

Abstract
This study investigated the potential antibacterial activity of three series of compounds synthesized from 12 linear and branched polyamines with 2–8 amino groups, which were substituted to produce the corresponding guanides, biguanides, or phenylguanides, against Acinetobacter baumannii, Enterococcus faecalis, Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus. Antibacterial activity was measured for each compound by determining the minimum inhibitory concentration against the bacteria, and the toxicity towards mammalian cells was determined. The most effective compound, THAM trisphenylguanide, was studied in time-to-kill and cytoplasmic leakage assays against methicillin-resistant Staphylococcus aureus (MRSA, USA300) in comparison to chlorhexidine. Preliminary toxicity and MRSA challenge studies in mice were also conducted on this compound. THAM trisphenylguanide showed significant antibacterial activity (MIC ∼1 mg/L) and selectivity against MRSA relative to all the other bacteria examined. In time-to-kill assays it showed increased antimicrobial activity against MRSA versus chlorhexidine. It induced leakage of cytoplasmic content at concentrations that did not reduce cell viability, suggesting the mechanism of action may involve membrane disruption. Using an intraperitoneal mouse model of invasive MRSA disease, THAM trisphenylguanide reduced bacterial burden locally and in deeper tissues. This study has identified a novel guanide compound with selective microbicidal activity against Staphylococcus aureus, including a methicillin-resistant (MRSA) strain.
Introduction
Ever since antibiotics were introduced into clinical practice, bacterial pathogens have been developing resistance which reduces or eliminates their effectiveness. In addition, opportunistic pathogens with innate resistance to antibiotics have become emerging problems, particularly in hospital settings. In developed countries, such as the United States, bacterial species that have acquired multiple drug resistance include the ESKAPE pathogens, aptly named for their ability to escape the effects of our current antimicrobial drugs. These include E nterococcus faecium, S taphylococcus aureus, K lebsiella pneumoniae, A cinetobacter baumannii, P seudomonas aeruginosa and E nterobacter species [1]. As an example, methicillin-resistant Staphylococcus aureus (MRSA) was once thought to be problematic only in healthcare settings, but now community-associated MRSA infections are becoming more common. MRSA have become resistant to the multiple classes of antibiotics including beta lactams, macrolides, and quinolones [2], as well as the glycopeptide vancomycin, the common drug of last resort [3].
We have synthesized three series of compounds, derived from linear and branched polyamines with two to eight amino groups, which were substituted to produce the corresponding guanides, biguanides, or phenylguanides [4]. Some of these compounds were previously found to be effective as antagonists of CXCR4 and inhibited HIV infection by X4 strains [4], [5] and the formation of metastatic tumors [6]. The biguanide series of compounds are chemically related to some commercially important biocidal agents found in common household items such as disinfectants, mouthwashes, contact lens solutions, and cosmetics. Examples include polyhexamethylene biguanide (PHMB), a heterogeneous polymer containing multiple positively charged biguanides, and the small-molecule bis-biguanides chlorhexidine and alexidine. Our new compounds are also polycationic, and preliminary experiments had shown low cytotoxicity in vitro of many of the new compounds [4]; we hypothesized that our compounds might have more selective antibacterial activity than these other biguanides. In the current study we investigate the antimicrobial potential of these compounds and demonstrate the efficacy of THAM trisphenylguanide against MRSA both in vitro and in vivo.
Materials and Methods
Guanide, Biguanide and Phenylguanide Compounds
The compounds were synthesized and purified as described previously [4], with the following additions: Trisoctylaminomelamine (TOAM) was synthesized using 1,8-diaminooctane as previously described for trishexylaminomelamine (THAM) except that an equal volume of acetonitrile was added to the reaction after 2 h in refluxing water to help with solubility of the more non-polar compound and the reaction was allowed to reflux for an additional hour in 1∶1 acetonitrile/water. The TOAM guanide derivatives were synthesized as described previously for the corresponding THAM compounds [4].
Bacterial Strains
Bacterial strains used in these experiments were Escherichia coli (strain K91), Pseudomonas aeruginosa (strain PA01), Acinetobacter baumannii (strains ATCC 19606 and ATCC BAA1605), Enterococcus faecalis (strains MSU#10 and ATCC V583 [7]), and Staphylococcus aureus (strains RN4220 [8] and community-associated MRSA strain LAC, pulsed-field gel-electrophoresis type USA300 [9]). Three of the strains display known drug-resistant phenotypes. Specifically, A. baumannii BAA1605 is a multi-drug resistant isolate [10], [11], with resistance to ceftazidime, gentamicin, ticarcillin, piperacillin, aztreonam, cefepime, ciprofloxacin, imipenem, and meropenem. E. faecalis V583 is vancomycin and gentamicin resistant [7] and USA300 (strain LAC used in this study) is resistant to penicillin, oxacillin, erythromycin, and shows intermediate resistance to ciprofloxacin [9], [12], [13].
Minimum Inhibitory Concentration and Minimum Bactericidal Concentration Assays
Minimum inhibitory concentration (MIC) values were determined for those compounds showing the most antibacterial activity in the bacterial viability assays (see Supplemental Materials) against each of the strains of bacteria using a microdilution method in 96 well plates with LB media according to guidelines published by the British Society for Antimicrobial Chemotherapy [14]. Minimum bactericidal concentrations (MBC) were determined by plating 10 µL and 100 µL from each well showing no signs of growth after overnight incubation of the MIC plates and determining the minimum concentration showing a 3-log reduction of viable bacteria. MIC and MBC were also determined in cation-adjusted Mueller-Hinton broth (Hardy Diagnostics, Santa Maria, CA) for some compounds and strains.
Kill Curve Assay
Bacteria were cultured to mid-log phase (OD600 ∼0.4) in LB media with shaking and then diluted to approximately 104 to 105 CFU/mL. Bacteria were then treated with compound at the MIC, 2×MIC, and 4×MIC (as determined by the MIC assays). Over the course of 24 hours, bacteria were incubated at 37°C and aliquots removed at various time intervals, diluted, and plated on LB agar. Plated bacteria were incubated overnight at 37°C and CFU/mL were calculated for each time interval.
Cytoplasmic Leakage & Bacteriolysis Assay
Bacteria were cultured to mid-log phase (OD600 ∼0.4) in LB media with shaking. Bacteria were then harvested and resuspended in PBS to a concentration of ∼108 CFU/mL. Cultures were treated with compound at the MIC, 2×MIC, and 4×MIC and incubated at 37°C with shaking. Bacteria were monitored over time for cytoplasmic leakage via A260 measurements and for cell viability. For A260 readings, sample aliquots were centrifuged for 5 min at 10,000×g at room temperature to remove cellular debris. Viability was monitored by determining the CFU/mL during the first 3 hours post treatment as described above for the kill curve assays.
Cell Toxicity Assays
Compound cytotoxicity was evaluated using the CellTiter 96® Aqueous Non-Radioactive Cell Proliferation Assay (MTS) following the manufacturer’s instructions (Promega, Madison, WI) using both the human breast cancer cell line MDA-MB-231 (ATCC HTB-26) and HaCaT human keratinocytes [15]. The HaCaT cell line was grown in Epilife medium (MEPI500Ca) with human keratinocyte growth supplement (Invitrogen, San Diego, CA). Both types of cells were harvested using Tryspin/EDTA and diluted to 1×105 cells per well in a 96-well plate. Ten concentrations covering five orders of magnitude were tested with six replicates per concentration. Cells were incubated for 48–72 hours at 37°C with 5% CO2. MTS/PMS solution (Promega, Madison, WI) was added to the wells and absorbance was measured at 490 nm two to four hours later. Data was analyzed using GraphPad Prism version 5.0 (GraphPad Software, La Jolla, CA) using a variable slope inhibition dose response curve.
In vivo MRSA Challenge Experiments
Three groups of BALB/c mice (8 mice per group) were infected by intraperitoneal (i.p.) injection with 5×107 MRSA in 100 µl sterile PBS [16], [17]. The first treatment group was injected i.p. with 100 µl of 100 µM THAM trisphenylguanide in PBS immediately after infection. The second group was injected i.p. with 100 µl of 100 µM THAM trisphenylguanide one hour post-infection. The control group was injected with 100 µl PBS. At eight hours post-infection, all mice were euthanized. An intraperitoneal lavage using 10 ml PBS was performed to recover bacteria within the peritoneal cavity and plated at multiple dilutions on TSA plates. The heart and kidney were removed for evaluation of systemic infection. Briefly, tissues were weighed, homogenized, and plated at multiple dilutions on TSA plates. TSA plates were incubated overnight at 37°C at 5% CO2 and CFU enumerated the next day.
Ethical Approval
All studies conformed to National Institutes of Health guidelines and were approved by the Animal Care and Use Committee at Montana State University (MSU)-Bozeman, MT (Protocol #2011-60).
Statistics
Statistical analysis was performed using GraphPad Prism version 6.0 (GraphPad Software, La Jolla, CA). Data for each analysis represents a minimum of 2 repetitions. Data sets were analyzed using one-way analysis of variance with Bonferroni post-test or using Student’s t-test. Error bars represent the standard error of the mean.
Results
Antibacterial Activity In vitro
We initially screened all the guanide, biguanide, and phenylguanide compounds we had synthesized, and their parent amines, in a bacterial viability assay against three Gram- negative (E. coli, P. aeruginosa, and A. baumannii) as well as two Gram-positive species (S. aureus and E. faecalis). The IC50 values (Table S1) were used to estimate the relative activity level of the various compounds and determine which would be investigated further. Chlorhexidine was used as a positive control with known bactericidal activity [18].
The most active compounds were then examined in an in vitro assay to determine their minimum inhibitory concentrations (MIC) against these pathogens (Table 1). Trishexylaminomelamine trisphenylguanide (THAM-3ΦG, Figure 1), which had shown antibacterial activity against almost all the strains in the viability assay, showed notable activity only against E. coli and S. aureus in the MIC assay, with some selectivity against S. aureus (2 mg/L) compared to E. coli (8 mg/L). The anti-bacterial activity against MRSA shown by THAM-3ΦG is comparable to that shown by chlorhexidine. The slightly larger TOAM-3ΦG also displayed MIC values comparable to chlorhexidine against E. coli, E. faecalis, and S. aureus. The derivatives of the DNT2300 and DNT2200 dendrimers showed almost no activity against any of the bacteria in the MIC assay.
Table 1. Minimum Inhibitory Concentrations of the most active guanide (G) and phenylguanide (ΦG) compounds.
| MIC (mg/L) | ||||||||
| Bacteria/Strain | E. coli | P. aeruginosa | A. baumannii | A. baumannii | E. faecalis | E. faecalis | S. aureus | MRSA |
| Compound | K91 | PA01 | ATCC 19606 | ATCC BAA1605 | MSU#10 | V583 | RN4220 | USA300 |
| Chlorhexidine | 1 | 8 | 16 | 16 | 4 | 6 | 1 | 1 |
| THAM-3ΦG | 8 | 48 | >256 | >256 | 32 | 256 | 2 | 2 |
| DNT2300-6BG | >256 | >256 | ND | ND | >256 | ND | >256 | ND |
| DNT2300-6ΦG | 256 | >256 | >256 | ND | >256 | ND | 256 | 256 |
| DNT2200-8ΦG | >256 | >256 | ND | ND | ND | ND | >256 | ND |
| TOAM | >64 | 64 | >64 | ND | >64 | >64 | >64 | 48 |
| TOAM-3G | 8 | 16 | >64 | ND | 16 | >64 | 2 | 1 |
| TOAM-3ΦG | 2 | >64 | >64 | ND | 2 | 4 | 2 | 2 |
Values are the median of at least 3 determinations. >256 indicates no inhibition at the highest concentration tested. TOAM was insoluble at >64 mg/L.
Figure 1. Structures of chlorhexidine (top) and THAM-3ΦG.

The MIC assays were repeated using Mueller-Hinton broth (MHB) for THAM-3ΦG and chlorhexidine against E. coli and S. aureus (Table 2); the results were comparable to those obtained in LB (Table 1). The minimum bactericidal concentrations (MBC) of THAM-3ΦG, and chlorhexidine were also determined in MHB for S. aureus and E. coli (Table 2). For S. aureus, the MBCs were similar to the MICs in each case; for E. coli the MBC was significantly higher than the MIC for THAM-3ΦG (Table 2).
Table 2. Median MICs and MBCs for chlorhexidine and THAM-3ΦG in MHB.
| Bacteria/Strain | E. coli | S. aureus | MRSA | |||
| K91 | RN4220 | USA300 | ||||
| Compound | MIC | MBC | MIC | MBC | MIC | MBC |
| Chlorhexidine | 0.44 | 0.44 | 0.89 | 0.89 | 0.67 | 0.89 |
| THAM-3ΦG | 8.0 | 16 | 1.0 | 2.0 | 1.0 | 1.0 |
There were no significant differences between MIC and MBC for each S. aureus strain and no significant differences between the two S. aureus strains using 2-tailed unpaired T-tests (p>0.05, n≥7).
Cytotoxicity In vitro
The toxicity of the compounds to human cells was evaluated using an MTS dye reduction assay in both an epithelial (MDA-MB-231) and a keratinocyte (HaCat) derived cell line (Table 3). Many of the compounds were not toxic to the human cells at the highest concentrations tested, and almost all of them were less toxic than chlorhexidine. The increased cytotoxicity (Table 3) of the TOAM compounds compared to the THAM compounds makes them less attractive as antibacterials, although the TOAM-3ΦG toxicity appeared to be no greater than that of chlorhexidine.
Table 3. Cytotoxicities of the guanide, biguanide, phenylguanide derivatives, and the parent amines, against a human breast cancer cell line (MDA-231) and a human keratinocyte cell line (HaCat).
| Compound | CC50 a (mg/L) | Relative to chlorhexidineb | ||
| MDA-231 | HaCat | MDA-231 | HaCat | |
| Chlorhexidine | 2.6 | 1.9 | - | - |
| Spermidine | 22 | >150 | 8.5 | >79 |
| Spermidine-3ΦG | 430 | 400 | 170 | 210 |
| Spermine | 10 | >200 | 3.7 | >110 |
| Spermine-4ΦG | >110 | 110 | >43 | 56 |
| THAM | >770 | 200 | >290 | 100 |
| THAM-3G | 74 | 63 | 29 | 33 |
| THAM-BG | >1000 | 72 | >380 | 37 |
| THAM-3ΦG | 7.6 | 14 | 3.0 | 7.1 |
| DNT2300 | 29 | 13 | 11 | 6.9 |
| DNT2300-6G | >77 | 98 | >30 | 51 |
| DNT2300-6BG | 60 | 11 | 24 | 5.7 |
| DNT2300-6ΦG | 280 | 400 | 110 | 210 |
| DNT2200 | 69 | 16 | 27 | 8.3 |
| DNT2200-8G | >100 | 710 | >38 | 370 |
| DNT2200-8BG | 920 | 870 | 360 | 450 |
| DNT2200-8ΦG | >260 | 210 | >100 | 110 |
| TOAM | ND | 4.1 | ND | 2.1 |
| TOAM-3G | ND | 3.6 | ND | 1.9 |
| TOAM-2ΦG | ND | 3.1 | ND | 1.6 |
| TOAM-3ΦG | ND | 2.4 | ND | 1.3 |
CC50 is the concentration of compound which killed 50% of the cells in the MTS assay.
Factor by which cytotoxicity was reduced relative to chlorhexidine.
Time-to-Kill Assays
THAM-3ФG was comparable in effectiveness at killing MRSA to chlorhexidine at their MIC, and slightly more effective at 4×MIC (Figure 2). A 3-log reduction in CFU/mL was seen within 4 hours post treatment with THAM-3ФG at 4×MIC and no viable cells remained after 24 hours at the MIC. Chlorhexidine showed no significant dose-dependence of the reduction in bacterial viability over the range of 1× to 4×MIC, although bactericidal activity was seen by 24 hours. Chlorhexidine killed E. coli more rapidly and at lower concentrations (Figure 2a and b). At 2 mg/L it caused a >3 log reduction in CFU/mL within 4 hours of treatment, and no viable cells remained after 6 hours. At 4×MIC (32 mg/L), THAM-3ФG showed a >3 log reduction only after 4 hours of treatment, and it took 24 hours to kill all the cells at this concentration.
Figure 2. Time-to-kill assays.

MRSA (a–b) and E. coli (c–d) treated with chlorhexidine (a, c) and THAM-3ФG (b, d). Significant differences between THAM-3ФG and chlorhexidine treatments were only seen at 4×MIC at 4 and 6 h with MRSA (p-value<0.01) and at 1 and 2 h with E. coli (p-value<0.05). P-values were calculated for each time point and between compounds by one-way ANOVA.
Cytoplasmic Leakage Assay
The potential for THAM-3ФG to interact with the cellular membrane of MRSA in a way that would cause leakage of cellular contents was investigated and compared to that of chlorhexidine. The cellular contents leaking out of the cell, specifically amino acids and nucleic acids, were detected by an increase in the absorbance of the cell supernatant at 260 nm over time compared to untreated controls. As shown in Figure 3, treatment with both THAM-3ФG and chlorhexidine at 4×MIC caused a significant increase in A260 within the first 30 minutes of treatment, as compared to untreated controls. To verify that this leakage was not secondary to cellular death resulting from other mechanisms, the number of viable cells was also determined over the 3 hour period. There was no significant reduction in CFUs upon treatment with either THAM-3ФG or chlorhexidine (Figure S1). To test the efficacy of this assay, antibiotics with known mechanisms of action that are not membrane related were also tested. Ampicillin and gentamicin, which are cell wall and protein synthesis inhibitors, respectively, showed no increase in A260 measurements over the same time period at four times their MIC (Figure S2). It should be noted that the cell densities used in these assays were 4 logs higher than those used in MIC determination in order to provide sufficient sensitivity for the A260 measurements; this may explain the slight increase in MIC seen in this assay compared to data shown in Table 1.
Figure 3. Cytoplasmic Leakage.

Change in absorbance at 260(ΔA260) MRSA culture supernatants upon treatment with Chlorhexidine (a) and THAM-3ФG (b). Both compounds showed an immediate increase in A260 within a half hour after treatment at a concentration of 8 mg/L (4×MIC). Note that plots represent the difference between the absorbance of the supernatant from non-treated and treated cultures at each time point. *P-value<0.05, **p-value<0.01, and ***p-value<0.001 versus USA300 as analyzed by one-way ANOVA.
MRSA Challenge
THAM-3ΦG was chosen for the in vivo challenge experiments, because it had the best combination of antibacterial activity and low toxicity (Tables 1–3). In vivo toxicity studies were carried out prior to the start of experiments in mice challenged with MRSA. Groups of three Balb/c mice each were injected i.p. with 0.1 ml of 0, 10, 50, and 100 µM THAM-3ФG in PBS. The highest in vivo dose represented 0.36 mg/kg. Blood was drawn from the mice at two hours post injection and submitted for a standard panel of serum chemistry and enzyme levels. The chemistry and enzyme levels were not elevated for any of the concentrations tested compared to the mock-treated control group (data not shown).
Since the compound was not overtly toxic, we tested its ability to reduce bacterial burden in vivo using a mouse model of invasive staphylococcal disease [16], [17]. Mice were treated with 100 µM THAM-3ФG either at the same time or one hour after an i.p. challenge with a lethal dose of MRSA. The mice treated with THAM-3ФG immediately after the MRSA challenge demonstrated significantly reduced bacterial burdens in intraperitoneal lavage, kidneys, and heart. They averaged an approximately 3-log reduction in both organs and IP lavage compared to PBS treated mice (Figure 4). Treatment of mice with THAM-3ФG one hour after MRSA infection also reduced bacterial burdens compared to untreated mice at 8 h post infection. Mean CFUs were reduced 20-fold in kidneys, 3-fold in heart and 2-fold in IP lavages (Figure 4); these values were statistically significant for the kidney samples.
Figure 4. THAM-3ΦG reduces MRSA burden in vivo during peritonitis.
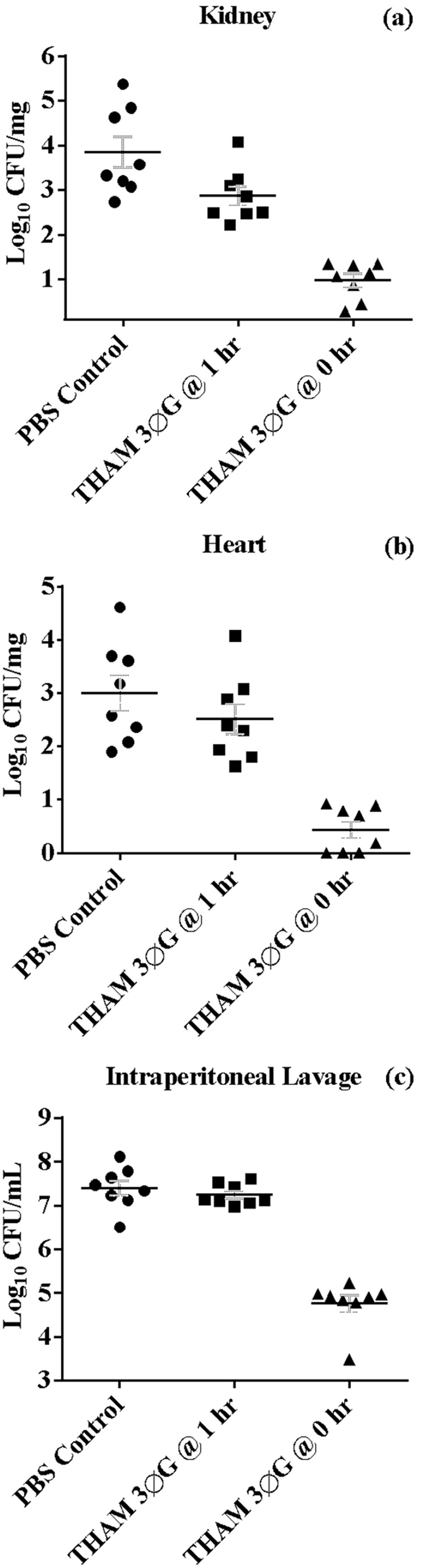
Mice were treated with PBS control or with THAM trisphenylguanide immediately after or at one hour post intraperitoneal infection. Bacterial burden was evaluated in the kidney (A) and heart (B); localized infection was assessed with peritoneal lavage fluid (C). The horizontal bar is the mean of the eight mice per treatment group; error bars represent standard error of the mean. Reductions in burden were significant (t-test) in kidneys for both the 1 hr (p = 0.028) and 0 hr (p = 2×10−6) treatments. Reductions were significant in the heart (p = 4×10−5) and IP lavage (p = 6×10−8) only for the 0 hr treatments.
Discussion
Initial screening of our library of guanide, biguanide, and phenylguanide compounds showed that the higher molecular weight compounds had greater antibacterial activity. The derivatives of the smaller amine starting materials (1,4-butanediamine, 1,6-hexanediamine), as well as the short chain melamine derivatives (trisethylaminomelamine and trisbutylaminomelamine), and the small dendrimer PAMAM G0 were inactive at the highest concentration tested (100 µM).
A wide variation in activity vs. species of bacteria for each of the compounds was observed. The compounds ranged from being inactive against all of the bacteria tested to having antibacterial activity against both Gram-positive and Gram-negative species in the bacterial viability assays. However, none of the compounds exhibited the broad antibacterial action of chlorhexidine (MIC values ≤16 mg/L against all of the pathogens) when the antibacterial activity was tested in a complex medium (LB) against actively growing bacteria. However, THAM-3ΦG was as effective against S. aureus (including MRSA) as chlorhexidine and slightly less toxic to mammalian cells; this increased selectivity for MRSA gives it potential for further development.
When all the compounds in Table 1, except TOAM and its derivatives, were tested against two eukaryotic pathogens, Candida albicans and Acanthamoeba castellanii, none of them showed any substantial toxicity at 100 µM against Candida, or at 200 µM against Acanthamoeba, whereas chlorhexidine and PHMB had IC50 values of approximately 10 µM and 10 µg/ml, respectively, against Acanthamoeba (data not shown). Others have previously shown that PHMB and chlorhexidine are effective against Acanthamoeba [19], [20], [21], and this provides further evidence for the selectivity of THAM-3ΦG compared to the biguanides. The reduced toxicity toward eukaryotic cells is consistent with the lack of in vivo toxicity observed in mice. However, given the preliminary nature of these experiments, it is difficult to know how the in vitro toxicity compares to the in vivo toxicity.
The mechanism of action for the commonly used biguanide antiseptics, chlorhexidine and PHMB, is not well-defined. Although they are thought to interact with the membrane surfaces [22] resulting in leakage of cellular contents, they do not have long alkyl chains that intercalate into the lipid bilayer like those of the common antibacterial cationic quaternary amine compounds. A PHMB study [23] found induction of the rhs genes in E. coli, implicating enzymes involved in the repair/binding of nucleic acids, and another study reported cooperative binding of PHMB to nucleic acids [24]. A chlorhexidine study [25] found that the mechanism of action in P. aeruginosa may be multifaceted, involving changes in outer membrane permeability, the attenuation of virulence and environmental adaptation processes, and energy deprivation through the repression of genes involved in aerobic respiration. Although the fact that some of our initially screened compounds were biguanides led us to infer a similarity to PHMB and chlorhexidine, the biguanides we synthesized had little or no antibacterial activity. This observation and the differences in specificity between the phenylguanides and chlorhexidine lead us to hypothesize that THAM-3ΦG may act by somewhat different mechanisms than chlorhexidine or PHMB. Recently described compounds with multiple guanide groups, p-guanidinoethylcalix [4] arene [26], and Akacid plus, a mixture of polymeric guanides [27], [28], also exhibit broad spectrum activity against bacterial and eukaryotic pathogens, unlike THAM-3ΦG.
Since chlorhexidine has been observed to cause leakage of cellular contents [22], we compared its action to that of THAM-3ΦG in a cytoplasmic leakage assay. For both compounds, leakage of molecules absorbing at 260 nm was observed at concentrations that did not decrease the number of viable cells, indicating that membrane leakage precedes cell death and likely causes it at higher concentrations. In summary, THAM-3ΦG may act to disrupt bacterial membranes, but it appears to be more specific against S. aureus than would be predicted by such a non-specific interaction. Future studies with our compounds are aimed at further exploring its mechanisms of action.
The structural and mechanistic similarities to chlorhexidine suggested potential applications of our compounds as topical microbicides. However, quantifying effects on topical bacterial infections in vivo is difficult. Therefore, a preliminary study was done using inhibition of a systemic infection as a model for activity in a complex in vivo environment. The results from this initial in vivo antibacterial assay performed with THAM-3ΦG in mice infected with lethal doses of MRSA were promising. Reductions in bacterial load were noted when the compound was given immediately after the bacterial inoculation as well as when the compound was given one hour post infection. The smaller reduction in load seen in the latter case was likely due to the bacteria experiencing a lower exposure (concentration×time) to the compound when they had an hour to disseminate before it was administered. Although this experiment did not assess the effective dose (ED50), it demonstrated that the compound still exhibited antibacterial properties in a complex in vivo environment. Furthermore, the administration of only one dose at only one time point was likely sub-optimal, since the pharmacokinetics are unknown. The proposed use of these compounds is to treat skin MRSA infections; further investigation will require optimizing dosage and topical application regimens in additional studies with appropriate infection models, such as skin and soft-tissue. Nonetheless, the current study demonstrates promise for in vivo application for this compound and potentially others with related structures.
Supporting Information
Cell viability. The number of viable cells was monitored throughout the absorbance assay. MRSA treated with Chlorhexidine (a) or THAM-3ФG (b) showed no significant reductions in CFUs.
(TIF)
Cell leakage with control antibiotics. Ampicillin (a) and gentamicin (b) were tested to confirm the efficacy of the absorbance assay as a means to monitor potential membrane disruption. Significant increases in A260 were not seen in either case at concentrations ranging from 1× to 4×MIC (The MICs determined using the assay described in Material and Methods were 32 mg/L for ampicillin and 0.5 mg/L for gentamicin, respectively).
(TIF)
Bacterial viability assays. Bacteria were grown at 37°C to mid-log phase (OD600 ∼0.4). The bacteria were diluted to approximately 104 to 105 colony forming units (CFU) per milliliter and added to a solution of phosphate buffered saline (PBS, 16 mM phosphate, 0.14 M NaCl, pH 7.2) containing the compound at varied concentrations. Samples were incubated at 37°C for one hour and then plated on Luria Bertani (LB) agar at appropriate dilutions in PBS. The plates were incubated overnight at 37°C and colonies enumerated the following day. IC50s were calculated from graphs of CFU as a function of compound concentration. Chlorhexidine was used as a positive control. Included are those guanide (G), biguanide (BG), phenylguanide (ΦG) compounds that had some antibacterial activity in an initial screen at 30 and 100 µM, as well as their and parent amines. Compounds synthesized from 1,4-butanediamine, 1,6-hexanediamine, trisethylaminomelamine, trisbutylaminomelamine, and the dendrimer PAMAM (G0) were not active against E. coli, P aeruginosa, or S. aureus so they were not tested against the remaining bacteria (data not shown).
(DOC)
Acknowledgments
We thank Garth James, Mike Franklin, and Karen Kelsey for providing bacterial strains, and Jean Starkey for MDA-MB-231 cells. We thank Ryan Bachover for technical assistance.
Funding Statement
This work was supported by institutional funds and by the following US Public Health Service grants: AI076965 (M.E.L.) from the National Institute of Allergy and Infectious Diseases, CoBRE grants P20RR024237 (M.T.) and P20RR020185 (C.C., E. E. and J.M.V.) from the National Center for Research Resources. S.K.W., R.L.W., and T.J.W. were supported by fellowships from INBRE grant P20RR016455 from the National Center for Research Resources. M.B.A. was supported by a fellowship funded by the Howard Hughes Medical Institute. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Boucher HW, Talbot GH, Bradley JS, Edwards JE, Gilbert D, et al. (2009) Bad bugs, no drugs: no ESKAPE! An update from the Infectious Diseases Society of America. Clin Infect Dis 48: 1–12. [DOI] [PubMed] [Google Scholar]
- 2. Lowy FD (2003) Antimicrobial resistance: the example of Staphylococcus aureus. J Clin Invest 111: 1265–1273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Vancomycin-resistant Staphylococcus aureus–Pennsylvania, 2002. MMWR Morb Mortal Wkly Rep 51: 902. [PubMed] [Google Scholar]
- 4. Wilkinson RA, Pincus SH, Shepard JB, Walton SK, Bergin EP, et al. (2011) Novel compounds containing multiple guanide groups that bind the HIV coreceptor CXCR4. Antimicrob Agents Chemother 55: 255–263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Wilkinson RA, Pincus SH, Song K, Shepard JB, Weaver A, et al. (2013) Improved guanide compounds which bind the CXCR4 co-receptor and inhibit HIV infection. Bioorg Med Chem Lett 23: 2197–2201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shepard JB, Wilkinson RA, Starkey JR, Teintze M (2013) Novel guanide substituted compounds bind to CXCR4 and inhibit breast cancer metastasis. Anti-Cancer Drugs in press. [DOI] [PubMed]
- 7. Sahm DF, Kissinger J, Gilmore MS, Murray PR, Mulder R, et al. (1989) In vitro susceptibility studies of vancomycin-resistant Enterococcus faecalis. Antimicrob Agents Chemother 33: 1588–1591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Nair D, Memmi G, Hernandez D, Bard J, Beaume M, et al. (2011) Whole-genome sequencing of Staphylococcus aureus strain RN4220, a key laboratory strain used in virulence research, identifies mutations that affect not only virulence factors but also the fitness of the strain. J Bacteriol 193: 2332–2335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Diep BA, Gill SR, Chang RF, Phan TH, Chen JH, et al. (2006) Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 367: 731–739. [DOI] [PubMed] [Google Scholar]
- 10. Chopra S, Torres-Ortiz M, Hokama L, Madrid P, Tanga M, et al. (2010) Repurposing FDA-approved drugs to combat drug-resistant Acinetobacter baumannii. J Antimicrob Chemother 65: 2598–2601. [DOI] [PubMed] [Google Scholar]
- 11. Tien HC, Battad A, Bryce EA, Fuller J, Mulvey M, et al. (2007) Multi-drug resistant Acinetobacter infections in critically injured Canadian forces soldiers. BMC Infect Dis 7: 95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Diep BA, Sensabaugh GF, Somboonna N, Carleton HA, Perdreau-Remington F (2004) Widespread skin and soft-tissue infections due to two methicillin-resistant Staphylococcus aureus strains harboring the genes for Panton-Valentine leucocidin. J Clin Microbiol 42: 2080–2084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, et al. (2007) Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 298: 1763–1771. [DOI] [PubMed] [Google Scholar]
- 14. Andrews JM (2001) Determination of minimum inhibitory concentrations. J Antimicrob Chemother 48 Suppl 1 5–16. [DOI] [PubMed] [Google Scholar]
- 15. Boukamp P, Petrussevska RT, Breitkreutz D, Hornung J, Markham A, et al. (1988) Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J Cell Biol 106: 761–771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Watkins RL, Pallister KB, Voyich JM (2011) The SaeR/S gene regulatory system induces a pro-inflammatory cytokine response during Staphylococcus aureus infection. PLoS One 6: e19939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Watkins RL, Zurek OW, Pallister KB, Voyich JM (2013) The SaeR/S two-component system induces interferon-gamma production in neutrophils during invasive Staphylococcus aureus infection. Microbes Infect 15: 749–754. [DOI] [PubMed] [Google Scholar]
- 18. Davies GE, Francis J, Martin AR, Rose FL, Swain G (1954) 1: 6-Di-4′-chlorophenyldiguanidohexane (hibitane); laboratory investigation of a new antibacterial agent of high potency. Br J Pharmacol Chemother 9: 192–196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Khunkitti W, Hann AC, Lloyd D, Furr JR, Russell AD (1998) Biguanide-induced changes in Acanthamoeba castellanii: an electron microscopic study. J Appl Microbiol 84: 53–62. [DOI] [PubMed] [Google Scholar]
- 20. Khunkitti W, Lloyd D, Furr JR, Russell AD (1998) Acanthamoeba castellanii: growth, encystment, excystment and biocide susceptibility. J Infect 36: 43–48. [DOI] [PubMed] [Google Scholar]
- 21. Turner NA, Russell AD, Furr JR, Lloyd D (2004) Resistance, biguanide sorption and biguanide-induced pentose leakage during encystment of Acanthamoeba castellanii. J Appl Microbiol 96: 1287–1295. [DOI] [PubMed] [Google Scholar]
- 22. Gilbert P, Moore LE (2005) Cationic antiseptics: diversity of action under a common epithet. J Appl Microbiol 99: 703–715. [DOI] [PubMed] [Google Scholar]
- 23. Allen MJ, White GF, Morby AP (2006) The response of Escherichia coli to exposure to the biocide polyhexamethylene biguanide. Microbiology 152: 989–1000. [DOI] [PubMed] [Google Scholar]
- 24. Allen MJ, Morby AP, White GF (2004) Cooperativity in the binding of the cationic biocide polyhexamethylene biguanide to nucleic acids. Biochem Biophys Res Commun 318: 397–404. [DOI] [PubMed] [Google Scholar]
- 25. Nde CW, Jang HJ, Toghrol F, Bentley WE (2009) Global transcriptomic response of Pseudomonas aeruginosa to chlorhexidine diacetate. Environ Sci Technol 43: 8406–8415. [DOI] [PubMed] [Google Scholar]
- 26. Grare M, Dibama HM, Lafosse S, Ribon A, Mourer M, et al. (2010) Cationic compounds with activity against multidrug-resistant bacteria: interest of a new compound compared with two older antiseptics, hexamidine and chlorhexidine. Clin Microbiol Infect 16: 432–438. [DOI] [PubMed] [Google Scholar]
- 27. Buxbaum A, Kratzer C, Graninger W, Georgopoulos A (2006) Antimicrobial and toxicological profile of the new biocide Akacid plus. J Antimicrob Chemother 58: 193–197. [DOI] [PubMed] [Google Scholar]
- 28. Kratzer C, Tobudic S, Graninger W, Buxbaum A, Georgopoulos A (2006) In vitro antimicrobial activity of the novel polymeric guanidine Akacid plus. J Hosp Infect 63: 316–322. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Cell viability. The number of viable cells was monitored throughout the absorbance assay. MRSA treated with Chlorhexidine (a) or THAM-3ФG (b) showed no significant reductions in CFUs.
(TIF)
Cell leakage with control antibiotics. Ampicillin (a) and gentamicin (b) were tested to confirm the efficacy of the absorbance assay as a means to monitor potential membrane disruption. Significant increases in A260 were not seen in either case at concentrations ranging from 1× to 4×MIC (The MICs determined using the assay described in Material and Methods were 32 mg/L for ampicillin and 0.5 mg/L for gentamicin, respectively).
(TIF)
Bacterial viability assays. Bacteria were grown at 37°C to mid-log phase (OD600 ∼0.4). The bacteria were diluted to approximately 104 to 105 colony forming units (CFU) per milliliter and added to a solution of phosphate buffered saline (PBS, 16 mM phosphate, 0.14 M NaCl, pH 7.2) containing the compound at varied concentrations. Samples were incubated at 37°C for one hour and then plated on Luria Bertani (LB) agar at appropriate dilutions in PBS. The plates were incubated overnight at 37°C and colonies enumerated the following day. IC50s were calculated from graphs of CFU as a function of compound concentration. Chlorhexidine was used as a positive control. Included are those guanide (G), biguanide (BG), phenylguanide (ΦG) compounds that had some antibacterial activity in an initial screen at 30 and 100 µM, as well as their and parent amines. Compounds synthesized from 1,4-butanediamine, 1,6-hexanediamine, trisethylaminomelamine, trisbutylaminomelamine, and the dendrimer PAMAM (G0) were not active against E. coli, P aeruginosa, or S. aureus so they were not tested against the remaining bacteria (data not shown).
(DOC)