

Abstract
Mesenchymal stem cells (MSCs) are present in diverse tissues and organs, including bone marrow, umbilical cord, adipose tissue, and placenta. MSCs can expand easily in vitro and have regenerative stem cell properties and potent immunoregulatory activity. They inhibit the functions of dendritic cells, B cells, and T cells, but enhance those of regulatory T cells by producing immunoregulatory molecules such as transforming growth factor-β, hepatic growth factors, prostaglandin E2, interleukin-10, indolamine 2,3-dioxygenase, nitric oxide, heme oxygenase-1, and human leukocyte antigen-G. These properties make MSCs promising therapeutic candidates for the treatment of autoimmune diseases. Here, we review the preclinical studies of MSCs in animal models for systemic lupus erythematosus, rheumatoid arthritis, Crohn's disease, and experimental autoimmune encephalomyelitis, and summarize the underlying immunoregulatory mechanisms.
Keywords: Mesenchymal stem cells, Immunoregulation, Autoimmune diseases
MESENCHYMAL STEM CELLS
Stem cells have self-renewal potential and differentiate into one or more specialized cell types (1). There are two types of stem cells, embryonic and adult. Embryonic stem cells, which are isolated from blastocysts, are able to differentiate into any organ-specific cells, whereas adult stem cells show restricted proliferation and lineage differentiation (2). Adult stem cells undergoing mesodermal lineage-specific differentiation to osteocytes, adipocytes, or chondrocytes are named mesenchymal stem cells (MSCs) (3). Although embryonic stem cells can differentiate into any cell types in humans, teratoma formation and ethical concerns limit their wide clinical application (4). In contrast, MSCs can be used for tissue repair and regeneration without these restrictions.
MSCs can be isolated from various human tissues, such as bone marrow, adipose tissue, umbilical cord blood, and placenta. Typical MSC markers include CD73, CD90, and CD105; however, MSCs do not express the hematopoietic markers CD34 and CD45. Upon exposure to specific differentiation media, MSCs differentiate into osteocytes, chondrocytes, adipocytes or other cell types (5,6). Cultured MSCs adhere to tissue culture plates (7,8). MSCs secrete various soluble factors that promote angiogenesis and mitosis and reduce apoptosis, and can be used for tissue repair or regeneration (9,10,11). MSCs can produce a number of immunoregulatory molecules and have advantages for clinical applications, such as easy preparation and low immunogenicity (12,13,14,15). Overall, these properties make MSCs good therapeutic candidates for the treatment of transplantation rejection, graft-versus-host diseases, and autoimmune diseases (13,16). In this review, we discuss the potential use of MSCs for the treatment of autoimmune diseases, such as systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), Crohn's disease, and experimental autoimmune encephalomyelitis (EAE).
EFFICACY OF MESENCHYMAL STEM CELLS IN ANIMAL MODELS OF AUTOIMMUNE DISEASES
MSCs show promising therapeutic activity in animal models of SLE (Table I), RA (Table II), Crohn's disease, and EAE (Table III). Overall experimental strategies can be summarized as follows. Human MSCs are usually isolated from BM, but sometimes from umbilical cord or adipose tissues. MSCs are injected intravenously or intraperitoneally at a dose of approximately 1×106 cells per mouse.
Table I.
Preclinical efficacy of mesenchymal stem cell (MSC) in animal models for systemic lupus erythematosus
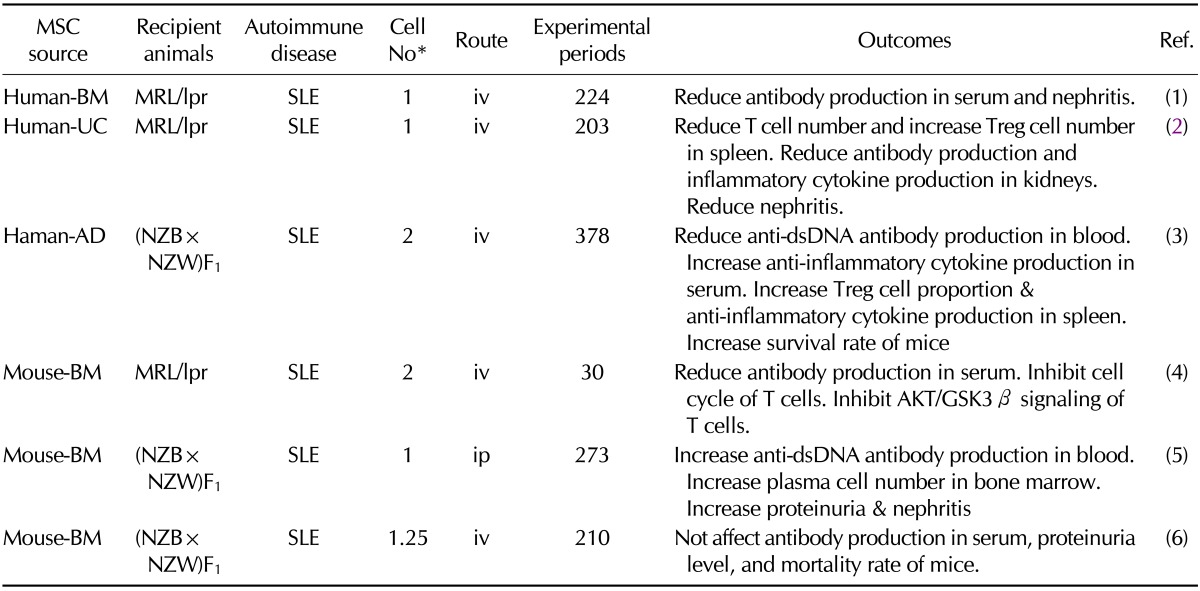
*(×106 cells/mouse). iv: intravenous injection, ip: intraperitoneal injection, Treg: regulatory T cells, SLE: systemic lupus erythematosus, BM: bone marrow, UC: umbilical cord blood, AD: adipose tissue, MSC: mesenchymal stem cell
Table II.
Preclinical efficacy of mesenchymal stem cells in animal models for rheumatoid arthritis
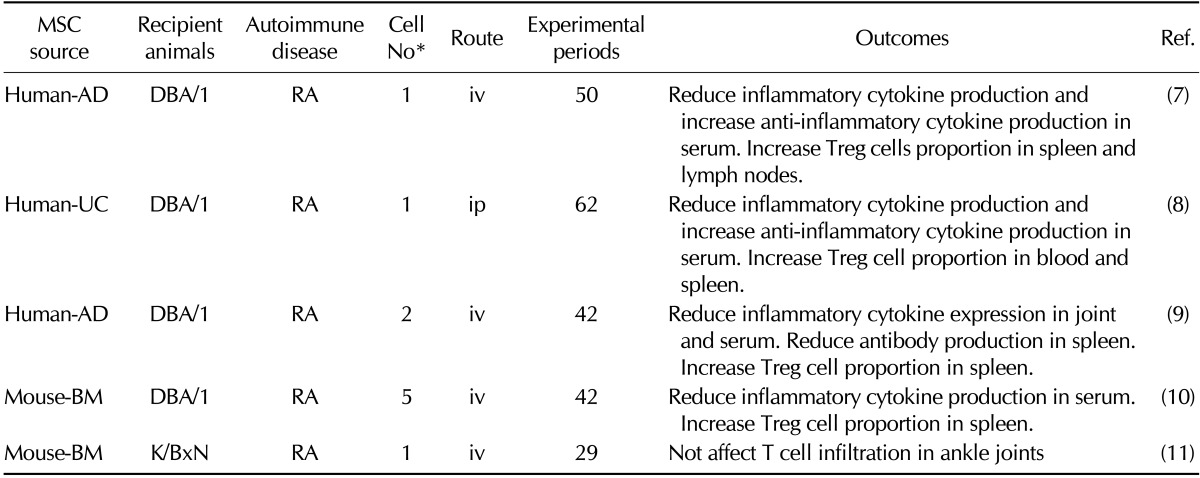
*(×106 cells/mouse). iv: intravenous injection, ip: intraperitoneal injection, Treg: regulatory T cells, RA: rheumatoid arthritis, BM: bone marrow, UC: umbilical cord blood, AD: adipose tissue, MSC: mesenchymal stem cell
Table III.
Preclinical efficacy of mesenchymal stem cells in animal models for Crohn's disease and experimental autoimmune encephalomyelitis
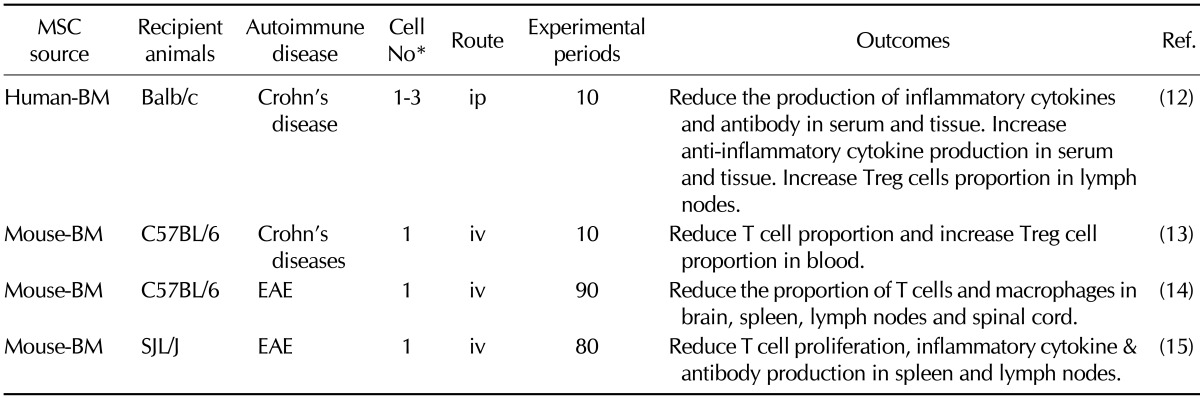
*(×106 cells/mouse). iv: intravenous injection, ip: intraperitoneal injection, Treg: regulatory T cells, EAE: experimental autoimmune encephalomyelitis, BM: bone marrow, UC: umbilical cord blood, AD: adipose tissue, MSC: mesenchymal stem cell
Systemic lupus erythematosus
SLE is a severe autoimmune disease characterized by multi-organ dysfunctions including renal, cardiovascular, neural, musculoskeletal, and cutaneous involvement (17). SLE is characterized by activation and proliferation of autoreactive T and B cells (18). MRL/lpr mice, which have a mutation in the lpr gene, spontaneously develop an autoimmune disease that is very similar to human SLE (19). Zhou and colleagues investigated the effect of human BM-derived MSCs on the pathogenesis of SLE in MRL/lpr mice (19). Human MSCs reduced proliferation of T cells from MRL/lpr mice in vitro. MSCs injected intravenously reduced serum levels of anti-dsDNA antibodies and proteinuria in MRL/lpr mice. Immunohistochemical analysis showed low expression levels of TGF-β, vascular endothelial growth factor, and complement C3 in renal tissues. Gu and colleagues isolated MSCs from human umbilical cord and injected them intravenously into MRL/lpr mice, which decreased the levels of proteinuria, serum creatinine and anti-dsDNA antibodies, and the extent of renal injury (17). In contrast, MSCs increased the number of Treg cells in the spleen. Ji and colleagues investigated how MSCs inhibited T cell proliferation in vivo (18). Intravenous injection of BM-derived MSCs reduced the serum anti-dsDNA antibody level. MSCs inhibited G1/S transition in T cells in the spleen and lymph nodes through a decrease in CDK2 expression. Furthermore, MSCs inhibited the Akt/GSK3β signaling pathway of T cells from MRL/lpr mice. These studies suggest that MSCs can ameliorate SLE pathogenesis by inhibiting the functions of T and B cells and activating Treg cells in the MRL/lpr mouse model. Controversial data were shown in NZB × NZW F1 (NZB/W) mouse models of SLE (20). MSCs from Balb/c mice were injected into NZB/W mice before and after disease onset, which showed worsen disease progression by showing increase of anti-dsDNA antibody production, plasma cell number, proteinuria level, and nephiritis (20). MSCs from C57BL/6 mice also did not affect the antibody production, proteinuria level, and the mortality rates (21). However, human MSCs ameliorated SLE without adverse effect in NZB/W mice (22). Human MSCs increased the survival rate and the proportion of Treg cells, but decreased the levels of anti-dsDNA antibody and proteinuria (22).
Rheumatoid arthritis
RA is characterized by the loss of self-tolerance, chronic inflammation in the joints, subsequent cartilage destruction, and bone erosion. The crucial process underlying RA initiation is the abnormal activation of DCs, T cells, B cells, macrophages, and neutrophils (23). The anti-inflammatory effects of MSCs in RA have been mainly studied in the collagen-induced arthritis model in DBA/1 mice. González et al. injected human adipose tissue-derived MSCs intravenously into collagen-injected DBA/1 mice. Systemic infusion of these cells significantly reduced the incidence and severity of experimental arthritis by decreasing production of various inflammatory cytokines and chemokines, and reducing the ratios of Th1/Th17 cells. MSCs induced production of anti-inflammatory IL-10 in lymph nodes and joints, and de novo generation of antigen-specific Treg cells. Liu and colleagues injected human umbilical cord-derived MSCs intraperitoneally into DBA/1 mice (24). Systemic infusion of these cells reduced the severity of arthritis. MSCs reduced the levels of proinflammatory cytokines and chemokines (TNF-α, IL-6 and monocyte chemoattractant protein-1), increased the levels of IL-10, shifted from Th2 to Th1 type responses, and induced Treg cells. Zhou and colleagues injected human adipose tissue-derived MSCs intravenously into DBA/1 mice (25). These MSCs reduced the incidence and severity of arthritis by inhibiting production of various inflammatory mediators and reducing antigen-specific Th1 cell expansion. MSCs also induced production of anti-inflammatory cytokines and generation of antigen-specific Treg cells. Human BM-derived MSCs showed similar efficacy in a collagen-induced arthritis model in DBA/1 mice as umbilical human cord- or adipose tissue-derived MSCs (26). These studies suggest that MSCs can ameliorate RA pathogenesis in DBA/1 mice by inhibiting the production of inflammatory cytokines (which are mainly produced by macrophages and T cells) and activating Treg cells. However, contradictory data was reported in adjuvant-induced and spontaneous (K/BxN) arthritis model and showed that MSCs were effective when administered only before disease onset, which suggested that MSCs lost their immunoregulatory properties when infused into inflammatory microenvironments (27).
Crohn's disease
Crohn's disease is a chronic form of inflammatory bowel disease and characterized by dysfunction of intestinal T cells, abnormal cytokine production, and inflammation in small intestine and the colonic mucosa (28). Pathogenesis of Crohn's disease is related to abnormal activation of Th1 cells, macrophages and neutrophils, uncontrolled production of inflammatory cytokines and chemokines, and an imbalance between effector T cells and suppressive Treg cells (29). In a preclinical study of the effect of MSCs on Crohn's disease, colitis was induced in mice by trinitrobenzene sulfonic acid, and human adipose tissue-derived MSCs were injected intraperitoneally after the onset of the disease (28). Systemic infusion of MSCs ameliorated the clinical and histopathologic severity of colitis, abrogated body weight loss, diarrhea and inflammation, and increased survival. MSCs down-regulated Th1-driven inflammatory responses and decreased the production of inflammatory cytokines and chemokines. MSCs also impaired Th1 cell expansion and activated Treg cells (28). Systemic infusion of human BM-derived MSCs ameliorated the pathogenesis of dextran sulfate sodium salt-induced experimental colitis and induced T cell apoptosis via the FASL-FAS pathway in vivo (30). These studies indicate that MSCs can ameliorate chemically induced colitis by inhibiting the inflammatory functions of macrophages and T cells in mice.
Experimental autoimmune encephalomyelitis
EAE is an autoimmune disease of the central nervous system, which is mediated by T cells and macrophages (31). EAE was induced in mice by injecting myelin oligodendrocyte glycoprotein-35-55 peptide, and human BM-derived MSCs were injected intravenously before and after the onset of the disease (31). MSCs ameliorated EAE progression showing low infiltration of inflammatory cells when injected before its onset, but their efficacy was relatively weak when they were injected after the onset (31). Human BM-derived MSCs were injected into another animal model of EAE, mice immunized with the peptide 139-151 of the proteolipid protein (32). MSC-injected mice ameliorated disease progression and low relapses, with decreased infiltration of inflammatory cells and decreased demyelination and axonal loss. MSCs reduced antigen-specific T-cell response and antibody titers. These studies suggest that MSCs can ameliorate neuro-inflammation by inhibiting the infiltration and functions of inflammatory cells in animal models.
IMMUNOREGULATORY MECHANISMS
MSCs have diverse immunoregulatory activities, which vary depending on immune cell subpopulations. MSCs affect differentiation, maturation, and function of dendritic cells (DCs) (33). MSCs inhibit the initial differentiation of CD14+ monocytes into immature DCs (34) and also inhibit DC maturation by decreasing the expression of HLA-DR, CD1a, CD80, CD83, and CD86, and the production of IL-1β, interleukin (IL)-12, and tumor necrosis factor (TNF)-α by DCs (35,36,37,38). MSCs impair the activation of lipopolysaccharide-treated DCs, reduce antigen presentation to CD4+ and CD8+ T cells and inhibit secretion of inflammatory cytokines (33,39). MSCs also reduce the expression of CCR7 and prevent DC homing to lymph nodes, where the main functions of DCs are to activate T cells (39). MSCs might affect DC functions via soluble mediators, such as IL-6, macrophage colony-stimulating factor (M-CSF), and prostaglandin E2 (PGE2) (40,41).
MSCs can regulate T cell functions via two ways. First, MSCs directly inhibit the functions of antigen-specific T cells (42,43). Second, MSCs inhibit T cell functions indirectly by stimulating the expansion of regulatory T (Treg) cells (36). The inhibitory effect of MSCs on T cells has been demonstrated in an animal model of graft-versus-host disease, which is mainly caused by abnormal activation of T cells (44). MSCs reduce allograft rejection in a mouse model (13). In an RA model, MSCs inhibit the activation of CD4+ and CD8+ T cells, and increase the number of and IL-10 production by Treg cells (45). MSCs affect T cell functions by producing transforming growth factor (TGF)-β, PGE2, indolamine 2,3-dioxygenase (IDO), heme oxygenase-1 (HO-1), and chemokines (46). Inflammatory cytokines, including interferon (IFN)-γ and tumor necrosis factor (TNF)-α released from cells in damaged tissues, strongly induce IDO expression by MSCs (47). HO-1 produced by human MSCs is able to promote the expansion of Treg cells (48). MSCs release CCL2, trigger T cell migration, directly contact with T cells, and induce T cell apoptosis through Fas-FasL interaction (30). Inflammatory cytokines, such as TNF-α and IL-1β, up-regulate the expression of ICAM-1 and VCAM-1 in MSCs, which strengthens their interaction with T cells (49).
MSCs can also modulate B cell functions (mainly antibody production). B cell proliferation is not affected by naïve MSCs, but is inhibited by IFN-γ-treated MSCs in transwell culture systems (47). MSCs inhibit B cell proliferation by arresting the G0/G1 phase of the cell cycle, but not by inducing apoptosis (50). MSCs inhibit B cell differentiation to plasma cells producing IgM, IgG, and IgA, and inhibit the expression of CXCR4, CXCR5, and CCR7 by B cells, resulting in reduced homing to lymph nodes (50). However, MSCs do not affect the expression of co-stimulatory molecules and cytokine production by B cells. MSCs directly inhibit B cell functions by producing soluble factors, as indicated by transwell experiments. MSC also indirectly affect B cell functions by inhibiting T cells, which play a key role in antigen-specific antibody production by B cells, and plasmacytoid DCs, which are crucial for B cell maturation (50,51).
CONCLUSIONS
MSCs are one of the promising therapeutic candidates for the treatment of autoimmune diseases and show beneficial effects in animal models of SLE, RA, Crohn's disease, and EAE. MSCs show beneficial therapeutic activity in animal models of these autoimmune diseases through inhibiting abnormally activated immune functions of dendritic cells, T cells, and B cells. However, several contradictory papers were also noted in preclinical efficacy evaluation study of MSCs in autoimmune animal models (20,21,22,27). Such contradiction may be due to several factors. First, properties and characteristics of MSCs might be diverse according to sources (organs or species) and culture conditions. Second, each laboratory used different experimental methods; route, number, and timing (before and after disease onset) of injection. Third, animal models were diverse according to the studies. Each animal model that is used in preclinical studies has typical pathophysiology and has both strength and weakness. Therefore, no single model may represent human disease and predict accurately the efficacy of MSCs. Overall, this observation strong emphasizes the importance of the usage of qualified or standardized MSCs and the usage of appropriate animal models. In addition, several basic questions remain to be answered. First, it is critical to understand the normal physiological functions of MSCs in vivo, since there is limited information on the physiological roles of MSCs in vivo. Second, with respect to the therapeutic mechanisms and efficacy, survival time, distribution and tissue homing, and action site (local or systemic) of MSCs should be further studied in vivo. Third, it is also important to understand the trafficking of MSCs into lymphoid tissues, where they interact with immune cells. Understanding the mechanisms of MSC action in treatment of autoimmune diseases will help to expand clinical application of MSCs to autoimmune diseases.
ACKNOWLEDGEMENTS
This research was supported by the grants from Korean government (13172MFDS303 & NRF 2008-0062275).
Abbreviations
- BM
bone marrow
- DC
dendritic cell
- EAE
experimental autoimmune encephalomyelitis
- HO
heme oxygenase
- IDO
indolamine 2,3-dioxygenase
- IFN
interferonIL, interleukin
- MSC
mesenchymal stem cell
- PG
prostaglandin
- RA
rheumatoid arthritis
- SLE
systemic lupus erythematosus
- TGF
tranforming growth factor
- Th
helper T
- TNF
tumor necrosis factor
- Treg
regulatory T
Footnotes
The authors have no financial conflict of interest.
References
- 1.Watt FM, Hogan BL. Out of Eden: stem cells and their niches. Science. 2000;287:1427–1430. doi: 10.1126/science.287.5457.1427. [DOI] [PubMed] [Google Scholar]
- 2.Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, Jones JM. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–1147. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- 3.Friedenstein AJ, Chailakhjan RK, Lalykina KS. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Tissue Kinet. 1970;3:393–403. doi: 10.1111/j.1365-2184.1970.tb00347.x. [DOI] [PubMed] [Google Scholar]
- 4.Nussbaum J, Minami E, Laflamme MA, Virag JA, Ware CB, Masino A, Muskheli V, Pabon L, Reinecke H, Murry CE. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. FASEB J. 2007;21:1345–1357. doi: 10.1096/fj.06-6769com. [DOI] [PubMed] [Google Scholar]
- 5.Kon E, Muraglia A, Corsi A, Bianco P, Marcacci M, Martin I, Boyde A, Ruspantini I, Chistolini P, Rocca M, Giardino R, Cancedda R, Quarto R. Autologous bone marrow stromal cells loaded onto porous hydroxyapatite ceramic accelerate bone repair in critical-size defects of sheep long bones. J Biomed Mater Res. 2000;49:328–337. doi: 10.1002/(sici)1097-4636(20000305)49:3<328::aid-jbm5>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- 6.Solchaga LA, Temenoff JS, Gao J, Mikos AG, Caplan AI, Goldberg VM. Repair of osteochondral defects with hyaluronan- and polyester-based scaffolds. Osteoarthritis Cartilage. 2005;13:297–309. doi: 10.1016/j.joca.2004.12.016. [DOI] [PubMed] [Google Scholar]
- 7.Horwitz EM, Le Blanc K, Dominici M, Mueller I, Slaper-Cortenbach I, Marini FC, Deans RJ, Krause DS, Keating A International Society for Cellular Therapy. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy. 2005;7:393–395. doi: 10.1080/14653240500319234. [DOI] [PubMed] [Google Scholar]
- 8.Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A, Prockop Dj, Horwitz E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–317. doi: 10.1080/14653240600855905. [DOI] [PubMed] [Google Scholar]
- 9.Amado LC, Saliaris AP, Schuleri KH, St John M, Xie JS, Cattaneo S, Durand DJ, Fitton T, Kuang JQ, Stewart G, Lehrke S, Baumgartner WW, Martin BJ, Heldman AW, Hare JM. Cardiac repair with intramyocardial injection of allogeneic mesenchymal stem cells after myocardial infarction. Proc Natl Acad Sci USA. 2005;102:11474–11479. doi: 10.1073/pnas.0504388102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lin YT, Chern Y, Shen CK, Wen HL, Chang YC, Li H, Cheng TH, Hsieh-Li HM. Human mesenchymal stem cells prolong survival and ameliorate motor deficit through trophic support in Huntington's disease mouse models. PLoS One. 2011;6:e22924. doi: 10.1371/journal.pone.0022924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Miyahara Y, Nagaya N, Kataoka M, Yanagawa B, Tanaka K, Hao H, Ishino K, Ishida H, Shimizu T, Kangawa K, Sano S, Okano T, Kitamura S, Mori H. Monolayered mesenchymal stem cells repair scarred myocardium after myocardial infarction. Nat Med. 2006;12:459–465. doi: 10.1038/nm1391. [DOI] [PubMed] [Google Scholar]
- 12.Ball LM, Bernardo ME, Roelofs H, Lankester A, Cometa A, Egeler RM, Locatelli F, Fibbe WE. Cotransplantation of ex vivo expanded mesenchymal stem cells accelerates lymphocyte recovery and may reduce the risk of graft failure in haploidentical hematopoietic stem-cell transplantation. Blood. 2007;110:2764–2767. doi: 10.1182/blood-2007-04-087056. [DOI] [PubMed] [Google Scholar]
- 13.Le Blanc K, Frassoni F, Ball L, Locatelli F, Roelofs H, Lewis I, Lanino E, Sundberg B, Bernardo ME, Remberger M, Dini G, Egeler RM, Bacigalupo A, Fibbe W, Ringden O Developmental Committee of the European Group for Blood and Marrow Transplantation. Mesenchymal stem cells for treatment of steroid-resistant, severe, acute graft-versus-host disease: a phase II study. Lancet. 2008;371:1579–1586. doi: 10.1016/S0140-6736(08)60690-X. [DOI] [PubMed] [Google Scholar]
- 14.Duijvestein M, Vos AC, Roelofs H, Wildenberg ME, Wendrich BB, Verspaget HW, Kooy-Winkelaar EM, Koning F, Zwaginga JJ, Fidder HH, Verhaar AP, Fibbe WE, van den Brink GR, Hommes DW. Autologous bone marrow-derived mesenchymal stromal cell treatment for refractory luminal Crohn's disease: results of a phase I study. Gut. 2010;59:1662–1669. doi: 10.1136/gut.2010.215152. [DOI] [PubMed] [Google Scholar]
- 15.Ciccocioppo R, Bernardo ME, Sgarella A, Maccario R, Avanzini MA, Ubezio C, Minelli A, Alvisi C, Vanoli A, Calliada F, Dionigi P, Perotti C, Locatelli F, Corazza GR. Autologous bone marrow-derived mesenchymal stromal cells in the treatment of fistulising Crohn's disease. Gut. 2011;60:788–798. doi: 10.1136/gut.2010.214841. [DOI] [PubMed] [Google Scholar]
- 16.Le Blanc K, Rasmusson I, Sundberg B, Gotherstrom C, Hassan M, Uzunel M, Ringden O. Treatment of severe acute graft-versus-host disease with third party haploidentical mesenchymal stem cells. Lancet. 2004;363:1439–1441. doi: 10.1016/S0140-6736(04)16104-7. [DOI] [PubMed] [Google Scholar]
- 17.Gu Z, Akiyama K, Ma X, Zhang H, Feng X, Yao G, Hou Y, Lu L, Gilkeson GS, Silver RM, Zeng X, Shi S, Sun L. Transplantation of umbilical cord mesenchymal stem cells alleviates lupus nephritis in MRL/lpr mice. Lupus. 2010;19:1502–1514. doi: 10.1177/0961203310373782. [DOI] [PubMed] [Google Scholar]
- 18.Ji S, Guo Q, Han Y, Tan G, Luo Y, Zeng F. Mesenchymal stem cell transplantation inhibits abnormal activation of Akt/GSK3beta signaling pathway in T cells from systemic lupus erythematosus mice. Cell Physiol Biochem. 2012;29:705–712. doi: 10.1159/000178590. [DOI] [PubMed] [Google Scholar]
- 19.Zhou K, Zhang H, Jin O, Feng X, Yao G, Hou Y, Sun L. Transplantation of human bone marrow mesenchymal stem cell ameliorates the autoimmune pathogenesis in MRL/lpr mice. Cell Mol Immunoly. 2008;5:417–424. doi: 10.1038/cmi.2008.52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Youd M, Blickarz C, Woodworth L, Touzjian T, Edling A, Tedstone J, Ruzek M, Tubo R, Kaplan J, Lodie T. Allogeneic mesenchymal stem cells do not protect NZBxNZW F1 mice from developing lupus disease. Clin Exp Immunol. 2010;161:176–186. doi: 10.1111/j.1365-2249.2010.04158.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Schena F, Gambini C, Gregorio A, Mosconi M, Reverberi D, Gattorno M, Casazza S, Uccelli A, Moretta L, Martini A, Traggiai E. Interferon-gamma-dependent inhibition of B cell activation by bone marrow-derived mesenchymal stem cells in a murine model of systemic lupus erythematosus. Arthritis Rheum. 2010;62:2776–2786. doi: 10.1002/art.27560. [DOI] [PubMed] [Google Scholar]
- 22.Choi EW, Shin IS, Park SY, Park JH, Kim JS, Yoon EJ, Kang SK, Ra JC, Hong SH. Reversal of serologic, immunologic, and histologic dysfunction in mice with systemic lupus erythematosus by long-term serial adipose tissue-derived mesenchymal stem cell transplantation. Arthritis Rheum. 2012;64:243–253. doi: 10.1002/art.33313. [DOI] [PubMed] [Google Scholar]
- 23.Firestein GS. Evolving concepts of rheumatoid arthritis. Nature. 2003;423:356–361. doi: 10.1038/nature01661. [DOI] [PubMed] [Google Scholar]
- 24.Liu Y, Mu R, Wang S, Long L, Liu X, Li R, Sun J, Guo J, Zhang X, Guo J, Yu P, Li C, Liu X, Huang Z, Wang D, Li H, Gu Z, Liu B, Li Z. Therapeutic potential of human umbilical cord mesenchymal stem cells in the treatment of rheumatoid arthritis. Arthritis Res Ther. 2010;12:R210. doi: 10.1186/ar3187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhou B, Yuan J, Zhou Y, Ghawji M, Jr, Deng YP, Lee AJ, Lee AJ, Nair U, Kang AH, Brand DD, Yoo TJ. Administering human adipose-derived mesenchymal stem cells to prevent and treat experimental arthritis. Clin Immunol. 2011;141:328–337. doi: 10.1016/j.clim.2011.08.014. [DOI] [PubMed] [Google Scholar]
- 26.Augello A, Tasso R, Negrini SM, Cancedda R, Pennesi G. Cell therapy using allogeneic bone marrow mesenchymal stem cells prevents tissue damage in collagen-induced arthritis. Arthritis Rheum. 2007;56:1175–1186. doi: 10.1002/art.22511. [DOI] [PubMed] [Google Scholar]
- 27.Papadopoulou A, Yiangou M, Athanasiou E, Zogas N, Kaloyannidis P, Batsis I, Fassas A, Anagnostopoulos A, Yannaki E. Mesenchymal stem cells are conditionally therapeutic in preclinical models of rheumatoid arthritis. Ann Rheum Dis. 2012;71:1733–1740. doi: 10.1136/annrheumdis-2011-200985. [DOI] [PubMed] [Google Scholar]
- 28.Gonzalez MA, Gonzalez-Rey E, Rico L, Buscher D, Delgado M. Adipose-derived mesenchymal stem cells alleviate experimental colitis by inhibiting inflammatory and autoimmune responses. Gastroenterology. 2009;136:978–989. doi: 10.1053/j.gastro.2008.11.041. [DOI] [PubMed] [Google Scholar]
- 29.Bouma G, Strober W. The immunological and genetic basis of inflammatory bowel disease. Nat Rev Immunol. 2003;3:521–533. doi: 10.1038/nri1132. [DOI] [PubMed] [Google Scholar]
- 30.Akiyama K, Chen C, Wang D, Xu X, Qu C, Yamaza T, Cai T, Chen W, Sun L, Shi S. Mesenchymalstem-cell-induced immunoregulation involves FAS-ligand-/FAS-mediated T cell apoptosis. Cell Stem Cell. 2012;10:544–555. doi: 10.1016/j.stem.2012.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zappia E, Casazza S, Pedemonte E, Benvenuto F, Bonanni I, Gerdoni E, Giunti D, Ceravolo A, Cazzanti F, Frassoni F, Mancardi G, Uccelli A. Mesenchymal tem cells ameliorate experimental autoimmune encephalomyelitis inducing T-cell anergy. Blood. 2005;106:1755–1761. doi: 10.1182/blood-2005-04-1496. [DOI] [PubMed] [Google Scholar]
- 32.Gerdoni E, Gallo B, Casazza S, Musio S, Bonanni I, Pedemonte E, Mantegazza R, Frassoni F, Mancardi G, Pedotti R, Uccelli A. Mesenchymal stem cells effectively modulate pathogenic immune response in experimental autoimmune encephalomyelitis. Ann Neurol. 2007;61:219–227. doi: 10.1002/ana.21076. [DOI] [PubMed] [Google Scholar]
- 33.Nauta AJ, Kruisselbrink AB, Lurvink E, Willemze R, Fibbe WE. Mesenchymal stem cells inhibit generation and function of both CD34+-derived and monocyte-derived dendritic cells. J Immunol. 2006;177:2080–2087. doi: 10.4049/jimmunol.177.4.2080. [DOI] [PubMed] [Google Scholar]
- 34.Jiang XX, Zhang Y, Liu B, Zhang SX, Wu Y, Yu XD, Mao N. Human mesenchymal stem cells inhibit differentiation and function of monocyte-derived dendritic cells. Blood. 2005;105:4120–4126. doi: 10.1182/blood-2004-02-0586. [DOI] [PubMed] [Google Scholar]
- 35.Aggarwal S, Pittenger MF. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood. 2005;105:1815–1822. doi: 10.1182/blood-2004-04-1559. [DOI] [PubMed] [Google Scholar]
- 36.Beyth S, Borovsky Z, Mevorach D, Liebergall M, Gazit Z, Aslan H, Galun E, Rachmilewitz J. Human mesenchymal stem cells alter antigen-presenting cell maturation and induce T-cell unresponsiveness. Blood. 2005;105:2214–2219. doi: 10.1182/blood-2004-07-2921. [DOI] [PubMed] [Google Scholar]
- 37.Maccario R, Podesta M, Moretta A, Cometa A, Comoli P, Montagna D, Daudt L, Ibatici A, Piaggio G, Pozzi S, Frassoni F, Locatelli F. Interaction of human mesenchymal stem cells with cells involved in alloantigen-specific immune response favors the differentiation of CD4+ T-cell subsets expressing a regulatory/suppressive phenotype. Haematologica. 2005;90:516–525. [PubMed] [Google Scholar]
- 38.Zhang W, Ge W, Li C, You S, Liao L, Han Q, Deng W, Zhao RC. Effects of mesenchymal stem cells on differentiation, maturation, and function of human monocyte-derived dendritic cells. Stem Cells Dev. 2004;13:263–271. doi: 10.1089/154732804323099190. [DOI] [PubMed] [Google Scholar]
- 39.Chiesa S, Morbelli S, Morando S, Massollo M, Marini C, Bertoni A, Frassoni F, Bartolome ST, Sambuceti G, Traggiai E, Uccelli A. Mesenchymal stem cells impair in vivo T-cell priming by dendritic cells. Proc Natl Acad Sci USA. 2011;108:17384–17389. doi: 10.1073/pnas.1103650108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Spaggiari GM, Abdelrazik H, Becchetti F, Moretta L. MSCs inhibit monocyte-derived DC maturation and function by selectively interfering with the generation of immature DCs: central role of MSC-derived prostaglandin E2. Blood. 2009;113:6576–6583. doi: 10.1182/blood-2009-02-203943. [DOI] [PubMed] [Google Scholar]
- 41.Wehner R, Wehrum D, Bornhauser M, Zhao S, Schakel K, Bachmann MP, Platzbecker U, Ehninger G, Rieber EP, Schmitz M. Mesenchymal stem cells efficiently inhibit the proinflammatory properties of 6-sulfo LacNAc dendritic cells. Haematologica. 2009;94:1151–1156. doi: 10.3324/haematol.2008.001735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bartholomew A, Sturgeon C, Siatskas M, Ferrer K, McIntosh K, Patil S, Hardy W, Devine S, Ucker D, Deans R, Moseley A, Hoffman R. Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp Hematol. 2002;30:42–48. doi: 10.1016/s0301-472x(01)00769-x. [DOI] [PubMed] [Google Scholar]
- 43.Krampera M, Glennie S, Dyson J, Scott D, Laylor R, Simpson E, Dazzi F. Bone marrow mesenchymal stem cells inhibit the response of naive and memory antigen-specific T cells to their cognate peptide. Blood. 2003;101:3722–3729. doi: 10.1182/blood-2002-07-2104. [DOI] [PubMed] [Google Scholar]
- 44.Maitra B, Szekely E, Gjini K, Laughlin MJ, Dennis J, Haynesworth SE, Koc ON. Human mesenchymal stem cells support unrelated donor hematopoietic stem cells and suppress T-cell activation. Bone Marrow Transplant. 2004;33:597–604. doi: 10.1038/sj.bmt.1704400. [DOI] [PubMed] [Google Scholar]
- 45.Gonzalez-Rey E, Gonzalez MA, Varela N, O'Valle F, Hernandez-Cortes P, Rico L, Buscher D, Delgado M. Human adipose-derived mesenchymal stem cells reduce inflammatory and T cell responses and induce regulatory T cells in vitro in rheumatoid arthritis. Ann Rheum Dis. 2010;69:241–248. doi: 10.1136/ard.2008.101881. [DOI] [PubMed] [Google Scholar]
- 46.Locatelli F, Maccario R, Frassoni F. Mesenchymal stromal cells, from indifferent spectators to principal actors. Are we going to witness a revolution in the scenario of allograft and immune-mediated disorders? Haematologica. 2007;92:872–877. doi: 10.3324/haematol.11479. [DOI] [PubMed] [Google Scholar]
- 47.Krampera M, Cosmi L, Angeli R, Pasini A, Liotta F, Andreini A, Santarlasci V, Mazzinghi B, Pizzolo G, Vinante F, Romagnani P, Maggi E, Romagnani S, Annunziato F. Role for interferon-gamma in the immunomodulatory activity of human bone marrow mesenchymal stem cells. Stem Cells. 2006;24:386–398. doi: 10.1634/stemcells.2005-0008. [DOI] [PubMed] [Google Scholar]
- 48.Mougiakakos D, Jitschin R, Johansson CC, Okita R, Kiessling R, Le Blanc K. The impact of inflammatory licensing on heme oxygenase-1-mediated induction of regulatory T cells by human mesenchymal stem cells. Blood. 2011;117:4826–4835. doi: 10.1182/blood-2010-12-324038. [DOI] [PubMed] [Google Scholar]
- 49.Ren G, Zhao X, Zhang L, Zhang J, L'Huillier A, Ling W, Roberts AI, Le AD, Shi S, Shao C, Shi Y. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J Immunol. 2010;184:2321–2328. doi: 10.4049/jimmunol.0902023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Corcione A, Benvenuto F, Ferretti E, Giunti D, Cappiello V, Cazzanti F, Risso M, Gualandi F, Mancardi GL, Pistoia V, Uccelli A. Human mesenchymal stem cells modulate B-cell functions. Blood. 2006;107:367–372. doi: 10.1182/blood-2005-07-2657. [DOI] [PubMed] [Google Scholar]
- 51.Tabera S, Perez-Simon JA, Diez-Campelo M, Sanchez-Abarca LI, Blanco B, Lopez A, Benito A, Ocio E, Sanchez-Guijo FM, Caaizo C, San Miguel JF. The effect of mesenchymal stem cells on the viability, proliferation and differentiation of B-lymphocytes. Haematologica. 2008;93:1301–1309. doi: 10.3324/haematol.12857. [DOI] [PubMed] [Google Scholar]