

Abstract
Purpose
The transcription factor Forkhead box M1 (FOXM1) plays critical roles in cancer development and progression. However, the regulatory role and underlying mechanisms of FOXM1 in cancer metabolism are unknown. In this study, we characterized the regulation of aerobic glycolysis by FOXM1 and its impact on pancreatic cancer metabolism.
Experimental Design
The effect of altered expression of FOXM1 on expression of glycolytic enzymes and tumor development and progression was examined using animal models of pancreatic cancer. Also, the underlying mechanisms of altered pancreatic cancer glycolysis were analyzed using in vitro molecular biology. The clinical relevance of aberrant metabolism caused by dysregulated FOXM1 signaling was determined using pancreatic tumor and normal pancreatic tissue specimens.
Results
We found that FOXM1 did not markedly change the expression of most glycolytic enzymes except for phosphoglycerate kinase 1 and lactate dehydrogenase A (LDHA). FOXM1 and LDHA were overexpressed concomitantly in pancreatic tumors and cancer cell lines. Increased expression of FOXM1 upregulated the expression of LDHA at both the mRNA and protein level and elevated LDH activity, lactate production, and glucose utilization, whereas reduced expression of FOXM1 did the opposite. Further studies demonstrated that FOXM1 bound directly to the LDHA promoter region and regulated the expression of the LDHA gene at the transcriptional level. Also, elevated FOXM1-LDHA signaling increased the pancreatic cancer cell growth and metastasis.
Conclusions
Dysregulated expression and activation of FOXM1 play important roles in aerobic glycolysis and tumorigenesis in pancreatic cancer patients via transcriptional regulation of LDHA expression.
Keywords: Metabolism, invasion, metastasis, transcription factor, signaling pathway
Introduction
Pancreatic cancer is the seventh leading cause of cancer-related deaths worldwide, and its incidence is increasing annually, especially in industrialized countries (1). In the United States, researchers estimated that 45,220 new pancreatic cancer cases would be diagnosed and that 38,460 patients would die of the disease in 2013 (2). Although early diagnosis of and surgery and systemic chemotherapy for pancreatic cancer have improved, the overall 5-year survival rate in pancreatic cancer patients remains below 5% (3). Thus, a better understanding of the underlying mechanisms that promote the pathogenesis of pancreatic cancer is urgently needed (4).
The Warburg effect, also known as aerobic glycolysis, is a shift from oxidative phosphorylation to glycolysis, a feature of which is increased lactate production even at normal oxygen concentrations, and is considered to be the root of cancer development and progression (5,6). In pancreatic cancer cases, investigators found that glycolytic flux was elevated and that uptake of glucose increased; thus, 18F-labeled fluorodeoxyglucose positron emission tomography can be used for diagnosis of and prognosis for pancreatic cancer (7,8). However, the molecular mechanism of increased glycolysis in pancreatic cancer cells is largely unknown. Recent studies demonstrated that many key oncogenic signaling pathways and factors play critical roles in the regulation of pancreatic cancer metabolism, including Kras, phosphoinositide 3-kinase/AKT, c-Myc, p53, and hypoxia-inducible factor-1 (9).
Forkhead box protein M1 (FOXM1) is an oncogenic transcription factor belonging to the Forkhead transcription factor superfamily. Human FOXM1 has three isoforms—FOXM1A, FOXM1B, and FOXM1C—as a result of differential splicing of exons Va and VIIa. FOXM1B and FOXM1C are transcriptionally active, whereas FOXM1A is transcriptionally inactive (10). FOXM1 plays essential roles in regulation of the cancer cell cycle, angiogenesis, invasion, and metastasis by directly promoting its target gene expression and networking with other factors (11). In previous studies, we found that FOXM1 was overexpressed in pancreatic cancer cells and promoted pancreatic cancer development and progression (12,13). Researchers also found evidence that FOXM1 participates in regulation of metabolism. For example, in leptin-knockout mice, overexpression of FOXM1 was related to elevated consumption of glucose, whereas low expression of it was not (14). However, the role of FOXM1 expression in pancreatic cancer aerobic glycolysis and the underlying mechanisms are unknown. Therefore, in the present study, we sought to determine the role of FOXM1 in regulation of the Warburg effect and its mechanism regarding this role in pancreatic cancer cases. We discovered that FOXM1B transcriptionally upregulated LDHA expression; increased LDH activity, lactate production, and glucose utilization; and promoted pancreatic tumorigenesis and tumor metastasis.
Materials and Methods
Human tissue specimens and immunohistochemical analysis
Expression of FOXM1 and LDHA was analyzed using two tissue microarrays: one (TMA) containing 57 primary pancreatic ductal adenocacinoma, 10 normal tumor-adjacent pancreatic tissue, and 10 normal pancreatic tissue specimens (US Biomax) and the other (TMA-P) containing 154 human pancreatic ductal adenocacinoma specimens obtained from the Pancreatic Cancer Tissue Bank at Shanghai Jiaotong University Affiliated First People’s Hospital (Shanghai, People’s Republic of China). The primary pancreatic cancers in the patients represented in TMA-P were diagnosed and later confirmed by at least two pathologists, and the patients were accepted for the patients underwent surgery at Affiliated First People’s Hospital, Jiangsu Province Hospital (Jiangsu, People’s Republic of China) and Shanghai East Hospital (Shanghai, People’s Republic of China) from 2004 to 2011. Tumor staging for the specimens was carried out using the American Joint Committee on Cancer staging criteria. The use of human specimens was approved by the proper institutional review boards. After hematoxylin and eosin staining of slides containing sections of optimal pancreatic tumor, normal tumor-adjacent pancreatic tissue up to 2 cm from the tumor, and normal pancreatic tissue up to 5 cm from the tumor, TMA-P slides were constructed (in collaboration with Shanghai Outdo Biotech, Shanghai, People’s Republic of China). Two punch cores 2 mm in greatest dimension were taken from nonnecrotic areas of each formalin-fixed, paraffin-embedded tumor, matched normal tumor-adjacent tissue, and normal tissue specimen. Primary tumor and matched normal tumor-adjacent tissue specimens obtained from 34 of these 154 patients were included in TMA-P; primary tumor and matched normal tumor-adjacent and normal tissue specimens from 22 of these patients were included. Sections (4-µm thick) of formalin-fixed, paraffin-embedded tumor specimens were prepared and processed for immunostaining using anti-FOXM1 (Santa Cruz Biotechnology) and anti-LDHA (Santa Cruz Biotechnology) antibodies. The staining results were scored by two investigators blinded to the clinical data as described previously (15).
Cell lines and reagents
The human pancreatic adenocarcinoma cell lines PANC-1, MiaPaCa-2, AsPC-1, BxPC-3, and PA-TU-8902 were purchased from the American Type Culture Collection. The pancreatic cancer cell line MDA Panc-28 was a gift from Dr. Paul J. Chiao (The University of Texas MD Anderson Cancer Center, Houston, TX). The human pancreatic adenocarcinoma cell line FG was obtained from Michael P. Vezeridis (The Warren Alpert Medical School of Brown University, Providence, RI) (16). The human metastatic pancreatic adenocarcinoma cell line COLO357 and its fast-growing liver-metastatic variant in nude mice, L3.7, were described previously (12). All of these cell lines were maintained in plastic flasks as adherent monolayers in Eagle's minimal essential medium supplemented with 10% fetal bovine serum (FBS), sodium pyruvate, nonessential amino acids, L-glutamine, and a vitamin solution (Flow Laboratories). PANC-1, MiaPaCa-2, AsPC-1, BxPC-3, and PA-TU-8902 were characterized or authenticated by the American Type Culture Collection using short tandem repeat profiling and passaged in our laboratory for fewer than 6 months after receipt. A competitive LDHA inhibitor, oxamate sodium, was purchased from Sigma-Aldrich (17).
Plasmids and small interfering RNAs
The plasmid pcDNA3.1-FOXM1 (pcDNA3.1-FOXM1B) and control vector pcDNA3.1 were described previously (18). A small interfering RNA (siRNA) targeting FOXM1 (siFOXM1) consisted of a pool of three target-specific 20- to 25-nt siRNAs designed to knock down FOXM1 expression was obtained from Santa Cruz Biotechnology.
Transient transfection
Transfection of plasmids and siRNAs into pancreatic cancer cells was performed using the transfection reagents Lipofectamine LTX and Lipofectamine 2000 CD, respectively (Invitrogen). For transient transfection, cells were transfected with plasmids or siRNAs at different concentrations as indicated for 48 hours before the performance of functional assays. Pancreatic cancer cells treated with the transfection reagents alone were included as mock controls.
Reverse transcription-polymerase chain reaction
Total RNA was extracted from tumor cells using a PureLink RNA Mini Kit (Life Technologies). Next, 2 µg of total RNA was reverse-transcribed using a First Strand cDNA Synthesis Kit (Promega) to synthesize cDNA specimens. Subsequently, 2 µl of the cDNA product was subjected to polymerase chain reaction (PCR) amplification with Taq DNA polymerase (Qiagen) using a thermal cycler with PCR primers to detect each factor (Supplementary Table S1). β-actin and α-tubulin were used as internal controls. The PCR products were loaded onto 2% agarose gels and visualized using ethidium bromide under ultraviolet light.
Western blot analysis
Standard Western blotting was carried out using whole-cell protein lysates and primary antibodies against FOXM1 and LDHA (Santa Cruz Biotechnology) and LDHB (Abcam) and a secondary antibody (anti-rabbit IgG or anti-mouse IgG; Santa Cruz Biotechnology). Equal protein specimen loading was monitored using an anti-α-tubulin antibody (Oncogene).
LDH activity, lactate production, and glucose utilization assay
Pancreatic tumor cells (1 × 106) were transfected with plasmids and siRNAs and prepared for LDH activity and lactate production assay using a Lactate Dehydrogenase Activity Assay Kit and Lactate Assay Kit (Sigma) according to the manufacturer’s protocol. For glucose utilization assay, tumor cells were transfected with plasmids and siRNAs, and cultures were incubated for 24 hours. The culture media were replaced with phenol-red free RPMI with 1% FBS or phenol-red free RPMI with 1% FBS and 20 mmol/l oxamate sodium in continuous culture for 3 days. Medium specimens were collected each day. Glucose concentrations in the media were measured using a colorimetric glucose assay kit (BioVision) and normalized according to cell number (19).
Construction of LDHA promoter reporter plasmids and mutagenesis
A 1.48-kb fragment containing LDHA 5' sequences from −1330 to +150 bp relative to the transcription initiation site was subcloned into the pGL3-basic vector (Promega). The final full-length reporter plasmid, which contained multiple FOXM1-binding sites, was designated pLuc-LDHA-1480. The deletion mutation reporter for this plasmid, pLuc-LDHA-493, was then generated. Both constructs were verified by sequencing the inserts and flanking regions of the plasmids.
Promoter reporter and dual luciferase assay
Pancreatic cancer cells were transfected with the indicated LDHA promoter reporter, siFOXM1, or expression plasmid. The LDHA promoter activity was normalized via co-transfection of a β-actin/Renilla luciferase reporter containing a full-length Renilla luciferase gene (20). The luciferase activity in the cells was quantified using a dual luciferase assay system (Promega) 24 hours after transfection.
Chromatin immunoprecipitation assay
Tumor cells (2 × 106) were prepared for chromatin immunoprecipitation (ChIP) assay using a ChIP assay kit (Millipore) according to the manufacturer’s protocol. The resulting precipitated DNA specimens were analyzed using PCR to amplify a 341-bp region of the LDHA promoter with the primers 5'-TATCTCAAAGCTGCACTGGGC-3' (forward) and 5'-TGCTGATTCCATTGCCTAGC-3' (reverse) and a 282-bp region of the LDHA promoter with the primers 5'-CTGCAGGAAGCCATGATCA-3' (forward) and 5'-TCCCACTCACAGTGAAGCCT-3' (reverse). The PCR products were resolved electrophoretically on a 2% agarose gel and visualized using ethidium bromide staining.
Animals
Pathogen-free female athymic nude mice were purchased from the National Cancer Institute. The animals were maintained in facilities approved by the Association for Assessment and Accreditation of Laboratory Animal Care International in accordance with the current regulations and standards of the U.S. Department of Agriculture and Department of Health and Human Services.
Tumor growth and metastasis
Pancreatic tumor cells (1 × 106) in 0.1 ml of Hank’s balanced salt solution were injected subcutaneously into the right scapular region of nude mice. The tumor-bearing mice were killed when they became moribund or on day 35 after inoculation, and their tumors were removed and weighed. Also, tumor cells (1 × 106) were injected intravenously into a separate group of mice via the ileocolic vein. The mice were killed on day 21 after inoculation or when they appeared to be moribund. Their livers were then removed, and surface metastases on them were counted after dissection of the livers into their individual lobes. Every surface was examined by two investigators who were unaware of the experimental protocol and scored separately (21).
Statistical analysis
The matching and significance of the patient specimen data were assessed using the Mann-Whitney U Test and Pearson correlation coefficient. A two-tailed χ2 test and the Fisher exact test were used to determine the significance of the differences among the covariates. The significance of the in vitro and in vivo data was determined using the Student t-test (two-tailed) or one-way analysis of variance. In all of the tests, P values less than 0.05 were considered statistically significant. The SPSS software program (version 13.0; IBM Corporation) was used for all statistical analysis.
Results
FOXM1-enhanced glycolysis in pancreatic cancer cells
Aerobic glycolysis is the primary aspect of altered cancer metabolism, so we focused on the effect of FOMX1 regulation of pancreatic cancer metabolism on glycolysis. Recent studies revealed that many key types of oncogenic signaling regulate cancer metabolism by regulating the expression and/or activity of metabolic enzymes. To further investigate the mechanism of FOXM1 regulation of glycolysis, we assessed the effect of FOXM1 on the expression of glycolytic enzymes. We treated PANC-1 cells with siFOXM1 and measured the levels of glycolytic enzyme mRNA expression using reverse transcription (RT)-PCR (Fig. 1A). We found that knockdown of FOXM1 expression did not change the expression of most of the enzymes except for phosphoglycerate kinase 1 (PGK-1) and LDHA.
Figure 1.
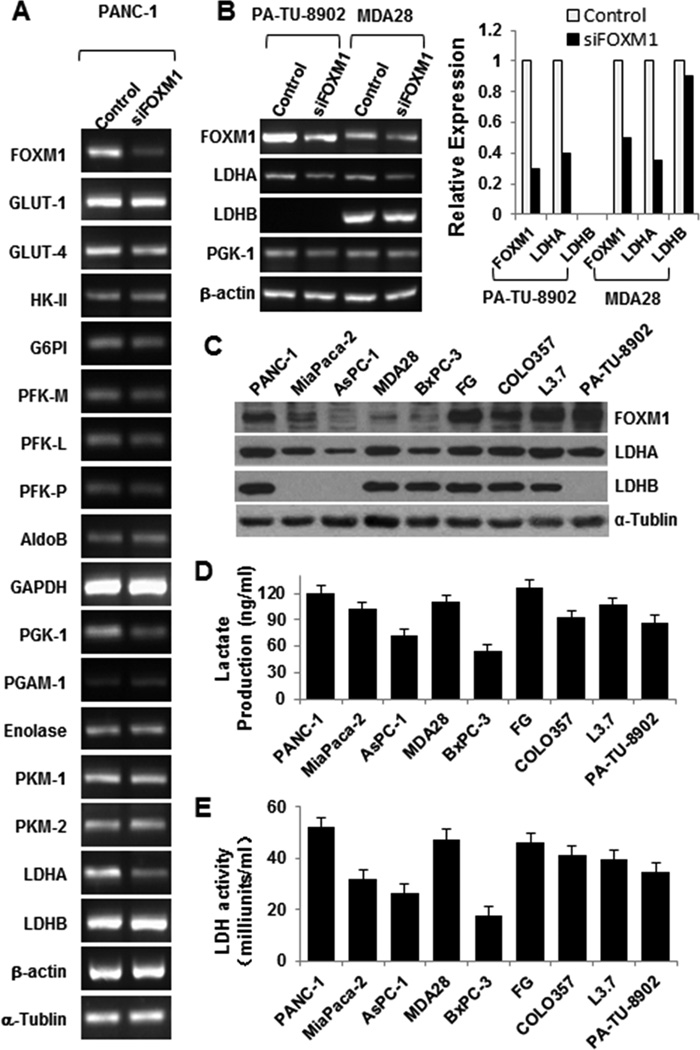
Regulation of the Warburg effect by FOXM1. A, RT-PCR analysis of the mRNA expression for glycolytic enzymes in PANC-1 cells after knockdown of FOXM1 expression. Expression of PGK-1 and LDHA was significantly downregulated. β-actin and α-tubulin were used as controls. B, RT-PCR analysis of the mRNA expression for FOXM1, LDHA, LDHB, and PGK-1 in PA-TU-8902 and MDA Panc-28 cells after knockdown of FOXM1 expression. C, Western blot analysis of FOXM1, LDHA, and LDHB expression in pancreatic cancer cells. D, analysis of the lactate production in pancreatic cancer cell lines using a Lactate Assay Kit. E, analysis of the LDH activity in pancreatic cancer cell lines using a Lactate Dehydrogenase Activity Assay Kit.
We then sought to confirm the effect of FOXM1 expression on PGK-1, LDHA, and LDHB mRNA expression in PA-TU-8902, MDA Panc-28, MiaPaCa-2, and COLO357 cells (Fig. 1B; Supplementary Fig. S1). LDHA mRNA expression decreased significantly, and PGK-1 mRNA expression decreased, but LDHB mRNA expression did not change. PGK-1 catalyzes 1,3-bisphosphoglycerate and ADP to form 3-phosphoglycerate and ATP, a reversible reaction primarily regulated by the ratio of ATP to ADP. In comparison, LDHA, a major subunit of LDH, converts pyruvate to lactate, which is the key characteristic of aerobic glycolysis. Furthermore, we observed that LDHA was overexpressed in pancreatic cancer cells and that inhibition of LDHA expression induced cancer cell oxidative stress and suppressed tumor growth (22, 23). In the present study, we focused on the effect of FOXM1 expression on glycolysis by regulation of LDHA expression. We then analyzed the association of FOXM1 expression with LDHA and LDHB expression, lactate production, and LDH activity in pancreatic cancer cell lines. We found that expression of FOXM1 was closely correlated with expression of LDHA, production of lactate, and activity of LDH but was not significantly correlated with expression of LDHB (Fig. 1C–E).
FOXM1 promotion of the Warburg effect in pancreatic cancer cells
Given that we observed that FOXM1 expression was associated with lactate production and LDH activity, to further explore the impact of FOXM1 on the Warburg effect in pancreatic cancer cells, we evaluated the correlation of altered FOXM1 expression with glucose utilization, lactate production, and LDH activity. We did so by knocking down FOXM1 expression in PANC-1 and FG cells, which had relatively high levels of FOXM1 and LDHA expression. We found significant decreases in glucose utilization, lactate production, and LDH activity (Fig. 2A; Supplementary Fig. S2). In comparison, FOXM1 overexpression markedly increased glucose utilization, lactate production, and LDH activity in BxPC-3 and AsPC-1 cells, which had relatively low levels of FOXM1 and LDHA expression (Fig. 2B; Supplementary Fig. S2). Furthermore, elevated glucose utilization, lactate production, and LDH activity induced by FOXM1 were dependent on LDHA, as specific inhibition of LDHA activity by oxamate sodium attenuated the increasing effect of FOXM1 on glucose utilization, lactate production, and LDH activity. These data indicated that FOXM1 may affect the utilization of glucose and production of lactate, which are the main features of the Warburg effect, via regulation of the activity of LDH.
Figure 2.
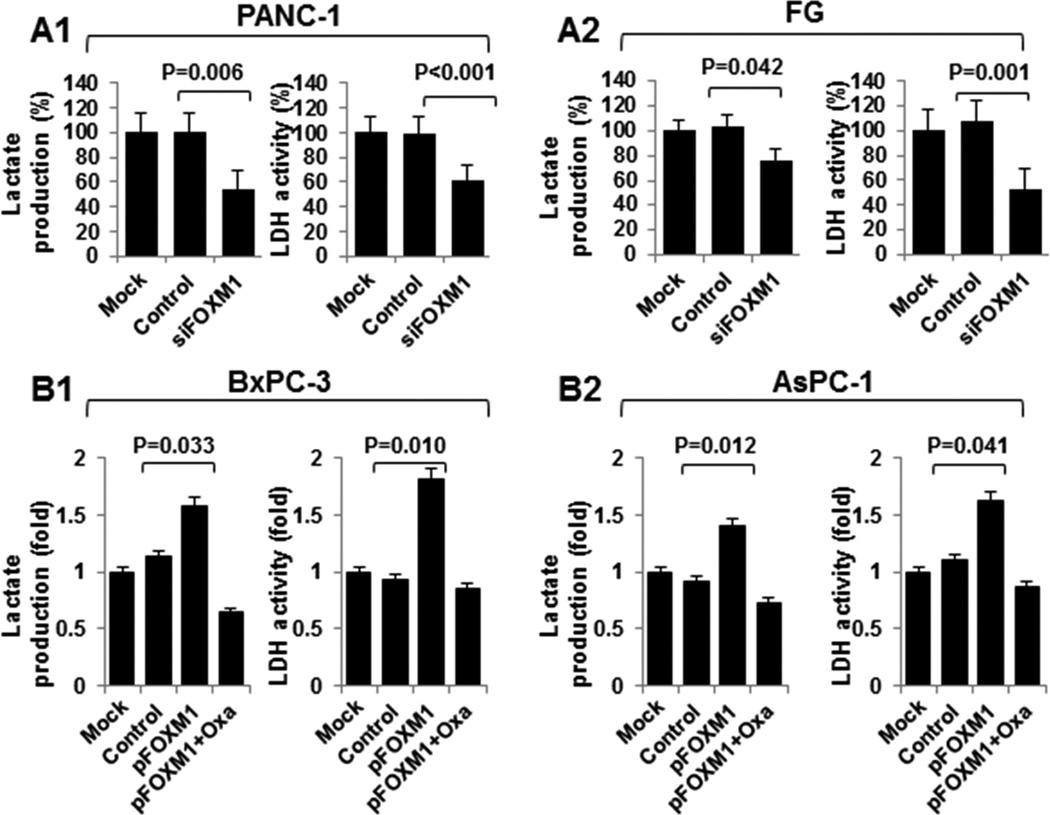
FOXM1 regulation of LDH activity and lactate production in pancreatic cancer cells. A and B, PANC-1 and FG cells were transfected with siFOXM1 or control siRNA (A), and BxPC-3 and AsPC-1 cells were transfected with pcDNA3.1, pcDNA3.1-FOXM1, or pcDNA3.1-FOXM1 and treated with 20 mmol/l oxamate sodium (B). The cultures were incubated for 48 hours. A Lactate Assay Kit was used to analyze the lactate production (left), and a Lactate Dehydrogenase Activity Assay Kit was used to analyze the LDH activity (right).
Transcriptional activation of LDHA expression by FOXM1 in pancreatic cancer cells
Our study demonstrated a direct association of FOXM1 expression with LDH activity and that FOXM1 knockdown can decrease LDHA mRNA expression. To further explore the mechanisms of FOXM1 regulation of LDH expression, we examined the effects of altered expression of FOXM1 on expression of LDHA and LDHB, the main subunits of LDH, at both the mRNA and protein level in the pancreatic cancer cell lines PANC-1, FG, BxPC-3, and AsPC-1. Silencing of FOXM1 expression significantly downregulated LDHA expression in PANC-1 and FG cells (Fig. 3A), whereas restored FOXM1 expression markedly upregulated LDHA expression in BxPC-3 and AsPC-1 cells (Fig. 3B). The expression of LDHB did not change with either overexpression or silencing of FOXM1. These data revealed that FOXM1 regulates LDH activity by regulating the expression of LDHA rather than LDHB.
Figure 3.
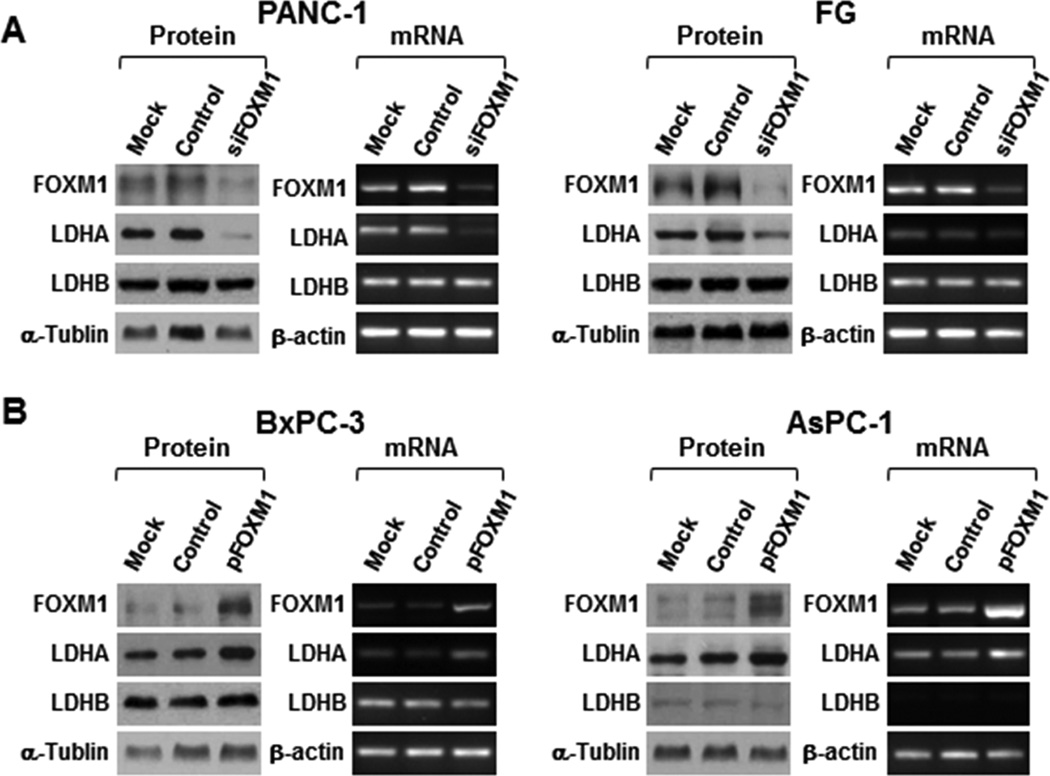
Regulation of LDHA expression by FOXM1. A and B, PANC-1 and FG cells were transfected with siFOXM1 or control siRNA (A), and BxPC-3 and AsPC-1 cells were transfected with pcDNA3.1-FOXM1 or pcDNA3.1 (B). The cultures were incubated for 48 hours. Total protein and RNA lysates were harvested to measure the FOXM1, LDHA, and LDHB expression using Western blot (left) and RT-PCR (right) analysis.
To identify the mechanism of FOXM1’s regulation of LDHA expression, we analyzed the sequence of the LDHA promoter for the potential FOXM1-binding elements 5'-A(C/T)AAA(C/T)AA-3', 5'-TAATCA-3', and 5'-AGATTGAGTA-3' (20, 24, 25). We identified four putative FOXM1-binding elements in the LDHA promoter region (Fig. 4A). We then generated two LDHA promoters—pLuc-LDHA-1480 and pLuc-LDHA-493—(Fig. 4A) and co-transfected the reporters with FOXM1 expression vectors or siFOXM1 into PANC-1, FG, BxPC-3, and AsPC-1 cells. As shown in Fig. 4E and F, overexpression of FOXM1 elevated the LDHA promoter activity, whereas knockdown of expression of FOXM1 decreased the LDHA promoter activity in the pLuc-LDA-1480 groups, and altered expression of FOXM1 did not change the promoter activity in the pLuc-LDHA-493 groups, which did not contain the potential FOXM1-binding site. To provide evidence that FOXM1 bound directly to the LDHA promoter, we conducted a ChIP assay using chromatins prepared from PANC-1 and FG cells. The results suggested that FOXM1 bound to all of the putative binding sites but mainly did so to sites #1 and #2 (Fig. 4B). To confirm these results, we knocked down FOXM1 expression in PANC-1 and FG cells, which led to decreased FOXM1 recruitment to the LDHA promoter (Fig. 4C), and induced FOXM1 overexpression in BxPC-3 and AsPC-1 cells, which led to increased FOXM1 recruitment to the LDHA promoter (Fig. 4D). These data strongly suggested that FOXM1 bound to the LDHA promoter and transcriptionally regulated LDHA expression.
Figure 4.
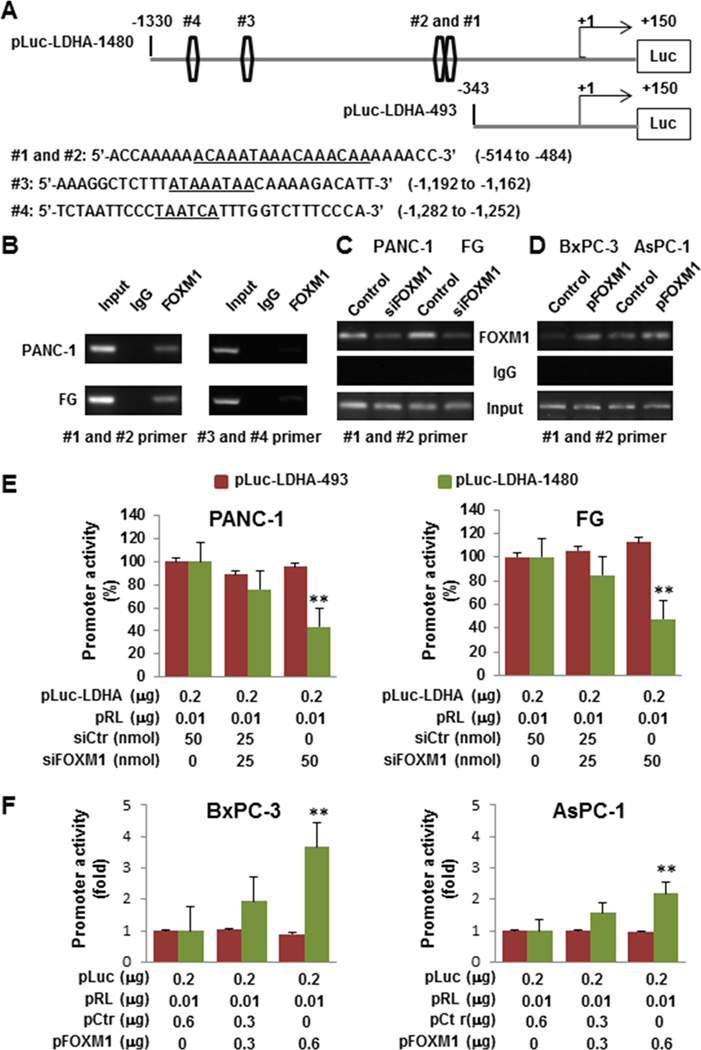
Direct binding of FOXM1 to the LDHA promoter. A, sequences and positions of putative FOXM1-binding elements on the LDHA promoter (#1, #2, #3, and #4). B, ChIP assay. Chromatins were isolated from PANC-1 and FG cells, and binding of FOXM1 to the LDHA promoter was examined using the ChIP assay. C, ChIP assay using chromatins isolated from PANC-1, PANC-1-siFOXM1, FG, and FG-siFOXM1 cells. D, ChIP assay using chromatins isolated from BxPC-3, BxPC-3-pFOXM1, AsPC-1, and AsPC-1-pFOXM1 cells. Normal IgG was used as a control, and 1% of the total cell lysates was subjected to PCR before immunoprecipitation (input control). E, Measurement of LDHA promoter activity. PANC-1 and FG cells were co-transfected with 0.2 µg of the LDHA promoter-luciferase construct pLuc-LDHA and 0, 25, and 50 nmol/l siFOXM1 or control siRNA. Promoter activity was examined using a dual luciferase assay kit. F, BxPC-3 and AsPC-1 cells were co-transfected with 0.2 µg of the LDHA promoter-luciferase construct pLuc-LDHA and 0, 0.3, and 0.6 µg of pcDNA3.1-FOXM1 or pcDNA3.1. Promoter activity was examined using a dual luciferase assay kit (**P < 0.01).
Direct impact of altered FOXM1-LDHA signaling on pancreatic tumor growth and metastasis in vivo
To determine the effect of altered FOXM1-LDHA signaling on pancreatic cancer biology, we induced overexpression of FOXM1 in AsPC-1 cells and knocked down expression of FOXM1 in FG cells and evaluated the ability of the cells with altered FOXM1 expression to grow and metastasize in animal models. Overexpression of FOXM1 significantly elevated the expression of LDHA in vivo (Fig. 5A), which was consistent with in vitro results, and promoted tumor growth and metastasis (Fig. 5B). Conversely, downregulation of expression of FOXM1 markedly inhibited the expression of LDHA (Fig. 5C) and tumor growth and metastasis (Fig. 5D). These data suggested that FOXM1-LDHA signaling regulated pancreatic cancer development and progression.
Figure 5.
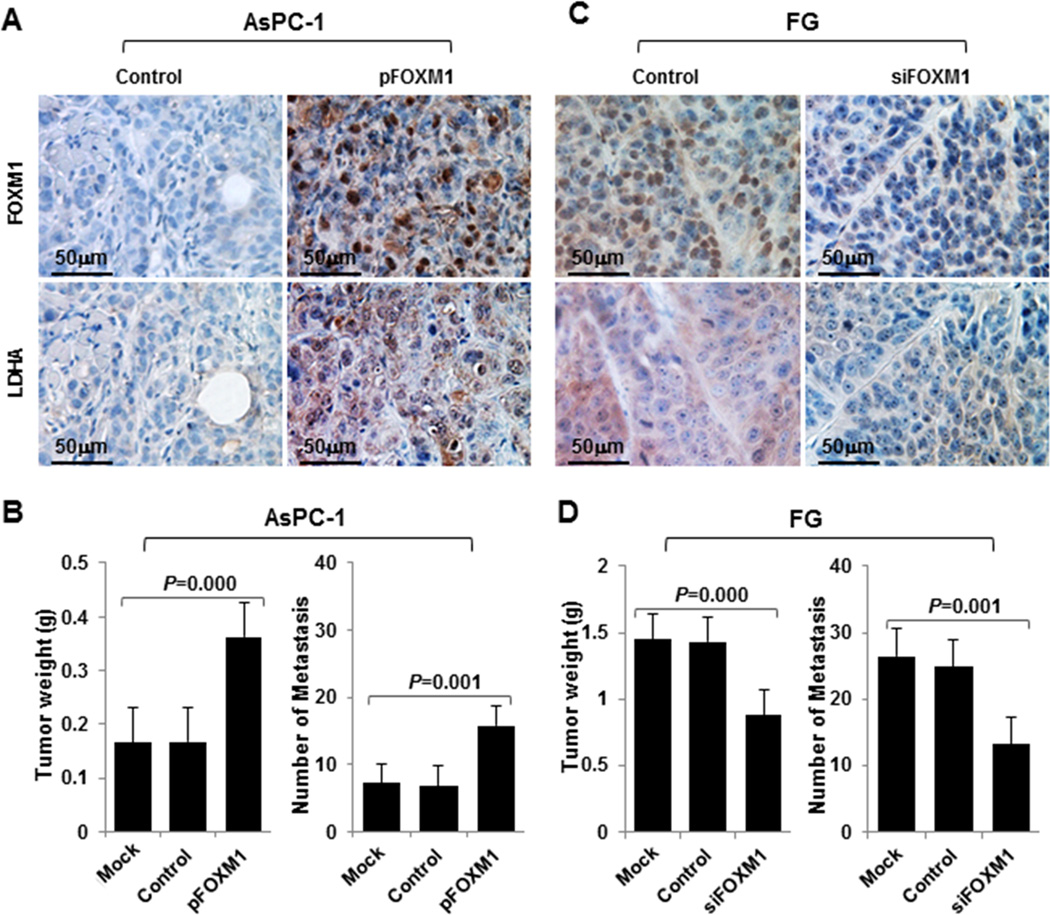
Influence of FOXM1-LDHA signaling on pancreatic cancer cell growth and metastasis in vivo. A–D, AsPC-1 cells with FOXM1 overexpression (A and B) or FG cells with knockdown of FOXM1 expression (C and D) were injected subcutaneously into the right scapular region in nude mice (1 × 106/mouse, n = 5) or intravenously into the ileocolic vein in nude mice (1 × 106/mouse, n = 5). The tumor-bearing mice were killed when they became moribund or on day 35 (subcutaneous) or day 21 (intravenous injection). Immunohistochemical staining of subcutaneous tumor specimens with specific anti-FOXM1 and anti-LDHA antibodies was performed. Shown are representative photos of the expression of FOXM1 and LDHA protein in AsPC-1 (A) and FG (C) cells, the weights of the tumors (B, left panel; D, left panel), and the numbers of liver surface metastases (B, right panel; D, right panel).
Direct correlation of concomitant expression of FOXM1 and LDH with pathologic features of pancreatic cancer
We provide evidence that FOXM1 transcriptionally regulated LDHA expression and glycolysis in pancreatic cancer cells. To further confirm our hypothesis, we investigated the expression of FOXM1 and LDHA in pancreatic tumor specimens using the TMA and TMA-P microarrays with different patient ethnicities. The clinicopathologic characteristics of the two arrays are described in Supplementary Table S2 and S3. We used the Mann-Whitney U test to analyze the matching of the clinicopathologic characteristics for the two arrays and found that most of the characteristics did not match (Supplementary Table S4). Thus, we analyzed TMA and TMA-P separately.
Using a specific anti-FOXM1 antibody, we observed FOXM1-positive staining in the nuclei and/or cytoplasm of pancreatic tumor cells and FOXM1-negative or weak FOXM1-positive staining in the cytoplasm of normal tumor-adjacent and normal pancreatic cells in TMA-P and TMA (Fig. 6A, upper panels; Fig. 6B, left panel; Supplementary Fig. S4A). We then analyzed the relationship between clinicopathologic characteristics and FOXM1 expression in pancreatic tumors. We observed that in TMA-P, FOXM1 expression was positively associated with pT classification (P < 0.001), regional lymph node metastasis (P < 0.001), TNM stage (P < 0.001) (Supplementary Fig. S3A1 and A2), and tumor differentiation (P = 0.005) (Supplementary Fig. S3B1 and B2; Supplementary Table S7). In TMA, FOXM1 expression was positively associated with regional lymph node metastasis (P = 0.035) and TNM stage (P = 0.015) but not other clinicopathologic characteristics (Supplementary Table S5), which may have resulted from the limited number of specimens. However, these data strongly suggested that elevated expression of FOXM1 played a critical role in pancreatic cancer development and progression.
Figure 6.
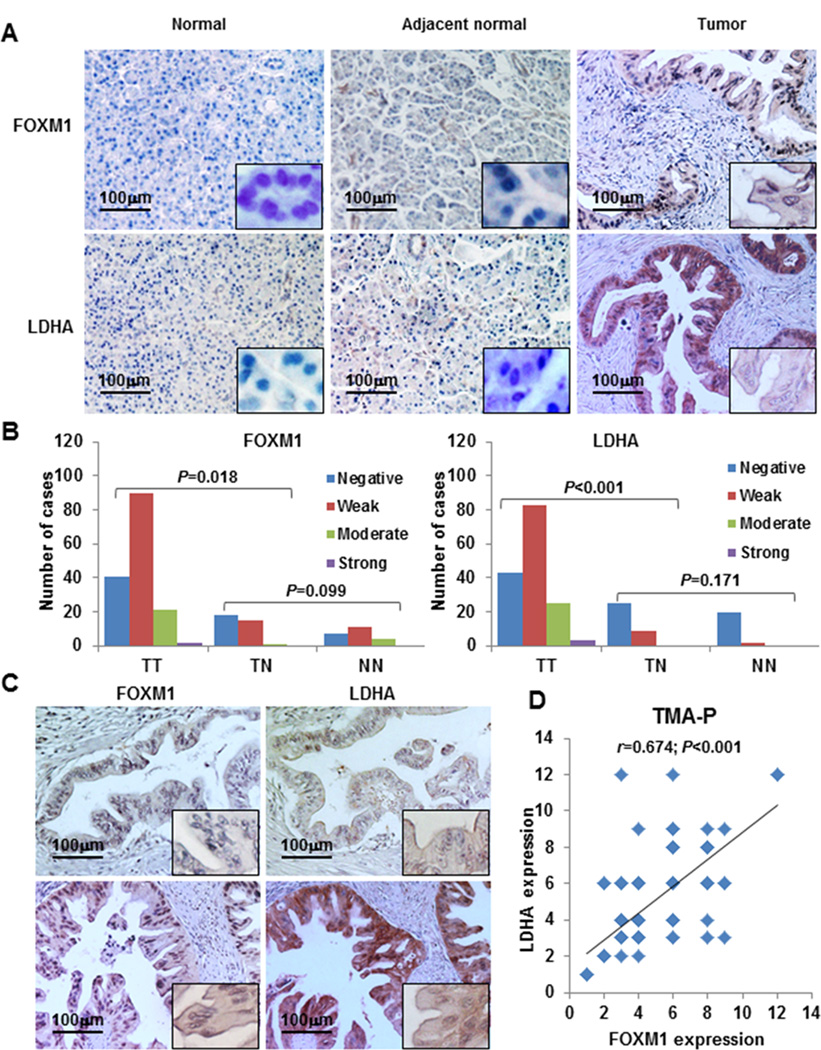
Concomitant expression of FOXM1 and LDHA in pancreatic tumors. Pancreatic tumor specimens in TMA and TMA-P were immunostained with specific anti-FOXM1 and anti-LDHA antibodies. A, representative images of FOXM1 and LDHA expression in normal pancreatic tissue, normal tumor-adjacent pancreatic tissue, and pancreatic tumor specimens (magnification, ×200). B, FOXM1 and LDHA expression in TMA-P. The expression levels were significantly higher in tumor (TT) than normal tumor-adjacent tissue (TN) and normal tissue (NN) specimens, whereas the expression of FOXM1 and LDHA did not differ between TN and NN. LDHA expression was associated with FOXM1 expression. C, representative photos of negative (upper two panels) and positive (lower two panels) FOXM1 and LDHA staining in pancreatic tumor specimens. D, direct correlation between FOXM1 expression and LDHA expression in TMA-P (n = 154; Pearson correlation coefficient, r = 0.674; P < 0.001). Some of the dots on the graph represent more than one specimen (overlapping scores).
Using the same cohort of TMA-P and TMA specimens, we analyzed the expression of LDHA and observed LDHA-positive staining in the cytoplasm of the cancer cells. We also saw increasingly LDHA-positive staining in normal pancreatic tissue, normal tumor-adjacent pancreatic tissue, and primary pancreatic tumor specimens (Fig. 6A, lower panels; Fig. 6B, right panel; Supplementary Fig. S4B). Expression of LDHA was also positively associated with pT classification (P = 0.001), TNM stage (P = 0.008) (Supplementary Fig. S3C1 and C2), and tumor differentiation (P = 0.018) (Supplementary Fig. S3D1 and D2; Supplementary Table S8) in TMA-P but positively associated only with TNM stage (P = 0.020) (Supplementary Table S6) in TMA.
Furthermore, we found direct correlations between FOXM1 and LDHA expression in both TMA-P and TMA, which were consistent with our findings for pancreatic cancer cell lines. In analyzing consecutive pancreatic tumor sections, we observed that pancreatic cancer cells expressed both FOXM1 and LDHA (Fig. 6C) and that FOXM1 expression was significantly correlated with LDHA expression in both TMA-P (r = 0.674, P < 0.001) (Fig. 6D) and TMA (r = 0.985, P < 0.001) (Supplementary Fig. S4C).
Discussion
In the present study, we determined the roles of FOXM1 in pancreatic cancer aerobic glycolysis and of FOXM1-LDHA signaling in pancreatic tumorigenesis. We provided four lines of evidence supporting a critical role for FOXM1 in regulation of aerobic glycolysis by the transcriptionally activated LDHA gene. First, we found that FOXM1 and LDHA were concomitantly overexpressed in pancreatic tumor specimens and cell lines. Also, the expression of FOXM1 and LDHA in pancreatic cancer cells was positively associated with lactate production and LDH activity. Second, overexpression of FOXM1 increased the expression of LDHA at both mRNA and protein level and increased glucose utilization, lactate production, and LDH activity, whereas reduced expression of FOXM1 did the opposite. These data indicated that FOXM1 regulated aerobic glycolysis by regulating the expression of LDHA. Third, FOXM1 bound directly to the LDHA promoter region and regulated expression of the LDHA gene at the transcriptional level. Fourth, increased FOXM1-LDHA signaling promoted pancreatic tumor cell growth and metastasis in vivo, whereas decreased FOXM1-LDHA signaling did the opposite. Thus, this novel FOXM1-LDHA signaling critically contributed to the Warburg effect in pancreatic cancer cells and to pancreatic cancer development and progression.
The oncogenic transcription factor FOXM1 is a key regulator of M-phase progression and G1/S- and G2/M-phase transition (26–29). Besides its essential roles in cell-cycle regulation, FOXM1 is involved in tumor angiogenesis, epithelial-mesenchymal transition, invasion, and metastasis (11). FOXM1 is overexpressed in many types of cancer cells, including liver cancer, prostate cancer, lung cancer, glioma, breast cancer, gastric cancer, and pancreatic cancer cells, and its overexpression is associated with tumor development and progression (18, 30–35). In the present study, we used two pancreatic cancer tissue microarrays with different patient ethnicities (TMA and TMA-P) and found that FOXM1 was overexpressed in pancreatic tumors and and that its overexpression was correlated with T stage, N stage, TNM stage, and tumor differentiation, which was consistent with results of our previous studies (12,13).
We have demonstrated the critical role of FOXM1 in cancer development and progression, but the literature contains little evidence revealing the function of FOXM1 in cancer metabolism. The Warburg effect, which researchers proposed more than 90 years ago, is considered a hallmark of cancer (36). Compared with normal cells, cancer cells take up and consume larger amounts of glucose and maintain much higher rates of glycolysis and lactate production even at normal oxygen concentrations (37). One of the explanations for the Warburg effect is overexpression of glucose transporters and the enzymes in the glycolytic pathway as a consequence of oncogene activation (9). In the present study, we found that knocking down expression of FOXM1 did not change the expression of most glycolytic enzymes except for PGK-1 and LDHA. Because LDHA mainly converts pyruvate to lactate, which is the key characteristic of aerobic glycolysis, we focused on the function of FOXM1 in glycolysis by regulating LDHA expression in this study.
LDHA is a major subunit of LDH, which is a tetrameric enzyme comprising the major subunits A and B, results in five isozymes: A4 (LDH5), A3B1 (LDH4), A2B2 (LDH3), A1B3 (LDH2), and B4 (LDH1). Recent studies revealed that LDHA favors catalysis of the conversion of pyruvate to lactate, whereas LDHB converts lactate to pyruvate; also, researchers linked an isozyme shift from LDH1 to LDH5 with cancer progression (38–40). Authors have reported that LDHA is overexpressed in many kinds of cancer cells, including breast, gynecologic, colorectal, and non-small cell lung cancer cells, and suggested that this overexpression promotes cancer development and progression (39, 41–43). In pancreatic cancer patients, the serum LDH level is elevated, and this elevation is an independent unfavorable prognostic factor (44). In the present study, we used the same cohorts of pancreatic tumor specimens used to analyze FOXM1 expression and found that LDHA expression was correlated with T stage, TNM stage, and tumor differentiation. These results suggested that LDHA expression was associated with pancreatic cancer development and progression. We then found that expression of LDHA was associated with that of FOXM1 in both TMA and TMA-P pancreatic tumor specimens regardless of patient ethnicity and type of cell lines, whereas we did not find a significant correlation between FOXM1 and LDHB expression. Furthermore, FOXM1 expression correlated with LDH activity and lactate production in pancreatic cancer cell lines. Next, we evaluated the effect of altered FOXM1 expression on LDHA and LDHB expression, glucose utilization, lactate production, and LDH activity. We found that FOXM1 overexpression led to a marked increase in LDHA expression, glucose utilization, lactate production, and LDH activity, whereas knockdown of FOXM1 expression did the opposite. Altered FOXM1 expression had no effect on LDHB expression. These date indicated that FOXM1 influenced the Warburg effect by regulating the expression of LDHA, which changed the activity of LDH.
Given the important roles of FOXM1 and LDHA expression in pancreatic cancer glycolysis, we sought to determine the underlying mechanisms responsible for co-expression of FOXM1 and LDHA in pancreatic cancer cells. Authors have reported that LDHA is the downstream target gene of hypoxia-inducible factor and c-Myc (45,46), and because FOXM1 is a transcription factor, we further studied FOXM1 to determine whether it regulates LDHA expression via transcription in the present study. We observed that FOXM1 bound directly to the LDHA promoter region and regulated the promoter activity of LDHA. Because LDHA is reported to promote the growth of pancreatic cancer cells (22), we further analyzed altered FOXM1-LDHA signaling to determine whether it regulates pancreatic tumor growth and metastasis in vivo. We found that elevated FOXM1-LDHA signaling increased both growth and metastasis, whereas decreased FOXM1-LDHA signaling did the opposite. Therefore, our results provide clinical and mechanistic evidence that FOXM1 regulates the Warburg effect and pancreatic cancer development and progression by transcriptionally regulating the expression of LDHA. Although our study demonstrated the role of FOXM1 in pancreatic cancer glycolysis and the mechanism of FOXM1 regulation of LDHA expression, further studies should be carried out to determine the role of FOXM1 in pancreatic cancer metabolism including the pentose phosphate pathway and glutamine, which also play key roles in cancer development and progression (47,48).
In summary, this study for the first time provided critical insight into the role of FOXM1 in pancreatic cancer glycolysis and identified a role for novel FOXM1-LDHA signaling in pancreatic cancer development and progression. We not only identified a novel molecular mechanism underlying mechanism of pancreatic cancer glycolysis and progression but also found that a new form of FOXM1-LDHA signaling is a promising molecular target for new therapeutic strategies for controlling pancreatic cancer.
Supplementary Material
Translational Relevance.
We used two pancreatic cancer tissue microarrays and molecular biologic and animal models to evaluate the activation and function of the FOXM1/lactate dehydrogenase A (LDHA) pathway in human pancreatic cancer cells. Our clinical and mechanistic findings indicated that LDHA is a direct transcriptional target of FOXM1 and that dysregulated FOXM1 expression, which occurs frequently in pancreatic cancer, leads to aberrant LDHA expression. Moreover, FOXM1 positively regulates pancreatic cancer cell aerobic glycolysis and growth, suggesting a novel molecular basis for the critical role of FOXM1 overactivation in pancreatic cancer metabolism and that dysregulated FOXM1-LDHA signaling is a promising new molecular target for novel preventive and therapeutic strategies for this malignancy. Therefore, our findings have a significant effect on clinical management of pancreatic cancer.
Acknowledgments
The authors thank Don Norwood for editorial comments.
Grant Support
This study was supported in part by grants R01-CA129956, R01-CA148954, R01CA152309, and R01CA172233 (to K.X.) from the National Institutes of Health; grants R01-CA116528 and R01-CA157933 (to S.H.) from the National Institutes of Health; and grants 81272917 and 81172022 (to Y.G.) from the National Natural Science Foundation of China.
Footnotes
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013;63:11–30. doi: 10.3322/caac.21166. [DOI] [PubMed] [Google Scholar]
- 3.Hidalgo M. Pancreatic cancer. N Engl J Med. 2010;362:1605–1617. doi: 10.1056/NEJMra0901557. [DOI] [PubMed] [Google Scholar]
- 4.Stathis A, Moore MJ. Advanced pancreatic carcinoma: current treatment and future challenges. Nat Rev Clin Oncol. 2010;7:163–172. doi: 10.1038/nrclinonc.2009.236. [DOI] [PubMed] [Google Scholar]
- 5.Warburg O. On the origin of cancer cells. Science. 1956;123:309–314. doi: 10.1126/science.123.3191.309. [DOI] [PubMed] [Google Scholar]
- 6.Garber K. Energy deregulation: licensing tumors to grow. Science. 2006;312:1158–1159. doi: 10.1126/science.312.5777.1158. [DOI] [PubMed] [Google Scholar]
- 7.Higashi T, Tamaki N, Torizuka T, Nakamoto Y, Sakahara H, Kimura T, et al. FDG uptake, GLUT-1 glucose transporter and cellularity in human pancreatic tumors. J Nucl Med. 1998;39:1727–1735. [PubMed] [Google Scholar]
- 8.Lee SM, Kim TS, Lee JW, Kim SK, Park SJ, Han SS. Improved prognostic value of standardized uptake value corrected for blood glucose level in pancreatic cancer using F-18 FDG PET. Clin Nucl Med. 2011;36:331–336. doi: 10.1097/RLU.0b013e31820a9eea. [DOI] [PubMed] [Google Scholar]
- 9.Chen JQ, Russo J. Dysregulation of glucose transport, glycolysis, TCA cycle and glutaminolysis by oncogenes and tumor suppressors in cancer cells. Biochim Biophys Acta. 2012;1826:370–384. doi: 10.1016/j.bbcan.2012.06.004. [DOI] [PubMed] [Google Scholar]
- 10.Ye H, Kelly TF, Samadani U, Lim L, Rubio S, Overdier DG, et al. Hepatocyte nuclear factor 3/fork head homolog 11 is expressed in proliferating epithelial and mesenchymal cells of embryonic and adult tissues. Mol Cell Biol. 1997;17:1626–1641. doi: 10.1128/mcb.17.3.1626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Koo CY, Muir KW, Lam EW. FOXM1: From cancer initiation to progression and treatment. Biochim Biophys Acta. 2012;1819:28–37. doi: 10.1016/j.bbagrm.2011.09.004. [DOI] [PubMed] [Google Scholar]
- 12.Huang C, Qiu Z, Wang L, Peng Z, Jia Z, Logsdon CD, et al. A novel FoxM1-caveolin signaling pathway promotes pancreatic cancer invasion and metastasis. Cancer Res. 2012;72:655–665. doi: 10.1158/0008-5472.CAN-11-3102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kong X, Li L, Li Z, Le X, Huang C, Jia Z, et al. Dysregulated expression of FOXM1 isoforms drive progression of pancreatic cancer. Cancer Res. 2013 doi: 10.1158/0008-5472.CAN-12-3859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Davis DB, Lavine JA, Suhonen JI, Krautkramer KA, Rabaglia ME, Sperger JM, et al. FoxM1 is up-regulated by obesity and stimulates beta-cell proliferation. Mol Endocrinol. 2010;24:1822–1834. doi: 10.1210/me.2010-0082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang L, Wei D, Huang S, Peng Z, Le X, Wu TT, et al. Transcription factor Sp1 expression is a significant predictor of survival in human gastric cancer. Clin Cancer Res. 2003;9:6371–6380. [PubMed] [Google Scholar]
- 16.Vezeridis MP, Tzanakakis GN, Meitner PA, Doremus CM, Tibbetts LM, Calabresi P. In vivo selection of a highly metastatic cell line from a human pancreatic carcinoma in the nude mouse. Cancer. 1992;69:2060–2063. doi: 10.1002/1097-0142(19920415)69:8<2060::aid-cncr2820690810>3.0.co;2-e. [DOI] [PubMed] [Google Scholar]
- 17.Zhai X, Yang Y, Wan J, Zhu R, Wu Y. Inhibition of LDH-A by oxamate induces G2/M arrest, apoptosis and increases radiosensitivity in nasopharyngeal carcinoma cells. Oncol Rep. 2013;30:2983–2991. doi: 10.3892/or.2013.2735. [DOI] [PubMed] [Google Scholar]
- 18.Liu M, Dai B, Kang SH, Ban K, Huang FJ, Lang FF, et al. FoxM1B is overexpressed in human glioblastomas and critically regulates the tumorigenicity of glioma cells. Cancer Res. 2006;66:3593–3602. doi: 10.1158/0008-5472.CAN-05-2912. [DOI] [PubMed] [Google Scholar]
- 19.Hatzivassiliou G, Zhao F, Bauer DE, Andreadis C, Shaw AN, Dhanak D, et al. ATP citrate lyase inhibition can suppress tumor cell growth. Cancer Cell. 2005;8:311–321. doi: 10.1016/j.ccr.2005.09.008. [DOI] [PubMed] [Google Scholar]
- 20.Zhang Y, Zhang N, Dai B, Liu M, Sawaya R, Xie K, et al. FoxM1B transcriptionally regulates vascular endothelial growth factor expression and promotes the angiogenesis and growth of glioma cells. Cancer Res. 2008;68:8733–8742. doi: 10.1158/0008-5472.CAN-08-1968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Marvin MR, Southall JC, Trokhan S, DeRosa C, Chabot J. Liver metastases are enhanced in homozygous deletionally mutant ICAM-1 or LFA-1 mice. J Surg Res. 1998;80:143–148. doi: 10.1006/jsre.1998.5322. [DOI] [PubMed] [Google Scholar]
- 22.Rong Y, Wu W, Ni X, Kuang T, Jin D, Wang D, et al. Lactate dehydrogenase A is overexpressed in pancreatic cancer and promotes the growth of pancreatic cancer cells. Tumour Biol. 2013;34:1523–1530. doi: 10.1007/s13277-013-0679-1. [DOI] [PubMed] [Google Scholar]
- 23.Le A, Cooper CR, Gouw AM, Dinavahi R, Maitra A, Deck LM, et al. Inhibition of lactate dehydrogenase A induces oxidative stress and inhibits tumor progression. Proc Natl Acad Sci U S A. 2010;107:2037–2042. doi: 10.1073/pnas.0914433107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wierstra I, Alves J. Despite its strong transactivation domain, transcription factor FOXM1c is kept almost inactive by two different inhibitory domains. Biol Chem. 2006;387:963–976. doi: 10.1515/BC.2006.120. [DOI] [PubMed] [Google Scholar]
- 25.Dai B, Kang SH, Gong W, Liu M, Aldape KD, Sawaya R, et al. Aberrant FoxM1B expression increases matrix metalloproteinase-2 transcription and enhances the invasion of glioma cells. Oncogene. 2007;26:6212–6219. doi: 10.1038/sj.onc.1210443. [DOI] [PubMed] [Google Scholar]
- 26.Wang X, Kiyokawa H, Dennewitz MB, Costa RH. The Forkhead Box m1b transcription factor is essential for hepatocyte DNA replication and mitosis during mouse liver regeneration. Proc Natl Acad Sci U S A. 2002;99:16881–16886. doi: 10.1073/pnas.252570299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Leung TW, Lin SS, Tsang AC, Tong CS, Ching JC, Leung WY, et al. Over-expression of FoxM1 stimulates cyclin B1 expression. FEBS Lett. 2001;507:59–66. doi: 10.1016/s0014-5793(01)02915-5. [DOI] [PubMed] [Google Scholar]
- 28.Fu Z, Malureanu L, Huang J, Wang W, Li H, van Deursen JM, et al. Plk1-dependent phosphorylation of FoxM1 regulates a transcriptional programme required for mitotic progression. Nat Cell Biol. 2008;10:1076–1082. doi: 10.1038/ncb1767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dai B, Gong A, Jing Z, Aldape KD, Kang SH, Sawaya R, et al. Forkhead box M1 is regulated by heat shock factor 1 and promotes glioma cells survival under heat shock stress. J Biol Chem. 2013;288:1634–1642. doi: 10.1074/jbc.M112.379362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kalinina OA, Kalinin SA, Polack EW, Mikaelian I, Panda S, Costa RH, et al. Sustained hepatic expression of FoxM1B in transgenic mice has minimal effects on hepatocellular carcinoma development but increases cell proliferation rates in preneoplastic and early neoplastic lesions. Oncogene. 2003;22:6266–6276. doi: 10.1038/sj.onc.1206640. [DOI] [PubMed] [Google Scholar]
- 31.Kalin TV, Wang IC, Ackerson TJ, Major ML, Detrisac CJ, Kalinichenko VV, et al. Increased levels of the FoxM1 transcription factor accelerate development and progression of prostate carcinomas in both TRAMP and LADY transgenic mice. Cancer Res. 2006;66:1712–1720. doi: 10.1158/0008-5472.CAN-05-3138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kim IM, Ackerson T, Ramakrishna S, Tretiakova M, Wang IC, Kalin TV, et al. The Forkhead Box m1 transcription factor stimulates the proliferation of tumor cells during development of lung cancer. Cancer Res. 2006;66:2153–2161. doi: 10.1158/0008-5472.CAN-05-3003. [DOI] [PubMed] [Google Scholar]
- 33.Madureira PA, Varshochi R, Constantinidou D, Francis RE, Coombes RC, Yao KM, et al. The Forkhead box M1 protein regulates the transcription of the estrogen receptor alpha in breast cancer cells. J Biol Chem. 2006;281:25167–25176. doi: 10.1074/jbc.M603906200. [DOI] [PubMed] [Google Scholar]
- 34.Li Q, Zhang N, Jia Z, Le X, Dai B, Wei D, et al. Critical role and regulation of transcription factor FoxM1 in human gastric cancer angiogenesis and progression. Cancer Res. 2009;69:3501–3509. doi: 10.1158/0008-5472.CAN-08-3045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Xia JT, Wang H, Liang LJ, Peng BG, Wu ZF, Chen LZ, et al. Overexpression of FOXM1 is associated with poor prognosis and clinicopathologic stage of pancreatic ductal adenocarcinoma. Pancreas. 2012;41:629–635. doi: 10.1097/MPA.0b013e31823bcef2. [DOI] [PubMed] [Google Scholar]
- 36.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 37.Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis? Nat Rev Cancer. 2004;4:891–899. doi: 10.1038/nrc1478. [DOI] [PubMed] [Google Scholar]
- 38.Koukourakis MI, Giatromanolaki A, Simopoulos C, Polychronidis A, Sivridis E. Lactate dehydrogenase 5 (LDH5) relates to up-regulated hypoxia inducible factor pathway and metastasis in colorectal cancer. Clin Exp Metastasis. 2005;22:25–30. doi: 10.1007/s10585-005-2343-7. [DOI] [PubMed] [Google Scholar]
- 39.Koukourakis MI, Giatromanolaki A, Sivridis E, Bougioukas G, Didilis V, Gatter KC, et al. Lactate dehydrogenase-5 (LDH-5) overexpression in non-small-cell lung cancer tissues is linked to tumour hypoxia, angiogenic factor production and poor prognosis. Br J Cancer. 2003;89:877–885. doi: 10.1038/sj.bjc.6601205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kim JH, Kim EL, Lee YK, Park CB, Kim BW, Wang HJ, et al. Decreased lactate dehydrogenase B expression enhances claudin 1-mediated hepatoma cell invasiveness via mitochondrial defects. Exp Cell Res. 2011;317:1108–1118. doi: 10.1016/j.yexcr.2011.02.011. [DOI] [PubMed] [Google Scholar]
- 41.Goldman RD, Kaplan NO, Hall TC. Lactic dehydrogenase in human neoplastic tissues. Cancer Res. 1964;24:389–399. [PubMed] [Google Scholar]
- 42.Koukourakis MI, Kontomanolis E, Giatromanolaki A, Sivridis E, Liberis V. Serum and tissue LDH levels in patients with breast/gynaecological cancer and benign diseases. Gynecol Obstet Invest. 2009;67:162–168. doi: 10.1159/000183250. [DOI] [PubMed] [Google Scholar]
- 43.Koukourakis MI, Giatromanolaki A, Sivridis E, Gatter KC, Harris AL. Lactate dehydrogenase 5 expression in operable colorectal cancer: strong association with survival and activated vascular endothelial growth factor pathway--a report of the Tumour Angiogenesis Research Group. J Clin Oncol. 2006;24:4301–4308. doi: 10.1200/JCO.2006.05.9501. [DOI] [PubMed] [Google Scholar]
- 44.Tas F, Aykan F, Alici S, Kaytan E, Aydiner A, Topuz E. Prognostic factors in pancreatic carcinoma: serum LDH levels predict survival in metastatic disease. Am J Clin Oncol. 2001;24:547–550. doi: 10.1097/00000421-200112000-00003. [DOI] [PubMed] [Google Scholar]
- 45.Hu CJ, Wang LY, Chodosh LA, Keith B, Simon MC. Differential roles of hypoxia-inducible factor 1alpha (HIF-1alpha) and HIF-2alpha in hypoxic gene regulation. Mol Cell Biol. 2003;23:9361–9374. doi: 10.1128/MCB.23.24.9361-9374.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Shim H, Dolde C, Lewis BC, Wu CS, Dang G, Jungmann RA, et al. c-Myc transactivation of LDH-A: Implications for tumor metabolism and growth. Proc Natl Acad Sci U S A. 1997;94:6658–6663. doi: 10.1073/pnas.94.13.6658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Riganti C, Gazzano E, Polimeni M, Aldieri E, Ghigo D. The pentose phosphate pathway: an antioxidant defense and a crossroad in tumor cell fate. Free Radic Biol Med. 2012;53:421–436. doi: 10.1016/j.freeradbiomed.2012.05.006. [DOI] [PubMed] [Google Scholar]
- 48.Shanware NP, Mullen AR, DeBerardinis RJ, Abraham RT. Glutamine: pleiotropic roles in tumor growth and stress resistance. J Mol Med (Berl) 2011;89:229–236. doi: 10.1007/s00109-011-0731-9. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.