

Abstract
Introduction
Probiotic use to prevent nosocomial gastrointestinal and potentially respiratory tract infections in critical care has shown great promise in recent clinical trials of adult and pediatric patients. Despite well-documented benefits of probiotic use in intestinal disorders, the potential for probiotic treatment to reduce lung injury following infection and shock has not been well explored.
Objective
Evaluate if Lactobacillus rhamnosus GG (LGG) or Bifidobacterium longum (BL) treatment in a weanling mouse model of cecal ligation and puncture (CLP) peritonitis will protect against lung injury.
Methods
3 week-old FVB/N mice were orally gavaged with 200 µl of either LGG, BL or sterile water (vehicle) immediately prior to CLP. Mice were euthanized at 24 h. Lung injury was evaluated via histology and lung neutrophil infiltration was evaluated by myeloperoxidase (MPO) staining. mRNA levels of IL-6, TNF-α, MyD88, TLR-4, TLR-2, NFΚB (p50/p105) and Cox-2 in the lung analyzed via real-time PCR. TNF-α and IL-6 in lung was analyzed via ELISA.
Results
LGG and BL treatment significantly improved lung injury following experimental infection and sepsis and lung neutrophil infiltration was significantly lower than in untreated septic mice. Lung mRNA and protein levels of IL-6 and TNF-α and gene expression of Cox-2 were also significantly reduced in mice receiving LGG or BL treatment. Gene expression of TLR-2, MyD88 and NFΚB (p50/p105) was significantly increased in septic mice compared to shams and decreased in the lung of mice receiving LGG or BL while TLR-4 levels remained unchanged.
Conclusions
Treatment with LGG and BL can reduce lung injury following experimental infection and sepsis and is associated with reduced lung inflammatory cell infiltrate and decreased markers of lung inflammatory response. Probiotic therapy may be a promising intervention to improve clinical lung injury following systemic infection and sepsis.
Introduction
Sepsis is a leading cause of death in infants and children despite the advances in medical and ICU care. Over 42,000 cases of severe sepsis are reported each year in the United States alone and millions are thought to occur worldwide [1]. Over 1 million deaths worldwide are associated with sepsis within the neonatal population [2], [3]. Low birth weight infants are particularly at risk, where the mortality is reported to be ∼50% [4]. Further the neonates who survive sepsis and septic shock continue to face substantial long term adverse effects [5]. Pediatric patients diagnosed with pneumonia or sepsis are also susceptible to acute lung injury or acute respiratory distress syndrome leading to a mortality rate of ∼25% [6], [7].
Critical illness and ICU care (broad spectrum antibiotics, poor nutrition deliver etc) creates a hostile environment in the gut and alter the microflora tilting the balance to favor pathogens [8]. Probiotics are living nonpathogenic bacteria colonizing intestine and providing benefit to the host with the potential to normalize the altered intestinal flora [9]. The use of probiotics in prevention of nosocomial gastrointestinal and respiratory tract infections in critical care has increased over the last few years and results from a growing number of randomized controlled trials within the adult and pediatric populations suggest their use as a promising treatment [10], [11], [12], [13]. The need for alternative, non-antimicrobial interventions for prevention of infection in an age of increasing antimicrobial resistance also make probiotics a promising strategy. Specifically, lactobacilli and bifidobacteria alone or in combination are the most frequently used strains in the treatment of various gastrointestinal disorders [14], [15], [16] or as therapy for different clinical conditions including antibiotic associated diarrhea [17], acute pancreatitis [18], ventilator associated pneumonia [11], [12], [19], sepsis and postoperative infections [20], [21].
Although probiotics are showing promise as an effective therapy in a growing number of illnesses, the mechanisms of their action are complex and still elusive [22]. Based on the results from several in vivo and in vitro studies, probiotics are able to decrease apoptosis in intestinal epithelial cells [23], [24], [25], [26], improve intestinal integrity [27], [28], [29], [30], prevent bacterial translocation [30], [31], reduce the overgrowth of pathogenic bacteria and suppress cytokine production [32], [33], [34], [35].
Despite the benefits of probiotic use in intestinal disorders, the effects of probiotic treatment to protect against lung injury following infection and sepsis are not well understood. We have recently shown the benefits of Lactobacillus rhamnosus GG (LGG) and Bifidobacterium longum (BL) on improved survival and intestinal homeostasis in weanling mouse model of cecal ligation and puncture (CLP) [36]. CLP is an experimental model of shock that mimics the pathology of sepsis occurring in the ICU patients [37]. Toll like receptors (TLRs) are pattern recognition receptors involved in the initial steps of signaling pathway leading to multiple organ failure in sepsis. TLRs bind to cell-wall components which activates nuclear factor (NF)-ΚB/IΚB system resulting in release of pro-inflammatory cytokines [38]. In addition to cytokines, pathogens activating TLRs were also reported to induce Cox-2 expression [39], [40].
In this study we hypothesized that LGG and BL will also have a protective effect against lung injury and will decrease the inflammatory response in the lungs potentially via the TLR/Myd88 pathway.
Methods
Probiotic treatment and septic peritonitis model
The animal protocol used in these studies was approved by the Institutional Animal Care and Use Committee of the University of Colorado Anschutz Medical Campus. Briefly, 3 weeks old FVB/N mice were orally gavaged with 200 µl of either LGG (1×109 CFU/ml), BL (1×107 CFU/ml), or sterile water (vehicle) immediately prior to initiation of the cecal ligation and puncture (CLP) procedure [41]. Briefly, a small midline abdominal incision was made, the cecum was ligated just distal to the ileocecal valve, and was then punctured twice with a 23-gauge needle. The cecum was squeezed to extrude a small amount of stool, replaced in the abdomen, and the peritoneum and skin were closed in layers. Sham mice were treated identically except the cecum was neither ligated nor punctured. All mice received 1.0 ml normal saline subcutaneously after the surgery to compensate for fluid loss. Mice received a single dose of probiotics prior to tissue collection. Animals were euthanized at 24 h.
Lactobacillus rhamnosus GG and Bifidobacterium longum culture
LGG (ATCC, Manassas, VA) was incubated in MRS broth (BD, Sparks, MD) for 24 hours at 37°C and 5% CO2. BL (ATCC, Manassas, VA) was cultured in Trypticase soy broth (BD, Sparks, MD) for 72 hours in an anaerobic chamber at 37°C. A600 was measured to determine the number of colony forming units (CFU) per 1 ml. BL and LGG were pelleted from the broth (10,000 rpm; 10 min) and resuspended in distilled water.
Immunohistology
Lung tissue was collected from each animal at 24 h and fixed overnight in 10% formalin, paraffin-embedded, and sectioned at 4–6 µm. Serial sections were stained with hematoxylin-eosin (H&E) and evaluated for severity of lung injury by blinded evaluator using a grading scale from 0 (no abnormality) to 4 (severe lung injury) as described previously [42].
Neutrophil infiltration into the lungs was evaluated by staining for myeloperoxidase (MPO). After deparaffinization and rehydration, sections were blocked with 1.5% rabbit serum (Vector Laboratories, Burlingame, CA) in phosphate-buffered saline for 30 min, then incubated with goat polyclonal MPO (1∶50; R&D Systems, Minneapolis, MN) antibody for 1 hour, washed with phosphate-buffered saline, and incubated with rabbit anti-goat biotinylated secondary antibody (Vector Laboratories) for 30 min. Vectastain Elite ABC reagent (Vector Laboratories) was then applied, followed by diaminobenzidine as substrate. Sections were counterstained with hematoxylin, dehydrated and cover-slipped. MPO positive cells were quantified in 10 random high-power fields per section. All counting was performed by a blinded evaluator.
RNA Preparation, RT, and Real-Time PCR
Total RNA was isolated from lung tissue (snap frozen in liquid N2, collected at 24 h) using the RNeasy Plus Mini Kit (Qiagen, Santa Clarita, CA) as described in the manufacturer's protocol. RNA concentrations were quantified at 260 nm, and the purity and integrity were determined using a NanoDrop. RT and real-time PCR assays were performed to quantify steady-state mRNA levels of IL-6, TNF-α, MyD88, TLR-4, TLR-2, NFΚB (p50/p105) and Cox-2. cDNA was synthesized from 0.2 µg of total RNA. Predeveloped TaqMan primers and probes (Applied Biosystems) were used for detection. Reporter dye emission was detected by an automated sequence detector combined with ABI Prism 7300 Real Time PCR System (Applied Biosystems). Real-time PCR quantification was performed with TaqMan GAPDH controls and relative mRNA expression calculated using the 2−ΔΔCT method [43].
IL-6 and TNF-α protein analysis in the lung tissue
Lung tissue was harvested and frozen immediately in liquid nitrogen. Samples were homogenized with a hand-held homogenizer in a 5× volume of ice-cold homogenization buffer (Tris HCl, 50 mm; pH, 7.4; NaCl, 100 mm; EDTA, 10 mm; Triton X-100, 0.5%) with added protease inhibitors (Roche Diagnostics, Mannheim, Germany). The homogenates were centrifuged at 10,000 rpm for 5 min at 4°C and the supernatant was collected. Total protein concentration was quantified using the Bradford protein assay. Enzyme-linked immunosorbent assay (ELISA) (R&D Systems, Mineapolis, MN) was used to determine the concentrations of TNF-α and IL-6 in lung tissue homogenates according to the manufacturer's instructions.
Statistics
Comparisons were performed with t test analysis (unpaired, two-tailed). To analyze the bacterial culture results, 2-tailed NPar, Mann-Whitney Test was used. No measurements or animals were lost to observation or missing in the analysis. Data were analyzed using Prism 4.0 (GraphPad Software, San Diego, CA) and reported as means ± SE. A p value≤0.05 was considered to be statistically significant.
Results
Probiotics improve lung injury and decrease the neutrophil infiltration during sepsis
We have previously shown in this model that probiotic treatment with LGG or BL can improve survival following CLP [36]. In this study we hypothesized this may be associated with or be related to reduction in lung injury. Thus, the effect of probiotic treatment on lung pathology was assessed. Sepsis led to marked histological injury 24 hours after CLP surgery in septic animals. This injury was significantly improved in septic animals treated with LGG or BL (Figure 1A, B). Sepsis-mediated lung injury was associated with a significantly higher number of infiltrating neutrophils, represented by the number of MPO positive cells in the septic animals when compared to shams (P<0.0001). Treatment with LGG and BL normalized (P<0.0001) the number of MPO positive cells in the lungs to that observed in sham mice (Figure 2A, B).
Figure 1. Probiotics improve lung pathology 24 hours post CLP.

(A) The severity of pneumonia (from 0: no abnormality to 4: severe lung injury) was significantly reduced in the lungs of mice treated with LGG and BL (P<0.05). (B) Representative H&E stained sections of lung are shown. Original magnification ×100. Shams n = 4 per group, Septic, Septic+LGG, Septic+BL n = 5–8 per group.
Figure 2. Probiotics decrease neutrophil infiltration in the lung 24 hours post CLP.

(A) Number of MPO positive cells in the lungs of septic mice were significantly increased compared to shams. LGG or BL treatment normalized these levels (P<0.0001). (B) Representative MPO stained sections of lung are shown. Original magnification ×200. Shams n = 4 per group, Septic, Septic+LGG, Septic+BL n = 6 per group.
Probiotics attenuate proinflammatory cytokine release in the lung after sepsis
To determine the effect of LGG and BL treatment on pro-inflammatory cytokine release in lungs after CLP-induced polymicrobial sepsis, mRNA levels of IL-6 and TNF-α were analyzed by Real-Time PCR and protein levels measured by ELISA. Gene expressions of IL-6 and TNF-α (Figure 3A, B) were significantly increased in lungs of septic animals (P<0.05) and normalized to sham levels in LGG or BL treated mice. Protein levels of IL-6 (P<0.01) (Figure 3C) and TNF-α (P<0.05) (Figure 3D) were also markedly elevated in septic mice and attenuated to sham levels in mice treated with either probiotic strain.
Figure 3. Probiotics attenuate proinflammatory cytokine release in lung 24 hours post CLP.

Reverse transcription and real-time PCR assays were performed to quantify steady-state mRNA levels of pro-inflammatory cytokines. (A) IL-6 and (B) TNF-α were significantly increased in the lung of septic animals compare to shams (P<0.05). LGG or BL treatment normalized these levels to shams (P<0.05). Shams n = 4 per group; Septic, Septic+LGG, Septic+BL n = 5 per group. Data are expressed as the mean ± SE. Enzyme-linked immunosorbent assay (ELISA) was used to determine the protein concentrations of IL-6 and TNF-α in the lung. (C) IL-6 and (D) TNF-α were significantly elevated in the lung of septic mice compared to shams (P<0.05). Treatment with LGG or BL prior to CLP led to significantly reduced (P<0.05) levels of both cytokines compared to untreated septic mice. Shams n = 3 per group; Septic, Septic+LGG, Septic+BL n = 4 per group. Data are expressed as the mean ± SE.
Probiotics decrease Cox-2 expression and regulate toll-like receptor (TLR) pathway in the lung during sepsis
Cox-2 is rapidly induced in response to cytokines and is elevated at sites of inflammation. Gene expression of Cox-2 was significantly increased in lungs of septic mice (P<0.05) and treatment with LGG or BL significantly decreased (P<0.05) Cox-2 levels to those observed in the lung of sham animals (Figure 4).
Figure 4. Probiotics downregulate Cox-2 expression in the lung 24 hours post CLP.
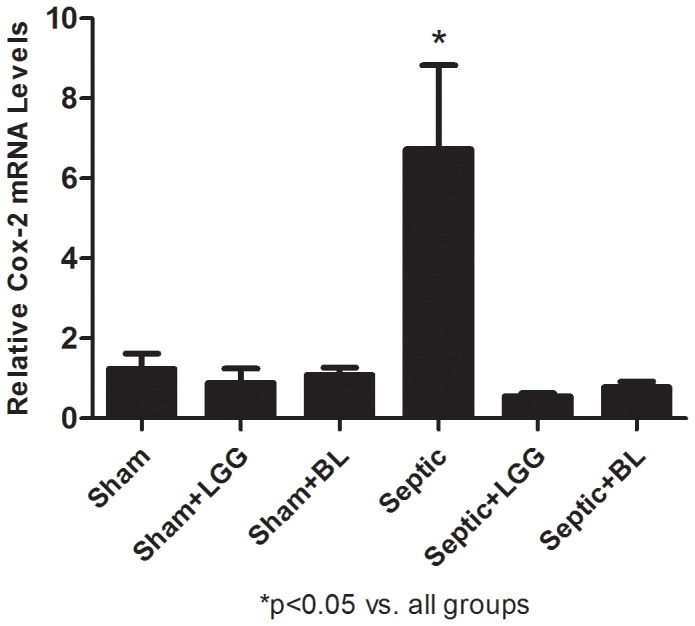
Reverse transcription and real-time PCR assays were performed to quantify steady-state mRNA levels of Cox-2. Cox-2 was significantly elevated in the septic group compared to sham groups (P<0.05). Treatment with LGG or BL significantly reduced mRNA levels of Cox-2 compared to untreated septic mice (P<0.05). Shams n = 4 per group; Septic, Septic+LGG, Septic+BL n = 4–5 per group. Data are expressed as the mean ± SE.
TLRs signal via the MyD88 pathway that includes the NFΚB transcriptional factor, which is a key activator of the cytokines involved in the innate immunity response. MyD88 has an important role in early recruitment of inflammatory cells and in the control of bacterial infection [44]. Gene expression of TLR-2, TLR-4, MyD88 and NFΚB (p50/p105) was analyzed by Real-Time PCR. There was significant increase of TLR-2 MyD88 and NFΚB (p50/p105) in the lungs of septic mice (P<0.05) (Figure 5A, C, D). LGG or BL treatment normalized the levels of TLR-2 and MyD88 to those in shams (Figure 5A, C). NFΚB (p50/p105) was significantly decreased in the lung of LGG treated mice (P<0.05). The levels in BL treated mice were also decreased but did not reach statistical significance (Figure 5D). TLR-4 remained unchanged regardless of treatment (Figure 5B).
Figure 5. Probiotics regulate Toll-like receptor (TLR) pathway in the lung 24 hours post CLP.

Reverse transcription and real-time PCR assays were performed to quantify steady-state mRNA levels of TLR-2, TLR-4, MyD88 and NFΚB (p50/p105). (A) TLR-2 and (C) MyD88 were significantly upregulated in the septic group compared to shams (P<0.05) and significantly downregulated in the lungs of LGG and BL treated septic mice compared to untreated septic mice. (B) mRNA levels of TLR-4 remained unchanged in all groups. (D) NFΚB (p50/p105) was significantly upregulated in the septic group compared to shams (P<0.05) and significantly downregulated in the lungs of LGG treated septic mice compared to untreated septic mice. The levels in BL treated mice were decreased but did not reach statistical significance. Shams n = 5 per group; Septic, Septic+LGG, Septic+BL n = 5–7 per group. Data are expressed as the mean ± SE.
Discussion
This work demonstrates two probiotic strains, Lactobacillus rhamnosus GG and Bifidobacterium longum, can reduce lung injury and attenuate the inflammatory response in the lungs of weanling mice subjected to CLP.
Not surprisingly, the gut has been identified as an origin and promoter of nosocomial sepsis and multiorgan failure in the critically ill, the major determinant of ICU outcome [45].Critical illness and ICU-based therapies, such as vasopressors and broad spectrum antibiotics, create a hostile environment in the gut and alter the microflora favoring the growth of pathogens. This is in part due to the loss of key beneficial lactic acid bacteria [46] otherwise called probiotics that can inhibit the overgrowth of pathogens by production of bacteriocins, hydrogen peroxide, organic acids, ammonia and by increased competition for adhesion sites on intestinal epithelia [47], [48]. Also, a number of bioactive factors secreted by probiotics, mainly by LGG, have been identified and their effects studied in intestinal injury as well as airway inflammation models [49], [50]. Soluble protein p40 derived from LGG as published by Polk et al preserves barrier function and reduces apoptosis in the colon epithelium in an EGF receptor-dependent manner [51]. A study performed in healthy adults suggests how three different lactobacilli induce differential gene-regulatory networks and pathways in the human mucosa, showing that mucosal responses to LGG involve would healing, IFN response and ion homeostasis [52]. A recent review article provides detailed information on several probiotic strains and their ability to stimulate the immune system including activation of macrophages, natural killer cells, T-lymphocytes and release of cytokines in strain specific, dose dependent manner [53]. Several randomized controlled trials within adult and pediatric populations suggest the use of probiotics as a promising therapy for nosocomial gastrointestinal and respiratory tract infections [10], [11], [12], [13] but there are still many questions to be answered about their mechanisms of action.
From current clinical studies of probiotic therapy, it appears that timing of probiotic administration may be important in their effectiveness with administration early in critical illness potentially being important. [27], [54], [55], [56]. In our mouse model of sepsis, the animals were given LGG or BL immediately before the surgery to better reflect the common clinical setting where a patient presenting with peritonitis could be treated at the time of surgery to attempt to prevent future hospital acquired infections and acute lung injury. Our recently published data describe significant improvement of several outcomes including survival, bacteremia, systemic inflammatory response and intestinal homeostasis with administration of these probiotic strains in this immediate “surgical” timeframe. [36].
The pathophysiology of septic shock syndrome is characterized by hyperactive and dysregulated endogenous inflammatory mediators including cytokines such as IL-6, TNF-α, IL-1β, IL-12 and interferon γ [57], [58]. It has been shown that early attenuation of transcription factor NFΚB activation and cytokine message expression correlates with improved outcome in polymicrobial sepsis [59]. Controlling inflammatory mediated injury to distant organs is a key goal in sepsis to prevent the multiple organ dysfunction syndrome (MODS) which carries quite a high mortality. This is often observed in generalized peritonitis (as studied in our model), which accompanies surgical conditions such as gastrointestinal perforation [58]. Clinical and experimental data support an important role of the lung during the initial stages of the multiple organ dysfunction syndrome (MODS) [60]. The release of pro-inflammatory mediators can cause acute lung injury [61] and it has been reported that levels of pro-inflammatory cytokines such as IL-6 and TNF-α are significantly elevated in the lungs after CLP-induced peritonitis [59], [62]. There are several publications reporting the protective effect of different probiotic strains against bacterial infection. A study done in the rat CLP peritonitis model demostrated a decrease of TNF-α and IL-1β in lungs of animals receiving a prolonged three week pre-treatment with probiotics and overall reduction of acute lung injury was also observed [35]. Racedo at al. used a mouse model of Streptococcus pneumoniae infection to evaluate the effect of L. casei and found that two days of pre-treatment could beneficially regulate the TNF-α and IL-10 balance, allowing a more effective immune response against infection and modulation the inflammatory response. This was associated with less damage to the lung in this model [63]. In our unique immediate pre-treatment model, we found significantly increased mRNA and protein levels of pro-inflammatory cytokines TNF-α, IL-6 in the lungs of septic mice. Treatment at the time of onset of peritonitis (rather than a prolonged pre-treatment period) with either Lactobacillus rhamnosus GG or Bifidobacterium longum normalized these cytokine levels to those seen in shams indicating the anti-inflammatory effect of both probiotic strains possibly contributing to better overall outcome.
In general, Cox-2 is not expressed in healthy tissues but is rapidly induced in response to cytokines and is elevated at sites of inflammation and injury [64] and is involved in pathogenesis of sepsis [65]. In mouse CLP model, Cox-2 expression was previously shown to increase in the lungs of septic mice [66], [67], [68], in addition dual inhibition of Cox-2 and 5-LOX successfully attenuated lung injury, reduced MPO activity and improved survival of these mice [69]. As shown in several in vitro and in vivo models, pathogens induce Cox-2 expression via activated Toll like receptors (TLRs) [39], [40]. TLRs play a central role in the initiation of innate immune responses and in the development of a subsequent pro-inflammatory response, which can lead to inflammation induced organ injury. TLRs are activated by specific microbial ligands leading to an association with TIR domain containing MyD88 factor which mediates a signaling cascade that activates NFΚB factor and results in upregulation of pro-inflammatory cytokines [70]. Markedly increased expression of TLR-2 and TLR-4 in monocytes [71], [72] and leukocytes [73] has been reported in septic patients. In mouse CLP peritonitis models, TLR-2 and TLR-4 expressions were significantly upregulated in hepatic and splenic macrophages [74], in the lungs and liver [75], [76] as well as in the intestine [77] when compared to sham mice. Here we demonstrate that Cox-2, TLR-2, MyD88 and NFΚB (p50/p105) were significantly higher in the lungs of septic mice compared to healthy shams and lower in the lungs of LGG and BL mice. NFΚB (p50/p105) in the lungs of BL treated septic mice showed only a decreasing trend. The expression of TLR-4 in the lungs remained unchanged among all experimental groups, similar to the observations of Williams et al. [75] in a CLP peritonitis model where TLR-4 expression increased at earlier time points but not at 24 hours. We speculate that downregulation of Cox-2 through TLR-2/TLR-4 (via MyD88) in the lungs of Lactobacillus rhamnosus GG or Bifidobacterium longum treated mice may play a protective role in attenuating inflammation induced lung injury following systemic sepsis and peritonitis.
In conclusion, probiotic therapy with LGG and BL can reduce lung injury following experimental peritonitis and sepsis and is associated with reduced lung inflammatory cell infiltrate and decreased markers of lung inflammatory response activation. Probiotic therapy may be a promising intervention to improve clinical lung injury following systemic infection and sepsis.
Funding Statement
This work was funded in part by National Institutes of Health (NIH) R01 GM078312 to P. Wischmeyer. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. No additional external funding was received for this study.
References
- 1. Watson RS, Carcillo JA, Linde-Zwirble WT, Clermont G, Lidicker J, et al. (2003) The epidemiology of severe sepsis in children in the United States. Am J Respir Crit Care Med 167: 695–701. [DOI] [PubMed] [Google Scholar]
- 2. Lukacs SL, Schoendorf KC, Schuchat A (2004) Trends in sepsis-related neonatal mortality in the United States, 1985–1998. Pediatr Infect Dis J 23: 599–603. [DOI] [PubMed] [Google Scholar]
- 3. Wynn JL, Scumpia PO, Delano MJ, O'Malley KA, Ungaro R, et al. (2007) Increased mortality and altered immunity in neonatal sepsis produced by generalized peritonitis. Shock 28: 675–683. [DOI] [PubMed] [Google Scholar]
- 4. Kermorvant-Duchemin E, Laborie S, Rabilloud M, Lapillonne A, Claris O (2008) Outcome and prognostic factors in neonates with septic shock. Pediatr Crit Care Med 9: 186–191. [DOI] [PubMed] [Google Scholar]
- 5. Adams-Chapman I, Stoll BJ (2006) Neonatal infection and long-term neurodevelopmental outcome in the preterm infant. Curr Opin Infect Dis 19: 290–297. [DOI] [PubMed] [Google Scholar]
- 6. Dahlem P, van Aalderen WM, Bos AP (2007) Pediatric acute lung injury. Paediatr Respir Rev 8: 348–362. [DOI] [PubMed] [Google Scholar]
- 7. Flori HR, Glidden DV, Rutherford GW, Matthay MA (2005) Pediatric acute lung injury: prospective evaluation of risk factors associated with mortality. Am J Respir Crit Care Med 171: 995–1001. [DOI] [PubMed] [Google Scholar]
- 8. Singhi SC, Baranwal A (2008) Probiotic use in the critically ill. Indian J Pediatr 75: 621–627. [DOI] [PubMed] [Google Scholar]
- 9. Hammerman C, Bin-Nun A, Kaplan M (2004) Germ warfare: probiotics in defense of the premature gut. Clin Perinatol 31: 489–500. [DOI] [PubMed] [Google Scholar]
- 10. Hojsak I, Abdovic S, Szajewska H, Milosevic M, Krznaric Z, et al. (2010) Lactobacillus GG in the prevention of nosocomial gastrointestinal and respiratory tract infections. Pediatrics 125: e1171–1177. [DOI] [PubMed] [Google Scholar]
- 11. Morrow LE, Kollef MH, Casale TB (2010) Probiotic prophylaxis of ventilator-associated pneumonia: a blinded, randomized, controlled trial. Am J Respir Crit Care Med 182: 1058–1064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Siempos II, Ntaidou TK, Falagas ME (2010) Impact of the administration of probiotics on the incidence of ventilator-associated pneumonia: a meta-analysis of randomized controlled trials. Crit Care Med 38: 954–962. [DOI] [PubMed] [Google Scholar]
- 13. Pitsouni E, Alexiou V, Saridakis V, Peppas G, Falagas ME (2009) Does the use of probiotics/synbiotics prevent postoperative infections in patients undergoing abdominal surgery? A meta-analysis of randomized controlled trials. Eur J Clin Pharmacol 65: 561–570. [DOI] [PubMed] [Google Scholar]
- 14. Bausserman M, Michail S (2005) The use of Lactobacillus GG in irritable bowel syndrome in children: a double-blind randomized control trial. J Pediatr 147: 197–201. [DOI] [PubMed] [Google Scholar]
- 15. Bin-Nun A, Bromiker R, Wilschanski M, Kaplan M, Rudensky B, et al. (2005) Oral probiotics prevent necrotizing enterocolitis in very low birth weight neonates. J Pediatr 147: 192–196. [DOI] [PubMed] [Google Scholar]
- 16. Drouault-Holowacz S, Bieuvelet S, Burckel A, Cazaubiel M, Dray X, et al. (2008) A double blind randomized controlled trial of a probiotic combination in 100 patients with irritable bowel syndrome. Gastroenterol Clin Biol 32: 147–152. [DOI] [PubMed] [Google Scholar]
- 17. Arvola T, Laiho K, Torkkeli S, Mykkanen H, Salminen S, et al. (1999) Prophylactic Lactobacillus GG reduces antibiotic-associated diarrhea in children with respiratory infections: a randomized study. Pediatrics 104: e64. [DOI] [PubMed] [Google Scholar]
- 18. Olah A, Belagyi T, Poto L, Romics L Jr, Bengmark S (2007) Synbiotic control of inflammation and infection in severe acute pancreatitis: a prospective, randomized, double blind study. Hepatogastroenterology 54: 590–594. [PubMed] [Google Scholar]
- 19. Schultz MJ (2010) Symbiotics as a preventive measure against ventilator-associated pneumonia. Crit Care Med 38: 1506–1507 author reply 1507. [DOI] [PubMed] [Google Scholar]
- 20. Giamarellos-Bourboulis EJ, Bengmark S, Kanellakopoulou K, Kotzampassi K (2009) Pro- and synbiotics to control inflammation and infection in patients with multiple injuries. J Trauma 67: 815–821. [DOI] [PubMed] [Google Scholar]
- 21. Kotzampassi K, Giamarellos-Bourboulis EJ, Voudouris A, Kazamias P, Eleftheriadis E (2006) Benefits of a synbiotic formula (Synbiotic 2000Forte) in critically Ill trauma patients: early results of a randomized controlled trial. World J Surg 30: 1848–1855. [DOI] [PubMed] [Google Scholar]
- 22. Shanahan F (2002) Probiotics and inflammatory bowel disease: from fads and fantasy to facts and future. Br J Nutr 88 Suppl 1: S5–9. [DOI] [PubMed] [Google Scholar]
- 23. Khailova L, Mount Patrick SK, Arganbright KM, Halpern MD, Kinouchi T, et al. (2010) Bifidobacterium bifidum reduces apoptosis in the intestinal epithelium in necrotizing enterocolitis. Am J Physiol Gastrointest Liver Physiol 299: G1118–1127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Tao Y, Drabik KA, Waypa TS, Musch MW, Alverdy JC, et al. (2006) Soluble factors from Lactobacillus GG activate MAPKs and induce cytoprotective heat shock proteins in intestinal epithelial cells. Am J Physiol Cell Physiol 290: C1018–1030. [DOI] [PubMed] [Google Scholar]
- 25. Yan F, Cao H, Cover TL, Whitehead R, Washington MK, et al. (2007) Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 132: 562–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Yan F, Polk DB (2002) Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J Biol Chem 277: 50959–50965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Alberda C, Gramlich L, Meddings J, Field C, McCargar L, et al. (2007) Effects of probiotic therapy in critically ill patients: a randomized, double-blind, placebo-controlled trial. Am J Clin Nutr 85: 816–823. [DOI] [PubMed] [Google Scholar]
- 28. Anderson RC, Cookson AL, McNabb WC, Kelly WJ, Roy NC (2010) Lactobacillus plantarum DSM 2648 is a potential probiotic that enhances intestinal barrier function. FEMS Microbiol Lett 309: 184–192. [DOI] [PubMed] [Google Scholar]
- 29. Khailova L, Dvorak K, Arganbright KM, Halpern MD, Kinouchi T, et al. (2009) Bifidobacterium bifidum improves intestinal integrity in a rat model of necrotizing enterocolitis. Am J Physiol Gastrointest Liver Physiol 297: G940–949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Zareie M, Johnson-Henry K, Jury J, Yang PC, Ngan BY, et al. (2006) Probiotics prevent bacterial translocation and improve intestinal barrier function in rats following chronic psychological stress. Gut 55: 1553–1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Luyer MD, Buurman WA, Hadfoune M, Speelmans G, Knol J, et al. (2005) Strain-specific effects of probiotics on gut barrier integrity following hemorrhagic shock. Infect Immun 73: 3686–3692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Aguero G, Villena J, Racedo S, Haro C, Alvarez S (2006) Beneficial immunomodulatory activity of Lactobacillus casei in malnourished mice pneumonia: effect on inflammation and coagulation. Nutrition 22: 810–819. [DOI] [PubMed] [Google Scholar]
- 33. Arribas B, Rodriguez-Cabezas ME, Comalada M, Bailon E, Camuesco D, et al. (2009) Evaluation of the preventative effects exerted by Lactobacillus fermentum in an experimental model of septic shock induced in mice. Br J Nutr 101: 51–58. [DOI] [PubMed] [Google Scholar]
- 34. Matsumoto T, Ishikawa H, Tateda K, Yaeshima T, Ishibashi N, et al. (2008) Oral administration of Bifidobacterium longum prevents gut-derived Pseudomonas aeruginosa sepsis in mice. J Appl Microbiol 104: 672–680. [DOI] [PubMed] [Google Scholar]
- 35. Tok D, Ilkgul O, Bengmark S, Aydede H, Erhan Y, et al. (2007) Pretreatment with pro- and synbiotics reduces peritonitis-induced acute lung injury in rats. J Trauma 62: 880–885. [DOI] [PubMed] [Google Scholar]
- 36. Khailova L, Frank DN, Dominguez JA, Wischmeyer PE (2013) Probiotic Administration Reduces Mortality and Improves Intestinal Epithelial Homeostasis in Experimental Sepsis. Anesthesiology [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Annane D, Bellissant E, Cavaillon JM (2005) Septic shock. Lancet 365: 63–78. [DOI] [PubMed] [Google Scholar]
- 38. Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124: 783–801. [DOI] [PubMed] [Google Scholar]
- 39. Xu F, Xu Z, Zhang R, Wu Z, Lim JH, et al. (2008) Nontypeable Haemophilus influenzae induces COX-2 and PGE2 expression in lung epithelial cells via activation of p38 MAPK and NF-kappa B. Respir Res 9: 16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Kirkby NS, Zaiss AK, Wright WR, Jiao J, Chan MV, et al. (2013) Differential COX-2 induction by viral and bacterial PAMPs: Consequences for cytokine and interferon responses and implications for anti-viral COX-2 directed therapies. Biochem Biophys Res Commun 438: 249–256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Baker CC, Chaudry IH, Gaines HO, Baue AE (1983) Evaluation of factors affecting mortality rate after sepsis in a murine cecal ligation and puncture model. Surgery 94: 331–335. [PubMed] [Google Scholar]
- 42. Robertson CM, Perrone EE, McConnell KW, Dunne WM, Boody B, et al. (2008) Neutrophil depletion causes a fatal defect in murine pulmonary Staphylococcus aureus clearance. J Surg Res 150: 278–285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402–408. [DOI] [PubMed] [Google Scholar]
- 44. Hajjar AM, Harowicz H, Liggitt HD, Fink PJ, Wilson CB, et al. (2005) An essential role for non-bone marrow-derived cells in control of Pseudomonas aeruginosa pneumonia. Am J Respir Cell Mol Biol 33: 470–475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. MacFie J, O'Boyle C, Mitchell CJ, Buckley PM, Johnstone D, et al. (1999) Gut origin of sepsis: a prospective study investigating associations between bacterial translocation, gastric microflora, and septic morbidity. Gut 45: 223–228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Wang X, Andersson R, Soltesz V, Leveau P, Ihse I (1996) Gut origin sepsis, macrophage function, and oxygen extraction associated with acute pancreatitis in the rat. World J Surg 20: 299–307 discussion 307–298. [DOI] [PubMed] [Google Scholar]
- 47. Cleveland J, Montville TJ, Nes IF, Chikindas ML (2001) Bacteriocins: safe, natural antimicrobials for food preservation. Int J Food Microbiol 71: 1–20. [DOI] [PubMed] [Google Scholar]
- 48. Lee YK, Lim CY, Teng WL, Ouwehand AC, Tuomola EM, et al. (2000) Quantitative approach in the study of adhesion of lactic acid bacteria to intestinal cells and their competition with enterobacteria. Appl Environ Microbiol 66: 3692–3697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Yan F, Cao H, Cover TL, Washington MK, Shi Y, et al. (2011) Colon-specific delivery of a probiotic-derived soluble protein ameliorates intestinal inflammation in mice through an EGFR-dependent mechanism. J Clin Invest 121: 2242–2253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Harb H, van Tol EA, Heine H, Braaksma M, Gross G, et al. (2013) Neonatal supplementation of processed supernatant from Lactobacillus rhamnosus GG improves allergic airway inflammation in mice later in life. Clin Exp Allergy 43: 353–364. [DOI] [PubMed] [Google Scholar]
- 51. Yan F, Polk DB (2012) Characterization of a probiotic-derived soluble protein which reveals a mechanism of preventive and treatment effects of probiotics on intestinal inflammatory diseases. Gut Microbes 3: 25–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. van Baarlen P, Troost F, van der Meer C, Hooiveld G, Boekschoten M, et al. (2011) Human mucosal in vivo transcriptome responses to three lactobacilli indicate how probiotics may modulate human cellular pathways. Proc Natl Acad Sci U S A 108 Suppl 1: 4562–4569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Ashraf R, Shah NP (2014) Immune system stimulation by probiotic microorganisms. Crit Rev Food Sci Nutr 54: 938–956. [DOI] [PubMed] [Google Scholar]
- 54. Dani C, Biadaioli R, Bertini G, Martelli E, Rubaltelli FF (2002) Probiotics feeding in prevention of urinary tract infection, bacterial sepsis and necrotizing enterocolitis in preterm infants. A prospective double-blind study. Biol Neonate 82: 103–108. [DOI] [PubMed] [Google Scholar]
- 55. McNaught CE, Woodcock NP, Anderson AD, MacFie J (2005) A prospective randomised trial of probiotics in critically ill patients. Clin Nutr 24: 211–219. [DOI] [PubMed] [Google Scholar]
- 56. Honeycutt TC, El Khashab M, Wardrop RM 3rd, McNeal-Trice K, Honeycutt AL, et al. (2007) Probiotic administration and the incidence of nosocomial infection in pediatric intensive care: a randomized placebo-controlled trial. Pediatr Crit Care Med 8: 452–458 quiz 464. [DOI] [PubMed] [Google Scholar]
- 57. Netea MG, van der Meer JW, van Deuren M, Kullberg BJ (2003) Proinflammatory cytokines and sepsis syndrome: not enough, or too much of a good thing? Trends Immunol 24: 254–258. [DOI] [PubMed] [Google Scholar]
- 58. Kono Y, Inomata M, Hagiwara S, Shiraishi N, Noguchi T, et al. (2011) A newly synthetic vitamin E derivative, E-Ant-S-GS, attenuates lung injury caused by cecal ligation and puncture-induced sepsis in rats. Surgery [DOI] [PubMed] [Google Scholar]
- 59. Williams DL, Ha T, Li C, Kalbfleisch JH, Laffan JJ, et al. (1999) Inhibiting early activation of tissue nuclear factor-kappa B and nuclear factor interleukin 6 with (1→3)-beta-D-glucan increases long-term survival in polymicrobial sepsis. Surgery 126: 54–65. [DOI] [PubMed] [Google Scholar]
- 60. Regel G, Grotz M, Weltner T, Sturm JA, Tscherne H (1996) Pattern of organ failure following severe trauma. World J Surg 20: 422–429. [DOI] [PubMed] [Google Scholar]
- 61. Farley KS, Wang LF, Razavi HM, Law C, Rohan M, et al. (2006) Effects of macrophage inducible nitric oxide synthase in murine septic lung injury. Am J Physiol Lung Cell Mol Physiol 290: L1164–1172. [DOI] [PubMed] [Google Scholar]
- 62. Singleton KD, Wischmeyer PE (2007) Glutamine's protection against sepsis and lung injury is dependent on heat shock protein 70 expression. Am J Physiol Regul Integr Comp Physiol 292: R1839–1845. [DOI] [PubMed] [Google Scholar]
- 63. Racedo S, Villena J, Medina M, Aguero G, Rodriguez V, et al. (2006) Lactobacillus casei administration reduces lung injuries in a Streptococcus pneumoniae infection in mice. Microbes Infect 8: 2359–2366. [DOI] [PubMed] [Google Scholar]
- 64. Appleby SB, Ristimaki A, Neilson K, Narko K, Hla T (1994) Structure of the human cyclo-oxygenase-2 gene. Biochem J 302 Pt 3: 723–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Rajapakse N, Kim MM, Mendis E, Kim SK (2008) Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 in lipopolysaccharide-stimulated RAW264.7 cells by carboxybutyrylated glucosamine takes place via down-regulation of mitogen-activated protein kinase-mediated nuclear factor-kappaB signaling. Immunology 123: 348–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Zhang LN, Zheng JJ, Zhang L, Gong X, Huang H, et al. (2011) Protective effects of asiaticoside on septic lung injury in mice. Exp Toxicol Pathol 63: 519–525. [DOI] [PubMed] [Google Scholar]
- 67. Li XH, Gong X, Zhang L, Jiang R, Li HZ, et al. (2013) Protective effects of polydatin on septic lung injury in mice via upregulation of HO-1. Mediators Inflamm 2013: 354087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68. Ang SF, Sio SW, Moochhala SM, MacAry PA, Bhatia M (2011) Hydrogen sulfide upregulates cyclooxygenase-2 and prostaglandin E metabolite in sepsis-evoked acute lung injury via transient receptor potential vanilloid type 1 channel activation. J Immunol 187: 4778–4787. [DOI] [PubMed] [Google Scholar]
- 69. Bitto A, Minutoli L, David A, Irrera N, Rinaldi M, et al. (2012) Flavocoxid, a dual inhibitor of COX-2 and 5-LOX of natural origin, attenuates the inflammatory response and protects mice from sepsis. Crit Care 16: R32. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 70. Akira S, Takeda K (2004) Toll-like receptor signalling. Nat Rev Immunol 4: 499–511. [DOI] [PubMed] [Google Scholar]
- 71. Tsujimoto H, Ono S, Hiraki S, Majima T, Kawarabayashi N, et al. (2004) Hemoperfusion with polymyxin B-immobilized fibers reduced the number of CD16+ CD14+ monocytes in patients with septic shock. J Endotoxin Res 10: 229–237. [DOI] [PubMed] [Google Scholar]
- 72. Armstrong L, Medford AR, Hunter KJ, Uppington KM, Millar AB (2004) Differential expression of Toll-like receptor (TLR)-2 and TLR-4 on monocytes in human sepsis. Clin Exp Immunol 136: 312–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Harter L, Mica L, Stocker R, Trentz O, Keel M (2004) Increased expression of toll-like receptor-2 and -4 on leukocytes from patients with sepsis. Shock 22: 403–409. [DOI] [PubMed] [Google Scholar]
- 74. Tsujimoto H, Ono S, Majima T, Kawarabayashi N, Takayama E, et al. (2005) Neutrophil elastase, MIP-2, and TLR-4 expression during human and experimental sepsis. Shock 23: 39–44. [DOI] [PubMed] [Google Scholar]
- 75. Williams DL, Ha T, Li C, Kalbfleisch JH, Schweitzer J, et al. (2003) Modulation of tissue Toll-like receptor 2 and 4 during the early phases of polymicrobial sepsis correlates with mortality. Crit Care Med 31: 1808–1818. [DOI] [PubMed] [Google Scholar]
- 76. Edelman DA, Jiang Y, Tyburski J, Wilson RF, Steffes C (2006) Toll-like receptor-4 message is up-regulated in lipopolysaccharide-exposed rat lung pericytes. J Surg Res 134: 22–27. [DOI] [PubMed] [Google Scholar]
- 77. Yu M, Shao D, Liu J, Zhu J, Zhang Z, et al. (2007) Effects of ketamine on levels of cytokines, NF-kappaB and TLRs in rat intestine during CLP-induced sepsis. Int Immunopharmacol 7: 1076–1082. [DOI] [PubMed] [Google Scholar]