

Abstract
Curcuma longa L. (Zingiberaceae family) and its polyphenolic compound curcumin have been subjected to a variety of antimicrobial investigations due to extensive traditional uses and low side effects. Antimicrobial activities for curcumin and rhizome extract of C. longa against different bacteria, viruses, fungi, and parasites have been reported. The promising results for antimicrobial activity of curcumin made it a good candidate to enhance the inhibitory effect of existing antimicrobial agents through synergism. Indeed, different investigations have been done to increase the antimicrobial activity of curcumin, including synthesis of different chemical derivatives to increase its water solubility as well ass cell up take of curcumin. This review aims to summarize previous antimicrobial studies of curcumin towards its application in the future studies as a natural antimicrobial agent.
1. Introduction
Curcumin or diferuloylmethane with chemical formula of (1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) (Figure 1) and other curcuminoids constitute the main phytochemicals of Curcuma longa L. (Zingiberaceae family) rhizome with the common name of turmeric [1]. This polyphenolic compound due to a variety of biological activities has been gained significant attention of researches all over the world [2–5]. Turmeric, an ancient coloring spice of Asia, as the main source of curcumin is traditionally used for many remedies [6]. As shown in Figure 2, curcumin due to a variety of specific characterizations is in interest of scientists in recent years. As many other plant materials, there are differences in the curcumin content for the Curcuma longa from different geographical regions and it could be due to hybridization with other Curcuma species which could be important fact to choose the plant with higher content of curcumin [4].
Figure 1.
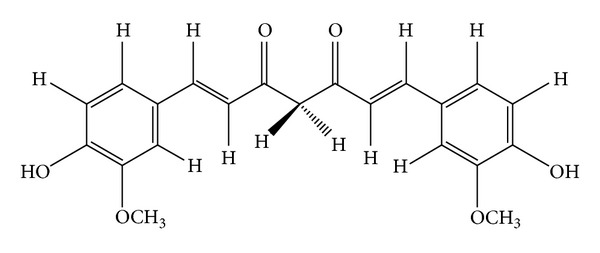
Chemical structure of curcumin.
Figure 2.
Importance of curcumin in antimicrobial studies.
Curcuma longa rhizome has been traditionally used as antimicrobial agent as well as an insect repellant [7]. Several studies have reported the broad-spectrum antimicrobial activity for curcumin including antibacterial, antiviral, antifungal, and antimalarial activities. Because of the extended antimicrobial activity of curcumin and safety property even at high doses (12 g/day) assessed by clinical trials in human, it was used as a structural sample to design the new antimicrobial agents with modified and increased antimicrobial activities through the synthesis of various derivatives related to curcumin [8, 9]. It was even studied as an antimicrobial agent suitable for textile materials. Results showed that curcumin in combination with aloe vera and chitosan could be a potential suppressor for microbial growth in cotton, wool, and rabbit hair assessed by the exhaustion method [10]. Either the continuous or batch dyeing process with curcumin provided textiles with antimicrobial properties beside the color. Curcumin finished wool had semidurable antimicrobial activity, less durable to light exposure than home laundering with 45% and 30% inhibition rates against Staphylococcus aureus and Escherichia coli, respectively, after 30 cycles of home laundering [11]. Mixture of curcumin with other antimicrobial agents is used for the development of antimicrobial skin gels and emulsions with improved skin protection and wound dressing properties [12]. Composition of curcumin with hydrogel silver nanoparticles is used to increase the function of hydrogel silver nanocomposites as marked substances for antimicrobial applications and wound dressing [12]. Curcumin-loaded myristic acid microemulsion with the 0.86 μg/mL of curcumin suitable for skin consumption inhibited 50% of the S. epidermidis growth as one of the nosocomial infectious agents. It showed 12-fold stronger inhibitory effect compared to curcumin activity dissolved in dimethyl sulfoxide (DMSO) [13].
2. Antibacterial Activity
Bacterial infections are among the important infectious diseases. Hence, over 50 years of extensive researches have been launched for achieving new antimicrobial medicines isolated from different sources. Despite progress in development of antibacterial agents, there are still special needs to find new antibacterial agents due to development of multidrug resistant bacteria [14]. The antibacterial study on aqueous extract of C. longa rhizome demonstrated the MIC (minimum inhibitory concentration) value of 4 to 16 g/L and MBC (minimum bactericidal concentration) value of 16 to 32 g/L against S. epidermis ATCC 12228, Staph. aureus ATCC 25923, Klebsiella pneumoniae ATCC 10031, and E. coli ATCC 25922 [15]. The methanol extract of turmeric revealed MIC values of 16 μg/mL and 128 μg/mL against Bacillus subtilis and Staph. aureus, respectively [16]. The study of hexane and ethanol turmeric extract and curcuminoids (from ethyl acetate extract of curcuminoids isolated from C. longa with 86.5% curcumin value) against 24 pathogenic bacteria isolated from the chicken and shrimp showed the highest antimicrobial activity for ethanol extract with the MIC value of 3.91 to 125 ppt [17]. The hexane and methanol extracts of C. longa demonstrated antibacterial effect against 13 bacteria, namely, Vibrio harveyi, V. alginolyticus, V. vulnificus, V. parahaemolyticus, V. cholerae, Bacillus subtilis, B. cereus, Aeromonas hydrophila, Streptococcus agalactiae, Staph. aureus, Staph. intermedius, Staph. epidermidis, and Edwardsiella tarda. However, curcuminoids elicited inhibitory activities against 8 bacteria of Str. agalactiae, Staph. intermedius, Staph. epidermidis, Staph. aureus, A. hydrophila, B. subtilis, B. cereus, and Ed. tarda. Hexane extract and curcuminoids exhibited the MIC values of 125 to 1000 ppt and 3.91 to 500 ppt, respectively [17]. Indeed, it was shown that the addition of 0.3% (w/v) of aqueous curcumin extract to the cheese caused the reduction in bacterial counts of Salmonella typhimurium, Pseudomonas aeruginosa, and E. coli 0157:H7. Moreover, it has decreased the Staph. aureus, B. cereus, and Listeria monocytogenes contamination after 14 days of cold storage period [18]. Turmeric oil as a byproduct from curcumin manufacture also was found effective against B. subtilis, B. coagulans, B. cereus, Staph. aureus, E. coli, and P. aeruginosa [19]. Curcumin also exhibited inhibitory activity on methicillin-resistant Staph. aureus strains (MRSA) with MIC value of 125–250 μg/mL [20]. The in vitro investigation of 3 new compounds of curcumin, namely, indium curcumin, indium diacetyl curcumin, and diacetyl curcumin, against Staph. aureus, S. epidermis, E. coli, and P. aeruginosa revealed that indium curcumin had a better antibacterial effect compared to curcumin itself and it may be a good compound for further in vivo studies. However, diacetylcurcumin did not exhibit any antibacterial effect against tested bacteria [21]. These results demonstrated promising antibacterial activity for different curcumin derivatives as well. The stability and assembly of FtsZ protofilaments as a crucial factor for bacterial cytokinesis are introduced as a possible drug target for antibacterial agents. Curcumin suppressed the B. subtilis cytokinesis through induction of filamentation. It also without significantly affecting the segregation and organization of the nucleoids markedly suppressed the cytokinetic Z-ring formation in B. subtilis [22]. It was demonstrated that curcumin reduces the bundling of FtsZ protofilaments associated with the binding ability to FtsZ with a dissociation constant of 7.3 μM. It showed that curcumin via inhibition of assembly dynamics of FtsZ in the Z-ring can possibly suppress the bacterial cell proliferation as one of the probable antibacterial mechanisms of action [22]. The study on E. coli and B. subtilis demonstrated that curcumin by the inhibitory effect against FtsZ polymerization could suppress the FtsZ assembly leading to disruption of prokaryotic cell division [23].
Also, curcumin showed significant antibacterial activity with MIC values between 5 and 50 μg/mL against 65 clinical isolates of Helicobacter pylori [41]. Curcumin also has an inhibitory effect on NF-κB activation and as a result on the release of IL-8 and cell scattering which led to a reduction in inflammation of gastric tissue as the main consequence for H. pylori in stomach. It inhibits the IκBα degradation, the activity of NF-κB DNA-binding and IκB kinase α and β (IKK α and β) [42]. Indeed, curcumin inhibited the matrix metalloproteinase-3 and metalloproteinase-9 activity (MMP-3 and MMP-9) as inflammatory molecules involved in H. pylori infection in mice and in cell culture with a dose dependent manner [43]. Curcumin showed more efficient therapeutic index than conventional triple therapy of H. pylori on MMP-3 and MMP-9 via reduction of activator protein-1 and proinflammatory molecule activation in H. pylori infected gastric tissues [43]. In vivo study of antibacterial effect of curcumin on H. pylori compared to OAM (Omeprazole, Amoxicillin, and Metronidazole) treatment revealed poor activity for eradication of H. pylori (5.9% versus 78.9% for OAM treatment). The reduction in inflammatory cytokine production was not reported from pylori-infected patients treated with curcumin [44]. The in vivo study of 1-week nonantibiotic therapy comprised of curcumin, pantoprazole, N-acetylcysteine, and lactoferrin against H. pylori infection was not effective for the eradication of H. pylori. However, the decrease in immunological criteria of gastric inflammation and dyspeptic symptoms was reported after 2 months of treatment schedule [45]. Nevertheless, the curcumin administration to the rats with H. pylori-induced gastric inflammation revealed a significant reduction in macromolecular leakage and NF-κB activation [46]. In an in vivo study of H. pylori-infected C57BL/6 mice administered with curcumin exhibited immense therapeutic potential and pronounced eradication effect against H. pylori infection associated with restoration of gastric damage [41].
2.1. Synergistic Antimicrobial Activity
The outburst of drug resistant microbial strains necessitates the studies for synergistic effects of antibiotics in combination with plant's derivatives to develop the antimicrobial cocktail with a wider spectrum of activity and reduction of adverse side effects of antimicrobial agents. Staph. aureus resistance to the penicillin group of antibiotics is increasing associated with appearance of adverse side effects such as hypersensitivity and anaphylactic reactions [47]. The synergistic activity of curcuminoids and ampicillin combination demonstrated pronounced reduction in the MIC of ampicillin against either clinical strain or Staph. aureus ATCC 25923 strain. Bacteriocin subtilosin isolated from B. amyloliquefaciens in combination with encapsulated curcumin revealed partial synergism against wild-type and nisin sensitive strains of L. monocytogenes Scott A [48]. In another in vivo study using 500 μg/disc of curcumin against clinical isolate of Staph. aureus the synergistic activity with antibiotics of cefixime, cefotaxime, vancomycin, and tetracycline was demonstrated [49]. The results proved that consumption of turmeric during the treatment of Staph. aureus infections with these antibiotics especially cefixime can be possibly helpful. Curcumin also demonstrated a synergistic effect in combination with some antibiotics, including ampicillin, oxacillin, and norfloxacin against methicillin-resistant Staph. aureus strain (MRSA) [20]. The synergistic effect of curcumin with ciprofloxacin against MRSA has also been reported, although there is an evidence of its antagonistic activity against S. typhi and S. typhimurium in combination with ciprofloxacin [49, 50].
Strongly bound metal complexes to antimicrobial agents are introduced as another possible way for synergistic activity of respective antimicrobial agents through elevation of the binding effect of them to the bacterial walls. Complexes of curcumin with cobalt nanoparticles showed increased antibacterial activity against E. coli [51]. Additionally, fabrication of silver nanocomposite films impregnated with curcumin showed the stronger antibacterial activity against E. coli. It was shown that the bactericidal activity of sodium carboxymethyl cellulose silver nanocomposite films (SCMC SNCFs) as an effective antibacterial material was improved by loading of curcumin with SCMC SNCFs [52]. In another in situ investigation, the synergistic effect of curcumin encapsulated chitosan-[poly (vinyl alcohol)] silver nanocomposite films was shown. The novel antimicrobial films with pronounced antimicrobial exhibition against E. coli proved to be potential antibacterial material for treating infections or wound dressing [53].
2.2. Anti-Biofilm Activity
Secretion of exopolysaccharide alginate via different stimulators such as aminoglycosides and imipenem consumption caused the increase in biofilm volume of P. aeruginosa. Anti-biofilm activity of curcumin against two strains of P. aeruginosa isolated from deep oropharyngeal swap samples of two cystic fibrosis patients with MIC values of 16 μg/mL was investigated by crystal violet staining method. The curcumin treatment of the strains with MIC concentrations did not reveal noteworthy elevation in biofilm optical density [54]. In addition, in another study curcumin showed the potential for reduction of biofilm initiation genes, inhibition of 31 quorum sensing (QS) genes, and downregulation of virulence factors including acyl homoserine lactone (HSL) production, elastase/protease activity, and pyocyanin biosynthesis. The antimicrobial activities led to reduction of pathogenicity in Arabidopsis thaliana and Caenorhabditis elegans as whole plant and animal infected models with P. aeruginosa [7]. The results exhibited that curcumin can be a potential candidate for P. aeruginosa infections in special infections characterized by biofilm formation, although further comprehensive studies are needed for the approval.
In some cases the adverse effects of curcumin against different antibiotics were shown. Ciprofloxacin is the most effective antibiotic against typhoidal and nontyphoidal infection of Salmonella. The main mechanism for antibacterial activity of ciprofloxacin is through SOS response, induction of chromosome fragmentation, and the production of ROS in the bacterial cell. The in vivo and in vitro investigations on curcumin together with ciprofloxacin showed that, through interference with ciprofloxacin activity, it caused an elevation in proliferation of Salmonella typhi and Salmonella enterica serovar Typhimurium (S. typhimurium). Although curcumin could not suppress the ciprofloxacin-induced gyrase inhibition, it protected Salmonella against oxidative burst induced by interferon γ (IFNγ) or ciprofloxacin via owing strong antioxidant effect. The results demonstrated the curcumin by suppressing the antibacterial effect of IFNγ or ciprofloxacin might increase the Salmonella pathogenesis [55]. The study of curcumin activity in a murine model of typhoid fever exhibited an elevation of Salmonella typhimurium pathogenicity and increased resistance to antimicrobial agents including antimicrobial peptides, nitrogen species, and reactive oxygen. Upregulation of genes involved in antioxidative function like mntH, sitA, and sodA as well as other genes involved in resistance to antimicrobial peptides including pmrD and pmrHFIJKLM was considered as a possible cause for the mentioned elevated tolerance. Curcumin also induced upregulation effect on SPI2 genes involved in intracellular survival and downregulation activity on SPI1 genes involved for entry within epithelial cells. This information proved that the indiscriminate use of curcumin should probably inhibit the pathogenesis of Salmonella [55]. Additionally, curcumin also at a dose 500 μg/disc showed antagonistic activity on the bactericidal effect of nalidixic acid against clinical strain of Staph. aureus investigated by disc diffusion method [49].
3. Antiviral Activity
Lack of effective therapeutics for the most of viral diseases, emergence of antiviral drug resistance, and high cost of some antiviral therapies necessitate finding new effective antiviral compounds [56, 57]. Additionally, the existing antiviral therapies are not always well-tolerated or quite effective and satisfactory [58]. Hence, the increasing requirement for antiviral substances will be more highlighted. Plants as a rich source of phytochemicals with different biological activities including antiviral activities are in interest of scientists [59, 60]. It has been demonstrated that curcumin as a plant derivative has a wide range of antiviral activity against different viruses. Inosine monophosphate dehydrogenase (IMPDH) enzyme due to rate-limiting activity in the de novo synthesis of guanine nucleotides is suggested as a therapeutic target for antiviral and anticancer compounds. Among the 15 different polyphenols, curcumin through inhibitory activity against IMPDH effect in either noncompetitive or competitive manner is suggested as a potent antiviral compound via this process [61]. The study of different bioconjugates of curcumin, namely, di-O-tryptophanylphenylalanine curcumin, di-O-decanoyl curcumin, di-O-pamitoyl curcumin, di-O-bis-(γ,γ)folyl curcumin, C4-ethyl-O-γ-folyl curcumin, and 4-O-ethyl-O-γ-folyl curcumin, against variety of viruses including parainfluenza virus type 3 (PIV-3), feline infectious peritonitis virus (FIPV), vesicular stomatitis virus (VSV), herpes simplex virus (HSV), flock house virus (FHV), and respiratory syncytial virus (RSV) assessed by MTT test showed the potent antiviral activity of curcumin and its bioconjugates against different viral pathogens for further studies. Also, di-O tryptophanylphenylalanine curcumin and di-O-decanoyl curcumin revealed remarkable antiviral activity against VSV and FIPV/FHV with EC50 values of 0.011 μM and 0.029 μM, respectively. However, bioconjugates did not exhibit significant antiviral activity against IIIB and ROD strains of type 1 human immunodeficiency virus (HIV-1) in MT-4 cells [62]. Table 1 summarizes the antiviral activity of C. longa and curcumin and possible mechanisms underlying inhibitory effects.
Table 1.
Antiviral activities of Curcuma longa L. and curcumin.
| Virus | Antiviral substances | Description of antiviral activity type | Reference |
|---|---|---|---|
| HIV | Curcumin | Inhibition of HIV-1 LTR-directed gene expression | [24] |
| Curcumin, reduced curcumin, allyl-curcumin, tocopheryl-curcumin | Inhibition of Tat-mediated transactivation of HIV-1 LTR | [25] | |
| Curcumin, curcumin boron complexes | Inhibition of HIV-1 and HIV-2 proteases | [26] | |
| Curcumin | Inhibition of HIV-1 Integrase | [27] | |
| Curcumin | Inhibition of Tat protein acetylation | [28] | |
| Curcumin | No antiviral effect in clinical trial | [29] | |
| Influenza | Curcumin | Inhibition of haemagglutination | [30] |
| HSV-1 | Curcumin, gallium-curcumin, Cu-curcumin | Reduction of HSV-1 replication | [31, 32] |
| HSV-2 | Curcumin | Significant protection in mouse model | [33] |
| Coxsackievirus | Curcumin | Replication inhibition through UPS dysregulation | [34] |
| HBV | Aqueous extract | Suppression of HBV replication by increasing the p53 level | [35] |
| HCV | Curcumin | Decrease of HCV replication by suppressing the Akt-SREBP-1 pathway | [36] |
| HPV | Curcumin | Inhibition expression of viral oncoproteins of E6 and E7 | [37] |
| HPV | Curcumin | Downregulation effect on the transcription of HPV-18 | [38] |
| JEV | Curcumin | Reduction in production of infective viral particles | [39] |
| HTLV-1 | Curcumin | Downregulation of JunD protein in HTLV-1-infected T-cell lines | [40] |
Viral long terminal repeat (LTR) has a critical role in transcription of type 1 human immunodeficiency virus (HIV-1) provirus. Inhibition of LTR activity can be a possible pathway for antiviral drug candidates in order to block HIV-1 replication [63, 64]. Curcumin proved to be an effective compound to inhibit the HIV-1 LTR-directed gene expression without any major effects on cell viability [24]. Curcumin and its derivatives, namely, reduced curcumin, allyl-curcumin, and tocopheryl-curcumin, revealed 70% to 85% inhibition in Tat protein transactivation of HIV-1 LTR measured by β-galactosidase activities of HeLa cells which in HIV-1 LTR was fused to the indictor of lacZ gene. Tocopheryl-curcumin demonstrated the most inhibition activity with 70% inhibition at 1 nM compared to 35% inhibition of curcumin at this concentration [25]. In addition, curcumin inhibited the acetylation of Tat protein of HIV significantly by p300 associated with suppression of HIV-1 multiplication. Curcumin by targeting the acetyltransferase proteins of p300/CREB-binding protein (CBP) can be a potent compound for combinatorial HIV therapeutics [28]. Curcumin was found to be an inhibitor of HIV-1 and HIV-2 protease with IC50 of 100 μM and 250 μM, respectively. The curcumin boron complexes exhibited noteworthy inhibition reduced to the IC50 value of 6 μM with time-dependent activity. The elevated affinity of boron derivatives of curcumin is possibly associated with the attachment of the orthogonal domains of the compound in intersecting sites within the substrate-binding cavity of the protease [26]. Integrase as another essential enzyme for HIV-1 replication was found to be inhibited by curcumin with IC50 value of 40 μM. Inhibition of deletion mutant of integrase containing only amino acids 50–212 indicated that curcumin possibly interacts with catalytic core of the enzyme. The study of energy minimization and the structural analogs of curcumin elicited that an intramolecular stacking of two phenyl rings of curcumin is possibly responsible for anti-integrase activity via bringing the hydroxyl groups into close proximity [27]. However, rosmarinic acid and dicaffeoyl methane as two curcumin analogs showed noteworthy inhibitory activity against integrase of HIV-1 with IC50 values less than 10 μM with the slow rate of binding to the enzyme assessed by kinetic studies [65]. However, through a clinical trial investigation on curcumin as an anti-HIV compound in 40 patients in eight weeks it was shown that there is no reduction in viral load or elevation in CD4 counts. But patients claimed that they preferred to take the curcumin in order to tolerate the minor gastrointestinal sufferings and feel better [29]. This demonstrated that clinical trials can possibly show up with the results completely different from in vitro studies. The clinical trial of clear liquid soap containing 0.5% w/v ethanol extract of C. longa rhizome on HIV patients reduced the wound infections and 100% decrease in itching symptom and it also affected the abscess to convert to dryness scabs (78.6%) within 2 weeks [16].
Curcumin showed the anti-influenza activity against influenza viruses PR8, H1N1, and H6N1. The results showed more than 90% reduction in virus yield in cell culture using 30 μM of curcumin. The plaque reduction test elicited the approximate EC50 of 0.47 μM for curcumin against influenza viruses [30]. In H1N1 and also H6N1 subtypes, the inhibition of haemagglutinin interaction reflected the direct effect of curcumin on infectivity of viral particles and this has proved by time of drug addiction experiment. Additionally, unlike amantadine, viruses developed no resistance to curcumin. The methoxyl derivatives of curcumin also did not show noteworthy role in the haemagglutination [30]. These results proved the significant potential of curcumin for inhibition of influenza.
In vitro study of curcumin and its derivatives, namely, gallium-curcumin and Cu-curcumin, exhibited remarkable antiviral activity against herpes simplex virus type 1 (HSV-1) in cell culture with IC50 values of 33.0 microg/mL, 13.9 microg/mL, and 23.1 microg/mL, respectively. The 50% cytotoxic concentration (CC50) of the respective compounds on Vero cell line showed to be 484.2 μg/mL, 255.8 μg/mL, and 326.6 μg/mL, respectively [31]. Curcumin considerably decreased the immediate early (IE) gene expression and infectivity of HSV-1 in cell culture assays. Curcumin has an effect on recruitment of RNA polymerase II to IE gene promoters through mediation of viral transactivator protein VP16, by an independent process of p300/CBP histone acetyl transferase effect [32]. In vitro replication of HSV-2 could be decreased by curcumin with ED50 value of 0.32 mg/mL [32]. Moreover, an in vivo study on mouse model with intravaginal HSV-2 challenge showed significant protection against HSV-2 infection due to administration of curcumin. This study showed that curcumin can be a good candidate for developing the antiviral products used intravaginally by women for protection against sexually transmitted herpes virus infection [33]. Indeed, a metallo-herbal complex of curcumin with copper (Cu2+) demonstrated microbicidal effect for further studies of vaginal gel with antiviral activity [66].
Coxsackieviruses cause a variety of diseases such as dilated cardiomyopathy and myocarditis. Coxsackievirus B3 (CVB3) in spite of extensive investigations is still a major human pathogen without specific effective and approved treatment [67, 68]. Curcumin exhibited the antiviral activity against coxsackievirus by reduction of viral RNA expression, protein synthesis, and virus titer. In addition, it was found to have a protective effect on cells against virus-induced apoptosis and cytopathic activity. Analysis of different pathways showed that curcumin forced its potent antiviral effect in inhibition of coxsackievirus replication through dysregulation of the ubiquitin-proteasome system (UPS) [34]. The recent studies proved that the UPS-mediated protein modification or degradation is an essential factor in the regulation of coxsackievirus replication [69].
Liver diseases associated with viral infections are major pandemics [70]. The fact that hepatitis B virus (HBV) elevates the possibility for the hepatocellular carcinoma (HCC) development some 100-fold and 695.900 deaths occurred due to liver cirrhosis and HCC worldwide in 2008 makes the need to find new antivirals against hepatitis viruses [71, 72]. The study of antiviral effect of aqueous extract of Curcuma longa rhizoma against HBV in HepG 2.2.15 cells containing HBV genomes showed repression of HBsAg secretion from liver cells without any cytotoxic effect. It also suppressed the HBV particles production and the rate of mRNA production of HBV on infected cells. The Curcuma longa extract suppressed HBV replication by increasing the rate of p53 protein through enhancing the stability of the protein as well as transactivating the transcription of p53 gene. It was understood that the extract has suppressed HBV enhancer I and X promoter leading to repression of HBx gene transcription by affecting p53 [35]. In vitro investigation of the antiviral activity of curcumin Huh7 replicon cells expressing the hepatitis C virus (HCV) indicated that curcumin can be a potent anti-HCV compound. Results showed the decrease in HCV gene expression and replication through suppressing the Akt-SREBP-1 pathway. In addition, the mixture of curcumin and IFNα as the known anti-HCV therapy induced profound inhibitory activity on HCV replication and demonstrated that curcumin can be possibly used as a complementary therapy for HCV [36].
High-risk human papillomaviruses (HPVs) infection via the expression of E6 and E7 viral oncoproteins has a critical role for development of cervical carcinoma. Curcumin showed the inhibitory activity against the expression of E6 and E7 genes of HPV-16 and HPV-18 as two main highly oncogenic human papilloma viruses [37]. The transcription factor AP-1 is a critical factor for transcriptional regulation of high-risk HPVs such as HPV-16 and HPV-18. Curcumin downregulates the AP-1 binding activity in HeLa cells with decreasing effect on the transcription of HPV-18 [38]. The results showed that curcumin through apoptosis modulation and also prevention of NFκB and AP-1 translocation associated with downregulation of viral oncogenes and decreasing the transcription of HPVs can be a good candidate for the management of highly oncogenic HPV infections [37, 38].
Japanese encephalitis virus (JEV) as an important endemic arbovirus in Southeast Asia is a major cause of acute encephalopathy which generally affects the children and leads to death in one third of patients. The permanent neuropsychiatric sequel is a complication for many survivors from JEV due to ineffective therapeutic measure [73]. The investigation of antiviral activity of curcumin on Neuro2a cell line infected with JEV showed reduction in production of infectious viral particles through inhibition of ubiquitin-proteasome system. The results of in vitro study indicated that curcumin through modulating cellular levels of stress-related proteins, reducing proapoptotic signaling molecules, restoration of cellular membrane integrity, and reduction in reactive oxygen species in cellular level imparts neuroprotection and can be a potential for further investigations [39].
Oncogenesis by human T-cell leukemia virus type 1 as an etiologic factor of adult T-cell leukemia (ATL) is critically dependent on the activation of the activator protein 1 (AP-1) [74]. The DNA binding and transcriptional effect of AP-1 in HTLV-1-infected T-cell lines were suppressed by curcumin treatment. Curcumin also inhibited the expression of JunD protein as an important factor in AP-1-DNA complex in HTLV-1-infected T-cells as well as HTLV-1 Tax-induced AP-1 transcriptional effect. Cell cycle arrest and inducing of apoptosis were found to be possible mechanisms against HTLV-1 replication in infected T-cell line by curcumin. Suppression of AP-1 activity possibly through decreasing the expression of JunD protein is introduced as a possible pathway for anti-ATL activity of curcumin [40].
4. Antifungal Activity
Substances and extracts isolated from different natural resources especially plants have always been a rich arsenal for controlling the fungal infections and spoilage. Due to extensive traditional use of turmeric in food products, various researches have been done in order to study the turmeric and curcumin with the aspect of controlling fungal related spoilage and fungal pathogens. The study of addition the turmeric powder in plant tissue culture showed that turmeric at the 0.8 and 1.0 g/L had appreciable inhibitory activity against fungal contaminations [75]. The methanol extract of turmeric demonstrated antifungal activity against Cryptococcus neoformans and Candida albicans with MIC values of 128 and 256 μg/mL, respectively [16]. The study of hexane extract of C. longa at 1000 mg/L demonstrated antifungal effect against Rhizoctonia solani, Phytophthora infestans, and Erysiphe graminis. It was also shown that 1000 mg/L of ethyl acetate extract of C. longa exhibitedinhibitory effect against R. solani, P. infestans, Puccinia recondita, and Botrytis cinerea. Curcumin at 500 mg/L also showed antifungal activity against R. solani, Pu. recondita, and P. infestans [76]. Curcumin and turmeric oil exert antifungal effect against two phytophagous fungi, namely, Fusarium solani and Helminthosporium oryzae. Turmeric oil exhibited the most effective antifungal activity against F. solani and H. oryzae with IC50 of 19.73 and 12.7 μg/mL, respectively [77]. The crude methanol extract of C. longa has inhibitory effect against some clinical isolates of dermatophytes. It was demonstrated that 18-month-old and freshly distilled oil isolated from rhizome of C. longa showed the most potent antifungal effect against 29 clinical isolates of dermatophytes with MIC values of 7.2 and 7.8 mg/mL, respectively [78]. Trichophyton rubrum, T. mentagrophytes, Epidermophyton floccosum, and Microsporum gypseum were suppressed by 1 : 40–1 : 320 dilutions of turmeric oil. An in vivo study on infected guinea pigs with T. rubrum demonstrated that dermal application of turmeric oil (dilution 1 : 80) induced an improvement in healing of the lesions after 2–5 days and it caused the lesions after 6-7 days of consumption to vanish. Turmeric oil also showed activity against pathogenic molds such as Sporothrix schenckii, Exophiala jeanselmei, Fonsecaea pedrosoi, and Scedosporium apiospermum with MIC values of 114.9, 459.6, 459.6, and 114.9 μg/mL, respectively [79]. However, curcumin showed more significant effect against Paracoccidioides brasiliensis than fluconazole, although it did not affect the growth of Aspergillus species [80]. The possible mechanism underlying the mentioned antifungal effect was found to be downregulation of Δ5,6 desaturase (ERG3) leading to significant reduction in ergosterol of fungal cell. Reduction in production of ergosterol results in accumulations of biosynthetic precursors of ergosterol which leads to cell death via generation of ROS [81]. Reduction in proteinase secretion and alteration of membrane-associated properties of ATPase activity are other possible critical factors for antifungal activity of curcumin [82].
Resistant strain development among the Candida species against existing antifungal drugs became a critical problem for therapeutic strategies. Thereby, finding new anti-Candida substances seems to be crucial [83]. The study of curcumin against 14 strains of Candida including 4 ATCC strains and 10 clinical isolates showed that curcumin is a potent fungicide compound against Candida species with MIC values range from 250 to 2000 μg/mL [82]. In another study, anti-Candida activity of curcumin was demonstrated against 38 different strains of Candida including some fluconazole resistant strains and clinical isolates of C. albicans, C. glabrata, C. krusei, C. tropicalis, and C. guilliermondii. The MIC90 values for sensitive and resistant strains were 250–650 and 250–500 μg/mL, respectively. Intracellular acidification via inhibition of H+-extrusion was identified as possible mechanism for cell death of Candida species [84]. The development of hyphae was proved to be inhibited by curcumin through targeting the global suppressor thymidine uptake 1 (TUP1) [81, 85]. Curcumin also showed inhibitory effect on Cryptococcus neoformans and C. dubliniensis with MIC value of 32 mg/L [80]. One of the major complications during therapies against chronic asthma is oropharyngeal candidiasis. Curcumin as a potential candidate for the treatment of candidosis with anti-inflammatory activity was studied in a murine model of asthma. Oral administrator of Curcumin is more effective than dexamethasone in reducing fungal burden in BALB/c mice. It also significantly decreased pathological changes in asthma [86]. Adhesion of Candida species isolated from AIDS patients to buccal epithelial cells is also markedly inhibited by curcumin and it was found to be more effective compared to fluconazole [80].
The investigation of curcumin mediation for photodynamic therapy can reduce the biofilm biomass of C. albicans, C. glabrata, and C. tropicalis. The results demonstrated that association of four LED fluences for light excitation with 40 μM concentration of curcumin at 18 J/cm2 inhibited up to 85% metabolic activity of the tested Candida species. The use of curcumin with light proved to be an effective method for noteworthy improvement in the antifungal activity against planktonic form of the yeasts [87]. Photodynamic effect considerably decreased C. albicans viability in either planktonic or biofilm cultures probably through increasing the uptake of curcumin by cells. However, to a lesser extent, photodynamic therapy was found to be phototoxic to the macrophages. [88]. A study on a murine model of oral candidiasis was done for gathering reliable data for curcumin-mediated photodynamic therapy efficacy in vivo. Results proved that all exposures to curcumin with LED light markedly inhibited the C. albicans viability after photodynamic therapy without harming the host tissue of mice. However, 80 μM of curcumin in association with light showed the best decrease in colony counts of C. albicans [89]. These results showed that curcumin is a high potential photosensitizer compound for fungicidal photodynamic therapy especially against Candida species.
The strong antifungal activity of C. longa rhizome and its low side effect were the main reasons to investigate its probable synergistic effect with existing fungicides. The synergistic activity of curcumin with five azole and two polyene drugs including voriconazole, itraconazole, ketoconazole, miconazole, fluconazole, amphotericin B, and nystatin showed 10–35-fold reduction in the MIC values of the fungicides against 21 clinical isolates of C. albicans. The synergistic activity of curcumin with amphotericin B and fluconazole could be associated with the accumulation of ROS which will be suppressed by adding an antioxidant [85]. The study of 200 clinical isolates of Candida species including C. tropicalis, C. kefyr, C. krusei, C. guilliermondii, C. glabrata, C. parapsilosis, and C. albicans demonstrated fungicidal activity for curcumin with MIC value of 32–128 μg/mL. Combination of curcumin with amphotericin B also exhibited synergistic activity against tested Candida species, although fluconazole and curcumin in some cases showed additive effects rather than synergistic activity. These results proved that combination of curcumin with existing fungicidal agents can provide more significant effect against systemic fungal infections like candidemia and candidosis [90]. In silico analysis demonstrated that curcumin by attaching to albumin serum in a separate binding site of amphotericin B and forming the complex alleviated the adverse side effect of amphotericin B via delaying the red cell lysis. The stability and aqueous solubility of the complex of curcumin and amphotericin B with albumin serum can be a potential candidate for the treatment of visceral leishmaniasis and systemic fungal infections [91]. The in vivo study of combination of curcumin and piperine in murine model of Candida infection also revealed synergistic effect with noteworthy fungal load reduction in kidney of Swiss mice [85]. The mixture of curcumin and ascorbic acid against different strains of Candida also exhibited 5- to 10-fold reduction of MIC values compared to the time that curcumin was tested alone [92]. These synergistic effects showed that curcumin in combination with different fungicide materials can significantly elicit synergistic activity to enhance the efficacy of existing antifungal strategies.
5. Enhancing the Bioavailability and Solubility of Curcumin to Improve Antimicrobial Activities
The optimum potential of curcumin is limited because of poor oral bioavailability and insufficient solubility in aqueous solvents leading to poor absorption, fast metabolism, and quick systemic elimination [5, 93]. For overcoming this obstacle, nanocarriers like curcumin-loaded PLGA (poly lactide-co-glycolide) and curcumin nanoparticle formulation were investigated and their better bioactivity and bioavailability as well as increased cellular uptake compared to curcumin were reported [5]. Another study revealed that heat-extracted curcumin elevated the solubility of curcumin 12-fold without significant disintegration due to heat treatment. Modification of 4-hydroxy-2-nonenal (HNE) as a critical oxidation by-product involved in disease pathogenesis via cytotoxicity, genotoxicity, and mutagenicity is inhibited 80% by heat-solubilized curcumin and suggested a possible mechanism for inducing bioactivity of curcumin [94]. The study of nanocurcumin as a nanoparticle of curcumin with the size of 2–40 nm processed by a wet-milling technique, showed curcumin to be more freely dispersible in water leading to more significant antimicrobial activity against Staph. aureus, E. coli, P. aeruginosa, B. subtilis, and two fungi of P. notatum and A. niger due to reduced particle size and enhanced bioavailability [95, 96]. However, nanocurcumin demonstrated more noteworthy activity against Gram-positive bacteria rather than Gram-negatives [95]. In another study to improve the stability and solubility of curcumin, microencapsulation process was investigated. Microcapsule of curcumin with improved solubility is suitable as a preservative and colorant in food industry and it exhibited potent antimicrobial effect against food-borne pathogens including E. coli, Staph. aureus, B. subtilis, B. cereus, Yersinia enterocolitica, Penicillium notatum, and Saccharomyces cerevisiae with MIC values ranging from 15.7 to 250 μg/mL. It was demonstrated that Gram-positive bacteria were more susceptible to the microcapsulated curcumin compared to Gram-negatives. However, antifungal effect was found to be stronger than the bactericidal effect [97, 98].
6. Conclusion
All previous investigations have shown the extensive antimicrobial activity of curcumin, although in vivo studies in some cases reported the less effective results of curcumin inhibitory effect. Among all former studies on antibacterial activity of curcumin the most promising result is against Helicobacter pylori, at least for using the curcumin as a complementary compound in combination with other existing medicines to decrease the symptoms of gastritis. The extensive antiviral effects of curcumin against different viral pathogens nominate this compound as an antiviral drug candidate to develop new antivirals from natural resources against sensitive viruses especially by developing different curcumin derivatives. However, using curcumin or its derivatives as antiviral compounds needs further investigations. Regarding the studies on antifungal activities of curcumin the most significant effect was found against Candida species and Paracoccidioides brasiliensis, although curcumin revealed fungicide effect against various fungi. In spite of various biological activities of curcumin, no real clinical uses have been reported for this compound and still clinical trials are undergoing for different ailments and diseases, namely, colon and pancreatic cancers, multiple myeloma, myelodysplastic syndromes, Alzheimer, and psoriasis [99]. Until 2013, more than 65 clinical trials on curcumin have been carried out, and still more is underway. This polyphenol compound is now used as a supplement in several countries, namely, China, India, Japan, Korea, South Africa, the United States, Thailand, and Turkey [100].
Acknowledgment
The authors would like to thank University of Malaya for University Malaya Research Flagship Grant (UMRG) (FL001-13HTM) and Postgraduate Research Grant (PPP) (PG005-2012B).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Ammon HPT, Wahl MA. Pharmacology of Curcuma longa . Planta Medica. 1991;57(1):1–7. doi: 10.1055/s-2006-960004. [DOI] [PubMed] [Google Scholar]
- 2.Lai PK, Roy J. Antimicrobial and chemopreventive properties of herbs and spices. Current Medicinal Chemistry. 2004;11(11):1451–1460. doi: 10.2174/0929867043365107. [DOI] [PubMed] [Google Scholar]
- 3.Maheshwari RK, Singh AK, Gaddipati J, Srimal RC. Multiple biological activities of curcumin: a short review. Life Sciences. 2006;78(18):2081–2087. doi: 10.1016/j.lfs.2005.12.007. [DOI] [PubMed] [Google Scholar]
- 4.Hayakawa H, Minanyia Y, Ito K, Yamamoto Y, Fukuda T. Difference of curcumin content in Curcuma longa L., (Zingiberaceae) caused by Hybridization with other Curcuma species. American Journal of Plant Sciences. 2011;2(2):111–119. [Google Scholar]
- 5.Anand P, Nair HB, Sung B, et al. Design of curcumin-loaded PLGA nanoparticles formulation with enhanced cellular uptake, and increased bioactivity in vitro and superior bioavailability in vivo . Biochemical Pharmacology. 2010;79(3):330–338. doi: 10.1016/j.bcp.2009.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 6.Araújo CAC, Leon LL. Biological activities of Curcuma longa L. Memorias do Instituto Oswaldo Cruz. 2001;96(5):723–728. doi: 10.1590/s0074-02762001000500026. [DOI] [PubMed] [Google Scholar]
- 7.Rudrappa T, Bais HP. Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. Journal of Agricultural and Food Chemistry. 2008;56(6):1955–1962. doi: 10.1021/jf072591j. [DOI] [PubMed] [Google Scholar]
- 8.LaColla P, Tramontano E, Musiu C, Marongiu ME, Novellino E, Greco G. Curcumin-like derivatives with potent activity against HIV-1 integrase: synthesis, biological evaluation and molecular modeling. Antiviral Research. 1998;37(3):57–57. [Google Scholar]
- 9.Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB. Bioavailability of curcumin: problems and promises. Molecular Pharmaceutics. 2007;4(6):807–818. doi: 10.1021/mp700113r. [DOI] [PubMed] [Google Scholar]
- 10.Ammayappan L, Jeyakodi Moses J. Study of antimicrobial activity of aloevera, chitosan, and curcumin on cotton, wool, and rabbit hair. Fibers and Polymers. 2009;10(2):161–166. [Google Scholar]
- 11.Han S, Yang Y. Antimicrobial activity of wool fabric treated with curcumin. Dyes and Pigments. 2005;64(2):157–161. [Google Scholar]
- 12.Varaprasad K, Vimala K, Ravindra S, Narayana Reddy N, Venkata Subba Reddy G, Mohana Raju K. Fabrication of silver nanocomposite films impregnated with curcumin for superior antibacterial applications. Journal of Materials Science: Materials in Medicine. 2011;22(8):1863–1872. doi: 10.1007/s10856-011-4369-5. [DOI] [PubMed] [Google Scholar]
- 13.Liu CH, Huang HY. Antimicrobial activity of curcumin-loaded myristic acid microemulsions against Staphylococcus epidermidis . Chemical and Pharmaceutical Bulletin. 2012;60(9):1118–1124. doi: 10.1248/cpb.c12-00220. [DOI] [PubMed] [Google Scholar]
- 14.Wise R, Hart T, Cars O, et al. Antimicrobial resistance. Is a major threat to public health. British Medical Journal. 1998;317(7159):609–610. doi: 10.1136/bmj.317.7159.609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Niamsa N, Sittiwet C. Antimicrobial activity of Curcuma longa aqueous extract. Journal of Pharmacology and Toxicology. 2009;4(4):173–177. [Google Scholar]
- 16.Ungphaiboon S, Supavita T, Singchangchai P, Sungkarak S, Rattanasuwan P, Itharat A. Study on antioxidant and antimicrobial activities of turmeric clear liquid soap for wound treatment of HIV patients. Songklanakarin Journal of Science and Technology. 2005;27(2):269–578. [Google Scholar]
- 17.Lawhavinit O-A, Kongkathip N, Kongkathip B. Antimicrobial activity of curcuminoids from Curcuma longa L. on pathogenic bacteria of shrimp and chicken. Kasetsart Journal—Natural Science. 2010;44(3):364–371. [Google Scholar]
- 18.Hosny IM, El Kholy WI, Murad HA, El Dairouty RK. Antimicrobial activity of Curcumin upon pathogenic microorganisms during manufacture and storage of a novel style cheese ‘Karishcum’. Journal of American Science. 2011;7:611–618. [Google Scholar]
- 19.Negi PS, Jayaprakasha GK, Rao LJM, Sakariah KK. Antibacterial activity of turmeric oil: a byproduct from curcumin manufacture. Journal of Agricultural and Food Chemistry. 1999;47(10):4297–4300. doi: 10.1021/jf990308d. [DOI] [PubMed] [Google Scholar]
- 20.Mun SH, Joung DK, Kim YS, et al. Synergistic antibacterial effect of curcumin against methicillin-resistant Staphylococcus aureus . Phytotherapy Research. 2013;19(7):599–604. [Google Scholar]
- 21.Tajbakhsh S, Mohammadi K, Deilami I, et al. Antibacterial activity of indium curcumin and indium diacetylcurcumin. African Journal of Biotechnology. 2008;7(21):3832–3835. [Google Scholar]
- 22.Rai D, Singh JK, Roy N, Panda D. Curcumin inhibits FtsZ assembly: an attractive mechanism for its antibacterial activity. Biochemical Journal. 2008;410(1):147–155. doi: 10.1042/BJ20070891. [DOI] [PubMed] [Google Scholar]
- 23.Kaur S, Modi NH, Panda D, Roy N. Probing the binding site of curcumin in Escherichia coli and Bacillus subtilis FtsZ—a structural insight to unveil antibacterial activity of curcumin. European Journal of Medicinal Chemistry. 2010;45(9):4209–4214. doi: 10.1016/j.ejmech.2010.06.015. [DOI] [PubMed] [Google Scholar]
- 24.Li CJ, Zhang LJ, Dezube BJ, Crumpacker CS, Pardee AB. Three inhibitors of type 1 human immunodeficiency virus long terminal repeat-directed gene expression and virus replication. Proceedings of the National Academy of Sciences of the United States of America. 1993;90(5):1839–1842. doi: 10.1073/pnas.90.5.1839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Barthelemy S, Vergnes L, Moynier M, Guyot D, Labidalle S, Bahraoui E. Curcumin and curcumin derivatives inhibit Tat-mediated transactivation of type 1 human immunodeficiency virus long terminal repeat. Research in Virology. 1998;149(1):43–52. doi: 10.1016/s0923-2516(97)86899-9. [DOI] [PubMed] [Google Scholar]
- 26.Sui Z, Salto R, Li J, Craik C, Ortiz de Montellano PR. Inhibition of the HIV-1 and HIV-2 proteases by curcumin and curcumin boron complexes. Bioorganic & Medicinal Chemistry. 1993;1(6):415–422. doi: 10.1016/s0968-0896(00)82152-5. [DOI] [PubMed] [Google Scholar]
- 27.Mazumder A, Raghavan K, Weinstein J, Kohn KW, Pommier Y. Inhibition of human immunodeficiency virus type-1 integrase by curcumin. Biochemical Pharmacology. 1995;49(8):1165–1170. doi: 10.1016/0006-2952(95)98514-a. [DOI] [PubMed] [Google Scholar]
- 28.Balasubramanyam K, Varier RA, Altaf M, et al. Curcumin, a novel p300/CREB-binding protein-specific inhibitor of acetyltransferase, represses the acetylation of histone/nonhistone proteins and histone acetyltransferase-dependent chromatin transcription. The Journal of Biological Chemistry. 2004;279(49):51163–51171. doi: 10.1074/jbc.M409024200. [DOI] [PubMed] [Google Scholar]
- 29.James JS. Curcumin: clinical trial finds no antiviral effect. AIDS Treatment News. 1996;(242):1–2. [PubMed] [Google Scholar]
- 30.Chen D-Y, Shien J-H, Tiley L, et al. Curcumin inhibits influenza virus infection and haemagglutination activity. Food Chemistry. 2010;119(4):1346–1351. [Google Scholar]
- 31.Zandi K, Ramedani E, Mohammadi K, et al. Evaluation of antiviral activities of curcumin derivatives against HSV-1 in Vero cell line. Natural Product Communications. 2010;5(12):1935–1938. [PubMed] [Google Scholar]
- 32.Kutluay SB, Doroghazi J, Roemer ME, Triezenberg SJ. Curcumin inhibits herpes simplex virus immediate-early gene expression by a mechanism independent of p300/CBP histone acetyltransferase activity. Virology. 2008;373(2):239–247. doi: 10.1016/j.virol.2007.11.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bourne KZ, Bourne N, Reising SF, Stanberry LR. Plant products as topical microbicide candidates: assessment of in vitro and in vivo activity against herpes simplex virus type 2. Antiviral Research. 1999;42(3):219–226. doi: 10.1016/s0166-3542(99)00020-0. [DOI] [PubMed] [Google Scholar]
- 34.Si X, Wang Y, Wong J, Zhang J, McManus BM, Luo H. Dysregulation of the ubiquitin-proteasome system by curcumin suppresses coxsackievirus B3 replication. Journal of Virology. 2007;81(7):3142–3150. doi: 10.1128/JVI.02028-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim HJ, Yoo HS, Kim JC, et al. Antiviral effect of Curcuma longa Linn extract against hepatitis B virus replication. Journal of Ethnopharmacology. 2009;124(2):189–196. doi: 10.1016/j.jep.2009.04.046. [DOI] [PubMed] [Google Scholar]
- 36.Kim K, Kim KH, Kim HY, Cho HK, Sakamoto N, Cheong J. Curcumin inhibits hepatitis C virus replication via suppressing the Akt-SREBP-1 pathway. FEBS Letters. 2010;584(4):707–712. doi: 10.1016/j.febslet.2009.12.019. [DOI] [PubMed] [Google Scholar]
- 37.Divya CS, Pillai MR. Antitumor action of curcumin in human papillomavirus associated cells involves downregulation of viral oncogenes, prevention of NFkB and AP-1 translocation, and modulation of apoptosis. Molecular Carcinogenesis. 2006;45(5):320–332. doi: 10.1002/mc.20170. [DOI] [PubMed] [Google Scholar]
- 38.Prusty BK, Das BC. Constitutive activation of transcription factor AP-1 in cervical cancer and suppression of human papillomavirus (HPV) transcription and AP-1 activity in HeLa cells by curcumin. International Journal of Cancer. 2005;113(6):951–960. doi: 10.1002/ijc.20668. [DOI] [PubMed] [Google Scholar]
- 39.Dutta K, Ghosh D, Basu A. Curcumin protects neuronal cells from japanese encephalitis virus-mediated cell death and also inhibits infective viral particle formation by dysregulation of ubiquitin-proteasome system. Journal of Neuroimmune Pharmacology. 2009;4(3):328–337. doi: 10.1007/s11481-009-9158-2. [DOI] [PubMed] [Google Scholar]
- 40.Tomita M, Kawakami H, Uchihara J-N, et al. Curcumin suppresses constitutive activation of AP-1 by downregulation of JunD protein in HTLV-1-infected T-cell lines. Leukemia Research. 2006;30(3):313–321. doi: 10.1016/j.leukres.2005.08.004. [DOI] [PubMed] [Google Scholar]
- 41.De R, Kundu P, Swarnakar S, et al. Antimicrobial activity of curcumin against helicobacter pylori isolates from India and during infections in mice. Antimicrobial Agents and Chemotherapy. 2009;53(4):1592–1597. doi: 10.1128/AAC.01242-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Foryst-Ludwig A, Neumann M, Schneider-Brachert W, Naumann M. Curcumin blocks NF-κB and the motogenic response in Helicobacter pylori-infected epithelial cells. Biochemical and Biophysical Research Communications. 2004;316(4):1065–1072. doi: 10.1016/j.bbrc.2004.02.158. [DOI] [PubMed] [Google Scholar]
- 43.Kundu P, De R, Pal I, Mukhopadhyay AK, Saha DR, Swarnakar S. Curcumin alleviates matrix metalloproteinase-3 and -9 activities during eradication of Helicobacter pylori infection in cultured cells and mice. PLoS ONE. 2011;6(1) doi: 10.1371/journal.pone.0016306.e16306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Koosirirat C, Linpisarn S, Changsom D, Chawansuntati K, Wipasa J. Investigation of the anti-inflammatory effect of Curcuma longa in Helicobacter pylori-infected patients. International Immunopharmacology. 2010;10(7):815–818. doi: 10.1016/j.intimp.2010.04.021. [DOI] [PubMed] [Google Scholar]
- 45.Di Mario F, Cavallaro LG, Nouvenne A, et al. A curcumin-based 1-week triple therapy for eradication of Helicobacter pylori infection: something to learn from failure? Helicobacter. 2007;12(3):238–243. doi: 10.1111/j.1523-5378.2007.00497.x. [DOI] [PubMed] [Google Scholar]
- 46.Sintara K, Thong-Ngam D, Patumraj S, Klaikeaw N, Chatsuwan T. Curcumin suppresses gastric NF-κB activation and macromolecular leakage in Helicobacter pylori-infected rats. World Journal of Gastroenterology. 2010;16(32):4039–4046. doi: 10.3748/wjg.v16.i32.4039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Odds FC. Synergy, antagonism, and what the chequerboard puts between them. Journal of Antimicrobial Chemotherapy. 2003;52(1):p. 1. doi: 10.1093/jac/dkg301. [DOI] [PubMed] [Google Scholar]
- 48.Amrouche T, Noll KS, Wang Y, Huang Q, Chikindas ML. Antibacterial activity of subtilosin alone and combined with curcumin, poly-lysine and zinc lactate against listeria monocytogenes strains. Probiotics and Antimicrobial Proteins. 2010;2(4):250–257. doi: 10.1007/s12602-010-9042-7. [DOI] [PubMed] [Google Scholar]
- 49.Moghaddam KM, Iranshahi M, Yazdi MC, Shahverdi AR. The combination effect of curcumin with different antibiotics against Staphylococcus aureus . International Journal of Green Pharmacy. 2009;3(2):141–143. [Google Scholar]
- 50.Marathe SA, Kumar R, Ajitkumar P, Nagaraja V, Chakravortty D. Curcumin reduces the antimicrobial activity of ciprofloxacin against Salmonella Typhimurium and Salmonella Typhi . Journal of Antimicrobial Chemotherapy. 2013;68(1):139–152. doi: 10.1093/jac/dks375. [DOI] [PubMed] [Google Scholar]
- 51.Hatamie S, Nouri M, Karandikar SK, et al. Complexes of cobalt nanoparticles and polyfunctional curcumin as antimicrobial agents. Materials Science and Engineering C. 2012;32(2):92–97. [Google Scholar]
- 52.Varaprasad K, Mohan YM, Vimala K, Mohana Raju K. Synthesis and characterization of hydrogel-silver nanoparticle-curcumin composites for wound dressing and antibacterial application. Journal of Applied Polymer Science. 2011;121(2):784–796. [Google Scholar]
- 53.Vimala K, Mohan YM, Varaprasad K, et al. Fabrication of curcumin encapsulated chitosan-PVA silver nanocomposite films for improved antimicrobial activity. Journal of Biomaterials and Nanobiotechnology. 2011;2(1):55–64. [Google Scholar]
- 54.Karaman M, Fırıncı F, Arıkan Ayyıldız Z, Bahar IH. Effects of Imipenem, Tobramycin and Curcumin on biofilm formation of Pseudomonas aeruginosa strains. Mikrobiyoloji Bulteni. 2013;47(1):192–194. doi: 10.5578/mb.3902. [DOI] [PubMed] [Google Scholar]
- 55.Marathe SA, Ray S, Chakravortty D. Curcumin increases the pathogenicity of Salmonella enterica serovar typhimurium in Murine model. PLoS ONE. 2010;5(7) doi: 10.1371/journal.pone.0011511.e11511 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Tomei L, Altamura S, Paonessa G, De Francesco R, Migliaccio G. HCV antiviral resistance: the impact of in vitro studies on the development of antiviral agents targeting the viral NS5B polymerase. Antiviral Chemistry & Chemotherapy. 2005;16(4):225–245. doi: 10.1177/095632020501600403. [DOI] [PubMed] [Google Scholar]
- 57.Lemoine M, Nayagam S, Thursz M. Viral hepatitis in resource-limited countries and access to antiviral therapies: current and future challenges. Future Virology. 2013;8(4):371–380. doi: 10.2217/fvl.13.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.De Clercq E. Strategies in the design of antiviral drugs. Nature Reviews Drug Discovery. 2002;1(1):13–25. doi: 10.1038/nrd703. [DOI] [PubMed] [Google Scholar]
- 59.Jassim SAA, Naji MA. Novel antiviral agents: a medicinal plant perspective. Journal of Applied Microbiology. 2003;95(3):412–427. doi: 10.1046/j.1365-2672.2003.02026.x. [DOI] [PubMed] [Google Scholar]
- 60.Zorofchian Moghadamtousi S, Hajrezaei M, Abdul Kadir H, Zandi K. Loranthus micranthus Linn.: biological activities and phytochemistry. Evidence-Based Complementary and Alternative Medicine. 2013;2013:9 pages. doi: 10.1155/2013/273712.273712 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Dairaku I, Han Y, Yanaka N, Kato N. Inhibitory effect of curcumin on IMP dehydrogenase, the target for anticancer and antiviral chemotherapy agents. Bioscience, Biotechnology and Biochemistry. 2010;74(1):185–187. doi: 10.1271/bbb.90568. [DOI] [PubMed] [Google Scholar]
- 62.Singh RK, Rai D, Yadav D, Bhargava A, Balzarini J, De Clercq E. Synthesis, antibacterial and antiviral properties of curcumin bioconjugates bearing dipeptide, fatty acids and folic acid. European Journal of Medicinal Chemistry. 2010;45(3):1078–1086. doi: 10.1016/j.ejmech.2009.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Nabel GJ, Rice SA, Knipe DM, Baltimore D. Alternative mechamisms for activation of human immunodeficiency virus enhancer in T cells. Science. 1988;239(4845):1299–1302. doi: 10.1126/science.2830675. [DOI] [PubMed] [Google Scholar]
- 64.Cullen BR, Greene WC. Regulatory pathways governing HIV-1 replication. Cell. 1989;58(3):423–426. doi: 10.1016/0092-8674(89)90420-0. [DOI] [PubMed] [Google Scholar]
- 65.Mazumder A, Neamati N, Sunder S, et al. Curcumin analogs with altered potencies against HIV-1 integrase as probes for biochemical mechanisms of drug action. Journal of Medicinal Chemistry. 1997;40(19):3057–3063. doi: 10.1021/jm970190x. [DOI] [PubMed] [Google Scholar]
- 66.Chauhan G, Rath G, Goyal AK. In-vitro anti-viral screening and cytotoxicity evaluation of copper-curcumin complex. Artificial Cells, Nanomedicine and Biotechnology. 2013;41(4):276–281. doi: 10.3109/21691401.2012.742096. [DOI] [PubMed] [Google Scholar]
- 67.Kawai C. From myocarditis to cardiomyopathy: mechanisms of inflammation and cell death: learning from the past for the future. Circulation. 1999;99(8):1091–1100. doi: 10.1161/01.cir.99.8.1091. [DOI] [PubMed] [Google Scholar]
- 68.Lee C-K, Kono K, Haas E, et al. Characterization of an infectious cDNA copy of the genome of a naturally occurring, avirulent coxsackievirus B3 clinical isolate. Journal of General Virology. 2005;86(1):197–210. doi: 10.1099/vir.0.80424-0. [DOI] [PubMed] [Google Scholar]
- 69.Si X, McManus BM, Zhang J, et al. Pyrrolidine dithiocarbamate reduces coxsackievirus B3 replication through inhibition of the ubiquitin-proteasome pathway. Journal of Virology. 2005;79(13):8014–8023. doi: 10.1128/JVI.79.13.8014-8023.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Ganem D, Prince AM. Hepatitis B virus infection—natural history and clinical consequences. The New England Journal of Medicine. 2004;350(11):1118–1129. doi: 10.1056/NEJMra031087. [DOI] [PubMed] [Google Scholar]
- 71.Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics, 2002. CA: A Cancer Journal for Clinicians. 2005;55(2):74–108. doi: 10.3322/canjclin.55.2.74. [DOI] [PubMed] [Google Scholar]
- 72.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA: A Cancer Journal for Clinicians. 2011;61(2):69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 73.Chen C-J, Raung S-L, Kuo M-D, Wang Y-M. Suppression of Japanese encephalitis virus infection by non-steroidal anti-inflammatory drugs. Journal of General Virology. 2002;83(8):1897–1905. doi: 10.1099/0022-1317-83-8-1897. [DOI] [PubMed] [Google Scholar]
- 74.Fujii M, Niki T, Mori T, et al. HTLV-1 tax induces expression of various immediate early serum responsive genes. Oncogene. 1991;6(6):1023–1029. [PubMed] [Google Scholar]
- 75.Upendra RS, Khandelwal P, Reddy AHM. Turmeric powder (Curcuma longa Linn.) as an antifungal agent in plant tissue culture studies. International Journal of Engineering Science. 2011;3(11):7899–7904. [Google Scholar]
- 76.Kim M-K, Choi G-J, Lee H-S. Fungicidal property of Curcuma longa L. rhizome-derived curcumin against phytopathogenic fungi in a greenhouse. Journal of Agricultural and Food Chemistry. 2003;51(6):1578–1581. doi: 10.1021/jf0210369. [DOI] [PubMed] [Google Scholar]
- 77.Chowdhury H, Banerjee T, Walia S. In vitro screening of Curcuma longa L and its derivatives sa antifungal agents against Helminthosporrum oryzae and Fusarium solani . Pesticide Research Journal. 2008;20(1):6–9. [Google Scholar]
- 78.Wuthi-udomlert M, Grisanapan W, Luanratana O, Caichompoo W. Antifungal activity of Curcuma longa grown in Thailand. The Southeast Asian Journal of Tropical Medicine and Public Health. 2000;31(1):178–182. [PubMed] [Google Scholar]
- 79.Apisariyakul A, Vanittanakom N, Buddhasukh D. Antifungal activity of turmeric oil extracted from Curcuma longa (Zingiberaceae) Journal of Ethnopharmacology. 1995;49(3):163–169. doi: 10.1016/0378-8741(95)01320-2. [DOI] [PubMed] [Google Scholar]
- 80.Martins CVB, Da Silva DL, Neres ATM, et al. Curcumin as a promising antifungal of clinical interest. Journal of Antimicrobial Chemotherapy. 2009;63(2):337–339. doi: 10.1093/jac/dkn488. [DOI] [PubMed] [Google Scholar]
- 81.Sharma M, Manoharlal R, Puri N, Prasad R. Antifungal curcumin induces reactive oxygen species and triggers an early apoptosis but prevents hyphae development by targeting the global repressor TUP1 in Candida albicans . Bioscience Reports. 2010;30(6):391–404. doi: 10.1042/BSR20090151. [DOI] [PubMed] [Google Scholar]
- 82.Neelofar K, Shreaz S, Rimple B, Muralidhar S, Nikhat M, Khan LA. Curcumin as a promising anticandidal of clinical interest. Canadian Journal of Microbiology. 2011;57(3):204–210. doi: 10.1139/W10-117. [DOI] [PubMed] [Google Scholar]
- 83.Jianhua W, Hai W. Antifungal susceptibility analysis of berberine, baicalin, eugenol and curcumin on Candida albicans . Journal of Medical Colleges of PLA. 2009;24(3):142–147. [Google Scholar]
- 84.Khan N, Shreaz S, Bhatia R, et al. Anticandidal activity of curcumin and methyl cinnamaldehyde. Fitoterapia. 2012;83(3):434–440. doi: 10.1016/j.fitote.2011.12.003. [DOI] [PubMed] [Google Scholar]
- 85.Sharma M, Manoharlal R, Negi AS, Prasad R. Synergistic anticandidal activity of pure polyphenol curcumin i in combination with azoles and polyenes generates reactive oxygen species leading to apoptosis. FEMS Yeast Research. 2010;10(5):570–578. doi: 10.1111/j.1567-1364.2010.00637.x. [DOI] [PubMed] [Google Scholar]
- 86.Karaman M, Arıkan Ayyıldız Z, Fırıncı F, et al. Effects of curcumin on lung histopathology and fungal burden in a mouse model of chronic asthma and oropharyngeal candidiasis. Archives of Medical Research. 2011;42(2):79–87. doi: 10.1016/j.arcmed.2011.01.011. [DOI] [PubMed] [Google Scholar]
- 87.Dovigo LN, Pavarina AC, Carmello JC, MacHado AL, Brunetti IL, Bagnato VS. Susceptibility of clinical isolates of Candida to photodynamic effects of curcumin. Lasers in Surgery and Medicine. 2011;43(9):927–934. doi: 10.1002/lsm.21110. [DOI] [PubMed] [Google Scholar]
- 88.Dovigo LN, Pavarina AC, Ribeiro APD, et al. Investigation of the photodynamic effects of curcumin against Candida albicans . Photochemistry and Photobiology. 2011;87(4):895–903. doi: 10.1111/j.1751-1097.2011.00937.x. [DOI] [PubMed] [Google Scholar]
- 89.Dovigo LN, Carmello JC, de Souza Costa CA, et al. Curcumin-mediated photodynamic inactivation of Candida albicans in a murine model of oral candidiasis. Medical Mycology. 2013;51(3):243–251. doi: 10.3109/13693786.2012.714081. [DOI] [PubMed] [Google Scholar]
- 90.Tsao S-M, Yin M-C. Enhanced inhibitory effect from interaction of curcumin with amphotericin B or fluconazole against Candida species. Journal of Food and Drug Analysis. 2000;8(3):208–212. [Google Scholar]
- 91.Kudva AK, Manoj MN, Swamy BN, Ramadoss CS. Complexation of amphoterecin B and curcumin with serum albumin: solubility and effect on erythrocyte membrane damage. Journal of Experimental Pharmacology. 2011;3:1–6. doi: 10.2147/JEP.S15078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Khalil OAK, de Faria Oliveira OMM, Vellosa JCR, et al. Curcumin antifungal and antioxidant activities are increased in the presence of ascorbic acid. Food Chemistry. 2012;133(3):1001–1005. [Google Scholar]
- 93.Mohanty C, Das M, Sahoo SK. Emerging role of nanocarriers to increase the solubility and bioavailability of curcumin. Expert Opinion on Drug Delivery. 2012;9(11):1347–1364. doi: 10.1517/17425247.2012.724676. [DOI] [PubMed] [Google Scholar]
- 94.Kurien BT, Singh A, Matsumoto H, Scofield RH. Improving the solubility and pharmacological efficacy of curcumin by heat treatment. Assay and Drug Development Technologies. 2007;5(4):567–576. doi: 10.1089/adt.2007.064. [DOI] [PubMed] [Google Scholar]
- 95.Bhawana B, Basniwal RK, Buttar HS, Jain VK, Jain N. Curcumin nanoparticles: preparation, characterization, and antimicrobial study. Journal of Agricultural and Food Chemistry. 2011;59(5):2056–2061. doi: 10.1021/jf104402t. [DOI] [PubMed] [Google Scholar]
- 96.Shailendiran D, Pawar N, Chanchal A, Pandey RP, Bohidar HB, Verma AK. Characterization and antimicrobial activity of nanocurcumin and curcumin. Proceedings of the International Conference on Nanoscience, Technology and Societal Implications (NSTSI '11); December 2011; IEEE; pp. 1–7. [Google Scholar]
- 97.Wang Y, Lu Z, Wu H, Lv F. Study on the antibiotic activity of microcapsule curcumin against foodborne pathogens. International Journal of Food Microbiology. 2009;136(1):71–74. doi: 10.1016/j.ijfoodmicro.2009.09.001. [DOI] [PubMed] [Google Scholar]
- 98.Wang Y-F, Shao J-J, Zhou C-H, et al. Food preservation effects of curcumin microcapsules. Food Control. 2012;27(1):113–117. [Google Scholar]
- 99.Hatcher H, Planalp R, Cho J, Torti FM, Torti SV. Curcumin: from ancient medicine to current clinical trials. Cellular and Molecular Life Sciences. 2008;65(11):1631–1652. doi: 10.1007/s00018-008-7452-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Gupta SC, Kismali G, Aggarwal BB. Curcumin, a component of turmeric: from farm to pharmacy. Biofactors. 2013;39(1):2–13. doi: 10.1002/biof.1079. [DOI] [PubMed] [Google Scholar]