

Abstract
We investigated whether time courses of fMRI BOLD activity in recollection-sensitive brain regions varied according to the time over which recollected information was maintained. Human subjects studied word-picture pairs and were subsequently tested with studied and unstudied pictures during a scanned test phase. The test requirement was to judge whether each picture was old or new and, if old, to retrieve its study associate and hold it in mind until a response cue appeared. The interval between the test item and cue varied between 2 and 8 seconds. Separate responses were required when items were deemed new or the associate was not retrieved. Whereas recollection-related activity in the posterior cingulate, medial temporal, and medial prefrontal cortices was transient and unrelated to the maintenance interval, activity in the left anterior angular gyrus (aLAG) tracked the interval. Thus, as in a prior study, recollection-sensitive regions could be temporally dissociated.
Keywords: angular gyrus, episodic memory, hippocampus, recollection, working memory
Introduction
Successful recollection engages a network of brain regions that includes the hippocampus, left posterior parietal cortex, medial prefrontal cortex (mPFC), parahippocampal cortex, and retrosplenial/posterior cingulate cortex. These regions appear to be engaged regardless of the nature of the study and test materials employed or the type of retrieval test (see Rugg and Vilberg, 2013; Kim, 2010). This ‘core’ recollection network is thought to support cognitive processes contributing to successful recollection, but the specific roles played by the different regions comprising the network have not been fully elucidated. In a previous study (Vilberg and Rugg, 2012), we evaluated whether recollection-related activity in different regions of the network differed in their sensitivity to the duration of an interval over which recollected information was to be maintained. A dissociation within the core recollection network was evident between the medial temporal and retrosplenial cortices, where recollection-related activity was transient and independent of the duration of the maintenance interval, and the left angular gyrus (LAG), where recollection-related activity was sustained across the maintenance interval. This latter finding is consistent with proposals that the LAG contributes in some way to the representation of recollected content in a consciously accessible form (see Vilberg and Rugg, 2008).
In the present study, we sought to extend our prior findings to another class of recollected information. Whereas in the prior study participants recollected previously paired pictures in response to word cues, here, participants were cued with pictures to recollect previously paired words. As in our prior study, retrieved content had to be maintained over a variable delay prior to making a judgment about the recollected associate. By varying the maintenance interval as well as the inter-trial interval, we were able differentiate regions demonstrating transient and sustained recollection effects. At issue is whether our prior findings were material-selective or whether, consistent with the proposal that the core recollection network is material- and content-independent, a dissociation between transient recollection effects in medial temporal and midline cortical regions, and sustained effects in the LAG, is still evident. This issue is particularly important because it is not possible, based on our prior findings alone, to adjudicate between effects selective for pictures of objects, and effects that generalize to other materials. We adopted a similar strategy in our prior studies of the relationship between LAG recollection effects and amount of information retrieved (cf. Vilberg & Rugg, 2007; 2009).
Materials and Methods
Subjects
Twenty four (12 female) right-handed, native English speakers, aged 18 to 29 years (mean = 23), participated. Subjects reported themselves to be free of neurological and psychiatric illness and were screened for contraindications for MRI. All subjects gave informed consent prior to participating in the study, which had been approved by the Institutional Review Boards of UT Southwestern and UT Dallas. Two subjects’ data were excluded because they contributed insufficient trials (< 10) to conditions of interest. The results reported below utilize data from the 22 remaining subjects.
Experimental materials and procedure
The experiment consisted of 4 study-test cycles, with fMRI data acquired during each test block. Figure 1 shows a schematic of the structure of the study and test blocks, as well as the timing of trials. Study stimuli were the same word-picture pairs employed in Vilberg and Rugg (2012). Pictures were presented in color on a grey background and all text was presented in black Helvetica font. Test items comprised single pictures. For each subject, 120 word-picture pairs were randomly assigned to serve as study items (30 pairs per block), and an additional 60 pictures were randomly assigned to be used as new test items (15 words per block). Items were displayed on a screen positioned at the back of the scanner bore visible via a mirror attached to the head coil.
Figure 1.
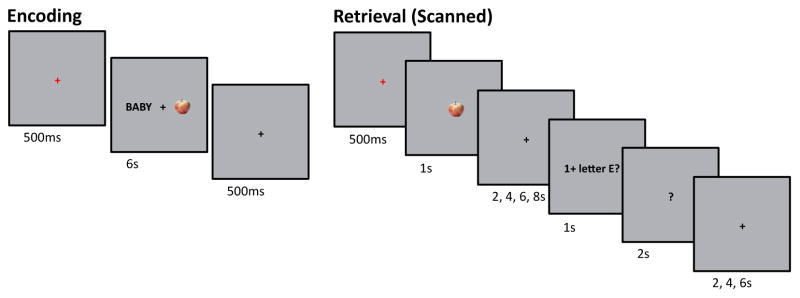
Trial schematics are shown for a single retrieval trial (top) and a single encoding trial (bottom).
Prior to performing the first study block, subjects performed practice versions of the study and test tasks outside the scanner (see Vilberg and Rugg, 2012 for details of the practice session). Responses during study and test blocks in the scanner were made using MRI-compatible button boxes.
At study, the task requirement was to signal with a button-press which of the two denoted objects was the smaller. Participants were instructed to respond prior to the onset of the red fixation character signaling the beginning of the next trial. Each study block consisted of 30 trials. For each test picture, participants were instructed to decide whether it was old or new, and if old, to try to recollect and “hold in mind” the visual form of the word that had been paired with the picture at study until queried about one of three possible features of the word. Following the presentation of the picture, a cue appeared on the screen after a variable delay period. The cue indicated which of three questions the subject should answer regarding an attribute of the word associated with the test picture, should it have been recollected. The three cues were ‘1+ letter E’, ‘2+ syllables’, and ‘Alphabetical’, representing the following questions, respectively: Does the recollected word have one or more Es in it? Does the recollected word have two or more syllables? Are the first and last letters of the recollected word in alphabetical order? By cueing these three different questions in an unpredictable sequence we aimed to prevent the adoption of a strategy whereby subjects merely classified the words as soon as they were recollected and then maintained the appropriate response over the delay period instead of a representation of the recollected word. If the test picture was judged new, the new key was to be pressed. If the test picture was judged old but the associated study word could not be recollected, the ‘old don’t remember’ key was to be pressed. If the test picture was judged old and the study word was recollected, participants were to press the key corresponding to a yes or no judgment to indicate their answer to the question posed by the cue. Subjects were instructed to withhold responding until the appearance of the cue but to make a response prior to the offset of the question mark. Hand assignment was counterbalanced such that half of all participants used their left hand to answer the attribute question and the other half used their right hand.
A short break (< 5 min) was given between each study-test cycle. The ordering of the test trials was pseudo-randomized for each subject such that no more than 4 old or new trials occurred in succession. The durations of the intervals between presentation of test items and response cues were randomly assigned to old and new trials separately for each subject such that approximately equal proportions of each duration (2, 4, 6, and 8s) occurred for each of these trial types.
MRI data acquisition and analysis
MRI data were acquired and preprocessed using the same procedures described in Vilberg and Rugg (2012). Functional analysis was performed using a General Linear Model (GLM) in which a delta function was used to model neural activity at item onset (item-related activity). Delay-related activity was modeled with a boxcar that onset concurrently with the fixation character that followed presentation of the test picture. The boxcar varied in length with the duration of the fixation character (2, 4, 6, or 8s following the offset of the test item). The delta and boxcar functions were each convolved with a canonical hemodynamic response function (HRF) to model the predicted BOLD response. Four event types were modeled: Associative hits (old trials for which a correct response was given to the cue terminating the trial), item hits (old trials for which an ‘old don’t remember’ response was given), correct rejections (new trials for which a new response was given), and events of no interest. The model thus contained 4 event types for each of the item- and delay-related regressors, for a total of 8 event types in the model. The model also included as covariates six regressors representing motion-related variance (three for rigid-body translation and three for rotation), regressors modeling the separate scan sessions, and the across-scan mean.
The subject-specific item and delay parameter estimates for associative hits, item hits and correct rejections were taken forward to two-way repeated measures ANOVA as implemented in SPM, employing the factors of type of trial type and regressor (item versus delay). Unless otherwise noted, contrasts derived from the ANOVA model were thresholded at p < 0.001, uncorrected, with a 26 voxel extent threshold (the cluster extent threshold giving a corrected whole-brain cluster-wise significance level of p < .05 on the basis of a Monte Carlo simulation using 10,000 iterations of randomized data; http://afni.nimh.gov/afni). Exclusive masks were thresholded at p < .05 one-tailed. Peak coordinates of significant effects are reported in MNI space. In addition to the previously described GLM, a finite impulse response (FIR) model was employed to estimate the time courses of the responses elicited by the three event types of interest, with each maintenance delay modeled separately for each condition. The details of this model are described in Vilberg and Rugg (2012), where an identical procedure was employed. Finally, to directly compare the present results with those of our prior study, we employed an additional ANOVA model using the same event types as in the original GLM, but with experiment as an across-subjects factor.
Results
Behavioral results
The mean old item hit rate was 90.2%, against a mean correct rejection rate of 86.5%. The proportions of old and new items attracting each class of response and their associated response times (RTs) with respect to cue onset are given in Table 1. The mean numbers of associative hit, item hit, and correct rejection trials were 45, 49, and 50 respectively, with ranges of 11–86, 18–71, and 24–60, respectively. A repeated-measures ANOVA on test RTs (associative hits, item hits, and correct rejections) revealed a main effect of response type, F(2, 41.1) = 140.72, p < .001. Subsequent pair-wise comparisons revealed that correct rejections were significantly faster than associative hits and item hits, F(1, 21) = 310.29 and 42.56, each p < .001, respectively. Additionally, associative hits were associated with longer RTs than item hits, F(1, 21) = 91.38. p < .001.
Table 1.
Response proportions and RTs for old and new trials
| Response | Proportions | RTs | ||
|---|---|---|---|---|
| Old | New | Old | New | |
| Yes/No correct | .39 | ---- | 1716 (331) | ---- |
| Yes/No incorrect | .09 | .01 | 2006 (447) | 2369 (743) |
| Old don’t remember | .42 | .06 | 1117 (359) | 1366 (643) |
| New | .06 | .87 | 886 (356) | 742 (177) |
Note: Response proportion columns do not sum to 1.00 due to exclusion of response omissions and trials upon which multiple key presses were made. RTs are displayed in milliseconds, with standard deviations in parentheses.
fMRI results
Transient recollection effects were identified in a two-stage procedure. First, using the error term from the ANOVA model described in the Methods section, we employed the item regressor to identify regions that demonstrated greater activity for associative hits than for item hits. Then, we exclusively masked the outcome of this contrast with the corresponding contrast for the delay regressor. Thus, the outcome of this procedure identified clusters where recollection effects were transient (exclusively associated with the item regressor). Sustained effects were identified by the associative hit > item hit contrast for the delay regressor alone.
Regions demonstrating transient recollection effects were confined to medial prefrontal, posterior cingulate, and left and right parahippocampal cortices. Sustained recollection effects included clusters in bilateral inferior frontal gyrus, right middle frontal gyrus, left inferior temporal gyrus, striatum, midbrain, and bilateral superior/inferior parietal cortex, where they extended from the intraparietal sulcus into the AG. Figure 2 illustrates these two classes of effects.
Figure 2.

Top: Regions showing transient recollection effects identified by the GLM analysis. Bottom: Regions showing sustained recollection effects are displayed. Effects are projected onto SPM’s single subject rendered brain and are also displayed on sections of the mean across-subjects normalized anatomical image. Mean parameter estimates for two representative regions are plotted across time from −2 to 10s from test item onset. The (MNI) coordinates of the individual voxels from which time courses were derived are (A) −27, −34, −14, and (B) −33, −67, 46. These FIR-derived time courses were extracted separately for each maintenance interval trial type, and, prior to collapsing across maintenance interval, were truncated at 8s, 10s, 12s, and 14s post item onset (for 2, 4, 6, and 8s maintenance interval trials, respectively) to avoid potential contamination of the time course data with BOLD activity associated with the following trial (see Vilberg & Rugg, 2012 where the same procedure was employed). Standard errors are displayed for each estimate. CR = correct rejection.
The present findings seemingly differ from those of our prior study, where transient recollection effects were not evident in mPFC. To directly contrast the two sets of findings we employed a between-experiment ANOVA (see Methods). Regions demonstrating transient effects common to each experiment were identified by inclusively masking the transient recollection effect from each experiment (defined as described above) at p < .01 for each contrast, giving a conjoint significance level of p < .001 (according to Fisher’s procedure). This analysis identified clusters in posterior cingulate cortex, left parahippocampal cortex extending into the hippocampus, right mammillary body, and mPFC (see Figure 3). Regions demonstrating common sustained recollection effects were identified by inclusively masking the associative hit > item hit effect for the delay regressor from each experiment at p < .01. Overlapping regions were extensive, encompassing over 6000 voxels (see Figure 3). Regions demonstrating differences in recollection effects between the two experiments were identified using the F contrast of the interaction between experiment and condition, thresholded at p < .001. For transient effects, the contrast on the item regressor was exclusively masked with the across-experiment main effect for the delay regressor at p < .05. For sustained effects, interactions were identified using the F contrast of the interaction with experiment on the delay regressor alone, thresholded at p < .001. No regions exhibited differential transient recollection effects. There was, however, a 191-voxel cluster in the retrosplenial cortex (x, y, z = −3, −58, 13) where sustained effects differed by experiment. Whereas in the prior study the region exhibited a transient recollection effect, in the current experiment the region exhibited item > associative hit effects at late (8+s) time points.
Figure 3.
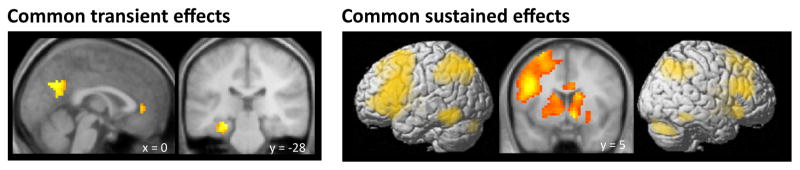
Regions showing transient (top) and sustained (bottom) recollection effects common to both the current and our prior study (Vilberg & Rugg, 2012). Effects are projected onto SPM’s single subject rendered brain and are also displayed on sections of the mean across-subjects normalized anatomical image.
Finally, to address the question whether, as in our prior study, recollection-related activity in the LAG in the present experiment co-varied with maintenance interval (see Introduction) we extracted the mean parameter estimates from two anatomically defined regions of interest (ROIs) encompassing anterior (aLAG) and posterior (pLAG) aspects of the LAG (see Figure 4, top). As in our prior study (see Vilberg & Rugg, 2012), the ROIs were generated using Anatomy toolbox v1.8 (Eickhoff, Stephan, Mohlberg, Grefkes, Fink, Amunts, & Zilles, 2005). Data were extracted for each participant for the different delay conditions and trial types using the FIR model (see Methods). Figure 4 (bottom) illustrates mean across-ROI time courses, truncated as described in Figure 2 and collapsed across the four maintenance interval durations. We subjected the associative hit and item hit time course data to an ANOVA, with factors of condition (associative hits vs. associative miss), time point (0 to 10s), and region (anterior/posterior). The ANOVA revealed significant main effects of each factor, as well as interactions between region and condition, and region and time point (each F > 4.00, p < .05). Crucially, the analysis also gave rise to a three way interaction, F(2.4, 49.4) = 10.97, p < .001. Follow-up pairwise comparisons at each time point for each region revealed significantly greater activity for associative than item hits for aLAG from 4s to 10s following picture onset, each one-tailed p < .01. For pLAG, associative hit > item hit effects were only apparent at the 4 and 6s post-picture onset time points, each one-tailed p < .05.
Figure 4.

Top: Left anterior (red; aLAG) and posterior (blue; pLAG) angular gyrus ROIs are depicted on sections of the mean across-subjects normalized anatomical image. Bottom: Parameter estimates averaged across all voxels within each ROI are shown from 2s prior to picture onset. CR = correct rejection. ** p < .025, * p < .05.
General Discussion
The aim of the present study was to dissociate regions belonging to the core recollection network according to the time-courses of their retrieval-related activity, building on the findings of our prior study (Vilberg and Rugg, 2012). As in that study, where recollected items were pictures rather than words, activity in regions of the core recollection network dissociated temporally. Whereas medial temporal and midline cortical regions demonstrated transient recollection effects, activity in the aLAG tracked the interval over which recollected information was maintained. Thus, the temporal dissociation observed in our prior study does not depend upon retrieval of pictorial information, but instead generalizes to the retrieval of concrete words.
Although not all regions of the core recollection network survived the statistical thresholds applied in the present and our prior studies separately, a combined analysis revealed transient recollection effects common to both experiments in the mPFC, retrosplenial cortex, and left parahippocampal gyrus (see Figure 3). The only region of the core recollection network exhibiting a common sustained effect was the aLAG. Together, these results testify to the robustness of the temporal dissociation that exists between medial temporal and midline components of the network and its sole lateral cortical component, the LAG.
Several proposals for the role of the hippocampus in episodic memory retrieval converge on the idea that the region is responsible for the reinstatement of patterns of cortical activity engaged at the time of initial encoding (e.g., Alvarez and Squire, 1994; Norman and O’Reilly, 2003). Our findings that the hippocampus demonstrated transient recollection effects are compatible with these models, suggesting that the role of this structure is limited to the retrieval of information and does not extend to its subsequent maintenance (see also Ranganath, Cohen, and Brozinsky, 2005). Other regions of the core recollection network demonstrating transient effects are each strongly interconnected with the hippocampus, and the similar patterns of activity in these regions likely reflects the fact that they form a functional network that, along with the hippocampus, supports episodic memory retrieval (Aggleton, 2012).
To date, little work has addressed the question of how reactivated mnemonic content is maintained in a consciously accessible form. This issue is of particular importance given that the adaptive use of retrieved episodic information relies on the ability to maintain and integrate the retrieved content with other information to inform decision-making processes. Given previous findings of task- and material-general recollection effects in LAG (Kim, 2010), and our prior findings that activity in this region tracks a delay interval, the region seems likely to play a generic role in support of the maintenance of recollected content. There are at least two plausible accounts for how LAG might function in this respect. The accounts differ primarily in how interactions between episodic and working memory are characterized. One account proposes that accessibility of recollected content is a consequence of the role played by LAG in integrating neural activity across distributed cortical sites where the different features of a retrieved episode are represented (for a similar proposal with respect to semantic memory, see Binder and Desai, 2011). Alternatively, LAG may play a more direct representational role, perhaps supporting a consciously accessible ‘buffer’ in which recollected information is re-represented (cf. Baddeley, 2000). Current evidence does not allow adjudication between these two classes of accounts.
Unlike our prior study, here activity in the aLAG and pLAG dissociated with respect to their time-courses, with only the anterior sub-region showing a sustained effect (Figure 4). This finding raises the possibility that LAG recollection effects may be material-sensitive, although it is also possible that the finding reflects a correlated factor such as the amount of information that had to be retained to satisfy task demands (cf. Vilberg and Rugg, 2007). Further work is necessary to evaluate the extent to which sustained recollection effects in LAG vary according to material and richness of retrieved content.
Several regions that are thought to fall outside of the core recollection network also demonstrated sustained recollection-related activity. One of these regions, left lateral inferior temporal cortex (LITC; see Figure 2), was identified as consistently demonstrating recollection effects across studies in a recent meta-analysis (Kim, 2010), and demonstrated sustained effects in our prior study also. In view of evidence suggesting that this region supports conceptual representations of concrete objects (e.g., Martin and Chao, 2001; Binder and Desai, 2011), we have been reluctant to grant it membership to the core recollection network. Rather, we conjecture that the consistency with which recollection effects have been identified in this region reflects the popularity of objects and concrete words as experimental items. From this perspective, the sustained activity identified in LITC in the present and our prior study may reflect the role of the region in supporting conceptual representations of recollected items over the maintenance interval. Nonetheless, future studies that address the question of whether recollection-related activity in this region is indeed restricted to retrieval of concrete information would be of value.
Finally, as was also the case in our prior study, sustained recollection effects were evident in bilateral dorsal parietal and lateral prefrontal cortex, as well as in the striatum. This fronto-parietal network has been identified in numerous prior studies that have employed delayed-match-to-sample and similar tasks to investigate working memory (e.g., Riggall and Postle, 2012), and we assume that its engagement in the present study reflects the similar demands placed upon fronto-parietally mediated attentional and control processes by the current task and tasks employed in the study of working memory.
In summary, the present findings converge with prior results (Vilberg and Rugg, 2012) to demonstrate that retrieval-related activity in the different members of the core recollection network can be temporally dissociated. A major challenge for the future will be to elucidate the specific functional roles played by these different regions, and the nature of their interactions with the disparate cortical areas within which recollected content is reinstated. Additional studies are also necessary to elucidate the extent to which the neural correlates of the maintenance of recollected content overlap with those associated with the maintenance of information directly presented to participants.
Acknowledgments
This research was supported by NIMH grant 5R01MH072966.
References
- Aggleton JP. Multiple anatomical systems embedded within the primate medial temporal lobe: Implications for hippocampal function. Neuroscience & Biobehavioral Reviews. 2012;36:1579–96. doi: 10.1016/j.neubiorev.2011.09.005. [DOI] [PubMed] [Google Scholar]
- Alvarez P, Squire LR. Memory consolidation and the medial temporal lobe: a simple network model. Proceedings of the National Academy of Sciences: USA. 1994;91:7041–5. doi: 10.1073/pnas.91.15.7041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baddeley A. The episodic buffer: A new component of working memory? Trends in Cognitive Science. 2000;4:417–23. doi: 10.1016/s1364-6613(00)01538-2. [DOI] [PubMed] [Google Scholar]
- Binder JR, Desai RH. The neurobiology of semantic memory. Trends in Cognitive Sciences. 2011;15:527–36. doi: 10.1016/j.tics.2011.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eickhoff S, Stephan KE, Mohlberg H, Grefkes C, Fink GR, Amunts K, Zilles K. A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. NeuroImage. 2005;25:1325–35. doi: 10.1016/j.neuroimage.2004.12.034. [DOI] [PubMed] [Google Scholar]
- Kim H. Dissociating the roles of the default-mode, dorsal, and ventral networks in episodic memory retrieval. NeuroImage. 2010;50:1648–57. doi: 10.1016/j.neuroimage.2010.01.051. [DOI] [PubMed] [Google Scholar]
- Martin A, Chao LL. Semantic memory and the brain: structure and processes. Current Opinion in Neurobiology. 2001;11:194–201. doi: 10.1016/s0959-4388(00)00196-3. [DOI] [PubMed] [Google Scholar]
- Norman KA, O’Reilly RC. Modeling hippocampal and neocortical contributions to recognition memory: A complementary-learning-systems approach. Psychological Review. 2003;110:611–46. doi: 10.1037/0033-295X.110.4.611. [DOI] [PubMed] [Google Scholar]
- Ranganath C, Cohen MX, Brozinsky CJ. Working memory maintenance contributes to long-term memory formation: neural and behavioral evidence. Journal of Cognitive Neuroscience. 2005;17:994–1010. doi: 10.1162/0898929054475118. [DOI] [PubMed] [Google Scholar]
- Nyberg L. Any novelty in hippocampal formation and memory? Current Opinion in Neurobiology. 2005;18:424–28. doi: 10.1097/01.wco.0000168080.99730.1c. [DOI] [PubMed] [Google Scholar]
- Riggall AC, Postle BR. The relationship between working memory storage and elevated activity as measured with functional magnetic resonance imaging. Journal of Neuroscience. 2012;32:12990–8. doi: 10.1523/JNEUROSCI.1892-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rugg MD, Vilberg KL. Brain networks underlying episodic memory retrieval. Current Opinion in Neurobiology. 2013;23:255–60. doi: 10.1016/j.conb.2012.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilberg KL, Rugg MD. Dissociation of the neural correlates of recognition memory according to familiarity, recollection, and amount of recollected information. Neuropsychologia. 2007;45:2216–25. doi: 10.1016/j.neuropsychologia.2007.02.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilberg KL, Rugg MD. Memory retrieval and the parietal cortex: A review of evidence from a dual-process perspective. Neuropsychologia. 2008;46:1787–99. doi: 10.1016/j.neuropsychologia.2008.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilberg KL, Rugg MD. Left parietal cortex is modulated by amount of recollected verbal information. Neuro Report. 2009;20:1295–9. doi: 10.1097/WNR.0b013e3283306798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilberg KL, Rugg MD. The neural correlates of recollection: transient versus sustained fMRI effects. Journal of Neuroscience. 2012;32:15679–87. doi: 10.1523/JNEUROSCI.3065-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]