

Abstract
The serotonin and circadian systems are two important interactive regulatory networks in the mammalian brain that regulate behavior and physiology in ways that are known to impact human mental health. Previous work on the interaction between these two systems suggests that serotonin modulates photic input to the central circadian clock (the suprachiasmatic nuclei; SCN) from the retina and serves as a signal for locomotor activity, novelty, and arousal to shift the SCN clock, but effects of disruption of serotonergic signaling from the raphe nuclei on circadian behavior and on SCN function are not fully characterized. In this study, we examined the effects on diurnal and circadian behavior, and on ex vivo molecular rhythms of the SCN, of genetic deficiency in Pet-1, an ETS transcription factor that is necessary to establish and maintain the serotonergic phenotype of raphe neurons. Pet-1−/− mice exhibit loss of rhythmic behavioral coherence and an extended daily activity duration, as well as changes in the molecular rhythms expressed by the clock, such that ex vivo SCN from Pet-1− /− mice exhibit period lengthening and sex-dependent changes in rhythmic amplitude. Together, our results indicate that Pet-1 regulation of raphe neuron serotonin phenotype contributes to the period, precision and light/dark partitioning of locomotor behavioral rhythms by the circadian clock through direct actions on the SCN clock itself, as well as through non-clock effects.
Introduction
The serotonin (5-hydroxytryptamine, or 5-HT) and circadian systems are two important, interactive regulatory networks in the mammalian brain (for review, see [1]). Both systems exert influence on multiple neural centers to regulate behavior, and originate in specific nuclei in the nervous system. Serotonin, a monoamine neurotransmitter, is synthesized in the brain primarily in neurons of the midbrain and medullary raphe nuclei. The dorsal and median raphe nuclei send projections from the brainstem and midbrain throughout the brain, and interact both directly and indirectly with the central circadian clock, the suprachiasmatic nuclei (SCN) of the hypothalamus, from which they receive reciprocal indirect connections. The serotonergic phenotype of neurons of the raphe requires the expression of Pet-1, an ETS proto-oncogene transcription factor that activates the expression of the enzyme tryptophan hydroxylase (TpH2), a key molecule in the 5-HT biosynthetic pathway, as well as other genes that are necessary for serotonergic specification during development [2]. Dysregulation of serotonergic signaling profoundly impacts human mental and physical health, and is a primary contributor to mood and developmental disorders.
Mammalian circadian rhythms are organized by a ∼24 hour biological clock located in the suprachiasmatic nuclei of the hypothalamus, which are composed of some 20,000 tightly packed neurons [3]. Individual SCN neurons can act as self-sustained pacemakers [4], expressing rhythms in spike frequency and gene expression in an autoregulatory timing loop that cycles over the course of a day [5]. The SCN acts to organize the intrinsic rhythms of brain regions and peripheral tissues to regulate behavior and physiology. Like the serotonergic system, circadian dysfunction is associated with a range of sleep, mood, and developmental disorders (for review, see [6], [7]).
In rodents, wheel-running, novelty, and arousal can reset the SCN clock [8], [9], although the clock is set primarily by light [10]. These non-photic behavioral inputs are thought to arise partially from the median raphe nucleus and partially from the intergeniculate leaflet and use serotonin and NPY as neurotransmitters, respectively [11], [12], [13], [14], [15], [16], [17], [18], where the IGL itself receives serotonergic input from the dorsal raphe [19]. SCN neurons themselves express the serotonin re-uptake transporter (SERT) and the serotonin 5-HT1B, 5-HT7 and 5-HT2C receptors [13], [20], [21], [22], [23], [24], [25], [26] where they mediate serotonin's effects on circadian light responses [15], [27], [28] and circadian rhythms [11], [12], [13], [14], [15], [16], [17]. In the SCN, serotonin modulates glutamatergic light signaling [28] and can phase shift rhythms directly [13], [29] (for review, see [30]). Chemical depletion of serotonin in hamsters, results in maintained rhythms and increases their activity duration in light cycles [31]; likewise, chemical depletion of 5-HT in mice also increases activity duration and alters their ability to entrain to forced treadmill running, even when activity levels are kept constant [32]. Furthermore, behavioral serotonin release, such as that caused by novelty-induced exercise, can inhibit light-induced behavioral phase shifts [33]. Together, these results suggest that serotonin works antagonistically to photic stimulation of the SCN [34], [35], and its depletion in adult rodents alters the clock control of activity in relationship to the prevailing light cycle. The effects on the circadian system of Pet-1 deficiency and altered serotonin signaling during brain development have not been extensively studied.
In the brain, serotonin signaling is turned on early in mammalian development—E10–12 in mouse [36] or during the first month of gestation in primates [37]. Circadian rhythms, in contrast, appear significantly later during postnatal development. Here, we have used mice with germline knockout of ETS transcription factor Pet-1 that is necessary for the development of raphe serotonin neurons [38]. Pet-1 −/− mice have about an 75% reduction in the number of dorsal and median raphe serotonin neurons that express key serotonin signaling genes, such as Tph2, SERT and 5HT1a. The serotonin projection from the raphe nuclei to the SCN is specifically lost in Pet-1−/− mice [39], [40].
Pet-1− /− mice on a mixed genetic background exhibit circadian behavioral abnormalities and abnormal temporal organization [39], [41]. We have examined the effects of this brain specific serotonergic deficiency on diurnal and circadian behavior in a congenic mouse strain (C57Bl/6J), and on ex vivo SCN bioluminescence rhythms. Our results indicate that genetic loss of Pet-1 and serotonergic innervation of the SCN leads to specific changes in diurnal and circadian behavior such as an expansion of activity duration, rhythmic instability, and a treatment-dependent change in free-running period. There are also changes in the period of the isolated SCN ex vivo, although these changes are not consistently mirrored by the characteristics of Pet-1−/− mouse behavior, indicating both SCN and non-SCN effects on circadian control of locomotor behavior by genetic disruption of serotonergic signaling. Sex-dependent effects of the genetic Pet-1 deficiency on the SCN pacemaker are also evident, suggesting interactions of serotonergic and hormonal regulation of this clock tissue.
Materials and Methods
Ethics Statement
All handling of animals was approved by the Vanderbilt University Institutional Animal Care and Use Committee (IACUC) in accordance with National Institutes of Health (NIH) guidelines.
Animals and Housing
129sv mice containing a Pet-1 null allele [38] were backcrossed twelve generations to C57BI/6J (Jackson) mice to yield a congenic strain for behavioral characterization (males only). These mice were then crossed to a congenic C57Bl/6J Per1::Luciferase [42] reporter mouse line, and both males and females were used for LumiCycle experiments. To avoid maternal-rearing complications from Pet-1− /− females [43], Pet-1+/− males and females were bred to attain Pet-1 +/+, Pet-1 +/− and Pet-1− /− offspring. The breeding Pet-1 +/− females were not naïve mothers, having had at least one litter prior to the use of subsequent litters in these experiments. For behavioral characterization, mice were bred in a 12∶12 Light:Dark (LD) cycle. Pups were weaned at postnatal day ∼21 and kept together for 1–2 weeks in a litter-filled group cage with food and water ad libitum while they were genotyped as described previously [38]. Animals were then placed individually into litter-filled poly-propylene wheel cages (Coulbourn Instruments, Whitehall, PA) equipped with infrared (IR) motion detector (Spy2 from Visonic, Tel Aviv, Israel) and food and water ad libitum. For organotypic slice culture, male and female mice were bred and kept in one of three photoperiods: a short LD 8∶16, an equinox LD 12∶12, or a long LD 16∶8 photoperiod for 50 days. At weaning, mice were housed individually in litter-filled poly-propylene wheel cages with food and water ad libitum.
Behavioral Characterization
To determine the effect of Pet-1 deficiency on circadian behavioral properties in conjunction with and independently of wheel activity, we monitored activity in 5-min bins using ClockLab Software (Actimetrics, Evanston, IL) using simultaneous wheel and IR. Specifically, male mice were given 1-2 weeks to acclimate to their cages and entrain to a LD 12∶12 light-cycle (phase 1), after which the mice were split into two groups that were subjected to an identical light paradigm, but with differing wheel-state to control for order-effects. Each animal was subjected to 5 distinct phases in which the mouse had access to a freely turning wheel or in which the wheel was locked ( Figure 1 ). The first group (Paradigm A), had the following order of wheel-state: open for about 2 weeks (phase 2), locked for about 3–4 weeks (phase 3: same LD 12∶12 as phases 1–2; phase 4: constant darkness, DD), then finally open in constant darkness for about 2 weeks (phase 5). The second group (Paradigm B) had exactly the opposite order, to test or control for the possibility of order-effects ( Figure 1a ). The final 12 days from each phase were analyzed in ClockLab. We used IR recordings rather than wheel recordings for analysis because it provided activity monitoring during all components of each paradigm.
Figure 1. Wheel open/lock circadian behavioral paradigms.

a | Double-plotted graphics representing two equal light cycle paradigms (12L:12D→DD) with opposite order of wheel access in each phase. b | Representative double-plotted actograms showing the monitored activity of mice that are, from left to right, wildtype, heterozygote, or knockout for Pet-1, as monitored by infrared (IR) motion detection (top) and wheel (bottom). Note that there is no wheel activity during wheel-locked phases. The Pet-1 +/+ mouse (left) represents paradigm A, while the Pet-1 +/− and Pet-1 −/− mice (middle and right, respectively) represents paradigm B. Y-axis represents days; X-axis represents time-of-day.
Behavioral Analysis
IR behavior was analyzed using ClockLab Analysis software (Actimetrics), and activity was quantified as the number of IR activations occurring during 5-min bins. The free-running period (in DD, over 12 days), rhythmic power, alpha, amount of activity during lights-on and lights-off, as well as the total amount of activity per day were determined for each phase using the chi-squared periodogram and activity profile options in the “Batch Analysis” function, and by eye-fit. Phases of activity onset and offset were eye-fit using ClockLab's actogram function. In addition, rhythmic power was determined by using a Chi-squared periodogram that compared amplitude of behavioral rhythms (as in [44]).
Tissue Preparation for ex vivo Organotypic Slice Cultures
The mice were euthanized by cervical dislocation and rapid decapitation between ZT 9-12. The brains were extracted and blocked in cold sampling media as previously described [45]. 300µm hypothalamic coronal sections containing the suprachiasmatic nuclei (SCN) were cut using a vibroslicer (Campden Instruments Ltd., Lafayette, IN) in Hank's balanced salt solution (HBSS, Gibco, Carlsbad, CA) enriched with 25 U/mL penicillin/streptomycin, NaHCO3 (7.5%; Sigma, St. Louis, MO) and 1.0 M HEPES kept at ∼4°C. Slices were then trimmed and transferred to culture membranes (Millicell, Millipore, Billerica, MA) in 35 mm culture dishes containing 1 mL of Dulbecco's modified Eagle's medium – low glucose (DMEM D2902-10x1L; Sigma-Aldrich, St. Louis, MO) supplemented with 25 U/mL penicillin/streptomycin, 1.0 M HEPES, 4.5 g/L glucose, 7.5% NaHCO3, N-2 supplement (Invitrogen), and 0.1 mM beetle luciferin (Promega, Madison, WI). Dishes were sealed and placed directly into a LumiCycle (Actimetrics) kept at 35.5°C. Rhythms were analyzed using the LumiCycle data analysis software (Actimetrics) where a baseline subtraction was performed using a fitted polynomial curve to the first six cycles of the raw data with an order value within one cycle of the six cycles to determine the best fit line. The period length was then calculated by determining the maximum and minimum time points (in hours) for Per1::LUC expression [42]. Rhythmic power was also examined using the LumiCycle and ClockLab data analysis software (Actimetrics) using a Chi-squared periodogram that compared amplitude to rhythms.
Statistical Analysis
Means were statistically compared in IBM SPSS 20 (IBM, Armonk, NY) by univariate analyses including repeated measures, with post-hoc tests. For data with non-homogeneous distributions, the Sheirer-Ray-Hare extension of the nonparametric Kruskal-Wallis Test was performed [46]. Post-hoc Fisher's Least Significant Difference tests were performed on parametric data and post-hoc Dunnett's T3 test were performed on non-parametric data. Significance was ascribed at p<0.05.
Results
To determine the influence of genetic disruption of brain serotonergic signaling by Pet-1 deficiency on circadian behavior and how it may modulate the feedback effects of wheel-running on circadian rhythms, we examined knockout (Pet-1−/−; N = 16), heterozygous (Pet-1+/−; N = 14) and wildtype (Pet-1+/+; N = 9) mice in a series of light/dark cycles combined with locking or unlocking the running wheels available to these individually housed mice ( Figure 1 ). We assayed activity both with the running wheel (when unlocked = open) and with infrared motion detection. The activity signal readout by infrared motion detectors (IR) persists with both open and locked wheels, and records both on-wheel and off-wheel locomotor activity ( Figure 1 ). Visual inspection of the actograms revealed striking alterations in the circadian behavior of Pet-1 −/− mice. There were clear differences in the partitioning of day/night activity in LD and in the coherence of free-running rhythms in DD that were quantified with more detailed analysis below.
Pet-1−/− mice exhibit expanded locomotor activity duration
Pet-1−/− mice exhibited clear differences in the distribution of activity across the day in LD, with significant decrements in night-time activity and increased day-time activity ( Figure 2 ). Pet-1 −/− mice exhibited an expanded alpha, or activity duration, in both a light-dark cycle (LD) and constant darkness (DD), as measured with both an infrared motion detector (IR; Figure 3a,e ) and with a wheel ( Figure 4b,e ). Specifically in LD, an earlier activity onset and a later activity offset is an obvious feature of Pet-1 −/− mouse locomotor behavior ( Figure 2 , Table 1 , and Figure S1). Wheel-running did not affect the length of alpha, or the timing of activity onsets or offsets in either lighting condition, when compared to the locked wheel condition ( Figure 3a,e and Table 2 ). Pet-1−/− mice exhibited significantly less wheel-running activity overall ( Table 1 ), but there was no significant difference in total activity between genotypes as measured by IR ( Table 1 ).
Figure 2. Averaged activity profiles across genotypes as monitored by (a) wheel and (b-d) infrared motion detector (IR) in LD.

Each point represents the average across days and across animals within that particular genotype while in a 12L:12D cycle. Blue points represent Pet-1 +/+ mice; orange points represent Pet-1 +/− mice; black points represent Pet-1 −/− mice. Colored bar (top) represents 24 hour light cycle, where the grey is 12 hours of dark, and the yellow is 12 hours of light. Activity measured in revolutions/hour (a) or counts/hour (b–d). b | Overall IR activity profile of animals regardless of wheel state. These results are further broken-down into IR behavior with a free wheel (c) and with a locked wheel (d). See inset legend (a). Asterisks (*) denote significance at p≤0.05. Note the prominent early morning activity in Pet-1 −/− mice.
Figure 3. Main effects of Pet-1 deficiency and running wheel access on circadian behavioral characteristics in 12L:12D (a-b) and constant darkness (c-f) as measured by an infrared motion detector (IR).

a | Pet-1 −/− mice exhibit a significant increase in length of daily activity duration (alpha) in LD. b | Rhythmic power is unaffected by genotype, as measured by χ2 periodogram, but exercise leads to a significant increase. c | Free-running period of Pet-1 −/− mice is significantly decreased in DD. Furthermore, locking a wheel significantly decreases this period. d | There is a significant interaction between genotype and wheel-access such that as Pet-1 gene dosage decreases, locking the wheel decreases period to an even greater extent. e | As in LD, Pet-1 −/− mice in DD exhibit a greater duration of daily activity. f | Pet-1 −/− mice exhibit reduced rhythmic power, and overall, mice allowed to exercise on a wheel exhibit greater rhythmic power. Error bars represent SEM; asterisks (*) denote significance at p≤0.05.
Figure 4. Main effects of genotype on circadian behavioral characteristics in 12L:12D (a–c) and constant darkness (d–f).

a | The percent of wheel-running activity in the light-phase is significantly increased in Pet-1 −/− mice. b | Pet-1 −/− mice exhibit a significant increase in length of daily wheel-running duration (alpha) in LD for Pet-1 −/− mice. c | Rhythmic power is significantly decreased in Pet-1 −/− mice. d | Free-running period of Pet-1 −/− mice is significantly decreased in DD. As in LD, Pet-1 −/− mice in DD exhibit (e) a greater duration of daily activity and (f) reduced rhythmic power. Error bars represent SEM; asterisks (*) denote significance at p ≤0.05.
Table 1. Main effects of genotype (and photoperiod) on circadian behavioral phenotypes.
| Genotype | main-effects | |||||
| Characteristic | Pet-1+/+ | Pet-1+/− | Pet-1−/− | d.f. | F or Χ2 | p |
| IR Behavioral - LD | ||||||
| Onset (ZT) | 12.0±0.02 | 12.0±0.03 | 11.5±0.09 | 2 | 33.856 | <0.001 |
| OnsetError (h) | 0.18±0.01 | 0.16±0.01 | 0.30±0.03 | 2 | 17.614 | <0.001 |
| Offset (ZT) | 0.4±0.08 | 0.1±0.06 | 1.9±0.13 | 2, 75 | 2.361 | 0.101 |
| OffsetError (h) | 0.21±0.03 | 0.22±0.03 | 0.33±0.02 | 2, 75 | 0.802 | 0.452 |
| Duration (h) | 12.4±0.08 | 12.1±0.07 | 14.3±0.20 | 2 | 57.207 | <0.001 |
| Power (AU) | 1537.4±108.2 | 1588.1±94.6 | 1338.2±36.7 | 2 | 3.246 | 0.197 |
| % Light Activity | 52.0±3.78 | 46.7±1.73 | 53.6±1.08 | 2 | 14.543 | 0.001 |
| Average Activity (counts/min) | 1.41±0.25 | 1.16±0.08 | 1.16±0.06 | 2 | 0.055 | 0.973 |
| Total Activity (counts/day) | 2044.3±362.5 | 1698.5±121.5 | 1675.3±86.6 | 2 | 0.027 | 0.987 |
| Periodic Component Ratio (AU) | 0.192±0.03 | 0.138±0.02 | 0.480±0.02 | 2 | 55.128 | <0.001 |
| IR Behavioral - DD | ||||||
| Period (h) | 23.79±0.02 | 23.76±0.02 | 23.66±0.03 | 2, 75 | 7.886 | 0.001 |
| OnsetError (h) | 0.33±0.03 | 0.47±0.07 | 0.47±0.03 | 2 | 5.726 | 0.057 |
| OffsetError (h) | 0.55±0.03 | 0.53±0.04 | 0.60±0.03 | 2, 75 | 0.972 | 0.383 |
| Duration (h) | 13.0±0.30 | 13.6±0.20 | 16.3±0.20 | 2, 75 | 60.131 | <0.001 |
| Power (AU) | 1273.3±81.1 | 1315.8±90.3 | 1037.8±48.5 | 2 | 8.978 | 0.011 |
| Average Activity (counts/min) | 1.16±0.12 | 1.11±0.06 | 1.20±0.07 | 2 | 0.706 | 0.703 |
| Total Activity (counts/day) | 1661.4±178.9 | 1600.6±83.8 | 1727.3±97.0 | 2 | 0.705 | 0.703 |
| Periodic Component Ratio (AU) | 0.243±0.03 | 0.268±0.03 | 0.587±0.03 | 2,75 | 86.069 | <0.001 |
| Wheel Behavioral - LD | ||||||
| Onset (ZT) | 12.2±0.03 | 12.2±0.03 | 11.7±0.13 | 2 | 15.783 | <0.001 |
| OnsetError (h) | 0.09±0.01 | 0.10±0.01 | 0.40±0.11 | 2 | 7.521 | 0.023 |
| Offset (ZT) | 20.2±0.64 | 21.5±0.31 | 1.8±0.21 | 2, 36 | 73.379 | <0.001 |
| OffsetError (h) | 0.70±0.12 | 0.85±0.08 | 0.41±0.07 | 2, 36 | 7.915 | 0.001 |
| Duration (h) | 8.02±0.64 | 9.27±0.33 | 14.01±0.30 | 2, 36 | 70.566 | <0.001 |
| Power (AU) | 2251.3±70.7 | 2575.4±141.2 | 1589.8±120.1 | 2, 35 | 17.832 | <0.001 |
| % Light Activity | 1.47±0.28 | 0.82±0.29 | 20.10±3.82 | 2 | 25.419 | <0.001 |
| Average Activity (revs/min) | 13.0±0.88 | 16.8±1.46 | 7.5±0.91 | 2, 35 | 17.823 | <0.001 |
| Total Activity (revs/day) | 18790±1273 | 24151±2108 | 10865±1312 | 2, 35 | 17.823 | <0.001 |
| Periodic Component Ratio (AU) | 0.190±0.01 | 0.178±0.01 | 0.397±0.04 | 2 | 19.425 | <0.001 |
| Wheel Behavioral - DD | ||||||
| Period (h) | 23.78±0.05 | 23.78±0.03 | 23.65±0.04 | 2, 36 | 3.771 | 0.033 |
| OnsetError (h) | 0.48±0.20 | 0.36±0.08 | 0.58±0.07 | 2, 36 | 1.227 | 0.305 |
| OffsetError (h) | 0.53±0.07 | 1.00±0.11 | 0.58±0.09 | 2, 36 | 7.128 | 0.002 |
| Duration (h) | 9.61 1.07 | 8.58±0.43 | 15.93±0.33 | 2 | 27.74 | <0.001 |
| Power (AU) | 1961.7±153.8 | 1976.3±169.7 | 1143.0±68.0 | 2 | 16.632 | <0.001 |
| Average Activity (revs/min) | 12.1±0.96 | 13.0±1.49 | 7.2±1.02 | 2, 35 | 7.337 | 0.002 |
| Total Activity (revs/day) | 7008±3524 | 952±947 | 3341±1314 | 2 | 1.263 | 0.532 |
| Periodic Component Ratio (AU) | 0.289±0.03 | 0.252±0.01 | 0.455±0.03 | 2 | 17.922 | <0.001 |
| Ex Vivo Per1::Luc Expression | ||||||
| Period (h) | 24.23±0.07 | 24.24±0.07 | 24.50±0.07 | 2, 155 | 4.675 | 0.011 |
| Power (AU) | 1629.3±76.2 | 1655.6±74.0 | 1623.7±79.6 | 2 | 0.051 | 0.951 |
Activity monitored by infrared motion detector and wheel in LD and DD, as denoted. Ex vivo Per1::Luc expression is below. Significance is ascribed at p ≤0.05.
Table 2. Main effects of exercise on circadian behavioral phenotypes.
| Exercise | main-effects | ||||
| Characteristic | Wheel Open | Wheel Locked | d.f. | t or Z | p |
| IR Behavioral - LD | |||||
| Onset (ZT) | 11.82±0.05 | 11.75±0.08 | 38 | 1.338 | 0.189 |
| OnsetError (h) | 0.201±0.019 | 0.243±0.021 | 38 | −2.605 | 0.013 |
| Offset (ZT) | 0.936±0.171 | 0.844±0.143 | 38 | 0.943 | 0.352 |
| OffsetError (h) | 0.283±0.020 | 0.355±0.021 | 38 | −2.5 | 0.017 |
| Duration (h) | 13.07±0.213 | 13.06±0.207 | 37 | 0.047 | 0.963 |
| Power (AU) | 1611.6±72.90 | 1336.1±48.59 | 38 | 2.847 | 0.004 |
| % Light Activity | 50.12±1.18 | 48.32±2.08 | 38 | 1.341 | 0.188 |
| Average Activity (counts/min) | 1.238±0.113 | 1.196±0.081 | 38 | 0.626 | 0.535 |
| Total Activity (counts/day) | 1803.5±165.1 | 1734.1±116.4 | 38 | 0.703 | 0.486 |
| Periodic Component Ratio (AU) | 0.310±0.03 | 0.267±0.03 | 38 | 2.349 | 0.024 |
| IR Behavioral - DD | |||||
| Period (h) | 23.74±0.022 | 23.71±0.022 | 38 | 2.036 | 0.049 |
| OnsetError (h) | 0.420±0.042 | 0.454±0.042 | 38 | −0.579 | 0.566 |
| OffsetError (h) | 0.512±0.026 | 0.615±0.030 | 38 | −2.662 | 0.011 |
| Duration (h) | 14.64±0.285 | 14.54±0.309 | 38 | 0.535 | 0.595 |
| Power (AU) | 1307.4±74.41 | 1076.5±41.65 | 38 | −2.805 | 0.005 |
| Average Activity (counts/min) | 1.209±0.071 | 1.114±0.056 | 38 | 1.822 | 0.076 |
| Total Activity (counts/day) | 1736.5±101.8 | 1596.8±79.3 | 38 | 1.849 | 0.072 |
| Periodic Component Ratio (AU) | 0.395±0.03 | 0.383±0.03 | 37 | 0.655 | 0.516 |
Activity monitored by infrared motion detector in LD and DD. Significance is ascribed at p ≤0.05.
Pet-1−/− mice exhibit reduced rhythmic behavioral stability in LD
In addition to a significantly altered timing of activity onset and offset in relation to the light cycle, Pet-1 −/− mice exhibited a change in their day-to-day clock stability in LD. Onset error—the variability of when an animal begins its daily activity—was significantly greater in Pet-1 -/- mice in LD, whether measured by IR or wheel-running ( Table 1 ). In addition, while all genotypes of mice displayed greater day-to-day activity onset error when their running wheel was locked in LD, the same measure was not different in DD (p = 0.057; Table 1 ). Pet-1 −/− mice displayed a significant reduction in the coherence of their locomotor rhythms in DD, as measured by rhythmic power ( Figure 3f ), and a similar trend in LD ( Figure 3b ). Wheel-running significantly increased consolidation of rhythms, measured as rhythmic power, in both LD and DD conditions across all genotypes ( Figure 3b,f ). Using a χ2 periodogram analyses to measure periodic behavioral components, we found that Pet-1 −/− mice displayed an increase in the 12-hour component of their activity rhythms leading to significant bimodality of rhythms in a light/dark cycle and in constant darkness (black line, Figure 5 ). This increase in bimodality was present in both the free and locked wheel conditions. When expressed as a ratio of the 12-hour component to the 24-hour component, Pet-1 −/− mice were significantly different from control mice ( Figure 5 ).
Figure 5. χ2 periodogram analyses of genotype-specific and wheel-dependent effects on overall rhythmic amplitude (power) in LD (top) and DD (bottom).

See inset legend (top-left panel) for genotype. Purple line represents significance at p≤0.05. Periodic component ratios measure the ratio of 12-hour:24-hour components on the wheel. Error bars represent SEM; asterisks (*) denote significance at p≤0.05.
Pet-1−/− mice exhibit altered behavioral free-running period
In constant darkness, where the SCN's circadian output can be characterized independent of external clock-setting factors (zeitgebers) like light, we found that free-running period was decreased in Pet-1 −/− mice by about 0.1 hours ( Figures 3c and 4d ). We did not observe significant shortening of free-running period of any genotype in response to an open wheel (Figure S2c), although it has been previously reported that wheel-running shortens free-running period and that effect is dependent on serotonin [11], [14], [47], [48], [49]. We did, however, observe order-effects of wheel exposure (Figure S2a) and interactions of wheel-running after-effects on period with genotype that are consistent with the notion that serotonin mediates feedback of wheel-running activity on circadian behavior (Figure S2, Table S1).
Pet-1−/− mice exhibit altered ex vivo SCN free-running period and rhythmic power
To examine the properties of the SCN circadian pacemaker in isolation, we crossed the Pet-1 knockout mouse line with a circadian reporter mouse line in which the rhythmic activation of the Period1 clock gene promoter is read out as bioluminescence from firefly luciferase (Per1::Luc, [42]). SCN tissue explants from wildtype (N = 54), Pet-1 +/− (N = 57), and Pet-1 −/− (N = 53) genotypes displayed robust circadian rhythms in Per1::luciferase gene expression. Analysis revealed that SCN tissue explants from Pet-1 −/− mice have significantly longer periods (∼0.25 hours) across six circadian cycles in culture compared to controls ( Figure 6 , Table 1 ). We also tested the plasticity of the SCN pacemaker in response to photoperiod, breeding and housing Pet-1 +/+ Per1::Luc, Pet-1 +/− Per1::Luc, and Pet-1 −/− Per1::Luc mice in LD 12∶12 (N = 58), 8∶16 (N = 49), and 16∶8 (N = 57) photoperiods and then assaying SCN bioluminescence rhythms ex vivo. There was a main effect of the photoperiod such that SCN isolated from Pet-1 −/− mice expressed shortened free-running periods of about 0.3 hours following exposure to long, summer-like photoperiods of LD 16∶8 ( Figure 6c ), in agreement with previous results [50].
Figure 6. SCN from Pet-1 −/− Per1::LUC mice exhibit longer period circadian rhythms ex vivo.
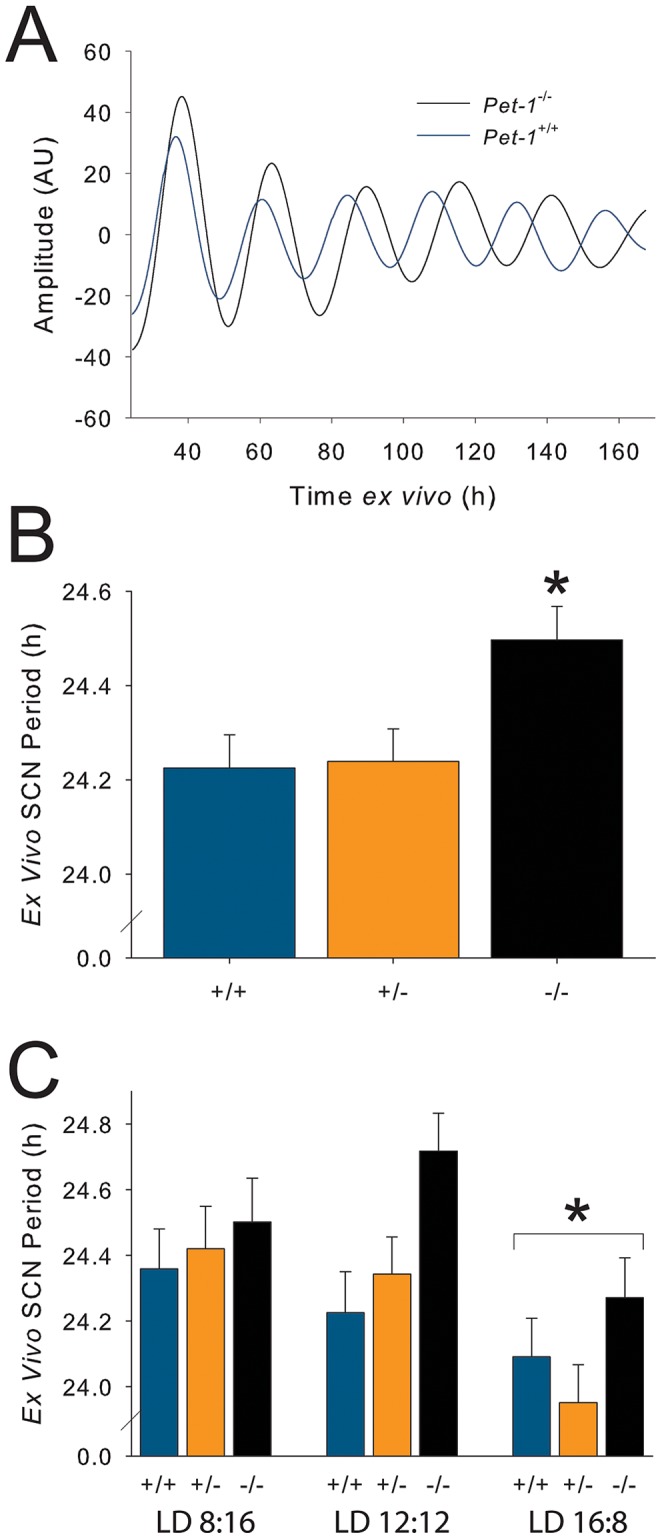
a | Blue line is a representative wildtype SCN; black line is a representative Pet-1 −/− SCN; both taken from a LD 12∶12 cycle. Ex vivo SCN period as a function of Pet-1 genotype (b; main effect across photoperiods) or photoperiod (c). Asterisks (*) denote significance at p ≤0.05.
While only male mice were used in behavioral experiments, as is common practice to avoid the interactive effects of the female estrus cycle on rodent circadian behavioral studies [51], [52], for ex vivo experiments, SCN were isolated from the brains of both sexes. As indicated above, sex did not influence the SCN period in culture, however rhythmic power analysis of the ex vivo SCN rhythms demonstrated an intriguing phenotype. There was an interaction between sex and genotype such that when a Pet-1 null allele is present (Pet-1 +/− and Pet-1 −/−), female (N = 28) SCN showed increased rhythmic power while male (N = 24) SCN rhythms exhibited decreased rhythmic power (p = 0.029; Figure S3).
Discussion
In this study, we sought to address questions regarding the influence of Pet-1, a key gene for central nervous system serotonin signaling on the circadian system. We approached this question by assaying locomotor activity rhythms of mice with genetic knockout of Pet-1 in LD and DD while simultaneous manipulating their ability to use a running-wheel. We then assayed ex vivo cultures of SCN from these mice to assess any differences that a genetic knockdown of serotonin signaling might have on the molecular expression of the rhythm and plasticity in response to photoperiod. Our results indicate that the circadian behaviors of Pet-1 −/− mice are significantly altered, but that these changes are not consistently reflected in the isolated clock nucleus, although its properties are also changed.
Pet-1 −/− mice exhibit significantly expanded locomotor activity over the course of LD 12∶12 light cycles, with increased rhythmic fragmentation and onset error, and increased bimodality in their rhythms. These results are similar to the results obtained with chemical depletion of serotonin in adult hamster [31], in which increased activity duration and fragmentation were also characteristic. While the previous study did not analyze locomotor activity with the same depth, the similarity of results in that study, in which serotonin was only depleted in the adult, and the present study in which serotonin signaling was disrupted during development and in the adult, suggests that the effects seen were primarily due to the ongoing depletion of brain serotonin. In addition the similarity suggests that there may be no additional detectable effect of serotonergic signaling disruption on the circadian system during development. Similar results were also reported by Paulus and Mintz [41] in Pet-1 −/− mice on a mixed genetic background suggesting these aspects of the effects of Pet-1 deficiency on circadian behavior are robust and consistent. Even with the increase in alpha duration, however, there is no increase in total activity over the course of the day in LD or DD ( Table 1 ), dispelling concerns that these mice may be hyperactive. Rather, in LD there is a redistribution of activity during the late night and early day. The fact that the most striking effects are seen in rhythms in LD cycles is consistent with the previously described roles and actions of serotonin in modulating SCN light responses. Consistent with this, is a recent report detailing altered acute light-induced clock resetting in Pet-1 −/− mice on mixed genetic background [41].
The altered period, duration, and fragmentation of behavioral rhythms suggested the potential for a direct effect on the SCN clock of genetic serotonin deficiency. Interestingly, the rhythmic properties of the SCN itself are indeed modified in Pet-1 deficiency, perhaps due to loss of serotonin input during development. However, surprisingly, ex vivo SCN cultures from Pet-1 −/− mice did not reveal altered properties consistent with the observed behavioral effects (e.g. shortened period, waveform damping). In fact, the SCN explants showed lengthening of period, the opposite of the behavioral rhythms. This suggests that although Pet-1 knockout does indeed have genotypic effects on the SCN, those effects are either 1) not preserved in the de-afferented SCN slice, or 2) are preserved, but do not fully account for behavioral effects. There are previous reports of instances in which altered light inputs to the SCN (photoperiod or cycle period) produce opposing effects on the period of circadian behavior and on isolated SCN rhythms [53], [54], [55], [56]. Thus our results are consistent with a loss of Pet-1 perturbing the regulation of light signaling to the SCN. In addition, our results suggest that the perturbations in circadian behavior are mediated in part by disruption of serotonin signaling to other areas of the brain, since they are not accounted for by the phenotype of the Pet-1 −/− SCN itself. We also found that in different photoperiods, the changes in rhythmic period in ex vivo Pet-1 −/− SCN cultures agree with previous results on after-effects of proximal photoperiod in wildtype SCN [10], [50], indicating that the SCN pacemaker retains photoperiodic plasticity even in the absence of serotonin input.
A Pet-1 genotype x sex interaction in setting the rhythmic power of the ex vivo SCN suggests an interaction between serotonergic and hormonal regulation of the SCN pacemaker that may differ between males and females. While a number of studies have reported subtle sex differences in behavioral circadian rhythms and clear effects of gonadectomy on circadian organization (for review, see [62], [63]), studies of hormonal effects on SCN physiology are limited [64]. Both estrogenic and androgenic signaling impacts the serotonergic raphe nuclei [52], and thus may modulate the SCN through the serotonergic median raphe projection that is lost in the Pet-1 −/− mouse. In particular, estrogen acts to increase the circadian light responses of Raphe neurons [65] and thus may influence SCN properties through this serotonergic circuit. However, androgenic, but not estrogenic signaling can also directly influence SCN neurons, with androgen receptors localized in the SCN core region, which also receives the raphe input [62], [66]. Loss of convergent 5HT/androgenic input to the core region of the SCN in males may account for the sex-dependent effect on SCN rhythms.
It is intriguing that the period change of the ex vivo Pet-1 −/− SCN cultures mimics the direction of change seen behaviorally in mixed background Pet-1 −/− mice [39], [41], but not that of the C57Bl/6J background. Paulus & Mintz [39], [41] examined the Pet-1 knockout on a mixed strain of sv129/C57Bl mice that exhibited a significantly longer behavioral period in DD than our results in C57Bl/6J congenic mice. We also behaviorally tested this same mixed strain before backcrossing these mice in our lab, and observed lengthened free-running periods virtually identical to [39], [41] (data not shown), so we are confident that the shortened behavioral free-running period we report here is a phenotype of Pet-1 −/− on the C57Bl/6J background strain. Finally, although we did not observe the previously reported shortening of period with wheel activity in wildtype mice, nor its expected loss in Pet-1 −/− mice [11], [14], [47], [48], [49], we did observe order-effects of wheel exposure on period indicating that there were interactions between wheel-running and serotonergic tone, but perhaps the relatively short duration wheel exposures we used did not reveal their full extent.
It should be noted that the Per1::LUC mouse line we used reports the transcriptional activation of the Per1 promoter in the circadian transcription-translation feedback loop that forms the core molecular clockworks [42]. Whereas it does not directly report the relative phase of other clock genes, it is subject to transcriptional regulation by them and reports the ongoing period of the molecular clockwork oscillations as a whole. Expression of reporters like the one used here have been shown to correlate highly with neuroelectrical properties and in response to rhythmic perturbations [42], [57], [58], [59], [60], [61].
Here we have demonstrated that a loss of behavioral rhythmic coherence and an extended daily activity duration are robust aspects of the Pet-1 −/− circadian phenotype and persist in the face of background strain differences (C57Bl/6J congenic vs. mixed sv129 [39], [41]). These aspects are cardinal features of loss of serotonergic input to the SCN as they are also recapitulated by chemical depletion of serotonin in hamsters [30]. However, specific aspects of the Pet-1 −/− phenotype, such as altered free-running period, are dependent on background strain. Finally, we have demonstrated with ex vivo SCN explants that loss of serotonergic input to the SCN alters clock period and amplitude (sex-dependent), but that neither the disrupted rhythmic coherence, nor the behavioral period change can be explained by effects of Pet-1 deficiency directly on the SCN alone because the isolated SCN rhythms do not recapitulate the fragmentation or period change of the Pet-1 −/− behavioral phenotype, despite being the same genetic background. Thus, the effects of Pet-1 −/− must involve the SCN and other brain regions as well.
Overall, our results suggest that Pet-1 and its regulation of raphe serotonin phenotype contribute to circadian organization in several distinct ways. Its presence enhances the precision of clock-controlled activity onset and offsets, as well as the coherence of behavioral rhythms. In its absence, activity bouts tend to extend into the light phase of each day, rather than being restricted to the dark phase for nocturnally active mice. In addition, Pet-1 regulates both behavioral and SCN period, although these effects were not concordant in our study. Disruption of sleep and circadian rhythms is a hallmark of human mood disorders in which deficiency of serotonergic signaling is also implicated. Pet-1 −/− mice recapitulate features of circadian disruption typical of mood disorders such as extension of activity into the normal sleep/rest phase (i.e. insomnia) as well as exhibiting increased anxiety-like behaviors [38] and therefore may be a productive experimental system for exploring the relationship between circadian disruption and mood disorders.
Supporting Information
Rayleigh plots of activity onset and offset as monitored by IR ( top ) and wheel-running ( bottom ), and when the wheel was open ( left ) and locked ( right ). Blue arrowheads represent Pet-1 +/+ mice; orange arrowheads represent Pet-1 +/− mice; black arrowheads represent Pet-1 +/+ mice; arrows represent the mean phase vector of arrowheads of the same respective color, where length is inversely proportional to the phase variance. Circle represents 24 hours, where the grey is 12 hours of dark, and the yellow is 12 hours of light.
(TIF)
Mice experience behavioral after-effects in period dependent on genotype and the order in which they gained access to a wheel. a | Interaction between the order in which mice of all genotypes received access to a wheel and their resultant free-running period during the time the wheel was open or locked. b | Interaction between Pet-1 genotype and the order in which mice received access to a wheel leads to changes in free-running period. c | While not a statistically significant interaction, this plot illustrates the open/lock components that make up B. Purple icons represent mice that had the wheel open first, then locked; Yellow icons represent mice that had the wheel locked first, then opened. Projection of arrows indicates wheel order-effect. Significance is ascribed at p≤0.05.
(TIF)
Rhythmic power analyses in ex vivo Per1 ::LUC mouse SCN demonstrates an interaction between Pet-1 genotype and sex.
(TIF)
Descriptive statistics demonstrating interactions between wheel order, wheel access and Pet-1 genotype, as well as between Pet-1 genotype and sex. Significance is ascribed at p ≤ 0.05.
(TIF)
Acknowledgments
The authors would like to thank K.L. Gamble and A.J. Ciarleglio for statistical guidance, and R.M. Penney IV for husbandry.
Funding Statement
This work was supported by NIMH P50 MH096972 (R. Blakely, ESD and DGM), and by NIMH R01 MH062723 (ESD). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Ciarleglio CM, Resuehr HE, McMahon DG (2011) Interactions of the serotonin and circadian systems: nature and nurture in rhythms and blues. Neuroscience 197: 8–16. [DOI] [PubMed] [Google Scholar]
- 2. Deneris ES, Wyler SC (2012) Serotonergic transcriptional networks and potential importance to mental health. Nature neuroscience 15: 519–527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Reppert SM, Weaver DR (2002) Coordination of circadian timing in mammals. Nature 418: 935–941. [DOI] [PubMed] [Google Scholar]
- 4. Welsh DK, Logothetis DE, Meister M, Reppert SM (1995) Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms. Neuron 14: 697–706. [DOI] [PubMed] [Google Scholar]
- 5. Lowrey PL, Takahashi JS (2011) Genetics of circadian rhythms in Mammalian model organisms. Advances in genetics 74: 175–230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Malhi GS, Kuiper S (2013) Chronobiology of mood disorders. Acta psychiatrica Scandinavica Supplementum: 2–15. [DOI] [PubMed]
- 7. Glickman G (2010) Circadian rhythms and sleep in children with autism. Neurosci Biobehav Rev 34: 755–768. [DOI] [PubMed] [Google Scholar]
- 8. Mrosovsky N (1996) Locomotor activity and non-photic influences on circadian clocks. Biol Rev Camb Philos Soc 71: 343–372. [DOI] [PubMed] [Google Scholar]
- 9. Antle MC, Mistlberger RE (2000) Circadian clock resetting by sleep deprivation without exercise in the Syrian hamster. J Neurosci 20: 9326–9332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Pittendrigh C, Daan S (1976) A Functional Analysis of Circadian Pacemakers in Nocturnal Rodents: V. Pacemaker Structure: A Clock for All Seasons. J Comp Physiol A 106: 333–355. [Google Scholar]
- 11. Edgar DM, Martin CE, Dement WC (1991) Activity feedback to the mammalian circadian pacemaker: influence on observed measures of rhythm period length. J Biol Rhythms 6: 185–199. [DOI] [PubMed] [Google Scholar]
- 12. Mistlberger RE, Bossert JM, Holmes MM, Marchant EG (1998) Serotonin and feedback effects of behavioral activity on circadian rhythms in mice. Behav Brain Res 96: 93–99. [DOI] [PubMed] [Google Scholar]
- 13. Prosser RA, Dean RR, Edgar DM, Heller HC, Miller JD (1993) Serotonin and the mammalian circadian system: I. In vitro phase shifts by serotonergic agonists and antagonists. J Biol Rhythms 8: 1–16. [DOI] [PubMed] [Google Scholar]
- 14. Edgar DM, Miller JD, Prosser RA, Dean RR, Dement WC (1993) Serotonin and the mammalian circadian system: II. Phase-shifting rat behavioral rhythms with serotonergic agonists. J Biol Rhythms 8: 17–31. [DOI] [PubMed] [Google Scholar]
- 15. Smith BN, Sollars PJ, Dudek FE, Pickard GE (2001) Serotonergic modulation of retinal input to the mouse suprachiasmatic nucleus mediated by 5-HT1B and 5-HT7 receptors. J Biol Rhythms 16: 25–38. [DOI] [PubMed] [Google Scholar]
- 16. Glass JD, Grossman GH, Farnbauch L, DiNardo L (2003) Midbrain raphe modulation of nonphotic circadian clock resetting and 5-HT release in the mammalian suprachiasmatic nucleus. J Neurosci 23: 7451–7460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Morin LP, Allen CN (2006) The circadian visual system, 2005. Brain Res Rev 51: 1–60. [DOI] [PubMed] [Google Scholar]
- 18. Deurveilher S, Semba K (2005) Indirect projections from the suprachiasmatic nucleus to major arousal-promoting cell groups in rat: implications for the circadian control of behavioural state. Neuroscience 130: 165–183. [DOI] [PubMed] [Google Scholar]
- 19. Mintz EM, Gillespie CF, Marvel CL, Huhman KL, Albers HE (1997) Serotonergic regulation of circadian rhythms in Syrian hamsters. Neuroscience 79: 563–569. [DOI] [PubMed] [Google Scholar]
- 20. Manrique C, Francois-Bellan AM, Segu L, Becquet D, Hery M, et al. (1994) Impairment of serotoninergic transmission is followed by adaptive changes in 5HT1B binding sites in the rat suprachiasmatic nucleus. Brain research 663: 93–100. [DOI] [PubMed] [Google Scholar]
- 21. Manrique C, Segu L, Hery F, Hery M, Faudon M, et al. (1993) Increase of central 5-HT1B binding sites following 5,7-dihydroxytryptamine axotomy in the adult rat. Brain research 623: 345–348. [DOI] [PubMed] [Google Scholar]
- 22. Kiss J, Leranth C, Halasz B (1984) Serotoninergic endings on VIP-neurons in the suprachiasmatic nucleus and on ACTH-neurons in the arcuate nucleus of the rat hypothalamus. A combination of high resolution autoradiography and electron microscopic immunocytochemistry. Neuroscience letters 44: 119–124. [DOI] [PubMed] [Google Scholar]
- 23. Bosler O (1989) Ultrastructural relationships of serotonin and GABA terminals in the rat suprachiasmatic nucleus. Evidence for a close interconnection between the two afferent systems. Journal of neurocytology 18: 105–113. [DOI] [PubMed] [Google Scholar]
- 24. Bosler O, Beaudet A (1985) VIP neurons as prime synaptic targets for serotonin afferents in rat suprachiasmatic nucleus: a combined radioautographic and immunocytochemical study. J Neurocytol 14: 749–763. [DOI] [PubMed] [Google Scholar]
- 25. Lovenberg TW, Baron BM, de Lecea L, Miller JD, Prosser RA, et al. (1993) A novel adenylyl cyclase-activating serotonin receptor (5-HT7) implicated in the regulation of mammalian circadian rhythms. Neuron 11: 449–458. [DOI] [PubMed] [Google Scholar]
- 26. Amir S, Robinson B, Ratovitski T, Rea MA, Stewart J, et al. (1998) A role for serotonin in the circadian system revealed by the distribution of serotonin transporter and light-induced Fos immunoreactivity in the suprachiasmatic nucleus and intergeniculate leaflet. Neuroscience 84: 1059–1073. [DOI] [PubMed] [Google Scholar]
- 27. Pickard GE, Smith BN, Belenky M, Rea MA, Dudek FE, et al. (1999) 5-HT1B receptor-mediated presynaptic inhibition of retinal input to the suprachiasmatic nucleus. J Neurosci 19: 4034–4045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Quintero JE, McMahon DG (1999) Serotonin modulates glutamate responses in isolated suprachiasmatic nucleus neurons. Journal of neurophysiology 82: 533–539. [DOI] [PubMed] [Google Scholar]
- 29. Prosser RA (2003) Serotonin phase-shifts the mouse suprachiasmatic circadian clock in vitro. Brain Res 966: 110–115. [DOI] [PubMed] [Google Scholar]
- 30. Morin LP (1999) Serotonin and the regulation of mammalian circadian rhythmicity. Ann Med 31: 12–33. [DOI] [PubMed] [Google Scholar]
- 31. Meyer-Bernstein EL, Morin LP (1996) Differential serotonergic innervation of the suprachiasmatic nucleus and the intergeniculate leaflet and its role in circadian rhythm modulation. J Neurosci 16: 2097–2111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Marchant EG, Watson NV, Mistlberger RE (1997) Both neuropeptide Y and serotonin are necessary for entrainment of circadian rhythms in mice by daily treadmill running schedules. J Neurosci 17: 7974–7987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Mistlberger RE, Antle MC (1998) Behavioral inhibition of light-induced circadian phase resetting is phase and serotonin dependent. Brain Res 786: 31–38. [DOI] [PubMed] [Google Scholar]
- 34. Kawakami F, Okamura H, Tamada Y, Nakajima T, Ibata Y (1995) Changes in vasoactive intestinal peptide mRNA levels in the rat suprachiasmatic nucleus following p-chlorophenylalanine (PCPA) treatment under light/dark conditions. Neuroscience letters 200: 171–174. [DOI] [PubMed] [Google Scholar]
- 35. Belenky MA, Pickard GE (2001) Subcellular distribution of 5-HT(1B) and 5-HT(7) receptors in the mouse suprachiasmatic nucleus. J Comp Neurol 432: 371–388. [DOI] [PubMed] [Google Scholar]
- 36. Wallace JA, Lauder JM (1983) Development of the serotonergic system in the rat embryo: an immunocytochemical study. Brain research bulletin 10: 459–479. [DOI] [PubMed] [Google Scholar]
- 37. Levitt P, Rakic P (1982) The time of genesis, embryonic origin and differentiation of the brain stem monoamine neurons in the rhesus monkey. Brain research 256: 35–57. [DOI] [PubMed] [Google Scholar]
- 38. Hendricks TJ, Fyodorov DV, Wegman LJ, Lelutiu NB, Pehek EA, et al. (2003) Pet-1 ETS gene plays a critical role in 5-HT neuron development and is required for normal anxiety-like and aggressive behavior. Neuron 37: 233–247. [DOI] [PubMed] [Google Scholar]
- 39. Paulus EV, Mintz EM (2012) Developmental disruption of the serotonin system alters circadian rhythms. Physiology & behavior 105: 257–263. [DOI] [PubMed] [Google Scholar]
- 40. Kiyasova V, Fernandez SP, Laine J, Stankovski L, Muzerelle A, et al. (2011) A genetically defined morphologically and functionally unique subset of 5-HT neurons in the mouse raphe nuclei. The Journal of neuroscience: the official journal of the Society for Neuroscience 31: 2756–2768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Paulus EV, Mintz EM (2013) Photic and nonphotic responses of the circadian clock in serotonin-deficient Pet-1 knockout mice. Chronobiology international 30: 1251–1260. [DOI] [PubMed] [Google Scholar]
- 42. Herzog ED, Aton SJ, Numano R, Sakaki Y, Tei H (2004) Temporal precision in the mammalian circadian system: a reliable clock from less reliable neurons. J Biol Rhythms 19: 35–46. [DOI] [PubMed] [Google Scholar]
- 43. Lerch-Haner JK, Frierson D, Crawford LK, Beck SG, Deneris ES (2008) Serotonergic transcriptional programming determines maternal behavior and offspring survival. Nature neuroscience 11: 1001–1003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Colwell CS, Michel S, Itri J, Rodriguez W, Tam J, et al. (2003) Disrupted circadian rhythms in VIP- and PHI-deficient mice. Am J Physiol Regul Integr Comp Physiol 285: R939–949. [DOI] [PubMed] [Google Scholar]
- 45. Ohta H, Mitchell AC, McMahon DG (2006) Constant light disrupts the developing mouse biological clock. Pediatr Res 60: 304–308. [DOI] [PubMed] [Google Scholar]
- 46.Sokal RR, Rohlf FJ (1995) Biometry: the principles and practice of statistics in biological research. New York: W.H. Freeman. xix, 887 p.
- 47. Edgar DM, Dement WC (1991) Regularly scheduled voluntary exercise synchronizes the mouse circadian clock. Am J Physiol 261: R928–933. [DOI] [PubMed] [Google Scholar]
- 48. Edgar DM, Kilduff TS, Martin CE, Dement WC (1991) Influence of running wheel activity on free-running sleep/wake and drinking circadian rhythms in mice. Physiol Behav 50: 373–378. [DOI] [PubMed] [Google Scholar]
- 49. Edgar DM, Reid MS, Dement WC (1997) Serotonergic afferents mediate activity-dependent entrainment of the mouse circadian clock. Am J Physiol 273: R265–269. [DOI] [PubMed] [Google Scholar]
- 50. Ciarleglio CM, Axley JC, Strauss BR, Gamble KL, McMahon DG (2011) Perinatal photoperiod imprints the circadian clock. Nat Neurosci 14: 25–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Morin LP, Fitzgerald KM, Rusak B, Zucker I (1977) Circadian organization and neural mediation of hamster reproductive rhythms. Psychoneuroendocrinology 2: 73–98. [DOI] [PubMed] [Google Scholar]
- 52. Morin LP, Fitzgerald KM, Zucker I (1977) Estradiol shortens the period of hamster circadian rhythms. Science 196: 305–307. [DOI] [PubMed] [Google Scholar]
- 53. Yamazaki S, Alones V, Menaker M (2002) Interaction of the retina with suprachiasmatic pacemakers in the control of circadian behavior. J Biol Rhythms 17: 315–329. [DOI] [PubMed] [Google Scholar]
- 54. Aton SJ, Block GD, Tei H, Yamazaki S, Herzog ED (2004) Plasticity of circadian behavior and the suprachiasmatic nucleus following exposure to non-24-hour light cycles. J Biol Rhythms 19: 198–207. [DOI] [PubMed] [Google Scholar]
- 55. Pendergast JS, Friday RC, Yamazaki S (2009) Endogenous rhythms in Period1 mutant suprachiasmatic nuclei in vitro do not represent circadian behavior. J Neurosci 29: 14681–14686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Molyneux PC, Dahlgren MK, Harrington ME (2008) Circadian entrainment aftereffects in suprachiasmatic nuclei and peripheral tissues in vitro. Brain Res 1228: 127–134. [DOI] [PubMed] [Google Scholar]
- 57. Gamble KL, Allen GC, Zhou T, McMahon DG (2007) Gastrin-releasing peptide mediates light-like resetting of the suprachiasmatic nucleus circadian pacemaker through cAMP response element-binding protein and Per1 activation. J Neurosci 27: 12078–12087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Kuhlman SJ, Quintero JE, McMahon DG (2000) GFP fluorescence reports Period 1 circadian gene regulation in the mammalian biological clock. Neuroreport 11: 1479–1482. [PubMed] [Google Scholar]
- 59. Quintero JE, Kuhlman SJ, McMahon DG (2003) The biological clock nucleus: a multiphasic oscillator network regulated by light. J Neurosci 23: 8070–8076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Maywood ES, Reddy AB, Wong GK, O'Neill JS, O'Brien JA, et al. (2006) Synchronization and maintenance of timekeeping in suprachiasmatic circadian clock cells by neuropeptidergic signaling. Curr Biol 16: 599–605. [DOI] [PubMed] [Google Scholar]
- 61. Yoo SH, Yamazaki S, Lowrey PL, Shimomura K, Ko CH, et al. (2004) PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc Natl Acad Sci U S A 101: 5339–5346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Mong JA, Baker FC, Mahoney MM, Paul KN, Schwartz MD, et al. (2011) Sleep, rhythms, and the endocrine brain: influence of sex and gonadal hormones. The Journal of neuroscience: the official journal of the Society for Neuroscience 31: 16107–16116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Kriegsfeld LJ, Silver R (2006) The regulation of neuroendocrine function: Timing is everything. Horm Behav 49: 557–574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Karatsoreos IN, Silver R (2007) Minireview: The neuroendocrinology of the suprachiasmatic nucleus as a conductor of body time in mammals. Endocrinology 148: 5640–5647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Abizaid A, Mezei G, Thanarajasingam G, Horvath TL (2005) Estrogen enhances light-induced activation of dorsal raphe serotonergic neurons. Eur J Neurosci 21: 1536–1546. [DOI] [PubMed] [Google Scholar]
- 66. Karatsoreos IN, Wang A, Sasanian J, Silver R (2007) A role for androgens in regulating circadian behavior and the suprachiasmatic nucleus. Endocrinology 148: 5487–5495. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Rayleigh plots of activity onset and offset as monitored by IR ( top ) and wheel-running ( bottom ), and when the wheel was open ( left ) and locked ( right ). Blue arrowheads represent Pet-1 +/+ mice; orange arrowheads represent Pet-1 +/− mice; black arrowheads represent Pet-1 +/+ mice; arrows represent the mean phase vector of arrowheads of the same respective color, where length is inversely proportional to the phase variance. Circle represents 24 hours, where the grey is 12 hours of dark, and the yellow is 12 hours of light.
(TIF)
Mice experience behavioral after-effects in period dependent on genotype and the order in which they gained access to a wheel. a | Interaction between the order in which mice of all genotypes received access to a wheel and their resultant free-running period during the time the wheel was open or locked. b | Interaction between Pet-1 genotype and the order in which mice received access to a wheel leads to changes in free-running period. c | While not a statistically significant interaction, this plot illustrates the open/lock components that make up B. Purple icons represent mice that had the wheel open first, then locked; Yellow icons represent mice that had the wheel locked first, then opened. Projection of arrows indicates wheel order-effect. Significance is ascribed at p≤0.05.
(TIF)
Rhythmic power analyses in ex vivo Per1 ::LUC mouse SCN demonstrates an interaction between Pet-1 genotype and sex.
(TIF)
Descriptive statistics demonstrating interactions between wheel order, wheel access and Pet-1 genotype, as well as between Pet-1 genotype and sex. Significance is ascribed at p ≤ 0.05.
(TIF)