

Abstract
Peptide YY (PYY) is a gut hormone that activates the G protein-coupled neuropeptide Y (NPY) receptors, and because of its appetite reducing actions, it is evaluated as an antiobesity drug candidate. The C-terminal tail of PYY is crucial for activation of the NPY receptors. Here, we describe the design and preparation of a series of PYY(3–36) depsipeptide analogues, in which backbone amide-to-ester modifications were systematically introduced in the C-terminal. Functional NPY receptor assays and circular dichroism revealed that the ψ(CONH) bonds at positions 30–31 and 33–34 are particularly important for receptor interaction and that the latter is implicated in Y2 receptor selectivity.
Keywords: Peptide YY, PYY, amide-to-ester modifications, neuropeptide Y, NPY
Peptide YY (PYY), neuropeptide Y (NPY), and pancreatic peptide (PP) constitute the NPY family of peptides, which are involved in the regulation of gastro-intestinal functions, blood pressure, and feeding behavior.1 These peptides share a high degree of sequence homology and a common hairpin-like three-dimensional structure, known as the PP-fold.2 The peptides exert their physiological functions by interacting with four human G protein-coupled receptors (GPCRs), the NPY receptors, which belong to the class A superfamily and are designated Y1, Y2, Y4, and Y5.3
PYY is released postprandially from enteroendocrine L-cells as PYY(1–36), which is cleaved into PYY(3–36) by dipeptidyl peptidase IV.4 Whereas PYY(1–36) activates Y1, Y2, and Y5 receptors, removal of the N-terminal dipeptide Tyr–Pro results in increased selectivity for the Y2 receptor.5 Activation of Y2 receptors in the hypothalamic arcuate nucleus,6 brainstem,7 and vagal nerve fibers8 has been shown to be implicated in decreased food intake observed upon peripheral administration of PYY(3–36).6,9−12 These anorectic properties make PYY(3–36) a potential antiobesity agent.13
Alanine scans of both NPY and PYY have shown that the major determinants for binding to the Y2 receptor are located in the C-terminal of the peptide.14,15 The X-ray crystal structure of a class A chemokine GPCR bound to a cyclic peptide ligand demonstrated that several backbone amide functionalities of the peptide ligand are implicated in the interaction with the receptor.16 While the significance of the side chains of the C-terminal residues of PYY for receptor interaction is well established through numerous structure–activity relationship (SAR) studies,17 little is known about the contribution of backbone amide interactions for receptor recognition. In one study, substitution of backbone amides with reduced amide bonds, CH2NH, in the C-terminus of a truncated PYY derivative, PYY(22–36), demonstrated that this modification generally reduced affinities toward the Y2 receptor, except for the linkage between Arg35 and Tyr36, where no change in affinity was observed.18 This may indicate that the corresponding carbonyl groups are involved in receptor binding. However, the reduced affinities may also be caused by an increased flexibility, resulting in an enhanced magnitude of the conformational entropic penalty associated with receptor binding. Furthermore, reduced amide bonds do not address the potential interactions of the amide NH functionalities. Substitution of the C-terminal backbone amide bonds with N-methylated amide bonds showed that an N-methyl group was only allowed on Arg25 and Arg35 leading to moderate changes in affinity to the Y2 receptor.19 Yet, N-methyl peptide bonds are structurally different from amide bonds due to introduction of additional steric bulk and an increased tendency to adopt the cis conformation. In contrast, ester bonds are structurally similar to peptide bonds and address interactions of both the amide carbonyl and NH functionalities.20 We therefore decided to systematically explore the effect of the isosteric backbone amide-to-ester modifications in the C-terminus of PYY(3–36) on receptor interaction.
Introduction of amide-to-ester modifications is accomplished by replacing the amino acid of interest with the corresponding α-hydroxy acid.20 In the synthesis of such depsipeptides there are two main challenges: (i) only six of 19 α-hydroxy acids are commercially available and (ii) introduction of the α-hydroxy acid is not trivial. Here, we wanted to introduce backbone amide-to-ester mutations between the seven C-terminal residues, thus generating six depsipeptide analogues of PYY(3–36) (Figure 1). For this, four α-hydroxy acids corresponding to Arg, Gln, Tyr, and Thr were prepared, whereas α-hydroxy Val is commercially available.
Figure 1.
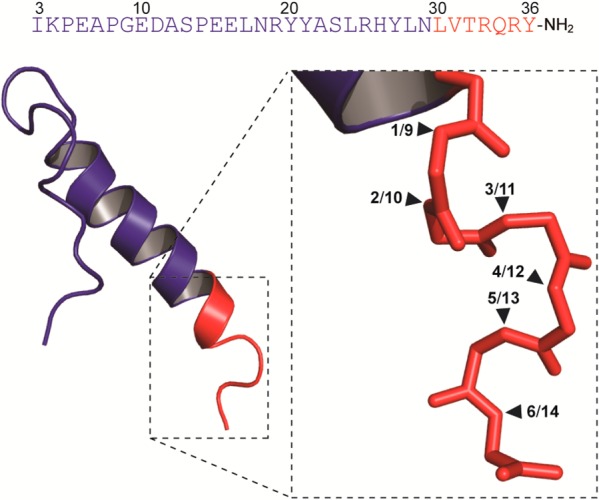
Sequence and NMR structure (PDB: 2DF0) of PYY(3–36) with the seven C-terminal residues in red and the remaining part of the sequence in blue. Close-up view shows sticks representation of the C-terminal backbone (without hydrogens) with the positions of NH to O modifications marked with black triangles and peptide numbers. Peptides 1–6 are hepta-depsipeptides used to generate full length depsipeptides 9–14, respectively.
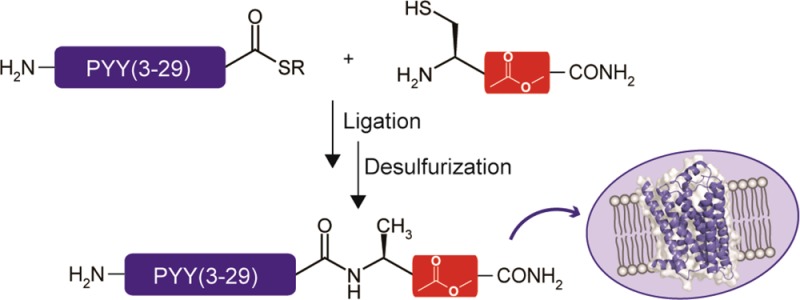
Rather than preparing 34-mer depsipeptides on solid-phase, we envisaged that a semisynthetic methodology based on native chemical ligation (NCL)21 could be exploited. This could be achieved by expressing a common precursor peptide thioester and subsequently ligate with hepta-depsipeptides bearing an N-terminal Cys. These hepta-depsipeptides 1–6 were synthesized by Fmoc- or Boc-based solid-phase peptide synthesis (SPPS) (Scheme 1 and Table S1, Supporting Information). The α-hydroxy acids were introduced by coupling with HOBt/DIC, as previously described,22 and the ester bonds formed by coupling the following amino acid using DIC/DMAP.23,24 This provided hepta-depsipeptides 1, 4, and 6 (Scheme 1), but synthesis of the remaining target peptides using this methodology did not succeed. In the synthesis of hepta-depsipeptide 2, incomplete reaction and epimerization was observed when coupling Boc-Val with α-hydroxy Thr. Thus, alternative strategies were examined, and the most efficient coupling was achieved using triple couplings with MSNT/N-methylimidazole (Table S2, Supporting Information).25 The challenge in the synthesis of 3 and 5 was that coupling of α-hydroxy Arg was incomplete; thus, PyBOP was used as a coupling reagent (Table S3, Supporting Information). In the subsequent ester formation, very low coupling yields were observed, and for the synthesis of 3, formation of the ester bond between Thr and α-hydroxy Arg was achieved by consecutive MSNT/N-methylimidazole couplings (Table S3, Supporting Information). The same approach was used for ester bond formation between Gln and α-hydroxy Arg in depsipeptide 5, although with decreased coupling times (Table S4, Supporting Information).
Scheme 1. General Synthesis of Hepta-depsipeptides 1–6.

Hepta-depsipeptides were prepared by either Fmoc- or Boc-based SPPS. Ester bonds were incorporated by coupling of α-hydroxy acids using HOBt/DIC/NEM for hepta-depsipeptides 1, 2, 4, and 6 and PyBOP/HOBt/DIPEA for hepta-depsipeptides 3 and 5. The following amino acid was coupled using DIC/DMAP/NEM for hepta-depsipeptides 1, 4, and 6 and MSNT/NMI for hepta-depsipeptides 2, 3, and 5. Fmoc-SPPS: R1 = tBu, R2 = Pbf. Boc-SPPS: R1 = 2-Br-Z, R2 = Tos.
To generate the PYY(3–36) depsipeptides, a recombinantly expressed PYY(3–29) α-thioester (7)26 was ligated to hepta-depsipeptides 1–6 (Figure 2A). This leaves a Cys at the ligation junction, which is subsequently reduced to Ala, knowing that the Leu30Ala mutation does not affect PYY(3–36) Y2 receptor potency.14,15 Initially, a control peptide, PYY(3–36), A30 (8), was prepared either by SPPS or semisynthesis, the latter validating the semisynthetic strategy. The PYY(3–36), A30 depsipeptides 9–14 were prepared by ligation of 7 with 1–6, respectively, using thiophenol as catalyst (Figure 2B). The thiophenol was removed by extraction with diethyl ether prior to conversion of Cys into Ala by radical desulfurization27 (Figure 2C). The semisynthetic analogues 9–14 were purified by semipreparative HPLC, resulting in purities of 98–99%. For analogues 12–14, some hydrolysis of the ester bonds was observed after desulfurization. Whereas the resulting impurities of analogues 12 and 14 were easily removed, for analogue 13, slightly optimized ligation and desulfurization conditions had to be used (see Supporting Information). Because of the observed decrease in hydrolytic stability during preparation, we also examined the stability of the depsi-peptide analogues under the conditions used for the functional assays, and gratifyingly, no degradation was observed for any of the analogues (Figure S1, Supporting Information).
Figure 2.
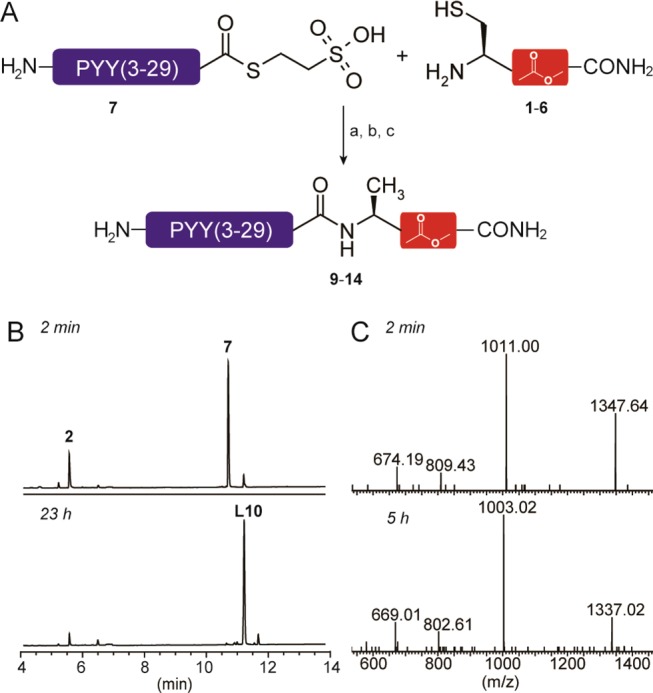
Generation of depsipeptide analogues of PYY(3–36), A30. (A) Scheme of the NCL based synthesis of the PYY(3–36), A30 depsipeptide analogues. Reagents and conditions: a, 1.8 equiv of 1–6, 0.6% thiophenol, pH 7.5, 5 °C; b, diethyl ether extraction; c, MESNa, VA-61, TCEP, 37 °C. (B) UPLC trace at 2 min and 23 h of ligation between 2 and 7, (L10: PYY(3–36)[COO31–32], 30C). (C) MS spectra at 2 min and 5 h of desulfurization of 10 (expected average masses: L10 = 4040.50 Da; 10 = 4008.43 Da).
The structure of PYY(3–36) is characterized by having an amphipathic α-helix from residues 15–32,28 which is expected to be important for receptor activation.29,30 The consequences of the amide-to-ester modifications on the integrity of the secondary structure were evaluated by circular dichroism (CD). The spectra were recorded in a sodium acetate buffer at pH 4.6, similar to the conditions used for determining the NMR structure of PYY(3–36).28 The CD spectra of the analogue with the Leu30Ala mutation, 8, generated by SPPS and semisynthesis were as expected identical (Figure S2, Supporting Information). Generally, the CD spectra for all tested peptides were highly similar with minima at 208 and 222 nm as well as maxima around 193 nm (Figure 3), which are characteristic for α-helical structures, and changes in molar CD, Δε, at 222 nm can be used to evaluate alterations in helicity.31
Figure 3.
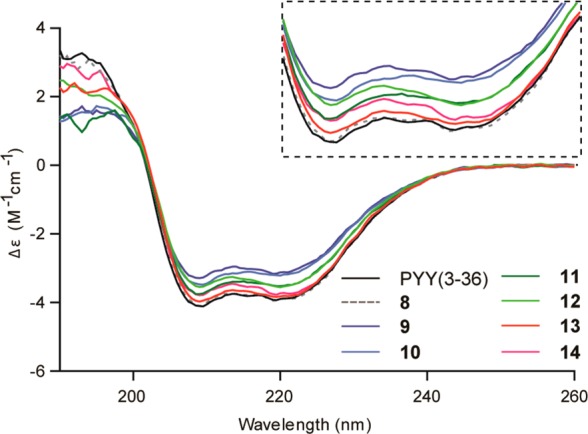
Circular dichroism (CD) of depsipeptide analogues 9–14 and controls in aqueous buffered solution (15 μM peptide, pH 4.6). The insert shows a close-up view of the spectra in the region 205–230 nm. Spectra are presented as mean of two independent experiments.
The helicity of analogue 8, bearing the Leu30Ala mutation, was not affected, but introduction of the amide-to-ester modification in analogues 9 and 10 reduced the helix content by approximately 20%. In these two analogues, the ester bond is positioned as part of the α-helix, as observed in the NMR structure of PYY(3–36) (Figure 1).28 In analogues 11 and 12, where the ester mutations are outside the α-helix, the helicity was 10% reduced, as estimated by CD. Thus, the ψ(CONH32–33) and ψ(CONH33–34) bonds may be important for stabilizing the α-helix, although they are located in the unstructured C-terminal. Finally, there was no change in the CD spectra of analogues 13 and 14, relative to PYY(3–36).
The consequence of insertion of amide-to-ester modifications in the C-terminal of PYY(3–36) for interaction with the NPY receptors was evaluated using a functional cAMP biosensor assay employing HEK293 cells expressing the human Y1, Y2, Y4, or Y5 receptor subtypes (Table 1). As expected from previous studies,14,15 the Leu30Ala mutation did not affect PYY(3–36) potency at the Y2 receptor. However, the mutation resulted in increased selectivity toward the Y2 receptor with approximate 7- and 20-fold decreases in potency observed for SPPS generated 8 and semisynthetic 8 relative to PYY(3–36) at the Y5 and Y1 receptor, respectively. Notably, the EC50 values of 8 were similar at all four receptor subtypes, demonstrating that the semisynthetic product is biologically equivalent to the fully synthetic product. For the depsipeptide analogues 9–14, the most pronounced effect on potency was observed for analogue 9 with an ester bond between Ala30 and Val31 as well as analogue 12 with an ester bond between Arg33 and Gln34. An approximate 200- and 60-fold reduction in potency at the Y2 receptor compared to 8 was observed, respectively. For analogue 9, reduced potency was further observed at the other three receptor subtypes with EC50 values above 500 nM. Interestingly, the ester bond in analogue 12 did not affect potency at Y1 and Y4, and only reduced potency at Y5 by 3-fold. Thus, whereas the backbone amide between positions 30 and 31 are important for interaction with all four receptor subtypes, the amide between positions 33 and 34 are only important for Y2 and to less extent Y5 interaction. In contrast, introduction of an ester bond at the most C-terminal position between Arg35 and Tyr36 in analogue 14 did not alter the potency at any of the receptor subtypes. Negligible to moderate decreases in Y2 receptor potencies were observed for analogues 10, 11, and 13.
Table 1. Potencies of Depsipeptide Analogues 9–14 at the NPY Receptors.
| EC50 (nM)a |
||||
|---|---|---|---|---|
| analogue | Y1 | Y2 | Y4 | Y5 |
| PYY(3–36) | 5.6 ± 0.8 | 1.0 ± 0.2 | 141 ± 40 | 8.2 ± 2.7 |
| PYY(3–36), A30 (8, SPPS) | 164 ± 34 | 1.0 ± 0.1 | 207 ± 47 | 42 ± 13 |
| PYY(3–36), A30 (8, semisynthesis) | 135 ± 30 | 1.3 ± 0.3 | 185 ± 38 | 37 ± 8 |
| PYY(3–36)[COO30–31], A30 (9) | >500 | 228 ± 46 | >500 | >500 |
| PYY(3–36)[COO31–32], A30 (10) | >500 | 8.4 ± 1.0 | >500 | >500 |
| PYY(3–36)[COO32–33], A30 (11) | >500 | 3.6 ± 0.8 | >500 | 112 ± 51 |
| PYY(3–36)[COO33–34], A30 (12) | 144 ± 3 | 62 ± 19 | 161 ± 35 | 172 ± 34 |
| PYY(3–36)[COO34–35], A30 (13) | 380 ± 58 | 2.2 ± 0.4 | 173 ± 20 | 187 ± 50 |
| PYY(3–36)[COO35–36], A30 (14) | 222 ± 60 | 0.3 ± 0.1 | 254 ± 42 | 29 ± 4 |
EC50 values are shown as mean ± SEM determined from at least three measurements.
The comparable effect on α-helicity of analogues 9 and 10, but varying influence on the potency at the Y2 receptor (200- vs 8-fold reduction), indicates that more than a structural effect may account for the highly reduced potency observed for analogue 9. Thus, the ψ(CONH30–31) bond may interact directly with the Y2 receptor subtype. Similar discrepancies are observed for analogues 11 and 12, indicating that the ψ(CONH33–34) bond may interact directly with the Y2 receptor subtype. Interestingly, this interaction is specific for the Y2 receptor since no or little effect in potency was observed at the other receptor subtypes. These data suggests that the ψ(CONH33–34) bond contributes to the increased selectivity of PYY(3–36) toward the Y2 versus the other receptor subtypes.
In conclusion, we have incorporated backbone amide-to-ester modifications in the C-terminal of PYY(3–36) using a semisynthetic strategy that results in a Leu30Ala substitution. A comparison of data from functional receptor assays with CD indicated that two backbone amide functionalities, namely, ψ(CONH30–31) and ψ(CONH33–34), are important for interaction of PYY(3–36) with the G protein-coupled Y2 receptor. The ψ(CONH30–31) bond may interact directly with all of the NPY receptors, whereas the ψ(CONH33–34) bond may interact with the Y2 receptor subtype in a selective manner. The results herein emphasize the significance of backbone hydrogen bonds in the interaction between peptide ligands and GPCRs and should be considered when developing subtype selective analogues and stabilized PYY(3–36) derivatives for obesity treatment.
Acknowledgments
Dr. N. S. Hansen and S. Padrah (University of Copenhagen) are acknowledged for preparation of the Boc-based α-hydroxy acids. We thank M. Mørkenborg (Novo Nordisk, Måløv) for preparation of synthetic PYY(3–36) and PYY(3–36), C30. We also thank A. L. Almind (Novo Nordisk, Måløv) for performing the NPY receptor assays and Berit B. Hansen (Novo Nordisk, Måløv) is acknowledged for performing circular dichroism.
Glossary
ABBREVIATIONS
- 2-Br-Z
2- bromobenzyloxycarbonyl
- Boc
tert-butyloxycarbonyl
- CD
circular dichroism
- DIC
N,N′-diisopropylcarbodiimide
- DIPEA
diisopropylethylamine
- DMAP
4-dimethylaminopyridine
- Fmoc
9-fluorenylmethyloxycarbonyl
- GPCR
G protein-coupled receptor
- HOBt
N-hydroxybenzotriazole
- MESNa
sodium 2-mercaptoethane-sulfonate
- MSNT
1-(mesitylene-2-sulfonyl)-3-nitro-1H-1,2,4-triazole
- NEM
N-ethylmorpholine
- NPY
neuropeptide Y
- Pbf
2,2,4,6,7-pentamethyldihydrobenzofuran-5-sulfonyl
- PyBOP
benzotriazole-1-yl-oxy-tris-pyrrolidino-phosphonium hexafluorophosphate
- PYY
peptide YY
- SPPS
solid-phase peptide synthesis
- tBu
tert-butyl
- TCEP
tris(2-carboxyethyl)phosphine
- Tos
tosyl
- VA-61
2,2′-azobis[2-(2-imidazolin-2-yl)propane]
Supporting Information Available
Experimental procedures for the synthesis and characterization of peptides, the in vitro cell-based functional assays and circular dichroism as well as data for optimization of ester bond formations and stability testing. This material is available free of charge via the Internet at http://pubs.acs.org.
We are grateful to the Danish Agency for Science, Technology and Innovation (Ministry of Science, Innovation and Higher Education), for the Ph.D. fellowship to L.A.
The authors declare no competing financial interest.
Supplementary Material
References
- Brothers S. P.; Wahlestedt C. Therapeutic potential of neuropeptide Y (NPY) receptor ligands. EMBO Mol. Med. 2010, 2, 429–439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blundell T. L.; Pitts J. E.; Tickle I. J.; Wood S. P.; Wu C. W. X-ray analysis (1.4-Å resolution) of avian pancreatic polypeptide: Small globular protein hormone. Proc. Natl. Acad. Sci. U.S.A. 1981, 78, 4175–4179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck-Sickinger A. G. Structural characterization and binding sites of G-protein-coupled receptors. Drug Discovery Today 1996, 1, 502–513. [Google Scholar]
- Mentlein R.; Dahms P.; Grandt D.; Kruger R. Proteolytic processing of neuropeptide Y and peptide YY by dipeptidyl peptidase IV. Regul. Pept. 1993, 49, 133–144. [DOI] [PubMed] [Google Scholar]
- Michel M. C.; Beck-Sickinger A.; Cox H.; Doods H. N.; Herzog H.; Larhammar D.; Quirion R.; Schwartz T.; Westfall T. XVI. International Union of Pharmacology recommendations for the nomenclature of neuropeptide Y, peptide YY, and pancreatic polypeptide receptors. Pharmacol. Rev. 1998, 50, 143–150. [PubMed] [Google Scholar]
- Batterham R. L.; Cowley M. A.; Small C. J.; Herzog H.; Cohen M. A.; Dakin C. L.; Wren A. M.; Brynes A. E.; Low M. J.; Ghatei M. A.; Cone R. D.; Bloom S. R. Gut hormone PYY3–36 physiologically inhibits food intake. Nature 2002, 418, 650–654. [DOI] [PubMed] [Google Scholar]
- Larsen P. J.; Kristensen P. Distribution of neuropeptide Y receptor expression in the rat suprachiasmatic nucleus. Mol. Brain Res. 1998, 60, 69–76. [DOI] [PubMed] [Google Scholar]
- Koda S.; Date Y.; Murakami N.; Shimbara T.; Hanada T.; Toshinai K.; Niijima A.; Furuya M.; Inomata N.; Osuye K.; Nakazato M. The role of the vagal nerve in peripheral PYY3–36-induced feeding reduction in rats. Endocrinology 2005, 146, 2369–2375. [DOI] [PubMed] [Google Scholar]
- Batterham R. L.; Cohen M. A.; Ellis S. M.; Le Roux C. W.; Withers D. J.; Frost G. S.; Ghatei M. A.; Bloom S. R. Inhibition of food intake in obese subjects by peptide YY3–36. N. Engl. J. Med. 2003, 349, 941–948. [DOI] [PubMed] [Google Scholar]
- Neary N. M.; Small C. J.; Druce M. R.; Park A. J.; Ellis S. M.; Semjonous N. M.; Dakin C. L.; Filipsson K.; Wang F.; Kent A. S.; Frost G. S.; Ghatei M. A.; Bloom S. R. Peptide YY3–36 and glucagon-like peptide-17–36 inhibit food intake additively. Endocrinology 2005, 146, 5120–5127. [DOI] [PubMed] [Google Scholar]
- Moran T. H.; Smedh U.; Kinzig K. P.; Scott K. A.; Knipp S.; Ladenheim E. E. Peptide YY(3–36) inhibits gastric emptying and produces acute reductions in food intake in rhesus monkeys. Am. J. Physiol.: Regul., Integr. Comp. Physiol. 2005, 288, R384–R388. [DOI] [PubMed] [Google Scholar]
- Halatchev I. G.; Ellacott K. L.; Fan W.; Cone R. D. Peptide YY3–36 inhibits food intake in mice through a melanocortin-4 receptor-independent mechanism. Endocrinology 2004, 145, 2585–2590. [DOI] [PubMed] [Google Scholar]
- Zac-Varghese S.; De S. A.; Bloom S. R. Translational studies on PYY as a novel target in obesity. Curr. Opin. Pharmacol. 2011, 11, 582–585. [DOI] [PubMed] [Google Scholar]
- Ahn J. S.; Nazarbaghi R.; D’Souza L. J.; Ghosh S.; Jodka C. M.; Lwin A. N.; Levy O. E. Synthesis & biological evaluation of PYY(3–36) analogs substituted with alanine. Adv. Exp. Med. Biol. 2009, 611, 515–516. [DOI] [PubMed] [Google Scholar]
- Beck-Sickinger A. G.; Wieland H. A.; Wittneben H.; Willim K. D.; Rudolf K.; Jung G. Complete l-alanine scan of neuropeptide Y reveals ligands binding to Y1 and Y2 receptors with distinguished conformations. Eur. J. Biochem. 1994, 225, 947–958. [DOI] [PubMed] [Google Scholar]
- Wu B.; Chien E. Y. T.; Mol C. D.; Fenalti G.; Liu W.; Katritch V.; Abagyan R.; Brooun A.; Wells P.; Bi F. C.; Hamel D. J.; Kuhn P.; Handel T. M.; Cherezov V.; Stevens R. C. Structures of the CXCR4 chemokine GPCR with small-molecule and cyclic peptide antagonists. Science 2010, 330, 1066–1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck-Sickinger A. G.; Jung G. Structure–activity relationships of neuropeptide Y analogues with respect to Y1 and Y2 receptors. Biopolymers 1995, 37, 123–142. [DOI] [PubMed] [Google Scholar]
- Balasubramaniam A.; Tao Z.; Zhai W.; Stein M.; Sheriff S.; Chance W. T.; Fischer J. E.; Eden P. E.; Taylor J. E.; Liu C. D.; McFadden D. W.; Voisin T.; Roze C.; Laburthe M. Structure–activity studies including a ψ(CH2-NH) scan of peptide YY (PYY) active site, PYY(22–36), for interaction with rat intestinal PYY receptors: development of analogues with potent in vivo activity in the intestine. J. Med. Chem. 2000, 43, 3420–3427. [DOI] [PubMed] [Google Scholar]
- Danho W.; Ehrlich G.; Fry D. C.; Khan W.; Swistok J.. Peptides with Neuropeptide-2 Receptor (Y2R) Agonist Activity. Patent WO06/077035.
- Powers E. T.; Deechongkit S.; Kelly J. W. Backbone-backbone H-bonds make context-dependent contributions to protein folding kinetics and thermodynamics: lessons from amide-to-ester mutations. Adv. Protein Chem. 2005, 72, 39–78. [DOI] [PubMed] [Google Scholar]
- Dawson P. E.; Muir T. W.; Clark-Lewis I.; Kent S. B. Synthesis of proteins by native chemical ligation. Science 1994, 266, 776–779. [DOI] [PubMed] [Google Scholar]
- Blankenship J. W.; Balambika R.; Dawson P. E. Probing backbone hydrogen bonds in the hydrophobic core of GCN4. Biochemistry 2002, 41, 15676–15684. [DOI] [PubMed] [Google Scholar]
- Kuisle O.; Quiñoá E.; Riguera R. A General methodology for automated solid-phase synthesis of depsides and depsipeptides. Preparation of a valinomycin analogue. J. Org. Chem. 1999, 64, 8063–8075. [DOI] [PubMed] [Google Scholar]
- Kuisle O.; Quiñoá E.; Riguera R. Solid phase synthesis of depsides and depsipeptides. Tetrahedron Lett. 1999, 40, 1203–1206. [DOI] [PubMed] [Google Scholar]
- Bannwarth W. Gene Technology: a challenge for a chemist. Chimia 1987, 41, 302–317. [Google Scholar]
- Albertsen L.; Østergaard S.; Paulsson J. F.; Norrild J. C.; Strømgaard K. A parallel semisynthetic approach for structure–activity relationship studies of peptide YY. ChemMedChem 2013, 8, 1505–13. [DOI] [PubMed] [Google Scholar]
- Wan Q.; Danishefsky S. J. Free-radical-based, specific desulfurization of cysteine: a powerful advance in the synthesis of polypeptides and glycopolypeptides. Angew. Chem., Int. Ed. 2007, 46, 9248–9252. [DOI] [PubMed] [Google Scholar]
- Nygaard R.; Nielbo S.; Schwartz T. W.; Poulsen F. M. The PP-fold solution structure of human polypeptide YY and human PYY3–36 as determined by NMR. Biochemistry 2006, 45, 8350–8357. [DOI] [PubMed] [Google Scholar]
- Lerch M.; Mayrhofer M.; Zerbe O. Structural similarities of micelle-bound peptide YY (PYY) and neuropeptide Y (NPY) are related to their affinity profiles at the Y receptors. J. Mol. Biol. 2004, 339, 1153–1168. [DOI] [PubMed] [Google Scholar]
- Zerbe O.; Neumoin A.; Mares J.; Walser R.; Walser R.; Zou C. Recognition of neurohormones of the NPY family by their receptors. J. Recept. Signal Transduction Res. 2006, 26, 487–504. [DOI] [PubMed] [Google Scholar]
- Chen Y. H.; Yang J. T.; Chau K. H. Determination of the helix and β form of proteins in aqueous solution by circular dichroism. Biochemistry 1974, 13, 3350–3359. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.