

Nettle et al. [1] explored the conditions under which it could be adaptive for humans to calibrate the development of life history traits (for example, reproductive timing) on the level of adversity experienced in early life. They concluded that external predictive adaptive responses (PARs)—in which early cues are employed to forecast the adult environment—can only evolve if environmental states show ‘almost perfect’ levels of year-to-year autocorrelation, in the order of 0.95 or greater. They suggested that annual autocorrelations could be empirically estimated from environmental time-series datasets; consistent findings of autocorrelations smaller than 0.95 would argue against accounts of human life-history plasticity that involve external PARs.
The model by Nettle et al. is a welcome contribution to the human literature on life history plasticity and the authors should be commended for their emphasis on testability and quantification. Indeed, their model offers conceptual clarity and quantitative predictions in an area where vague verbal claims are the unfortunate norm. However, it is important to understand to what extent the present results may depend on specific assumptions in the model, and critically evaluate the plausibility of the latter. This is especially true for quantitative predictions (for example, the 0.95 threshold) that lend themselves to be employed as heuristics for the interpretation of empirical findings.
Here, I argue that the model by Nettle et al. makes some unrealistic assumptions about the structure of environmental change and present a simple revised model in which past events can have both immediate and delayed effects on subsequent environmental states. I show that when delayed effects are incorporated in the model, the predictive value of early cues shows a notable increase and external PARs can evolve with annual autocorrelations in the order of 0.85 (or even 0.80) instead of 0.95. While the qualitative results obtained by Nettle et al. remain valid, the scope for the evolution of external PARs is larger than that implied in their paper. Future theoretical work should strive to clarify the temporal structure of human environments and systematically explore its implications for the evolution of life-history plasticity.
Nettle et al. modelled environmental fluctuations with a first-order autoregressive process of the form
| 1 |
with ϕ1 > 0. Coefficient ϕ1 in equation 1 is equivalent to parameter r in Nettle et al [1]. In this model, the state of the environment in year t is partly determined by the state of the environment in the previous year, t − 1 (immediate effects). However, earlier environmental states (years t − 2, t − 3,…) cannot affect the present state if not indirectly—that is, through their long shadow on the state of the environment at t − 1. In other words, the ‘memory’ of the environment only extends to the previous year, and there is no way for more distant events to exert delayed effects on the present. This assumption is unrealistic if one considers the structure of human social and physical environments. For example, infectious diseases can have incubation periods longer than one year, can be transmitted from mother to fetus and can have delayed effects on mortality (e.g. systemic infections in childhood may increase the risk of heart failure in adulthood). As humans are able to store and preserve food for long periods of time, a single year of abundance can have delayed positive effects on food availability over multiple years. Also, both intergroup and intragroup conflicts have long-term dynamics that include revenge, truces, alliances and so forth; because of these social dynamics, past conditions can have a wide range of delayed effects on present levels of violence and aggression. Even exogenous factors, for example temperature, usually exhibit small to moderate amounts of positive autocorrelation from one year to the next [2,3], violating the assumption that stochastic fluctuations (Xt in equation 1) are independently distributed across years.
The restrictive assumption that only immediate effects contribute to environmental stability can be relaxed by introducing higher-order terms in equation 1 [4,5]. The minimal extension of Nettle et al.'s model is a second-order autoregressive equation, in which the state of the environment at t is jointly influenced by previous states at t − 1 and t − 2
 |
2 |
with ϕ1,ϕ2 ≥ 0. As in equation 1, the distribution of environmental states generated by equation 2 has mean 0 and variance 1 (see the electronic supplementary material). The relative strength of immediate (t − 1) versus delayed (t − 2) effects in the revised model is determined by coefficients ϕ1 and ϕ2. When ϕ2 = 0, equation 2 reduces to equation 1 and the model describes a first-order autoregressive process (no delayed effects). When ϕ1 = ϕ2, delayed effects have the same weight as immediate effects (‘strong’ delayed effects). A reasonable intermediate case is that of ϕ1 = 2ϕ2, where immediate effects are twice as large as delayed effects (‘weak’ delayed effects). As discussed in the electronic supplementary material, the environmental fluctuations generated by equation 2 have a spectral distribution consistent with those observed in natural animal populations [6,7]. Furthermore, adding a second-order term captures the dynamics of correlated exogenous factors (i.e. non-independent Xt) when those factors can themselves be described by a first-order model [8,9]. Intriguingly, the temporal structure of exogenous climatic factors, such as temperature and precipitation, is often more consistent with a first-order autoregressive process than with a power-law function, in contrast with standard assumptions ([2]; see also the electronic supplementary material); this suggests that equation 2 provides a plausible model of stability and change in natural environments.
In second-order autoregressive models [4,5], the annual autocorrelation (ρ1) is given by
| 3 |
Thus, by choosing appropriate values of ϕ1 and ϕ2, it is possible to obtain the same annual autocorrelation from equations 1 and 2 (see the electronic supplementary material). For example, an annual autocorrelation of 0.80 may result from ϕ1 = 0.80 and ϕ2 = 0 (original model without delayed effects),  and
and 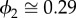 (revised model with weak delayed effects), or
(revised model with weak delayed effects), or  (revised model with strong delayed effects). However, even when the annual autocorrelation is the same, the autocorrelation function of equation 2 decays less steeply across successive years than that of equation 1 [5]. In other words, when the correlation between mt and mt−1 is the same in the two models, the revised model exhibits larger correlations between mt and mt−2, between mt and mt−3, and so forth (see the electronic supplementary material). As a consequence, early cues provide more information about future environmental states and their predictive value increases accordingly.
(revised model with strong delayed effects). However, even when the annual autocorrelation is the same, the autocorrelation function of equation 2 decays less steeply across successive years than that of equation 1 [5]. In other words, when the correlation between mt and mt−1 is the same in the two models, the revised model exhibits larger correlations between mt and mt−2, between mt and mt−3, and so forth (see the electronic supplementary material). As a consequence, early cues provide more information about future environmental states and their predictive value increases accordingly.
Figure 1 compares the results of Nettle et al.'s [1] original model with those of the revised model, for both weak (ϕ1 = 2ϕ2) and strong (ϕ1 = ϕ2) delayed effects. As shown in figure 1a,b, the predictive value of early cues shows a distinct increase in the revised model. For annual autocorrelations between 0.50 and 0.95, the average predictive value increases about 2.5-fold when delayed effects are weak (1-year sampling: 2.50; 5-year sampling: 2.40), and about 3.5-fold when delayed effects are strong (1-year sampling: 3.71; 5-year sampling: 3.52). Improved predictive values are reflected in the results of evolvability simulations: as shown in figure 1c,d, the region of parameter space in which PARs outperform genetically fixed strategies expands when delayed effects are included in the model, especially when individuals average cues over the first five years of life (figure 1d). With strong delayed effects and 5-year sampling, annual autocorrelations in the order of 0.85 can favour the evolution of external PARs. Whereas the parameter space of third- and higher-order autoregressive models is more difficult to examine systematically [4,5], exploratory simulations with third-order effects showed additional increases in predictive value compared with the equivalent second-order models, from about 1.5-fold with weak delayed effects to about twofold with strong delayed effects. As a result, external PARs could meet evolvability criteria with annual autocorrelations in the order of 0.80.
Figure 1.

(a,b) These graphs show the predictive value of early experience for adult environment as a function of the annual autocorrelation. Dotted black line, original model by Nettle et al. [1] (no delayed effects; ϕ2 = 0); dark grey line, revised model with weak delayed effects (ϕ1 = 2ϕ2); light grey line, revised model with strong delayed effects (ϕ1 = ϕ2). For simplicity, simulations in (a,b) assume perfect cue validity [1]. Data represent 10 000 simulated lifetimes for each 0.01 increment of the annual autocorrelation. (c,d) These figures show the regions of the parameter space in which an external PAR outperforms a genetically fixed strategy matched to the long-term environmental mean [1]. Black area, original model by Nettle et al.; dark grey area, revised model with weak delayed effects; light grey area, revised model with strong delayed effects. Data represent 2000 simulated lifetimes for each parameter combination.
In conclusion, the qualitative results of Nettle et al.'s [1] model are robust, but the conditions for PAR evolution derived by these authors should be partially revised. Whereas high year-to-year autocorrelations are still required for the evolution of external PARs, they do not have to be prohibitively high in order for prediction to be adaptive. Simple revisions of the model suggest a threshold of about 0.85 (or even 0.80) instead of 0.95. However, the revised models discussed here are still limited and somewhat unrealistic; more accurate models of temporal variation (for example, incorporating longer delays and long-term environmental cycles) might produce even lower estimates. The general point is that annual autocorrelations only provide a partial picture of the temporal structure of the environment; a more sophisticated description of environmental fluctuations will require a broader range of tools, including autocorrelation functions [4], spectral coefficients [2,3,6,7] and so forth (see the electronic supplementary material). Clarifying the temporal structure of human environments and exploring its implications for the evolution of life history plasticity should become top priorities for future work in this area.
Supplementary Material
Footnotes
The accompanying reply can be viewed at http://dx.doi.org/10.1098/rspb.2013.1343.
References
- 1.Nettle D, Frankenhuis WE, Rickard IJ. 2013. The evolution of predictive adaptive responses in human life history. Proc. R. Soc. B 280, 20131343 ( 10.1098/rspb.2013.1343) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vasseur DA, Yodzis P. 2004. The color of environmental noise. Ecology 85, 1146–1152. ( 10.1890/02-3122) [DOI] [Google Scholar]
- 3.Ruokolainen L, Lindén A, Kaitala V, Fowler MS. 2009. Ecological and evolutionary dynamics under coloured environmental variation. Trends Ecol. Evol. 24, 555–563. ( 10.1016/j.tree.2009.04.009) [DOI] [PubMed] [Google Scholar]
- 4.Chatfield C. 2000. Time-series forecasting. Boca Raton, FL: CRC press. [Google Scholar]
- 5.Wei WWS. 2006. Time series analysis: univariate and multivariate methods, 2nd edn Boston, MA: Pearson Addison-Wesley. [Google Scholar]
- 6.Inchausti P, Halley J. 2001. Investigating long-term ecological variability using the Global Population Dynamics database. Science 293, 655–657. ( 10.1126/science.293.5530.655) [DOI] [PubMed] [Google Scholar]
- 7.Inchausti P, Halley J. 2002. The long-term temporal variability and spectral colour of animal populations. Evol. Ecol. Res. 4, 1033–1048. [Google Scholar]
- 8.Royama T. 1981. Fundamental concepts and methodology for the analysis of animal population dynamics, with particular reference to univoltine species. Ecol. Monogr. 51, 473–493. ( 10.2307/2937325) [DOI] [Google Scholar]
- 9.Williams DW, Liebhold AM. 1995. Detection of delayed density dependence: effects of autocorrelation in an exogenous factor. Ecology 76, 1005–1008. ( 10.2307/1939363) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.