

Abstract
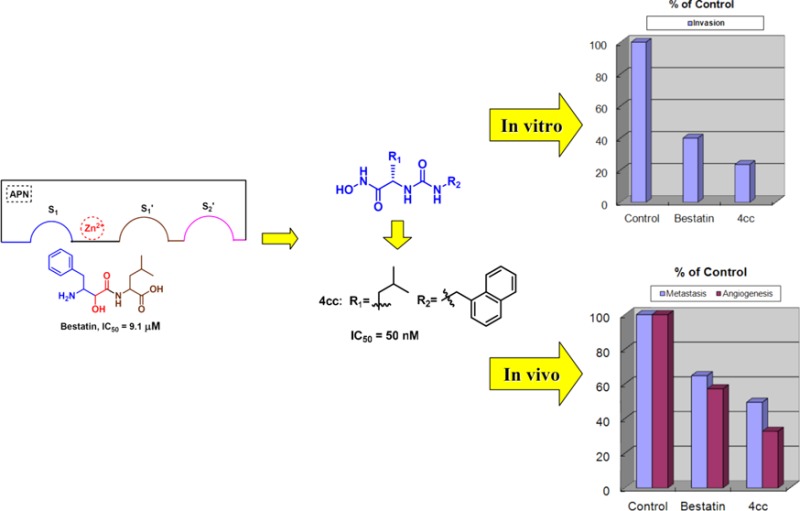
Cancer metastasis is a major barrier to its treatment and an important cause of patient death. Antimetastatic agents hold promise for patients with advanced metastatic tumors. Aminopeptidase N/CD13 (APN) is being pursued by many as an important target against cancer metastasis and angiogenesis, but there are few reports on the in vivo evaluation of synthetic APN inhibitors. Herein, a series of compounds targeting APN were synthesized and evaluated for their antimetastasis and antiangiogenesis potency both in vitro and in vivo. Excitingly, compounds 4m, 4t, and 4cc, with the most potent APN inhibitory activities, displayed significant antimetastasis and antiangiogenesis effects in vitro and in vivo, suggesting that those synthetic APN inhibitors have the potential to overcome cancer metastasis and angiogenesis.
Keywords: APN/CD13, inhibitors, anticancer, metastasis, angiogenesis
An enduring problem in cancer treatment is the high failure rates in patients with advanced metastatic cancers.1 Cancer metastasis is considered as a key cause of operation failure, postoperative relapse, and ultimately death.2 Metastasis is a complex, multistep process, in which cancer cells spread from primary sites to new places.3 Cancer cell invasion is the key stage, and angiogenesis is the prerequisite.4 The extracellular matrix (ECM) is the main barrier of malignant cell dissemination. The ECM is a substrate of aminopeptidase N (APN/CD13, EC 3.4.11.2) and matrix metalloproteinases (MMPs). Targeting MMPs in the treatment of cancer metastasis has yielded unsatisfactory results.5 Consequently, in recent years APN has become one of the most studied cancer therapeutic targets.6
APN is a zinc-dependent integral peptidase,7 belonging to the M1 family of the MA clan of peptidases.8 As an exopeptidase, APN preferentially cleaves neutral amino acids from the N-terminal of various oligopeptides, including enkephalins, neurokinins,9 angiotensins, and some cytokines.10 Human APN exists in many tissues, organs, and cell types, including kidney proximal tubule cells, stem cells, epithelial cells, endothelial cells, fibroblast cells, leukemia cells, and immune cells, etc.7,11 APN degrades ECM to promote malignant cell invasion into the bloodstream.11 In the angiogenesis process, neo-endothelial cells invading through the ECM also need APN.12 Therefore, APN would be a key target in cancer metastasis and angiogenesis.13 Meanwhile, the dysregulated expression and high-level exopeptidase activity of APN are detected in various mammalian cancer cell lines14 such as melanoma, prostate, ovarian, colon, renal and pancreas carcinomas, etc., as well as neo-endothelial cells. The statistical survival rates for patients with high APN expression solid tumors were reported to be significantly lower than those of patients with APN-negative solid tumors.15 So far, reported APN inhibitors can be divided into natural products and synthetic compounds. Among them, bestatin (Ubenimex16) may be the most studied one. It is isolated from a culture filtrate of Streptomyces olivoreticuli and is a cancer chemotherapeutic drug in some countries (Figure 1A).17 Bestatin is a potent APN inhibitor with a Ki value of 3.03 μM.18 Saiki et al. found that bestatin significantly inhibited cancer cell invasion in a dose-dependent manner.19 However, research on synthetic compounds only aimed at evaluating their APN and cell proliferation inhibition. There are few reports on their in vivo antimetastatic and antiangiogenic effects. Therefore, in this paper, potent synthetic small-molecule APN inhibitors were designed and synthesized, and their antimetastasis and antiangiogenesis effects were evaluated in vitro and in vivo.
Figure 1.

(A) Chemical structure of bestatin; (B) chemical structure of D24; (C) target compounds.
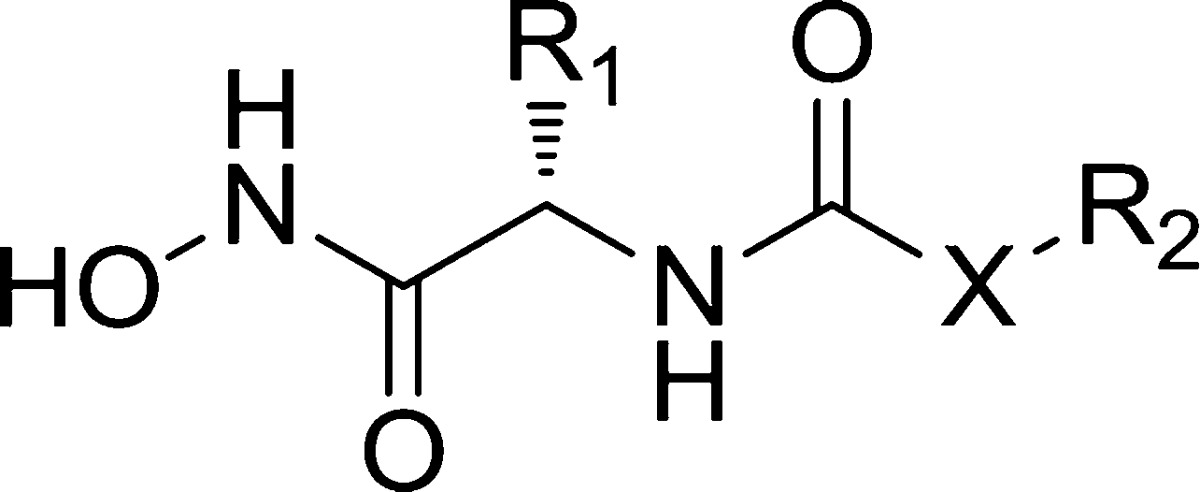
In 2006, Yoshimoto et al. reported the crystal structure of APN from Escherichia coli with the active site occupied by bestatin.20 Recently, Gu et al. engineered Thermoplasma acidophilum factor F3 (a homologous protein of human APN) with two-point directed mutation, which obtained the protein with an identical active site to human APN. Structural biology studies were then performed on the cocrystal structure of this protein and one active APN inhibitor (D24, Figure 1B) developed by our group. The result suggested that engineered factor F3 mutant has a larger binding pocket than E. coli APN.21 According to the chemical structures of bestatin and D24, both of them have a zinc-binding group (ZBG) and a neutral amino acid residue. In our ongoing studies, hydroxamic acid as strong ZBG in D24 was kept and five neutral amino acids (l-leucine, l-phenylglycine, l-isoleucine, l-phenylalanine, and l-methione) were introduced to the new target structures. The nonpeptide structure in D24 was replaced by various aromatic rings or an aromatic fragment with an alkyl linker so as to increase interaction with the larger pocket of human APN (Figure 1C).
The target compounds, 4a-4cc, 7, and 9 were first examined for their inhibitory activities toward APN from porcine kidney. Most compounds exhibited more potent inhibitory activities than bestatin, some of which had 10-fold or more improvement (Table 1). The substituents greatly influenced the compounds potency. Replacement of the ureido linker provides a serious potency decrease, such as for 7 and 9. With the same R2 substituent, l-leucine and l-phenylglycine residues contributed more to the potency than the other residues. As for R2 substituents, a long side chain between the aromatic ring and the ureido group decreased the potency, such as for 4i, 4j, 4d, and 4h. Their potencies increased along with the chain length decline, while not so regularly in 4k, 4a, and 4e. Most of the l-leucine based compounds with substituted phenyl or benzyl on the R2 position were much more potent. 4zb with phenyl was less potent than 4i with benzyl, which could also be seen in 4b versus 4za with fluorine, and in 4p versus 4x with an l-leucine residue. But the opposite phenomenon was seen in 4q versus 4w and in 4o versus 4v, with methyl or methoxy. As heterocycle R2 substituents, thiophene (4r) contributed more to the potency than furan (4l). For the l-leucine-based compounds with substituted benzyl at R2, electron donating groups seemed better than electron withdrawing groups. For instance, 4t was more potent than 4u, as well as 4w versus 4x, 4o and 4q versus 4p, also seen in 4y and 4z versus 4b, except 4v. This might also demonstrate that methyl or methoxy fit into the pocket better than fluorine. The substituted position on benzyl also influenced the potency. With methoxy substituted benzyl, 4t (meta-) seemed better than 4s (ortho-) and 4v (para-), while this is not so obvious in the fluorine substituted ones 4u and 4x. The most potent compounds were 4m (IC50 = 99 nM) with 1-naphthyl and 4cc (IC50 = 50 nM) with 1-naphthylmethyl, suggesting that a larger group was better here, though 4n with a biphenyl group was much less potent.
Table 1. Structures and IC50 Values of the Target Compoundsa.
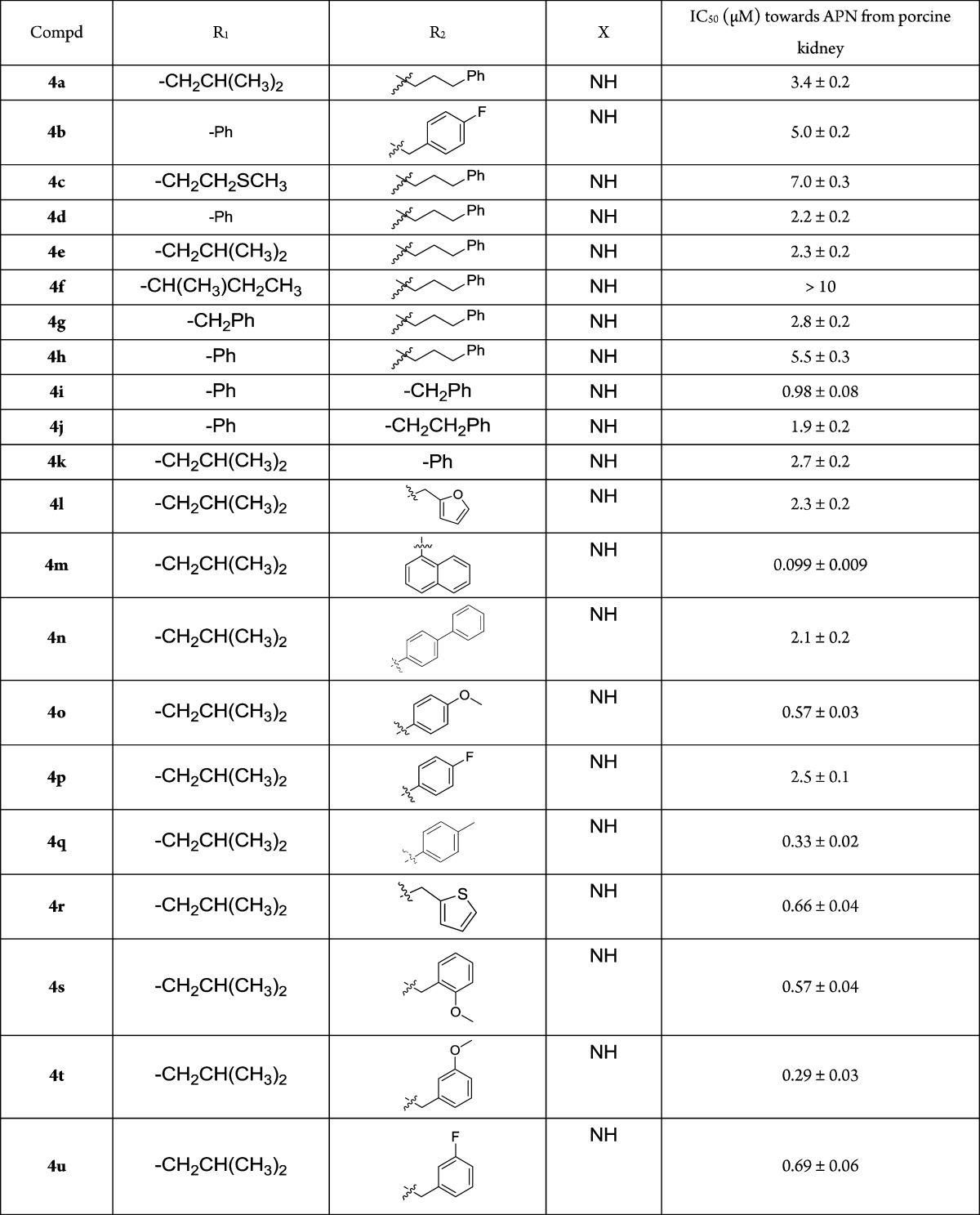
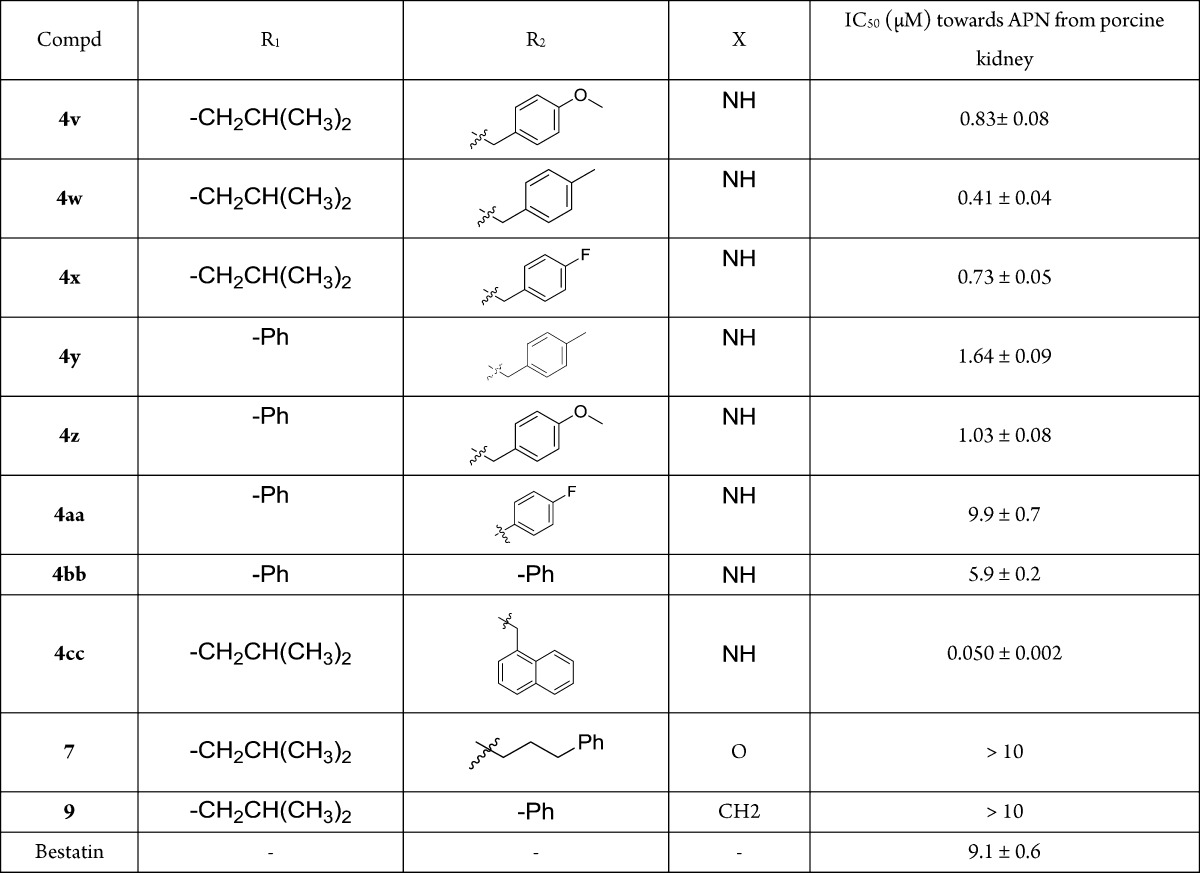
All compounds were assayed three times, and the results are expressed with standard deviations.
ES-2 cells with high APN expression were used as human APN, which makes more sense to the following in vitro and in vivo results. We only assessed the ability of the nanomolar range compounds to inhibit human APN on cultured ES-2 human ovarian clear cell carcinoma cells (Table 2). Those compounds were still better than bestatin, some even with more than 10-fold improvement. The effects of the more potent ones (4m, 4q, 4t, 4x, and 4cc) on ES-2 cell survival were evaluated, and Table 2 showed that at low concentration all compounds had almost no influence on ES-2 cell survival. At high concentration, there was only a slight antiproliferative effect for most of the compounds, except 4x. Matrix metalloproteinase-2 (MMP-2) is also a zinc-dependent metalloproteinase responsible for cancer invasion, so MMP-2 inhibitory activity was also tested. 4m, 4q, 4t, and 4cc exhibited very high selectivity for APN over MMP-2 (Table 2).
Table 2. IC50 Values and Inhibition Rates of the Compoundsa.
| growth inhibition rates toward ES-2 cells |
|||||
|---|---|---|---|---|---|
| compd | IC50 (μM) toward APN on the surface of ES-2 cells | 5 μg/mL | 50 μg/mL | IC50 (μM) toward MMP-2 | IC50 (MMP-2)/IC50 (APN from procine kidney) |
| 4m | 0.48 ± 0.05 | <1.0% | 3.63% | >1000 | >106 |
| 4o | 6.4 ± 0.6 | <1.0% | 6.85% | >1000 | >106 |
| 4q | 0.94 ± 0.08 | <1.0% | 1.78% | 573 ± 35 | 1740 |
| 4s | 1.1 ± 0.1 | <1.0% | 3.29% | >1000 | >106 |
| 4t | 0.79 ± 0.07 | <1.0% | <1.0% | >1000 | >106 |
| 4u | 3.0 ± 0.3 | <1.0% | 2.81% | >1000 | >106 |
| 4v | 6.9 ± 0.6 | <1.0% | <1.0% | >1000 | >106 |
| 4w | 1.5 ± 0.1 | <1.0% | 1.83% | >1000 | >106 |
| 4x | 0.91 ± 0.09 | <1.0% | 11.9% | >1000 | >106 |
| 4cc | 0.42 ± 0.03 | <1.0% | 6.67% | 545 ± 30 | 10900 |
| bestatin | 16.0 ± 1.6 | <1.0% | 2.89% | 276 ± 19 | 30.2 |
The compounds were assayed three times, and the results are expressed with standard deviations.
Cell migration is an important step during the invasion process. Agents blocking cell motility would also exhibit anti-invasion effects. A cell migration assay was performed on transwell chambers without Matrigel coating. Figure 2A showed that all the tested compounds (150 μM) had only a slight inhibitory tendency to ES-2 cell migration, without statistical significance. An anti-invasion assay was performed on transwell chambers coated with Matrigel. According to the result, ES-2 cells could freely invade and pass through Matrigel, but this was significantly blocked by bestatin, 4m, 4q, 4t, and 4cc. In 1989 Azuma et al. reported the tumor cell invasion block effects of bestatin in various conditions to mouse B16BL6 and 3LL cells. The result indicated that bestatin had approached 50–80% inhibitory rates at the 100 mg/mL level, with a dose-dependent tendency.22 According to our results on human ES-2 cell invasion inhibition, 4m and 4cc were more potent than bestatin at the concentrations of 15 μM and 150 μM (Figure 2B(g)). Figure 2 shows that 4m, 4q, 4t, and 4cc were able to significantly inhibit ES-2 cell invasion in a dose-dependent tendency without obvious influence on ES-2 cell survival and migration, which indicated their potential therapeutic application in overcoming cancer metastasis. The ECM gel (Matrigel)-induced human umbilical vascular endothelial cell (HUVEC) capillary tubular structure formation assay was used as an in vitro measurement of angiogenesis.23 Bestatin, 4m, 4t, and 4cc decreased the number of branch points formed by HUVECs (data not shown), which demonstrated the potential antiangiogenesis effects of the APN inhibitors.
Figure 2.
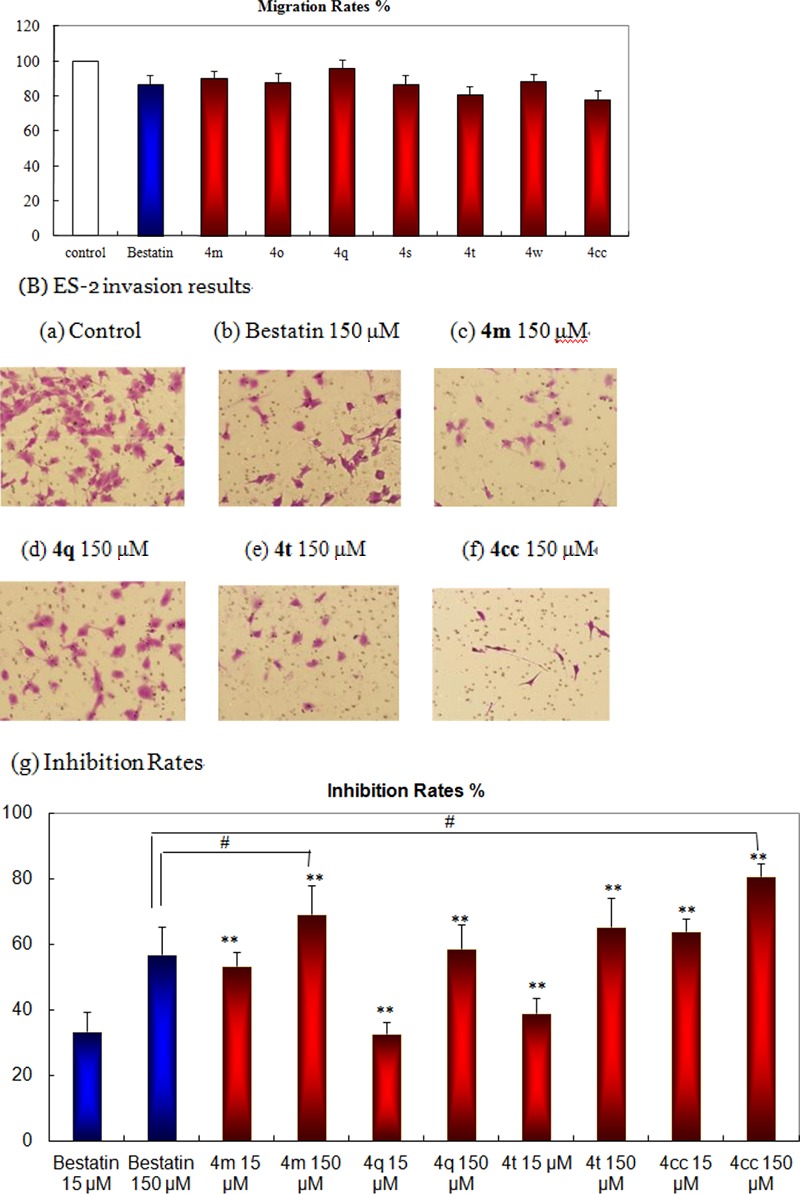
(A) ES-2 cell migration. (B) ES-2 cell invasion inhibition: (a) control; (b) bestatin 150 μM; (c) 4m, 150 μM; (d) 4q, 150 μM; (e) 4t, 150 μM; (f) 4cc, 150 μM; (g) statistical chart. Each column represents the mean values with SD for three independent experiments.***, P < 0.001; **, P < 0.01; *, P < 0.05, versus the control. ###, P < 0.001; ##, P < 0.01; #, P < 0.05, versus Bestatin treated groups.
B16BL6 cells, which represent an advanced malignant murine tumor more metastatic than B16,24 were selected to examine 4m, 4t, and 4cc on tumor-induced angiogenesis and experimental metastasis in vivo. 4m, 4t, and 4cc effectively inhibited murine APN activity and had quite low antiproliferative effects on B16BL6 (Figure 3A). In 2004, Saiki et al. demonstrated a significant antiangiogenesis effect of bestatin (100 or 200 mg/kg, p.o.) on the C57BL6 mouse dorsal air sac model with the B16BL6 cell line.25 In our assay, angiogenesis was induced by the implantation of B16BL6 cells into the intradermal site of C57BL/6 mouse backs,26 and it quantified by counting the number of vessels oriented toward the tumor mass27 under a dissecting microscope. Figure 3B showed that the intraperitoneal administration of 4m, 4t, and 4cc at the 100 mg/kg level led to significant antiangiogenesis effects, which were significantly better than those for bestatin (P < 0.01). In 2006, our group reported a murine H22 liver cancer experimental metastasis on Kunming mice, and bestatin showed potency (60–70% inhibitory rate) at 50 mg/kg orally.28 At this time, further study on APN inhibitor in vivo antimetastasis effects was performed through choosing a much higher advanced metastatic cancer cell line, B16BL6. The result showed that intravenous inoculation of B16BL6 resulted in the establishment of lung metastatic sites in C57BL/6 mice (as high as 170 nodes in the control group), much more serious than H22 (only 33 nodes). A significant decrease of the B16BL6 pulmonary colonies was detected in 4m, 4t, and 4cc treated groups (Figure 3C).
Figure 3.
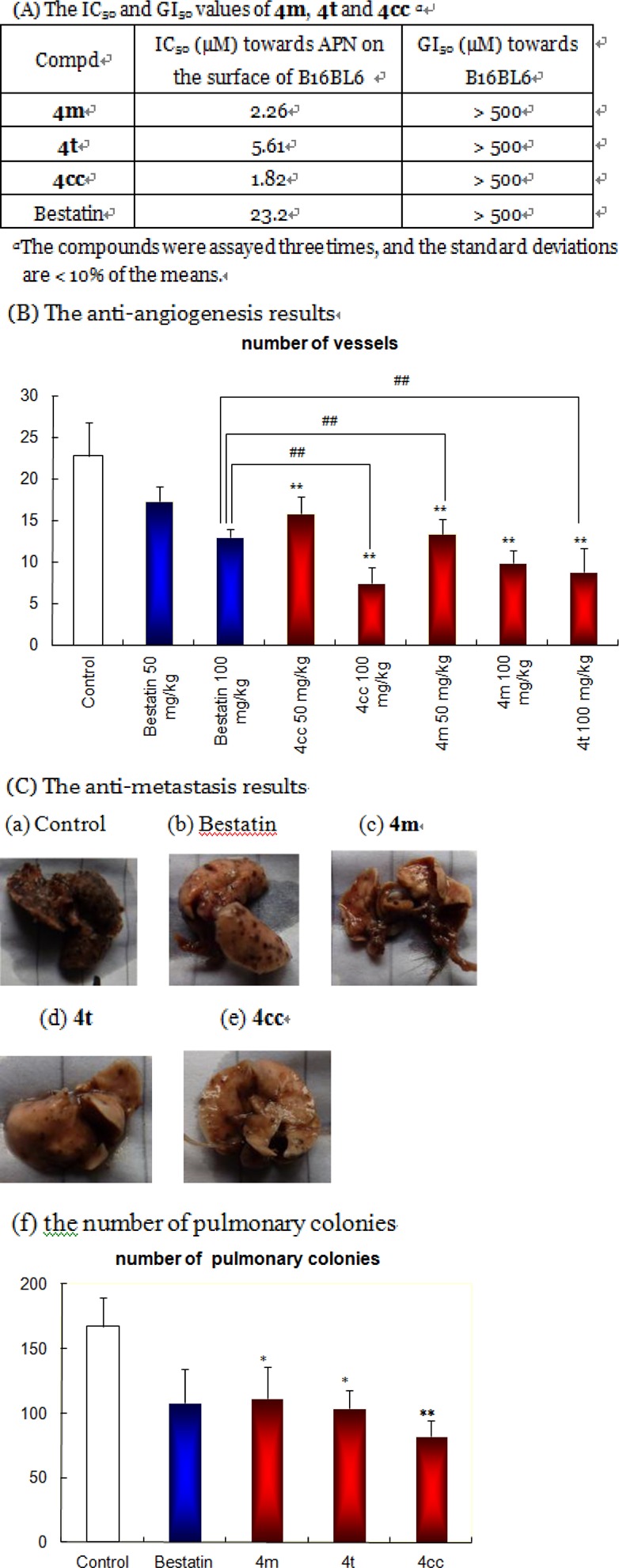
(A) The IC50 and GI50 values of 4m, 4t, and 4cc. (B) 4m, 4t, and 4cc inhibit B16BL6 induced angiogenesis in vivo. **, P < 0.01, versus the control. ##, P < 0.01, versus bestatin treated groups. (C) B16BL6 pulmonary colonies. (a) Control, (b) bestatin, (c) 4m, (d) 4t, (e) 4cc, (f) statistical chart. Each column represents the mean values with SD *, P < 0.05, versus the control, **, P < 0.01, versus the control.
The synthetic methods for compounds 4a–4cc are shown in Scheme 1A. The amines listed were transformed into isocyanates through triphosgene. The ureido linker was obtained from the isocyanates coupled with the amino acid methyl esters. Without further purification, they were directly transformed into hydroxamic acids as the target compounds. 7 was synthesized according to Scheme 1B. l-Leucine methyl ester was converted into isocyanate29 and coupled with 1-phenyl-1-propanol to obtain carbamate linker, which was finally transformed into hydroxamic acid. And the synthetic method of 9 was shown in Scheme 1C. The amide bond was formed through the coupling between l-leucine methyl ester and phenacetyl chloride, and the hydroxamic acid was obtained by the same method as above.
Scheme 1.

We have designed and synthesized a series of small-molecule synthetic APN inhibitors. The APN inhibitory activity demonstrated that most compounds were more potent than bestatin. The three most potent candidates, 4m, 4t, and 4cc, with a nanomolar range IC50, exhibited significant anti-invasion effects to ES-2 cells in vitro without obvious cytotoxicity, which is better than bestatin. In the in vivo evaluation including murine B16BL6 induced angiogenesis and experimental lung metastasis assays, 4m, 4t, and 4cc exhibited significant antiangiogenesis and antimetastasis effects without obvious cytotoxicity toward B16BL6 cells. Such results suggest that those three synthetic APN inhibitors are potential candidates which can be developed to overcome cancer metastasis and angiogenesis.
Acknowledgments
We thank Dr. Jiong Jia for 1H NMR and 13C NMR of the target compounds.
Supporting Information Available
Synthesis of compounds, experimental procedures, characterization of new compounds, 1H NMR, 13C NMR, and biological experiments. This material is available free of charge via the Internet at http://pubs.acs.org.
This work was supported by a National Natural Science Foundation Research Grant (Grant No. 21172134), the Ph.D. Programs Foundation of the Ministry of Education of China (Grant No. 20110131110037), Major Programs of the Chinese Academy of Sciences during the 12th Five-Year Plan Period (Grant No. 2011ZX09401-015), the Shandong Province Natural Science Foundation (Grant No. ZR2010HM028), the Natural Science Foundation for Young Scholars of Shandong Province (2006BS03021), and the Independent Innovation Foundation of Shandong University, IIFSDU (2012JC003).
The authors declare no competing financial interest.
Supplementary Material
References
- Francia G.; Cruz-Munoz W.; Man S.; Xu P.; Kerbel R. S. Mouse models of advanced spontaneous metastasis for experimental therapeutics. Nat. Rev. Cancer 2011, 11, 135–141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein C. A. The Metastasis Cascade. Science 2008, 321, 1785–1787. [DOI] [PubMed] [Google Scholar]
- Wirtz D.; Konstantopoulos K.; Searson P. C. The physics of cancer: the role of physical interactions and mechanical forces in metastasis. Nat. Rev. Cancer 2011, 11, 512–522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaffer C. L.; Weinberg R. A. A Perspective on Cancer Cell Metastasis. Science 2011, 331, 1559–1564. [DOI] [PubMed] [Google Scholar]
- Sang Q.; Jin Y.; Newcomer R.; Monroe S.; Fang X.; Hurst D.; Lee S.; Cao Q.; Schwartz M. Matrix metalloproteinase inhibitors as prospective agents for the prevention and treatment of cardiovascular and neoplastic diseases. Curr. Top. Med. Chem. 2006, 6, 289–316. [DOI] [PubMed] [Google Scholar]
- Wickstrom M.; Larsson R.; Nygren P.; Gullbo J.; Aminopeptidase N. (CD13) as a target for cancer chemotherapy. Cancer Sci. 2011, 102, 501–508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antczak C.; Meester I. D.; Bauvois B. Ectopeptidases in pathophysiology. Bioessays 2001, 23, 251–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlings N.; Barrett A. MEROPS. The peptidase database 1999, 271325–331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furuhashi M.; Mizutani S.; Kurauchi O.; Kasugai M.; Narita O.; Tomoda Y. In vitro degradation of opioid peptides by human placental aminopeptidase M. Exp. Clin. Endocrinol. 1988, 92, 235–237. [DOI] [PubMed] [Google Scholar]
- Hoffmann T.; Faust J.; Neubert K.; Ansorge S. Dipeptidyl peptidase IV (CD26) and Aminopeptidase N (CD13) catalyzed hydrolysis of cytokines and peptides with N-terminal cytokine sequences. FEBS Lett. 1993, 336, 61–64. [DOI] [PubMed] [Google Scholar]
- Antczak C.; De Meester I.; Bauvois B. Transmembrane proteases as disease markers and targets for therapy. J. Biol. Regul. Homeostasis Agents 2001, 15, 130–139. [PubMed] [Google Scholar]
- Carmelit P.; Jain R. Angiogenesis in cancer and other diseases. Circ. Res. 2000, 87, 176–178.10926865 [Google Scholar]
- Bauvois B.; Dauzonne D. Aminopeptidase-N/CD13 (EC 3.4.11.2) inhibitors: chemistry, biological evaluations, and therapeutic prospects. Med. Res. Rev. 2006, 26, 88–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoscato A.; Stamkoski R.; Babcock G.; Alexande J. Neutral surface aminopeptidasc activity of human tumor cell lines. Biochem. Biophys. Acta 1990, 1041, 317–319. [DOI] [PubMed] [Google Scholar]
- Kawamura J.; Shimada Y.; Kitaichi H.; Komoto I.; Hashimoto Y.; Kaganoi J.; Miyake M.; Yamasaki S.; Kondo K.; Imamura M. Clinicopathological significance of aminopeptidase N/CD13 expression in human gastric carcinoma. Hepatogastroenterology 2007, 54, 36–40. [PubMed] [Google Scholar]
- Rich D.; Moon B. J.; Harbeson S. Inhibition of aminopeptidases by amastatin and bestatin derivatives. Effect of inhibitor structure on slow-binding processes. J. Med. Chem. 1984, 27, 417–422. [DOI] [PubMed] [Google Scholar]
- Scornik O.; Botbol V. Bestatin as an experimental tool in mammals. Curr. Drug Metab. 2001, 2, 67–85. [DOI] [PubMed] [Google Scholar]
- Leyhausen G.; Schuster D.; Vaith P.; Zahn R.; Umezawa H.; Falke D.; Muller W. Identification and properties of the cell membrane bound leucine aminopeptidase interacting with the potential immuno-stimulant and chemotherapeutic agent bestatin. Biochem. Pharmacol. 1983, 32, 1051–1057. [DOI] [PubMed] [Google Scholar]
- Saiki I.; Yoneda J.; Azuma I.; Fujii H.; ; Abe F.; Nakajima M.; Tsuruo T. Role of aminopeptidase N (CD13) in tumor-cell invasion and extracellular matrix degradation. Int. J. Cancer 1993, 54, 137–143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito K.; Nakajima Y.; Onohara Y.; Takeo M.; Nakashima K.; Matsubara F.; Ito T.; Yoshimoto T. Aminopeptidase n (Proteobacteria alanyl aminopeptidase) from escherichia coli: crystal structure and conformational change of the methionine 260 residue involved in substrate recognition. J. Biol. Chem. 2006, 281, 33664–33676. [DOI] [PubMed] [Google Scholar]
- Su J.; Wang Q.; Feng J.; Zhang C.; Zhu D.; Wei T.; Xu W.; Gu L. Engineered Thermoplasma acidophilum factor F3 mimics human aminopeptidase N (APN) as a target for anticancer drug development. Bioorg. Med. Chem. 2011, 19, 2991–2996. [DOI] [PubMed] [Google Scholar]
- Saiki I.; Murata J.; Watanabe K.; Fujii H.; Abe F.; Azuma I. Inhibition of tumor cell invasion by Ubenimix (bestatin) in vitro. Jpn. J. Cancer Res. 1989, 80, 873–878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao H.; Liu H.; Chen Y.; Xin X.; Li J.; Hou Y.; Zhang Z.; Zhang X.; Xie C.; Geng M.; Ding J. Oligomannurarate sulfate, a novel heparanase inhibitor simultaneously targeting basic fibroblast growth factor, combats tumor angiogenesis and metastasis. Cancer Res. 2006, 66, 8779–8787. [DOI] [PubMed] [Google Scholar]
- Talmadge J.; Zbar B. Clonality of pulmonary metastases from the bladder 6 subline of the B16 melanoma studied by southern hybridization. J. Natl. Cancer Inst. 1987, 78, 315–320. [PubMed] [Google Scholar]
- Aozuka Y.; Koizumi K.; Saitoh Y.; Ueda Y.; Sakurai H.; Saiki I. Anti-tumor angiogenesis effect of aminopeptidase inhibitor bestatin against B16-BL6 melanoma cells orthotopically implanted into syngeneic mice. Cancer Lett. 2004, 216, 35–42. [DOI] [PubMed] [Google Scholar]
- Gude R. P.; Binda M. M.; Boquete A. L.; Bonfil R. D. Inhibition of endothelial cell proliferation and tumor-induced angiogenesis by pentoxifylline. J. Cancer Res. Clin. Oncol. 2001, 127, 625–630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidky Y. A.; Auerbach R. Lymphocyte-induced angiogenesis: a quantitative and sensitive assay of the graft-vs-host reaction. J. Exp. Med. 1975, 141, 1084–1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li Q.; Fang H.; Wang X.; Hu L.; Xu W. Novel cyclic-imide peptidomimetics as aminopeptidase N inhibitors. Design, chemistry and activity evaluation. Part I. Eur. J. Med. Chem. 2009, 44, 4819–4825. [DOI] [PubMed] [Google Scholar]
- Tsai J.; Takaoka L.; Powell N.; Nowick J. Synthesis of amino acid ester isocyanates. Org. Synth. 2002, 78, 220–224. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.