophages, NK cells, and neutrophils. Gate 3 is drawn around CD33bright/ILT3pos cells, so cDCs are characterized as CD33bright/ILT3pos/CD(14+16)dim to neg. (b) Regarding pDCs, initially all CD123pos cells are selected (Gate 1) and then gated on the basis of CD(14+16)neg expression (Gate 2) to exclude monocytes, lymphocytes, and most granulocytes. Gate 3 is drawn around CD123bright/ILT3bright cells, strictly selecting pDCs and excluding basophils, so pDCs are characterized as CD123bright/ILT3bright/CD(14+16)neg. Representative FACS plots are shown. The percentages of pDCs (c) and cDCs (d) in the peripheral blood at baseline and following in vivo allergen challenges are shown. Data are expressed as median with interquartile range (first and third quartiles).
3.3. Conventional DCs Are Recruited to the Skin Early Following In Vivo Allergen Challenge
DC subsets were examined in the skin at the allergen and diluent sites 24 h after the first and sixth challenges, time-points at which DC infiltration has been reported to peak [29]. Our findings demonstrated that pDCs and cDCs were mostly located in the subepithelium within 150 μm range from the epidermis but occasionally also extended deeper into the dermis (Figures 4(a)–4(c) and 5(a)–5(c), resp.). Pertinent to pDC recruitment, there was a trend for increased pDC numbers at the allergen site after the final allergen challenge (Figure 4(d)). In contrast, one-way ANOVA revealed significant differences between the distinct time-points for cDCs (P = 0.0015), with post hoc analysis demonstrating a significant increase at the allergen site 24 h after the first allergen skin challenge, compared to the diluent site (P < 0.05, Figure 5(d)). Although a trend for increased cDCs was also observed at the allergen site 24 h after the sixth challenge compared to the diluent site, this difference was not significant according to post hoc analysis (Figure 5(d)). Importantly, we observed that the percentages of circulating cDCs inversely correlated with those of skin-infiltrating cDCs (P = 0.0202; r = −0.6667, Figure 5(e)). No correlations were observed between DC subset numbers at the skin site and the magnitude of the early- or late-phase skin reactions (data not shown). In summary, these data provide evidence that cDCs are summoned to the inflamed skin site early following in vivo allergen exposure.
Figure 4.
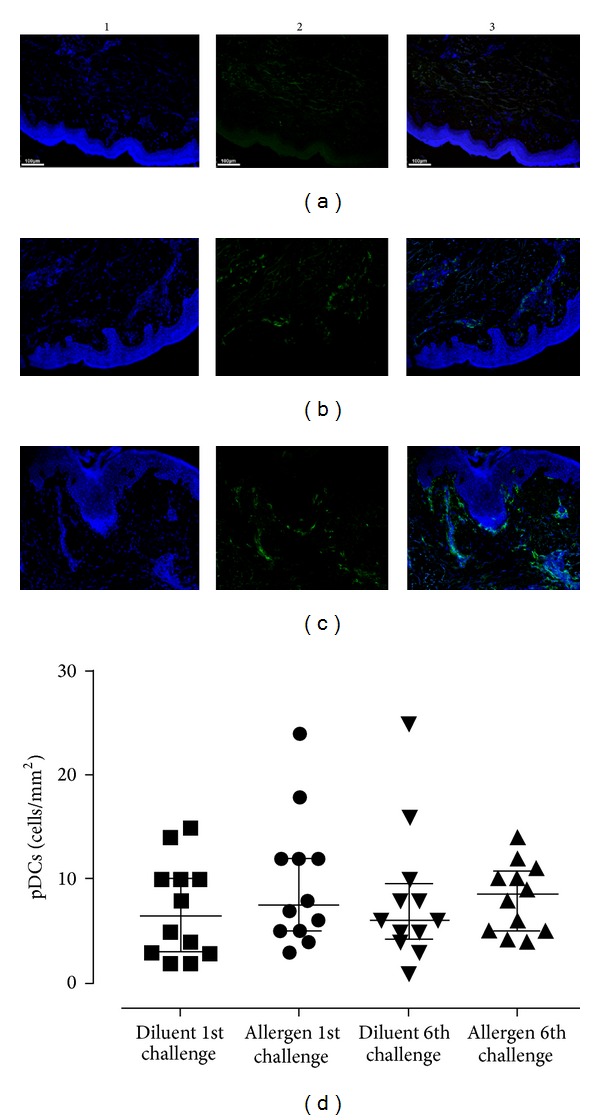
Plasmacytoid DC kinetics in the skin upon in vivo allergen challenge. Immunofluorescence staining was performed on skin biopsies and examined by confocal microscopy. Counterstaining was performed with Hoechst to visualize nuclear DNA (blue, column 1). PDCs were stained with a monoclonal antibody against BDCA-2 and envisioned with a secondary goat anti-mouse antibody conjugated with AF488 (green, column 2). Column 3 is the result of merging columns 1 and 2. Representative microphotographs (×100) of pDCs are shown 24 h after the first challenge at the diluent (a) and allergen sites (b) and after the sixth challenge at the allergen site (c). PDC numbers were not significantly altered between different time-points, although a trend for increased pDCs was observed after the sixth allergen challenge (d). Data are expressed as median with interquartile range (first and third quartiles). *P < 0.05, WBC: whole blood cells.
Figure 5.

Conventional DCs are recruited early to the skin upon in vivo allergen challenge. Immunofluorescence staining and data analysis were performed as described in Figure 4. Counterstaining was performed with Hoechst (blue, column 1). CDCs were stained with a monoclonal antibody against BDCA-1 and envisioned with a secondary goat anti-mouse antibody conjugated with AF568 (red, column 2). Column 3 is the result of merging columns 1 and 2. Representative microphotographs (×100) of cDCs are shown 24 h after the first challenge at the diluent (a) and allergen sites (b) and after the sixth challenge at the allergen site (c). Tissue cDCs significantly increased 24 h after the first allergen challenge compared to the diluent, and their numbers remained high after the sixth allergen challenge, although the difference was not significant (d). A significant inverse correlation was found between blood and skin tissue-infiltrating cDCs 24 h after the first allergen challenge (P = 0.0202; r = −0.667) (e). Data are expressed as median with interquartile range (first and third quartiles). *P < 0.05, WBC: whole blood cells.
3.4. Activin-A Is Increased in the Skin after Repeated In Vivo Allergen Challenges and Correlates with cDC Numbers
Our data showed that activin-A expression in normal skin (diluent site) was minimal and mostly present in a scattered fashion at the basal cells of the epidermis with very low levels observed in the dermis (Figure 6(a)). Interestingly, after the first in vivo allergen skin challenge, activin-A expression was more prominent and intense in the epidermis, as well as, in infiltrating inflammatory cells in the dermis (Figures 6(b) and 6(c)). Activin-A was also observed at low levels in the connective tissue. Following the sixth allergen challenge, activin-A in the epidermis and dermis at the allergen site was higher compared to the diluent site, but to a lesser extent, and intensity compared to that after the first allergen challenge (Figure 6(d)). As cDCs showed a similar kinetic pattern to that of activin-A following in vivo allergen challenge, we investigated the existence of possible correlations. Interestingly, the number of activin-A+ cells correlated with the number of BDCA-1+ cDCs at the 24 h time-point after allergen challenge (P = 0.0219; r = 0.662, Figure 6(e)). Together, these findings suggest a role for activin-A at the inflamed skin site during in vivo allergen challenge.
Figure 6.

Activin-A is increased in the skin after allergen challenge and correlates with BDCA+ cDCs. Activin-A in the skin was analysed using the alkaline phosphatase-antialkaline phosphatase method (red colour). (a) Activin A in normal skin (diluent site) was minimal and mostly located in a scattered fashion in the basal cells of the epidermis (black arrows) with minimal expression in the dermis (red arrows) (×100). (b) After the first allergen challenge, activin-A was more prominent and intense in the epidermis (black arrows) and in infiltrating inflammatory cells (red arrows) in the dermis (×100), also shown in (c) at higher magnification (×200). (d) Quantification of activin-A+ cells showed significantly higher expression at the allergen site 24 h after the first challenge compared to the diluent site. (e) The numbers of activin-A+ cells correlated with the numbers of BDCA-1+ cDCs in the dermis 24 h after allergen challenge (P = 0.0219; r = 0.662). Data are expressed as median with interquartile range (first and third quartiles). *P < 0.001.
3.5. Activin-A Modulates the Chemokine Receptor Profile of Allergen-Stimulated CD1c+ DCs
Previous studies have demonstrated that human cDCs express higher levels of activin-A type I and II signalling receptors compared to pDCs and respond actively to the ligand [24]. In view of our findings showing a strong correlation between activin-A expression and cDC trafficking at the inflamed skin site upon allergen encounter, we hypothesized that activin-A may affect the cDC chemotactic profile. To address this, we examined the effects of activin-A on chemokine receptor expression by cDCs isolated from the peripheral blood of atopics during stimulation with allergen in vitro. No significant differences were observed in the levels of CXCR3, CCR6, or CCR9 on cDCs upon activin-A treatment (Figure 7(a)). Activin-A, however, increased the expression of CCR10 on CD1c+ DCs, concomitant with a decrease in CCR4, as compared to control (PBS) (Figure 7(a)). In fact, activin-A significantly increased the ratio of CCR10/CCR4 expressing cDCs (Figure 7(b)). Chemokine receptor levels were very low in nonstimulated cDCs (medium alone) and increased upon LPS treatment (data not shown). Overall, these data suggest that activin-A may be involved in retaining cDCs in the skin after allergen challenge through the regulation of CCR4 and CCR10 expression.
Figure 7.
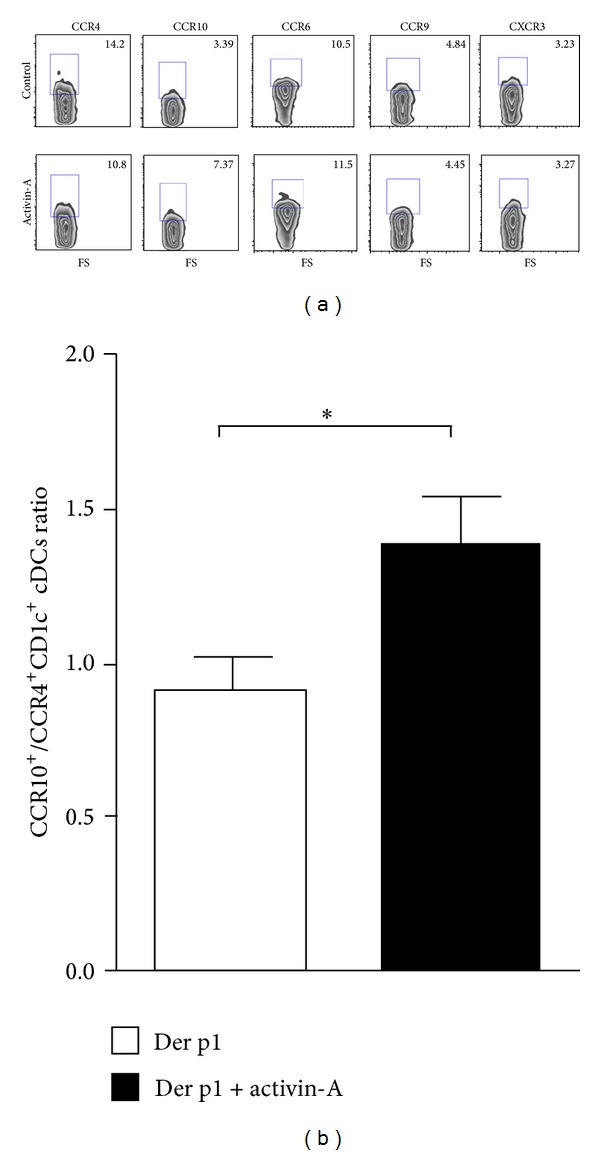
Activin-A modifies the chemokine receptor profile of CD1c+ cDCs towards a skin-homing phenotype. CD1c+ cDCs were isolated from the peripheral blood of individuals with atopy to Dermatophagoides pteronyssinus (Der p1), cultured for 24 h with 1 μg/mL Der p1 in the presence of 50 ng/mL recombinant activin-A or PBS (control). DCs were stained with fluorochrome-conjugated antibodies against human CCR4, CCR10, CCR6, CCR9, and CXCR3 and analysed by flow-cytometry. (a) Activin-A induced an increase in CCR10 levels, concomitant with a decrease in CCR4 on CD1c+ cDCs during stimulation with Der p1 in vitro. No differences were observed regarding the expression of CCR6, CCR9, and CXCR3 in Der p1-stimulated CD1c+ cDCs in the presence or absence of activin-A. (b) Activin-A significantly increased the ratio of CCR10/CCR4 expressing cDCs. Data are representative of two independent experiments. *P < 0.05.
4. Discussion
Human DC kinetics have been examined in the blood [12, 13], induced sputum [14], or bronchial tissue [15] of asthmatics, following a single allergen inhalation or segmental allergen challenge. Still, simultaneous evaluation of DC kinetics at the periphery and the challenged tissue site has not been investigated. A single study has examined DC kinetics in the blood and bronchoalveolar lavage fluid (BALF) in seven asthmatics after a single segmental allergen challenge but did not evaluate DC numbers at the inflamed bronchial tissue [16]. Jahnsen et al. investigated DC kinetics in the periphery and the challenged nasal tissue, but the study was limited to pDCs [17]. To our knowledge, this is the first human study evaluating both cDC and pDC kinetics concomitantly in the periphery and at the inflamed tissue site after repeated in vivo allergen skin challenges in atopic subjects, encapsulating both the initial and chronic stages of allergic inflammation.
Experimental skin allergen challenge has been widely used to examine the cellular processes associated with early- and late-phase allergic reactions [30] and to study the effects of repeated allergen exposure in the airways [26]. Moreover, these models imitate the chronicity of allergen exposure in a more standardized fashion and, as late-phase skin reactions exhibit similar histopathological inflammatory patterns to that of the airways [26], they can be used to recapitulate chronic allergic airway inflammation [31]. Furthermore, these models demand only atopic status as a prerequisite, which usually requires no treatment. In asthma, however, treatment with corticosteroids is often warranted, which can act as a confounding factor by decreasing pDC absolute numbers and cytokine responses upon in vivo bronchial allergen challenge [32]. Moreover, steroid treatment has been recently shown to increase the capacity of GILZ+ circulating DCs to activate allergen-specific IL-10 Tregs in allergic patients [33]. In addition, the difficulties and increased patient risk associated with performing multiple bronchial allergen challenges in conjunction with consecutive endobronchial biopsies represent critical restraining factors. Still, our study presents certain limitations, mainly pertaining to the fact that we did not perform challenges with an irrelevant allergen nor did we include healthy nonatopic controls to confirm that our findings were due to allergen exposure per se.
There has been contradicting evidence pertinent to the effects of repeated allergen challenges in the nose, airways, and skin of atopics. In certain studies, multiple challenges led to tolerance while in others led to enhanced inflammatory cell migration [30]. The time interval between skin challenges, the type of allergen used, and the dosing scheme is crucial in determining priming versus tolerance. Intervals longer than than one week seem to induce small late-phase reactions [26], while challenges with house dust mite often result in an increase in late skin reactions compared to grass pollen [30]. We observed early- and late-phase skin reactions after every challenge with house dust mite and detected no differences between challenges, as also shown by other investigators [31]. These data suggest that the possibility of desensitization is minimal.
Plasmacytoid DCs are primarily observed in peripheral blood, but they also migrate to lymphoid organs or tissues during inflammation [34]. The finding that there were no differences in the numbers of skin pDCs after repeated allergen challenges was rather unexpected. Previous studies of bronchial allergen challenge in asthmatics have also failed to show an accumulation of pDCs in the bronchial tissue [15]. One could argue that single allergen challenge models could be insufficient for the recruitment of pDCs; still, we did not observe significant changes in pDCs even after six weekly allergen challenges, although a trend was detected. The observation that pDC levels were decreased in the periphery within one hour after allergen challenge is suggestive of a rapid recruitment of these cells to the exposed skin site, possibly occurring long before 24 h. Moreover, pDCs have been shown to respond selectively to homeostatic chemokines, demonstrating an inherent tendency to migrate to secondary lymphoid organs rather than to inflammatory tissue sites [35]. It is, therefore, possible that peripheral pDCs were recruited predominantly to local draining lymph nodes (DLNs).
Conventional DCs, in contrast to pDCs, circulate in the blood or reside in peripheral tissues [34] and are quickly depleted from the circulation following allergen inhalation in asthmatics [12]. Upon local allergen challenge, cDCs have been reported to accumulate within the bronchial mucosa [15]. It is, however, not clear whether this is due to an enhanced cDC migration from the periphery, the differentiation of DCs in situ, or decreased migration of tissue-resident DCs towards DLNs upon allergen encounter [36]. Our findings demonstrate that cDCs accumulate at the inflamed skin site within 24 h after local allergen challenge and inversely correlate with their numbers in the periphery at the same time-point. These data support the notion that cDCs are recruited from the bloodstream to the skin early following allergen exposure. Still, this does not preclude the possibility of local DC differentiation from precursor populations. In fact, increased numbers of CD34+ progenitor cells have been noted in the airways of asthmatics [37]. In accordance with our findings, cDCs have also been reported to decrease in the periphery and increase in BALF 24 h after segmental allergen challenge in mild asthmatics; however, possible correlations were not examined [16]. Corrigan et al. also reported increased numbers of C11c+ BDCA-1+ cDCs bearing the thymic stromal lymphopoietin receptor in the skin, 24 h after allergen challenge, and their data supported that these cells were recruited from the circulation rather being differentiated de novo [29].
In order to investigate tissue mDCs, we used BDCA-1 as a cellular marker in the skin. CD1a has also been used to identify cDCs in normal skin; however, CD1c (BDCA-1) is considered a more useful marker as it colocalizes with nearly all CD11c+ cells [4, 5]. Upon inflammation, an additional population of infiltrating inflammatory DCs has been reported that is different from steady state dermal cDCs and is considered their potential precursor [4, 38]. In the acute phase of atopic dermatitis, these cells coexpress CD1b/c and produce chemokines that attract TH2 cells [4, 38, 39]. It is unknown whether the cDCs examined in the present studies contain also a population of these highly inflammatory DCs. Delineation of the precise phenotypic and functional properties of cDCs recruited to the skin upon in vivo allergen challenge will help resolve this question.
Our results also demonstrate that activin-A is greatly increased in the skin after in vivo allergen challenge. Importantly, its expression correlates with the numbers of infiltrating cDCs at 24 h after challenge, suggesting that activin-A may be involved in cDC recruitment to the skin. Activin-A induces DC migration and can act as a differentiating factor for blood cDCs and lung Langerhans cell-like DCs [24, 25]. Although cDCs express both type I and II receptors [40], activin-A has not been shown to exert direct chemotactic effects on these cells. Still, activin-A alters the chemokine and the chemokine receptor profile of human Langerhans dendritic cells [24, 25]. In agreement, we demonstrate that activin-A induced a reciprocal shift up on CCR10, with a shift down on CCR4 on allergen-stimulated CD1c+ cDCs obtained from atopic subjects. Both receptors are involved in DC and T cell trafficking to the skin and inflamed epithelia, including the lungs [41–43]. An inversion of the fold increase ratio in CCR4/CCR10 expression in skin allografts has been previously associated with T cell trafficking and allograft rejection [44]. Moreover, a recent single study has shown that circulating pDCs express increased levels of CCR4 and CCR10 in patients with allergic asthma [32]. Together, these findings prompt the speculation that activin-A may be involved in retaining cDCs at the inflamed skin site, at least partly, through modulating CCR10/CCR4 expression.
In summary, our study highlights key differences in the trafficking of human pDCs and cDCs upon in vivo allergen challenge in the skin, pointing to distinct roles in the allergic response. Importantly, our findings facilitate the understanding of human DC behaviour during allergen encounter in vivo, which is essential for controlling the balance between immunity and tolerance in allergic diseases.
Acknowledgments
The authors are grateful to all subjects participating in the present study. The authors would also like to thank Spyros Spiridakis (Beckman Coulter, Greece) for his technical advice on FACS experiments and analyses and Anna Agapaki, Eleni Rigana, and Stamatis Pagakis (Histochemistry Unit and Biological Imaging Facility, Biomedical Research Foundation of the Academy of Athens) for their expert technical assistance with confocal imaging.
Abbreviations
- AD:
Atopic dermatitis
- BALF:
Bronchoalveolar lavage fluid
- BDCA:
Blood dendritic cell antigen
- CCR:
C-C motif chemokine receptor
- cDC:
Conventional dendritic cell
- CXCR:
CXC motif chemokine receptor
- DC:
Dendritic cell
- Der p1:
Dermatophagoides pteronyssinus
- DLN:
Draining lymph node
- EPR:
Early-phase reaction
- HDM:
House dust mite
- IF:
Immunofluorescence
- IHC:
Immunohistochemistry
- LC:
Langerhans cell
- LPR:
Late-phase reaction
- LPS:
Lipopolysaccharide
- pDC:
Plasmacytoid dendritic cell
- SPT:
Skin prick test.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Authors' Contribution
Stelios Vittorakis and Konstantinos Samitas recruited subjects, conducted skin challenges, performed immunofluorescence/confocal imaging experiments, evaluated biopsy specimens, and wrote the paper. Sofia Tousa, Maria Aggelakopoulou, and Maria Semitekolou performed the in vitro experiments and FACS analyses. Eleftherios Zervas, and Vily Panoutsakopoulou contributed to the study design, assessed and interpreted experimental results, and reviewed the paper. Konstantinos Samitas and Eleftherios Zervas reviewed all final data and performed statistical analyses. Georgina Xanthou and Mina Gaga defined the research theme, designed the experiments, contributed to the assessments and interpretation of results, and wrote the paper. All authors have read and approved the final paper. Stelios Vittorakis and Konstantinos Samitas contributed equally.
References
- 1.Hammad H, de Vries VC, Maldonado-Lopez R, et al. Differential capacity of CD8α + or CD8α − dendritic cell subsets to prime for eosinophilic airway inflammation in the T-helper type 2-prone milieu of the lung. Clinical and Experimental Allergy. 2004;34(12):1834–1840. doi: 10.1111/j.1365-2222.2004.02133.x. [DOI] [PubMed] [Google Scholar]
- 2.Lambrecht BN, Peleman RA, Bullock GR, Pauwels RA. Sensitization to inhaled antigen by intratracheal instillation of dendritic cells. Clinical and Experimental Allergy. 2000;30(2):214–224. doi: 10.1046/j.1365-2222.2000.00818.x. [DOI] [PubMed] [Google Scholar]
- 3.Kool M, van Nimwegen M, Willart MAM, et al. An anti-inflammatory role for plasmacytoid dendritic cells in allergic airway inflammation. Journal of Immunology. 2009;183(2):1074–1082. doi: 10.4049/jimmunol.0900471. [DOI] [PubMed] [Google Scholar]
- 4.Zaba LC, Krueger JG, Lowes MA. Resident and “inflammatory” dendritic cells in human skin. Journal of Investigative Dermatology. 2009;129(2):302–308. doi: 10.1038/jid.2008.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Zaba LC, Fuentes-Duculan J, Steinman RM, Krueger JG, Lowes MA. Normal human dermis contains distinct populations of CD11c +BDCA-1+ dendritic cells and CD163+FXIIIA + macrophages. Journal of Clinical Investigation. 2007;117(9):2517–2525. doi: 10.1172/JCI32282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dzionek A, Sohma Y, Nagafune J, et al. BDCA-2, a novel plasmacytoid dendritic cell-specific type II C-type lectin, mediates antigen capture and is a potent inhibitor of interferon α/β induction. Journal of Experimental Medicine. 2001;194(12):1823–1834. doi: 10.1084/jem.194.12.1823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gregorio J, Meller S, Conrad C, et al. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type i interferons. Journal of Experimental Medicine. 2010;207(13):2921–2930. doi: 10.1084/jem.20101102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Möller GM, Overbeek SE, van Helden-Meeuwsen CG, et al. Increased numbers of dendritic cells in the bronchial mucosa of atopic asthmatic patients: downregulation by inhaled corticosteroids. Clinical and Experimental Allergy. 1996;26(5):517–524. [PubMed] [Google Scholar]
- 9.Spears M, Mcsharry C, Donnelly I, et al. Peripheral blood dendritic cell subtypes are significantly elevated in subjects with asthma. Clinical and Experimental Allergy. 2011;41(5):665–672. doi: 10.1111/j.1365-2222.2010.03692.x. [DOI] [PubMed] [Google Scholar]
- 10.Hagendorens MM, Ebot DG, Schuerwegh AJ, et al. Differences in circulating dendritic cell subtypes in cord blood and peripheral blood of healthy and allergic children. Clinical and Experimental Allergy. 2003;33(5):633–639. doi: 10.1046/j.1365-2222.2003.01649.x. [DOI] [PubMed] [Google Scholar]
- 11.KleinJan A, Willart M, van Rijt LS, et al. An essential role for dendritic cells in human and experimental allergic rhinitis. Journal of Allergy and Clinical Immunology. 2006;118(5):1117–1125. doi: 10.1016/j.jaci.2006.05.030. [DOI] [PubMed] [Google Scholar]
- 12.Upham JW, Denburg JA, O’Byrne PM. Rapid response of circulating myeloid dendritic cells to inhaled allergen in asthmatic subjects. Clinical and Experimental Allergy. 2002;32(6):818–823. doi: 10.1046/j.1365-2222.2002.01375.x. [DOI] [PubMed] [Google Scholar]
- 13.Farrell E, O’Connor TM, Duong M, et al. Circulating myeloid and plasmacytoid dendritic cells after allergen inhalation in asthmatic subjects. Allergy. 2007;62(10):1139–1145. doi: 10.1111/j.1398-9995.2007.01465.x. [DOI] [PubMed] [Google Scholar]
- 14.Dua B, Watson RM, Gauvreau GM, O’Byrne PM. Myeloid and plasmacytoid dendritic cells in induced sputum after allergen inhalation in subjects with asthma. Journal of Allergy and Clinical Immunology. 2010;126(1):133–139. doi: 10.1016/j.jaci.2010.04.006. [DOI] [PubMed] [Google Scholar]
- 15.Jahnsen FL, Moloney ED, Hogan T, Upham JW, Burke CM, Holt PG. Rapid dendritic cell recruitment to the bronchial mucosa of patients with atopic asthma in response to local allergen challenge. Thorax. 2001;56(11):823–826. doi: 10.1136/thorax.56.11.823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Bratke K, Lommatzsch M, Julius P, et al. Dendritic cell subsets in human bronchoalveolar lavage fluid after segmental allergen challenge. Thorax. 2007;62(2):168–175. doi: 10.1136/thx.2006.067793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Jahnsen FL, Lund-Johansen F, Dunne JF, Farkas L, Haye R, Brandtzaeg P. Experimentally induced recruitment of plasmacytoid (CD123high) dendritic cells in human nasal allergy. Journal of Immunology. 2000;165(7):4062–4068. doi: 10.4049/jimmunol.165.7.4062. [DOI] [PubMed] [Google Scholar]
- 18.Kariyawasam HH, Pegorier S, Barkans J, et al. Activin and transforming growth factor-β signaling pathways are activated after allergen challenge in mild asthma. Journal of Allergy and Clinical Immunology. 2009;124(3):454–462. doi: 10.1016/j.jaci.2009.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Semitekolou M, Alissafi T, Aggelakopoulou M, et al. Activin-A induces regulatory T cells that suppress T helper cell immune responses and protect from allergic airway disease. Journal of Experimental Medicine. 2009;206(8):1769–1785. doi: 10.1084/jem.20082603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kariyawasam HH, Semitekolou M, Robinson DS, Xanthou G. Activin-A: a novel critical regulator of allergic asthma. Clinical and Experimental Allergy. 2011;41(11):1505–1514. doi: 10.1111/j.1365-2222.2011.03784.x. [DOI] [PubMed] [Google Scholar]
- 21.Mukhopadhyay A, Sui YC, Lim IJ, Phillips DJ, Phan TT. The role of the activin system in keloid pathogenesis. The American Journal of Physiology—Cell Physiology. 2007;292(4):C1331–C1338. doi: 10.1152/ajpcell.00373.2006. [DOI] [PubMed] [Google Scholar]
- 22.Kypriotou M, Rivero D, Haller S, et al. Activin a inhibits antigen-induced allergy in murine epicutaneous sensitization. Frontiers in Immunology. 2013;4, article 246 doi: 10.3389/fimmu.2013.00246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Scutera S, Riboldi E, Daniele R, et al. Production and function of activin A in human dendritic cells. European Cytokine Network. 2008;19(1):60–68. doi: 10.1684/ecn.2008.0121. [DOI] [PubMed] [Google Scholar]
- 24.Salogni L, Musso T, Bosisio D, et al. Activin A induces dendritic cell migration through the polarized release of CXC chemokine ligands 12 and 14. Blood. 2009;113(23):5848–5856. doi: 10.1182/blood-2008-12-194597. [DOI] [PubMed] [Google Scholar]
- 25.Musso T, Scutera S, Vermi W, et al. Activin A induces langerhans cell differentiation in vitro and in human skin explants. PLoS ONE. 2008;3(9) doi: 10.1371/journal.pone.0003271.e3271e3271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Weller FR, Weller MS, Jansen HM, De Monchy JGR. Effect of local allergen priming on early, late, delayed-phase, and epicutaneous skin reactions. Allergy. 1996;51(12):883–886. doi: 10.1111/j.1398-9995.1996.tb04488.x. [DOI] [PubMed] [Google Scholar]
- 27.Warger T, Osterloh P, Rechtsteiner G, et al. Synergistic activation of dendritic cells by combined Toll-like receptor ligation induces superior CTL responses in vivo. Blood. 2006;108(2):544–550. doi: 10.1182/blood-2005-10-4015. [DOI] [PubMed] [Google Scholar]
- 28.Schaumann F, Müller M, Braun A, et al. Endotoxin augments myeloid dendritic cell influx into the airways in patients with allergic asthma. The American Journal of Respiratory and Critical Care Medicine. 2008;177(12):1307–1313. doi: 10.1164/rccm.200706-870OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Corrigan CJ, Jayaratnam A, Wang Y, et al. Early production of thymic stromal lymphopoietin precedes infiltration of dendritic cells expressing its receptor in allergen-induced late phase cutaneous responses in atopic subjects. Allergy. 2009;64(7):1014–1022. doi: 10.1111/j.1398-9995.2009.01947.x. [DOI] [PubMed] [Google Scholar]
- 30.de Bruin-Weller MS, Weller FR, de Monchy JGR. Repeated allergen challenge as a new research model for studying allergic reactions. Clinical and Experimental Allergy. 1999;29(2):159–165. doi: 10.1046/j.1365-2222.1999.00434.x. [DOI] [PubMed] [Google Scholar]
- 31.Ong YE, Menzies-Gow A, Barkans J, et al. Anti-IgE (omalizumab) inhibits late-phase reactions and inflammatory cells after repeat skin allergen challenge. Journal of Allergy and Clinical Immunology. 2005;116(3):558–564. doi: 10.1016/j.jaci.2005.05.035. [DOI] [PubMed] [Google Scholar]
- 32.Bratke K, Prieschenk C, Garbe K, Kuepper M, Lommatzsch M, Virchow JC. Plasmacytoid dendritic cells in allergic asthma and the role of inhaled corticosteroid treatment. Clinical & Experimental Allergy. 2013;43:312–321. doi: 10.1111/cea.12064. [DOI] [PubMed] [Google Scholar]
- 33.Karaki S, Garcia G, Tcherakian C, et al. Enhanced glucocorticoid-induced leucine zipper in dendritic cells induces allergen-specific regulatory CD4 T-cells in respiratory allergies. Allergy. 2014;69(5):624–631. doi: 10.1111/all.12379. [DOI] [PubMed] [Google Scholar]
- 34.Gros E, Novak N. Cutaneous dendritic cells in allergic inflammation. Clinical & Experimental Allergy. 2012;42:1161–1175. doi: 10.1111/j.1365-2222.2012.03964.x. [DOI] [PubMed] [Google Scholar]
- 35.Penna G, Vulcano M, Sozzani S, Adorini L. Differential migration behavior and chemokine production by myeloid and plasmacytoid dendritic cells. Human Immunology. 2002;63(12):1164–1171. doi: 10.1016/s0198-8859(02)00755-3. [DOI] [PubMed] [Google Scholar]
- 36.Bonasio R, von Andrian UH. Generation, migration and function of circulating dendritic cells. Current Opinion in Immunology. 2006;18(4):503–511. doi: 10.1016/j.coi.2006.05.011. [DOI] [PubMed] [Google Scholar]
- 37.Bossios A, Sjöstrand M, Dahlborn A-K, et al. IL-5 expression and release from human CD34 cells in vitro; ex vivo evidence from cases of asthma and Churg-Strauss syndrome. Allergy. 2010;65(7):831–839. doi: 10.1111/j.1398-9995.2009.02271.x. [DOI] [PubMed] [Google Scholar]
- 38.Segura E, Touzot M, Bohineust A, et al. Human inflammatory dendritic cells induce Th17 cell differentiation. Immunity. 2013;38:336–348. doi: 10.1016/j.immuni.2012.10.018. [DOI] [PubMed] [Google Scholar]
- 39.Guttman-Yassky E, Lowes MA, Fuentes-Duculan J, et al. Major differences in inflammatory dendritic cells and their products distinguish atopic dermatitis from psoriasis. Journal of Allergy and Clinical Immunology. 2007;119(5):1210–1217. doi: 10.1016/j.jaci.2007.03.006. [DOI] [PubMed] [Google Scholar]
- 40.Robson NC, Phillips DJ, McAlpine T, et al. Activin-A: a novel dendritic cell derived cytokine that potently attenuates CD40 ligand specific cytokine and chemokine production. Blood. 2008;111(5):2733–2743. doi: 10.1182/blood-2007-03-080994. [DOI] [PubMed] [Google Scholar]
- 41.Mikhak Z, Strassner JP, Luster AD. Lung dendritic cells imprint T cell lung homing and promote lung immunity through the chemokine receptor CCR4. Journal of Experimental Medicine. 2013;210:1855–1869. doi: 10.1084/jem.20130091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Reiss Y, Proudfoot AE, Power CA, Campbell JJ, Butcher EC. CC chemokine receptor (CCR)4 and the CCR10 ligand cutaneous T cell-attracting chemokine (CTACK) in lymphocyte trafficking to inflamed skin. Journal of Experimental Medicine. 2001;194(10):1541–1547. doi: 10.1084/jem.194.10.1541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Soler D, Humphreys TL, Spinola SM, Campbell JJ. CCR4 versus CCR10 in human cutaneous TH lymphocyte trafficking. Blood. 2003;101(5):1677–1682. doi: 10.1182/blood-2002-07-2348. [DOI] [PubMed] [Google Scholar]
- 44.McFarland HI, Puig M, Grajkowska LT, et al. Regulatory T cells in gamma irradiation-induced immune suppression. PLoS ONE. 2012;7 doi: 10.1371/journal.pone.0039092.e39092 [DOI] [PMC free article] [PubMed] [Google Scholar]