

Abstract
Objective
To elucidate the role of integrin α1β1 in chondrocyte responses to inflammatory interleukin-1α (IL-1) and anabolic transforming growth factor-β1 (TGF-β1) in the knee.
Methods
Intracellular calcium transient responses to IL-1 and TGF-β1 were measured in wild type and integrin α1-null chondrocytes using real time ex vivo confocal microscopy and immunohistochemistry was performed to analyze TGF-β1-mediated activation of Smad2/3 in tibial and femoral chondrocytes.
Results
Loss of integrin α1β1 reduces intracellular calcium transient response to IL-1, while it enhances chondrocyte responses to TGF-β1 as measured by intracellular calcium transients and activation of downstream Smad2/3.
Conclusions
Integrin α1β1 plays a vital role in mediating chondrocyte responses to two contrasting factors that are critical players in the onset and progression of osteoarthritis - inflammatory IL-1 and anabolic TGF-β. Further investigation into the molecular mechanisms by which integrin α1β1 mediates these responses will be an important next step in understanding the influence of increased expression of integrin α1β1 during the early stages of osteoarthritis on disease progression.
Keywords: integrin α1β1, TGF-β, IL-1, chondrocyte, intracellular calcium transient, Smad2/3
Introduction
Chondrocytes maintain articular cartilage by preserving the balance of catabolic and anabolic activity within the extracellular matrix1,2. The biological activity of chondrocytes can be mediated by inflammatory and fibrotic factors, an imbalance of which is implicated in the initiation and progression of osteoarthritis1–5. In general, inflammatory factors such as interleukin-1 (IL-1) promote the degradation of cartilage while fibrotic factors such as transforming growth factor-β1 (TGF-β) promote anabolic activity2,6–11. TGF-β also acts to prevent chondrocyte hypertrophy through activation of the canonical Smad2/3 pathway12,13. In addition to acting alone, IL-1 and TGF-β can also influence each other by modulating TGF-β type II receptor (TβRII) or IL-1 receptor (IL-1R) expression respectively14–17. The interplay between IL-1 and TGF-β is also evident in vivo where TGF-β antagonizes the effects of IL-1 when injected in combination into young (3 months) and old (18 months) mouse knees18.
Integrins are heterodimeric extracellular matrix receptors that can modulate the activation of growth factor receptors such as TβRII, IL-1R and epidermal growth factor receptor (EGFR)10,19–27. Of particular interest to this study is the collagen receptor integrin α1β1 that binds collagen II and chondron localized collagen VI, and is found more abundantly in osteoarthritic compared to healthy cartilage22,28–32. Integrin α1-null mice display no obvious phenotypical abnormalities into adulthood, although they develop spontaneous osteoarthritis earlier in life and more severely than wild type controls29,33. These results, together with the upregulation of integrin α1β1 in the early stages of disease, suggest that this receptor offers protection against osteoarthritis29. However the molecular mechanism(s) through which integrin α1β1 mediates this safeguard, is yet to be elucidated.
The dynamics (concentration, frequency, timing) of intracellular calcium ([Ca2+]i) transients are involved in regulating many cellular processes and an [Ca2+]i transient is often the first measurable biological responses of a cell to external stimuli5,34,35. By using calcium sensitive fluorescent dyes in conjunction with confocal microscopy, the real time [Ca2+]i transient response of live ex vivo murine chondrocytes can be determined36. Chondrocyte [Ca2+]i transients in response to IL-1 have been previously reported, however the effects of TGF-β on [Ca2+]i dynamics are unknown37.
Therefore, the purpose of this study was to compare the histology, morphology and chondrocyte responses to inflammatory IL-1 and anabolic TGF-β in the knees of skeletally mature wild type and integrin α1-null mice. Ex vivo whole femora assays were utilized to measure [Ca2+]i transients and basal activation of downstream canonical Smad2/3. We hypothesized that although the knees of integrin α1-null and wild type mice would be histologically and morphologically similar at the tissue level, integrin α1-null chondrocytes would have heightened responses to IL-1 and suppressed responses to TGF-β, accounting for the earlier development of spontaneous osteoarthritis in the integrin α1-null knee.
Materials and Methods
Animals
All animal procedures were approved by the University of Calgary Animal Care Committee. Breeder pairs of heterozygous integrin α1-null mice were backcrossed onto the BALB/c background strain for ten generations33. Homozygous breeder pairs of integrin α1-null and BALB/c mice were then used to produce the animals used in this study. Genotype was confirmed by polymerase chain reaction of ear punch tissue as previously described33. Equal numbers of male and female skeletally mature BALB/c (wild type) (age = 21 ± 5 weeks, mass = 27 ± 5 g (mean ± sd)) or integrin α1-null mice (age = 21 ± 3 weeks, mass = 30 ± 4 g (mean ± sd)) were used for this study29,33. Anesthetized mice were euthanized via cardiac puncture before their hind limbs were skinned and dislocated at the hip.
Micro Computed Tomography (microCT) Scanning – Bone Morphology
Hindlimbs were fixed at a physiological angle in formalin and high intensity medium resolution (16 μm) microCT scans were performed on each knee (microCT 35, SCANCO Medical, Wayne, PA). Regions of interest were contoured and corresponding bone parameters were evaluated (SCANCO Medical AG, Wayne, PA). Bone volume and density were evaluated for the calcified menisci, and the trabecular and subchondral bone of the femur and tibia [Fig. 1(E and F)]36. Subchondral bone thickness was measured utilizing a circle-drawing algorithm and the mean of five evenly spaced thickness measurements taken across the load bearing region of the knee was reported [Fig. 1(G and H)].
Fig. 1.

Histological images of a 184 day old wild type (A and C) and 182 day old integrin α1-null (B and D) medial condyle, stained with hematoxylin, fast green, and safranin-O (C and D are magnifications of the boxed areas in A and B respectively). Note that proteoglycan staining intensity, cartilage thickness and surface integrity and the size and abundance of chondrocytes are independent of genotype. Frontal (E and F) and sagittal through the medial condyle (G and H) microCT images of the same wild type (E and G) and integrin α1-null (F and H) knee as shown in A and B respectively. Trabecular bone is shaded red, subchondral bone blue, and ossified menisci green in E and F; cortical bone at the femoral condyles and tibial plateau was rendered transparent to show the analyzed trabecular regions. Sagittal sections (G and H) shaded with a thickness map with thicker regions shaded with colors located further right on the spectrum shown below. Trabecular bone architecture, subchondral bone thickness, and the size of ossified menisci and sesamoid bones are similar between the integrin α1-null and wild type knees.
Histology
After microCT scanning, the same hindlimbs were dissected free of muscle and decalcified (CalEX, Fisher Scientific, Ottawa, ON). Sagittal sections 8 μm thick were cut through the entire knee (Leica RM2255) after dehydration and paraffin embedding. Sections were stained with Weigert’s iron hematoxylin (Sigma), 0.2% fast green, and 0.01% safranin-O using an automated processor (Leica ST5010) prior to being coverslipped. One section from the center of each contact region was imaged (Zeiss Axiostart plus, AxioCam ICc1) and histologicaly graded by 3 blinded graders using a modified Mankin grading scheme38.
Media
All media and reconstitution ingredients were purchased from Invitrogen, Burlington, ON. Control media (pH 7.41) contained the following (final concentrations): Dulbecco’s Modified Eagle Medium High Glucose (24 mM D-glucose, no phenol red), L-Glutamine (4 mM), HEPES (15 mM), and Sodium Pyruvate (1 mM). Media osmolarity was measured using a freezing point osmometer (Model 3250, Advanced Instruments Inc., Norwood, MA) and adjusted to 300 mOsm by adding sucrose or distilled water.
Interleukin 1-α (IL-1) and transforming growth factor beta-1 (TGF-β) (R&D Systems, Burlington, ON) were reconstituted (10 μg/mL) in phosphate buffered saline (PBS) or a mixture of 0.1% (w/v) bovine albumin fraction V and 4 mM hydrochloric acid (HCl(aq)) respectively. These isoforms of IL-1 and TGF-β were chosen because; i) they are known to be in the synovial fluid of osteoarthritic patients, ii) IL-1α is more biologically active in the catabolism of meniscus and cartilage compared to IL-1β, and iii) TGF-β1 protects against IL-1 induced proteoglycan degredation3,4,39. Reconstituted treatment or vehicle was diluted in media to reach final treatment or vehicle control concentrations.
Confocal Microscopy
Intact femora were isolated using microdissection. Chondrocytes were incubated (37±1°C) with the calcium sensitive fluorescent dyes: Fura Red AM (60 μM) and Fluo-4 AM (15 μM) (Invitrogen, Grand Island, NY, USA) for 40 minutes and washed for 20 minutes immediately prior to imaging. The dyes were reconstituted in pluronic (20% v/v in DMSO, Invitrogen, Burlington, ON) to aid loading and decrease dye compartmentalization; Sulfinpyrazone (190 μM, MP Biomedicals, Lachine, QC) was added to increase cellular dye retention. Krazy Glue® was applied to the anterior side of the femoral shaft to fix the intact femora to a glass coverslip with condyles facing up. After the Krazy Glue® had cured, the coverslip with femur was placed inside a heated (37±1°C) perfusion chamber (Carl Zeiss Canada Ltd., Toronto, ON) and submersed in control media. Cell viability assays ensured >90% cell viability following the above described microdissection and in situ imaging procedure.
200 confocal images (512×512 pixels, Olympus Fluoview FV1000, Tokyo, Japan) of either lateral or medial condyle were collected at 3.6 second intervals using a 40x/0.8N.A. dipping objective lens (Olympus, Tokyo, Japan) [Fig. 2(A)]. A single excitation wavelength of 488 nm, and collection bands at 504–599 nm and 630–730 nm were utilized. After the first 9 scans, control media was withdrawn from the chamber and replaced by media (37±1°C) containing IL-1, TGF-β, or the appropriate vehicle control [Fig. 2(B–F)].
Fig. 2.
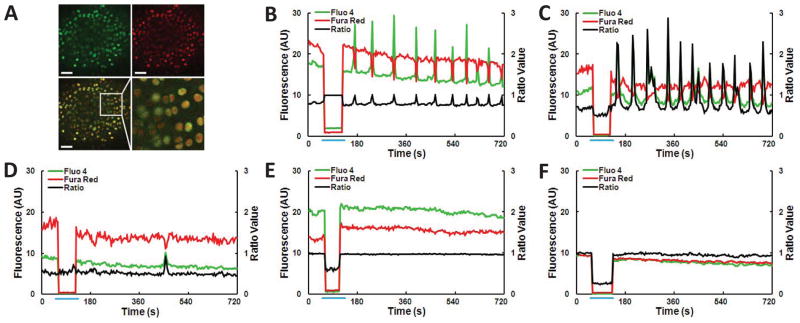
(A) Typical confocal image of an ex vivo wild type murine femora after incubation with calcium sensitive dyes; Fluo-4 (green), Fura Red (red), overlay of green and red fluorescence, magnified section of the boxed area of the overlay, depicting image resolution obtained during [Ca2+]i experiments. Scale bars are 50 μm. (B–F) Characteristic wild type chondrocyte [Ca2+]i ratio data of individual cells from the following treatments: (B, C) Multiple [Ca2+]i transients after (B) 1 ng/mL IL-1 treatment or (C) 5 ng/mL TGF-β; (D) Single [Ca2+]i transient after 5 ng/mL TGF-β; absence of [Ca2+]i transients from (E) 1 ng/mL IL-1 vehicle control or (F) 5 ng/mL TGF-β vehicle control. Fluorescence measured in arbitrary units (AU). Blue line indicates time of withdrawal and infusion of media from the chamber resulting in changes to fluorescence due to shifting focal plane.
Each cell was defined as a region of interest (ROI) and the average fluorescence of the two channels in each ROI was calculated using a custom MATLAB® code. A significant [Ca2+]i transient was defined as an increase in the Fluo-4/Fura Red ratio greater than 3.5 standard deviations above the mean fluctuation measured over the first nine control images of each ROI. Cells signaling during these first nine control images before withdrawal were excluded from analysis and reported results. The percentage of cells responding with single [Fig. 2(D)] or multiple [Fig. 2(B and C)] [Ca2+]i transients was then determined.
Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
Cartilage pieces from 8 wild type or integrin α1-null mice were harvested under sterile microdissection, suspended in TRIzol® (Life Technologies, Burlington, ON) and homogenized (Tissuemiser, Fisher Scientific, Ottawa, ON). RNA was extracted (RNeasy® Mini kit, Qiagen, Toronto, ON), quantified (Nanodrop 2000, Thermo Scientific, Burlington, ON) and reverse transcribed into cDNA using oligo dNTPs and random primers (Omniscript® RT kit, Qiagen, Toronto, ON). Primers for IL-1R1 (forward 5′-CGGCGCATGTGCAGTTAATA-3′, reverse 5′-TGTAGCCGTGAGGATGATAAAGC-3′), IL-1R2 (forward 5′-TGCAAAGTGTTTCTGGGAACC-3′, reverse 5′-ATATTGCCCCCACAACCAAG-3′), TβRI (forward 5′-CTGCCATAACCGCACTGTCAC-3′, reverse 5′-AAATGAAAGGGCGATCTAGTGATG-3′), TβRII (forward 5′-GCAGAACACCTCAGAGCAGTTT GC-3′, reverse 5′-ATCCACAGACAGAGTAGGGTCCAG-3′) and GAPDH (forward 5′-ACCACAGTCCATGCCATCACC-3′, reverse 5′-TCCACCACCCTGTTGCTGTA-3′) were used to measure the expression of these genes in triplicate using qRT-PCR (CFX96 Real-Time System, Bio-rad, Mississauga, ON). mRNA expression of each receptor evaluated by C(t) and normalized to the housekeeper, GAPDH.
Immunohistochemistry
Isolated femora and tibiae were fixed in 4% paraformaldehyde, decalcified (CalEX, Fisher Scientific, Ottawa, ON), then submerged in 25% sucrose. Samples were embedded in optimal cutting temperature compound and flash frozen. Sagittal sections (8 μm) were cut, washed (PBS) and blocked (normal goat serum and Triton X (Sigma, Oakville, ON)) before sequential wash (PBS and Tween 20 (Sigma, Oakville, ON)) and antibody application. A primary antibody against phosphorylated Smad2 (pSmad2) (Ser465/Ser467 which also cross reacts with phosphorylated Smad3, 1:100, Cell Signaling Technology, Danvers, MA) was applied followed by the secondary Alexa Fluor® 647 (Invitrogen, Burlington, ON) to visualize pSmad2/3. The nucleic acid stain Hoescht 33342 (0.1 mM, Invitrogen, Burlington, ON) was applied to all sections. Sections were then mounted in ProLong® Gold (Invitrogen, Burlington, ON) and coverslipped.
Slides were imaged using an oil immersion lens (40x/1.4 N.A.) on an LSM 7 DUO (Carl Zeiss Ltd., Toronto, ON) confocal microscope in either differential interference contrast (DIC) or channel scanning confocal modes. The Hoescht 33342 (excitation 405 nm, emission 429–684 nm) and Alexa Fluor® 647 (excitation 633 nm, emission 638–755 nm) dyes were sequentially scanned to maximize resolution. At least 14 sections from either the femoral condyle (≥244 chondrocytes) or tibial plateau (≥433 chondrocytes) of wild type or integrin α1-null were evaluated for the presence of pSmad2/3 by three experienced blinded graders.
Statistics
Multivariate analysis of variance (ANOVA) with Fisher’s Least Significant Difference (LSD) post-hoc was performed on all parametric morphological bone and histological data using STATISTICA software (StatSoft, Tulsa, OK). The percentage of cells responding with [Ca2+]i transients and the number of chondrocytes positive for pSmad2/3 was statistically analyzed using χ2 test. ANOVA with Fisher’s LSD was performed on the [Ca2+]i transient data using STATISTICA software (StatSoft, Tulsa, OK). A significance level of P<0.05 was used for all statistical analyses.
Results
Loss of integrin α1β1 does not affect knee morphology or histology
Qualitative evaluation of histological sections from the medial and lateral condyles of wild type and integrin α1-null and knees demonstrated similar size, abundance and organization of joint structures, and staining distribution and intensity in all tissues [Fig. 1(A–D)]. In particular, there were no phenotypic differences in cartilage thickness, staining intensity, surface integrity and the size and abundance of chondrocytes. Quantitative histological evaluation confirmed the absence of phenotypic differences (P=0.4607) (Table Ia). Histological signs of very mild OA were seen in one out of three mice of both genotypes and included mild hypertrophy of chondrocytes and undulating cartilage surfaces.
Table I.
Histology score and microCT parameters and for medial and lateral femora or tibiae from knees of wild type and integrin α1-null mice (a), and anterior or posterior regions of medial and lateral calcified menisci (b). Analysed regions of trabecular bone (red shading), subchondral bone (blue shading) and calcified menisci (green shading) are illustrated in [Fig. 1(E, F)].
| (a) | Femur
|
Tibia
|
||||||
|---|---|---|---|---|---|---|---|---|
| Medial | Lateral | Medial | Lateral | |||||
|
|
|
|||||||
| Histology | wild type | α1-null | wild type | α1-null | wild type | α1-null | wild type | α1-null |
| Histology Score (/30) | 6.11 ± 0.51 | 4.11 ± 1.02 | 6.00 ± 2.31 | 6.22 ± 2.22 | 6.22 ± 0.77 | 4.67 ± 0.88 | 6.00 ± 0.00 | 7.44 ± 1.68 |
| Trabecular Bone | ||||||||
| BV (mm3 ×10−1) †‡ | 2.66 ± 0.15 | 2.77 ± 0.20 | 1.70 ± 0.13 | 1.75 ± 0.16 | 1.03 ± 0.39 | 1.03 ± 0.12 | 1.61 ± 0.22 | 1.38 ± 0.27 |
| TV (mm3 ×10−1) †‡ | 4.97 ± 0.59 | 5.08 ± 0.55 | 4.16 ± 0.61 | 4.09 ± 0.66 | 2.32 ± 0.86 | 2.44 ± 0.40 | 5.26 ± 0.49 | 4.14 ± 0.63 |
| BV/TV ×10−2 †‡ | 54.1 ± 8.1 | 54.7 ± 3.0 | 41.0 ± 2.7 | 43.2 ± 4.2 | 44.6 ± 3.9 | 42.3 ± 2.5 | 30.8 ± 5.1 | 33.2 ± 3.5 |
| Conn. Density | 234.4 ± 45.5 | 229.8 ± 20.6 | 181.8 ± 37.7 | 178.1 ± 27.4 | 196.3 ± 11.0 | 244.3 ± 42.0 | 188.9 ± 48.2 | 228.2 ± 81.8 |
| Tb. Thickness (mm ×10−2) †‡ | 8.26 ± 1.58 | 7.81 ± 0.51 | 6.88 ± 0.31 | 7.04 ± 0.49 | 7.28 ± 0.30 | 6.57 ± 0.33 | 6.16 ± 0.55 | 6.07 ± 0.47 |
| Tb. Space (mm ×10−1) †‡ | 1.06 ± 0.08 | 1.01 ± 0.05 | 1.25 ± 0.04 | 1.19 ± 0.09 | 1.28 ± 0.19 | 1.24 ± 0.14 | 1.43 ± 0.23 | 1.29 ± 0.17 |
| Tb. Number | 7.29 ± 0.04 | 7.98 ± 0.20 | 6.94 ± 0.07 | 7.19 ± 0.21 | 8.09 ± 1.30 | 8.11 ± 0.92 | 7.07 ± 0.91 | 8.07 ± 0.84 |
| Bone Den. (mg Ha/ccm) | 925.3 ± 46.9 | 905.7 ± 15.7 | 906.8 ± 19.0 | 911.3 ± 20.4 | 943.3 ± 20.1 | 900.1 ± 13.6 | 889.0 ± 23.5 | 886.4 ± 14.6 |
| App. Den. (mg Ha/ccm) | 551.8 ± 71.9 | 544.4 ± 28.3 | 453.8 ± 22.5 | 471.3 ± 37.3 | 495.3 ± 34.0 | 467.7 ± 23.8 | 375.1 ± 43.5 | 396.2 ± 25.3 |
| Subchondral Bone | ||||||||
| Thickness (mm ×10−1) †‡ | 1.25 ± 0.08 | 1.39 ± 0.08 | 1.65 ± 0.45 | 1.45 ± 0.19 | 0.82 ± 0.08 | 1.00 ± 0.07 | 1.24 ± 0.23 | 1.04 ± 0.07 |
| TV (mm3 ×10−2) | 9.68 ± 0.53 | 9.68 ± 0.59 | 4.94 ± 0.68 | 5.87 ± 0.49 | 8.02 ± 1.83 | 6.78 ± 2.08 | 4.53 ± 1.20 | 6.91 ± 0.79 |
| App. Den. (mg Ha/ccm) † | 814.9 ± 25.7 | 798.4 ± 2.9 | 861.2 ± 20.5 | 875.9 ± 32.2 | 791.7 ± 26.2 | 809.6 ± 10.2 | 799.3 ± 23.2 | 768.9 ± 23.1 |
| (b) | Anterior Meniscus
|
Posterior Meniscus
|
||||||
|---|---|---|---|---|---|---|---|---|
| Medial | Lateral | Medial | Lateral | |||||
| Calcified Menisci | wild type | α1-null | wild type | α1-null | wild type | α1-null | wild type | α1-null |
| TV (mm3 ×10−2) Δ | 8.86 ± 3.41 | 7.13 ± 0.17 | 8.75 ± 0.70 | 9.27 ± 0.83 | 1.08 ± 0.64 | 1.61 ± 0.41 | 1.61 ± 0.32 | 2.19 ± 0.10 |
| App. Den. (mg Ha/ccm) Δ | 713.8 ± 14.4 | 671.6 ± 22.3 | 795.2 ± 6.0 | 820.0 ± 33.0 | 691.0 ± 25.4 | 668.1 ± 18.3 | 648.9 ± 30.5 | 683.1 ± 9.1 |
TV = Total Volume, BV = Bone Volume, BV/TV = Bone Volume/Total Volume, Conn. Density = Connectivity Density, Tb. = Trabecular, App. = Apparent, Den. = Density. Histology Score uses modified Mankin scheme1. N=3 independent measurements for each mean, ± 95% confidence interval. N=3 wildtype mice (age 184 days); N=3 integrin α1-null mice (age 182 days)
indicates femoral values significantly different from tibial values,
indicates medial values significantly different from lateral values,
indicates anterior values significantly different than posterior values.
P<0.05; specific P values in results section.
Qualitative comparisons of the ossified tissues of wild type and integrin α1-null knees including trabecular bone, subchondral bone, and calcified portions of the menisci [Fig. 1(E–H)] confirmed the aforementioned similarities in joint structures and lack of ossification in all ligament and tendon tissues. Quantitative evaluation confirmed the lack of genotypic differences (P=0.9758 (trabecular bone), P=0.8320 (subchondral bone) P=0.9543 (calcified menisci)), although statistically significant site (femur/tibia or anterior/posterior) and condyle (medial/lateral) differences were found (Table I). Bony signs of mild OA were seen in one out of three mice of both genotypes and manifest as small growths of osteophytes around patellar regions.
When both genotypes of mice were grouped, trabecular bone volume (BV), total volume (TV), and BV/TV were greater in the femur than the tibia by 0.096 mm3 (P<0.0001), 0.104 mm3 (P=0.0009), and 0.105 mm3 (P<0.0001) respectively, and on the medial compared to the lateral side by 0.026 mm3 (P=0.0123), 0.071 mm3 (P=0.0128), and 0.119 mm3 (P<0.0001) respectively. Tibial trabeculae were 0.010 mm thinner and 0.018 mm further apart than those on the femur (P=0.0043, P=0.0060), and lateral trabeculae were 0.009 mm thinner and 0.014 mm further apart than those on the medial side (P=0.0054, P=0.0220). The apparent density of the femoral trabecular bone was both 16.5% greater than that of tibial sections (P=0.0005), and 21.4% greater on the medial compared to lateral side (P<0.0001) (Table Ia). As there were no significant differences in corresponding bone density measurements (Table 1a), the smaller apparent density in the tibia compared to the femur was likely attributable to increased porosity rather than decreased density of solid bone in the tibia
The subchondral bone was more than 4 mm thicker on the femur than the tibia (P=0.0002), and 2 mm thicker on the lateral compared to medial side (P=0.0140). The apparent subchondral bone density of the femur was 7.7% greater on the lateral compared to medial side, and was 10.8% greater than the tibia on the lateral side. (P=0.0002, P<0.0001) (Table Ia).
Interestingly, the ossified volume of the anterior menisci was 0.069 mm3 larger than the posterior menisci (P<0.0001) and the apparent density of the anterior portion of the calcified lateral menisci was larger than all other meniscal regions by at least 114.85 mg HA/cm3 (16.6%) (P<0.0001) (Table Ib). Taken together with the observation of higher bone density in the lateral femur, this result indicates a preference for anterio-lateral femoral load bearing during murine gait.
Genotype or concentration do not affect control experiment parameters
Control experiments for IL-1 (PBS) resulted in half as many cells responding when compared to the TGF-β controls (HCl(aq) and BSA) (P<0.0001) [Fig. 3(A and B)]. Furthermore, [Ca2+]i transients following PBS application were shorter and took longer to occur compared to the HCl(aq) and BSA application in the TGF-β controls. Together these observations suggest that the HCl(aq) and BSA solution increases chondrocyte sensitivity to mobilizing [Ca2+]i when compared to exposure to only PBS. Importantly, neither genotype nor concentration affected the percentage of chondrocytes responding with [Ca2+]i transients, nor the analyzed [Ca2+]i transient parameters within either IL-1 or TGF-β control experiments. Any differences observed in [Ca2+]i transient numbers or properties between concentrations or genotypes reported below are thus likely attributable to cytokine exposure.
Fig. 3.
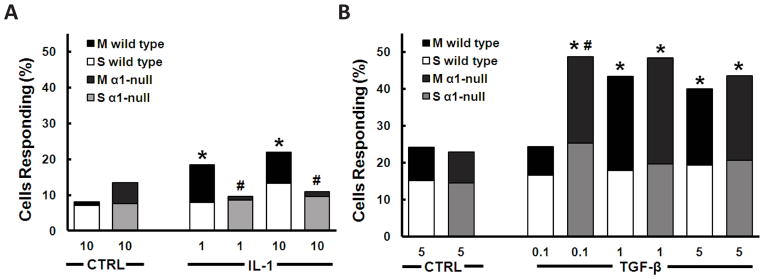
Representation of the percentage of chondrocytes responding with multiple (M) or single (S) [Ca2+]i transients as a function of stimulant IL-1 (A), TGF-β (B), or vehicular control (CTRL)), concentration (ng/mL), and genotype (wild type or integrin α1-null (α1-null)). Note that integrin α1-null chondrocytes are insensitive to IL-1 but hypersensitive to TGF-β, and that the additional responses with treatment are largely multiple [Ca2+]i transients. ‘*’ indicates significantly different from corresponding vehicular control (some data not shown) (wild type 1.0 ng/mL IL-1 P=0.0004, wild type 1.0 ng/mL TGF-β P=0.0003, wild type 5.0 ng/mL TGF-β P=0.0017). ‘#’ indicates significantly different from corresponding wild type stimulus (integrin α1-null 1.0 ng/mL P=0.0090, integrin α1-null 10.0 ng/mL IL-1 P=0.0011) respectively. P<0.0001 unless otherwise indicated. N=3 independent femora, n≥127 cells were used in each treatment.
Integrin α1β1 regulates [Ca2+]i transient responses to IL-1 and TGF-β
The percentage of wild type chondrocytes responding with [Ca2+]i transients more than doubled when exposed to 1.0 or 10.0 ng/mL of IL-1, in a dose independent manner [Fig. 3(A)]. Multiple signals [Fig. 2(B)] were the primary constituent of the response [Fig. 3(A)], likely indicating the involvement of calcium from internal stores34,35. Conversely, when integrin α1-null chondrocytes were exposed to the same concentrations of IL-1, the percentage of cells signaling remained at control levels [Fig. 3(A)] suggesting that integrin α1β1 is necessary for chondrocyte signal transduction of IL-1 measured by [Ca2+]i transients.
In a similar manner to IL-1, the percentage of wild type chondrocytes responding with an [Ca2+]i transient almost doubled after treatment with 1.0 or 5.0 ng/mL of TGF-β, in a dose independent manner [Fig. 3(B)]; the majority of the additional signals were multiple in nature [Fig. 2(C)]. In integrin α1-null chondrocytes however, the threshold of TGF-β required to elicit a response was tenfold lower than wild type [Fig. 3(B)], suggesting a role for integrin α1β1 in regulating the ability of chondrocytes to sense and respond to the presence of very low physiological levels of TGF-β3. All concentrations at or above 0.1 ng/mL TGF-β doubled the percentage of integrin α1-null chondrocytes responding with [Ca2+]i transients [Fig. 3(B)]. Comparable to wild type cells, the majority of responses elicited by TGF-β in integrin α1-null chondrocytes likely involved calcium from intracellular stores as they were multiple in nature34,35. Interestingly, genotype, concentration, or cytokine type did not affect this multiplicity of [Ca2+]i signaling events [Fig. 3(B)].
IL-1 and TGF-β evoke different intracellular calcium transient parameters
All [Ca2+]i transients analyzed were symmetrical, as interpreted by rise time being approximately half of the total duration of the transient (data not shown). On average, treatment with either cytokine, IL-1 or TGF-β, resulted in [Ca2+]i transients that were 3 seconds shorter compared to wild type control chondrocyte transients (P=0.0002, data not shown). The magnitude of wild type chondrocyte [Ca2+]i transients was at least 1.3 fold larger in the presence of IL-1 or TGF-β compared to respective wild type controls [Fig. 4(A)]. The high concentration of TGF-β (5 ng/mL) caused the transient magnitude to increase to over 250% of the control value, a result unique to the TGF-β stimulus [Fig. 4(A)] (P<0.0001).
Fig. 4.

(A) [Ca2+]i transient magnitude of wild type chondrocytes as a function of concentration (low or high concentration that elicited a response) or treatment (vehicle (Control) or Molecule (IL-1 or TGF-β)), measured as the change in Fluo-4/Fura Red fluorescence ratio above baseline in arbitrary units (AU). Only chondrocytes that experienced [Ca2+]i transients were included in the peak analysis from the aforementioned treatments. Note the increase in magnitude in molecule compared to control transients in both TGF-β and IL-1 stimulated cells, and the greater increase of transient magnitude at the higher concentration of TGF-β but not IL-1. (B) [Ca2+]i transient magnitude as a function of genotype (wild type or integrin α1-null (‘α1-null’)) in either TGF-β or corresponding vehicular control treatments. In wild type chondrocytes, transient magnitude was larger in all concentrations when compared to corresponding vehicular controls, in contrast to integrin α1-null chondrocytes, where no increase of magnitude was observed. ‘*’ indicates point is significantly different than all other points on the same graph (P<0.0001), ‘#’ indicates point is significantly different than corresponding vehicular control (Low TGF-β concentration P=0.0118, Low IL-1 concentration P=0.0213, 1.0 ng/mL TGF-β P=0.0082). N=3 independent femora and n≥144 cells for each mean; error bars indicate 95% confidence intervals.
In TGF-β experiments genotype, concentration, or cytokine presence did not influence the duration of [Ca2+]i transients. Wild type chondrocytes displayed a trend of increased [Ca2+]i transient magnitude in TGF-β treatments compared to controls [Fig. 4(B)]; transients induced by 5 ng/mL TGF-β were significantly larger than any other concentration [Fig. 4(B)] (P<0.0001). In contrast, the magnitude of [Ca2+]i transients for controls and all concentrations of TGF-β were similar in integrin α1-null chondrocytes [Fig. 4(B)]. In addition, TGF-β resulted in faster [Ca2+]i mobilization in wild type chondrocytes compared to vehicle controls by 28 seconds (P=0.0094), and this phenomenon was exacerbated with the loss of the integrin α1 subunit (responses 59 second sooner than controls, P<0.0001) (data not shown).
IL-1R2 expression is reduced but IL-1R1 and TβR levels are similar in integrin α1-null chondrocytes at the mRNA level
In light of diminished [Ca2+]i transient responses of integrin α1-null chondrocytes to IL-1 and enhanced responses to TGF-β, we investigated basal chondrocyte expression of the receptors IL-1R1, IL-1R2, TβRI and TβRII associated with these responses. At the mRNA level, expression of IL-1R2 was reduced in integrin α1-null compared to wild type chondrocytes, however there was no difference in expression of IL-1R1, TβRI and TβRII between the two genotypes [Fig. 5(A and B)].
Fig. 5.
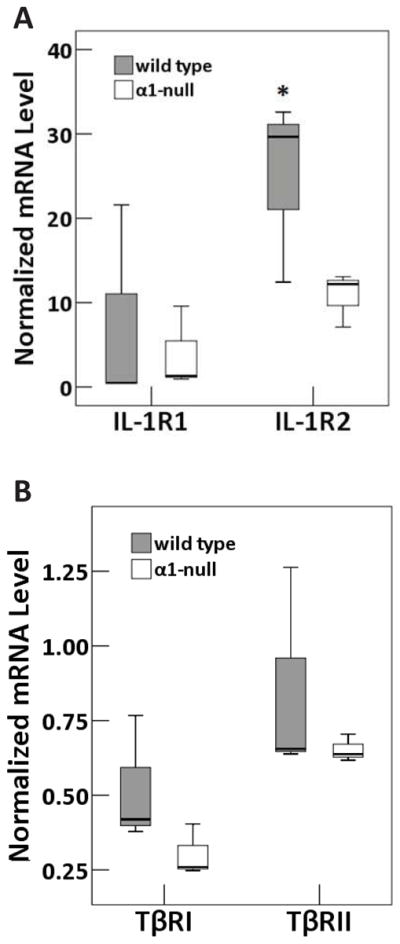
Box and whisker plots representing normalized mRNA levels of (A) IL-1R1 and IL-1R2 and (B) TβRI and TβRII from wild type or integrin α1-null mice. Absolute values of mRNA were calculated for each gene of interest and normalized to GAPDH housekeeping gene. ‘*’ indicates significantly different from corresponding integrin α1-null value (P<0.01). Triplicates of three independent mRNA samples were evaluated in each group. Each independent mRNA sample was extracted from pooled cartilage samples harvested from 8 mice.
Enhanced pSmad2/3 levels in integrin α1-null chondrocytes
Having observed the hypersensitivity of the [Ca2+]i transient response of integrin α1-null chondrocytes to TGF-β treatment, but no difference in basal TβR expression at the mRNA level, we investigated the canonical pathway of TGF-β that requires the phosphorylation of Smad2 and Smad3. In all sections stained by immunohistochemistry, pSmad2/3 staining was punctate and colocalized to both the chondrocytic cytoplasm and nucleus [Fig. 6(A–H)]. Qualitatively more intense stain was found in femoral compared to tibial chondrocytes [Fig. 6(I–P)]. More integrin α1-null chondrocytes stained positively for pSmad2/3 compared to wild type in both femoral (78% vs. 73%) and tibial (78% vs. 68%) cartilage, reaching statistical significance in the tibia (P<0.0001) [Fig. 7]. In the thicker tibial sections, the pSmad2/3 staining expanded to include chondrocytes in the superficial and deep layers of the cartilage in addition to the middle layer that was stained in wild type sections [Fig. 6(M–P)].
Fig. 6.

Basal phosphorylated Smad2/3 (pSmad2/3) in wild type and integrin α1-null chondrocytes. Wild type (A–D) and integrin α1-null (E–H) chondrocytes or sections of femoral condyle (I–L) and tibial plateau (M–P) chondrocytes from wild type (I, K, M, O) and integrin α1-null (J, L, N, P) mice stained with anti-pSmad2/3 antibodies (red) (A, E, I, J, M, N), Hoescht 33342 (blue) (B, F), overlay of both (C, G), DIC channel (D, H) and DIC channel overlaid on fluorescence (K, L, O, P). Scale bar = 5 μm (A – H) or 20 μm (I – P).
Fig. 7.

Graph of percent of basal pSmad2/3 stained chondrocytes in the femoral or tibial cartilage from wild type or integrin α1-null mice. A larger proportion of integrin α1-null chondrocytes (α1-null) stained positively for pSmad2/3 compared to wild type controls, reaching statistical significance in the tibia. ‘*’ indicates significantly (P<0.001) different from corresponding wild type control. Data from a total of at least 244 femoral (n≥244) and 433 tibial chondrocytes (n≥433) from N=3 mice and n≥14 sections were graded in each group.
Discussion
The goal of this paper was to investigate the role of integrin α1β1 in chondrocyte transduction of IL-1 and TGF-β. Concerning IL-1, we have shown that physiological concentrations of IL-1 (1.0 or 10.0 ng/mL) double the number of chondrocytes responding with [Ca2+]i transients ex vivo and that integrin α1β1 is necessary for this response3. Previous studies have shown that focal adhesions are required for IL-1 induced [Ca2+]i signaling and that IL-1 stimulation causes remodeling of the actin cytoskeleton in chondrocytes, suggesting that integrins, which link the cytoskeleton to the extracellular matrix, may play a crucial role in chondrocyte IL-1 signal transduction37,40. Here we identify integrin α1β1 as a critical player in the mediation of chondrocyte responses to IL-1. Furthermore, we show that basal levels of IL-1R2 mRNA expression are diminished in integrin α1-null compared to wild type chondrocytes, though no difference was found in IL-1R1 expression between the genotypes. This suggests that integrin α1β1 can influence chondrocyte responses to IL-1 by changing receptor expression levels, however, further investigation is required to identify the mechanism(s) through which integrin α1β1 exerts this influence.
The multiple nature of the [Ca2+]i transient response to IL-1 we report points to the recruitment of internal [Ca2+]i stores and is in agreement with other studies, however the dose independent nature of the response and the percentage of cells responding in our study contrast previous findings34,35,37. Specifically, the percentage of cells responding to 10 ng/mL IL-1 with [Ca2+]i transient is lower than previously reported (20% vs. ≥91%)37. Differences in species (murine/porcine), IL-1 diffusion (intact femora/explant) and Ca2+ sensitive dyes may have contributed to this inconsistency41.
A key finding of this study is that physiologically relevant concentrations of TGF-β (≥1 ng/mL) double the number of ex vivo chondrocytes responding with an [Ca2+]i transient3. The involvement of [Ca2+]i in enabling TGF-β mediated cell shape change and modulation of protein kinase C dependent intracellular systems has been documented in other cell types42,43. Here we provide evidence of [Ca2+]i involvement in TGF-β signaling in chondrocytes, and that this involves oscillations of [Ca2+]i. The oscillatory nature of TGF-β induced [Ca2+]i likely indicates the involvement of calcium from internal stores34,35 though further investigation is necessary to identify the precise mechanism of [Ca2+]i release. TGF-β is known to influence collagen and glycosaminoglycan production and chondrogenic differentiation in a dose dependent manner44,45. The increase of [Ca2+]i transient magnitude at higher concentrations of TGF-β measured in our study is dose dependent, however the percent of cells signaling was not. Together these results suggest that dose dependent TGF-β responses may arise from prolonged responses of the same cells rather than the recruitment of more cells.
Of particular interest are our findings that integrin α1-null chondrocytes are hypersensitive to TGF-β and also demonstrate increased basal activation of Smad 2/3 despite no differences in basal TβR expression at the mRNA level. Together these results may suggest that integrin α1β1 is able to regulate TβR mediated responses, however further investigation, for example measuring pSmad2/3 expression after TGF-β treatment, may provide further support to this hypothesis. It is known that integrin α1β1 negatively regulates EGFR-mediated signaling by binding and activating the tyrosine phosphatase TCPTP, thus inhibiting the dephosphorylating EGFR25. It is possible that integrin α1β1 could regulate TβR levels and/or activation in a similar manner, however further investigation is required to identify the specifics of this interplay.
When considered within the context of the significant influence of integrin α1β1 on chondrocyte transduction of both IL-1 and TGF-β, it is remarkable that, in agreement with others, we found no significant histological and/or morphological differences between the knees of wild type and integrin α1-null mice at 6 months of age29. It is possible that the genotypic differences in chondrocyte responses to IL-1 and TGF-β may only become evident at the tissue level after a meniscal or ligamentous insult when the differences in signaling characteristics may become vital to the initiation and progression of osteoarthritic degradation29. In a similar vein, one might expect the upregulation of TGF-β- and downregulation of IL-1-responses in integrin α1-null chondrocytes to protect the integrin α1-null mouse from osteoarthritis however this is not the case29,46–48. Integrin α1-null mice develop spontaneous osteoarthritis earlier and more severely than wild type controls29,46. It is possible that the heightened chondrocyte response to TGF-β we report may contribute to the spontaneous osteoarthritis observed in the integrin α1-null mouse, similar to that which occurs after multiple intra-articular injections of TGF-β1 or the over expression of active TGF-β1 in the mouse knee7,49,50. These studies describe ‘chondrophytes’ developing in both the cartilaginous margins of femoro-patellar and femoro-tibial joint spaces, the collateral ligaments and the menisci four days post treatment, a phenotype shared with spontaneous osteoarthritis in the integrin α1-null mouse7,46,49,50. Alternatively, integrin α1β1 may influence other, as of yet unidentified, degradative pathways in the chondrocyte that override the presumed protection afforded by heightened TGF-β and suppressed IL-1 responses measured in the present study.
Despite the remarkable similarity at the tissue level of integrin α1-null and wild type knees, we have shown that α1-null chondrocytes have heightened TGF-β mediated responses and diminished IL-1 responses as measured by [Ca2+]i transients and the relative activation levels of downstream Smad2/3. Together, these results implicate integrin α1β1 in chondrocyte transduction of two important mediators of osteoarthritis and thus the development and progression of this disease. The specific mechanisms by which integrin α1β1 modulates these signaling pathways currently remain unknown.
Acknowledgments
The authors would like to thank Carin Pihl for immunohistochemical tissue processing and animal procedure assistance, Charlie Shin, Christina Jablonski, and Jordan Laudon for RNA harvest and PCR procedure assistance, and Azim Jinha for MATLAB® expertise.
Role of the funding sources
This work was supported by the Canadian Institutes of Health Research Operating Grant Bridge Funding IMH-102815 (AC), the Hunter Family Foundation (AC), University of Calgary Faculty of Kinesiology (AC), the Markin USRP in Health in Wellness (RP, ML, SS), Veterans Affairs Merit Reviews 1I01BX002025-01 (AP) and the National Institutes of Health grants DK095761 (AP).
Footnotes
Author Contributions
All authors made substantial contributions to the design, acquisition of data, and/or analysis and interpretation of data, and were involved in drafting or critically revising the article for important intellectual content. All authors approved the final version to be published. Dr. Clark had full access to all of the data in this study and takes responsibility for both the integrity and accuracy of the data and data analysis.
Study conception and design: Pozzi, Clark.
Acquisition of data: Parekh, Lorenzo, Shin, Clark.
Analysis and interpretation of data: Parekh, Lorenzo, Shin, Pozzi, Clark.
Conflict of interest
None of the authors declare any conflict of interest.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Contributor Information
R Parekh, Email: parekhr@ucalgary.ca.
MK Lorenzo, Email: mklorenz@ucalgary.ca.
SY Shin, Email: ssy080@gmail.com.
A Pozzi, Email: ambra.pozzi@vanderbilt.edu.
AL Clark, Email: alclark@ucalgary.ca.
References
- 1.Goldring MB. The role of the chondrocyte in osteoarthritis. Arthritis Rheum. 2000;43:1916–26. doi: 10.1002/1529-0131(200009)43:9<1916::AID-ANR2>3.0.CO;2-I. [DOI] [PubMed] [Google Scholar]
- 2.Westacott CI, Sharif M. Cytokines in osteoarthritis: Mediators or markers of joint destruction? Semin Arthritis Rheum. 1996;25:254–72. doi: 10.1016/s0049-0172(96)80036-9. [DOI] [PubMed] [Google Scholar]
- 3.Schlaak JF, Pfers I, Meyer Zum Büschenfelde KH, Märker-Hermann E. Different cytokine profiles in the synovial fluid of patients with osteoarthritis, rheumatoid arthritis and seronegative spondylarthropathies. Clin Exp Rheumatol. 1996;14:155–62. [PubMed] [Google Scholar]
- 4.McNulty AL, Rothfusz NE, Leddy HA, Guilak F. Synovial fluid concentrations and relative potency of interleukin-1 alpha and beta in cartilage and meniscus degradation. J Orthop Res. 2013;31:1039–45. doi: 10.1002/jor.22334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mobasheri A, Barrett-Jolley R, Carter SD, Martin-Vasallo P, Schulze-Tanzil G, Shakibaei M. Functional Roles of Mechanosensitive Ion Channels, β1 Integrins and Kinase Cascades in Chondrocyte Mechanotransduction. In: Kamikin A, Kiseleva I, editors. Mechanosensitivity in Cells and Tissues. Moscow: Academia; 2005. [PubMed] [Google Scholar]
- 6.Pettipher ER, Higgs GA, Henderson B. Interleukin 1 induces leukocyte infiltration and cartilage proteoglycan degradation in the synovial joint. Proc Natl Acad Sci U S A. 1986;83:8749– 53. doi: 10.1073/pnas.83.22.8749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Van Beuningen HM, van der Kraan PM, Arntz OJ, van den Berg WB. Transforming growth factor-beta 1 stimulates articular chondrocyte proteoglycan synthesis and induces osteophyte formation in the murine knee joint. Lab Invest. 1994;71:279–90. [PubMed] [Google Scholar]
- 8.Van Beuningen HM, Glansbeek HL, van der Kraan PM, van den Berg WB. Osteoarthritis-like changes in the murine knee joint resulting from intra-articular transforming growth factor-beta injections. Osteoarthr Cartil. 2000;8:25–33. doi: 10.1053/joca.1999.0267. [DOI] [PubMed] [Google Scholar]
- 9.Poleni PE, Etienne S, Velot E, Netter P, Bianchi A. Activation of PPARs alpha, beta/delta, and gamma Impairs TGF-βeta1-Induced Collagens’ Production and Modulates the TIMP-1/MMPs Balance in Three-Dimensional Cultured Chondrocytes. PPAR Res. 2010:635912. doi: 10.1155/2010/635912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Seifarth C, Csaki C, Shakibaei M. Anabolic actions of IGF-I and TGF-beta1 on Interleukin-1beta-treated human articular chondrocytes: evaluation in two and three dimensional cultures. Histol Histopathol. 2009;24:1245–62. doi: 10.14670/HH-24.1245. [DOI] [PubMed] [Google Scholar]
- 11.Homandberg GA, Hui F, Wen C, Purple C, Bewsey K, Koepp H, et al. Fibronectin-fragment-induced cartilage chondrolysis is associated with release of catabolic cytokines. Biochem J. 1997;321 (Pt 3):751–7. doi: 10.1042/bj3210751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Van der Kraan PM, Blaney Davidson EN, Blom A, van den Berg WB. TGF-β signaling in chondrocyte terminal differentiation and osteoarthritis: modulation and integration of signaling pathways through receptor-Smads. Osteoarthr Cartil. 2009;17:1539–45. doi: 10.1016/j.joca.2009.06.008. [DOI] [PubMed] [Google Scholar]
- 13.Yang X, Chen L, Xu X, Li C, Huang C, Deng CX. TGF-βeta/Smad3 signals repress chondrocyte hypertrophic differentiation and are required for maintaining articular cartilage. J Cell Biol. 2001;153:35–46. doi: 10.1083/jcb.153.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Harvey AK, Hrubey PS, Chandrasekhar S. Transforming growth factor-beta inhibition of interleukin-1 activity involves down-regulation of interleukin-1 receptors on chondrocytes. Exp Cell Res. 1991;195:376– 85. doi: 10.1016/0014-4827(91)90387-a. [DOI] [PubMed] [Google Scholar]
- 15.Pronost S, Segond N, Macro M, Rédini F, Penfornis H, Jullienne A, et al. Modulation of interleukin-1 receptor expression by transforming growth factor-beta in cultured rabbit articular chondrocytes: analysis by reverse transcription-polymerase chain reaction. Osteoarthr Cartil. 1995;3:147–55. doi: 10.1016/s1063-4584(05)80049-4. [DOI] [PubMed] [Google Scholar]
- 16.Lim WH, Toothman J, Miller JH, Tallents RH, Brouxhon SM, Olschowka ME, et al. IL-1 beta inhibits TGFbeta in the temporomandibular joint. J Dent Res. 2009;88:557–62. doi: 10.1177/0022034509336823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Baugé C, Legendre F, Leclercq S, Elissalde JM, Pujol JP, Galéra P, et al. Interleukin-1beta impairment of transforming growth factor beta1 signaling by down-regulation of transforming growth factor beta receptor type II and up-regulation of Smad7 in human articular chondrocytes. Arthritis Rheum. 2007;56:3020–32. doi: 10.1002/art.22840. [DOI] [PubMed] [Google Scholar]
- 18.Van Beuningen HM, van der Kraan PM, Arntz OJ, van den Berg WB. Protection from interleukin 1 induced destruction of articular cartilage by transforming growth factor beta: studies in anatomically intact cartilage in vitro and in vivo. Ann Rheum Dis. 1993;52:185–91. doi: 10.1136/ard.52.3.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hynes RO. Integrins: Bidirectional, Allosteric Signaling Machines. Cell. 2002;110:673–87. doi: 10.1016/s0092-8674(02)00971-6. [DOI] [PubMed] [Google Scholar]
- 20.Loeser RF. Integrin-mediated attachment of articular chondrocytes to extracellular matrix proteins. Arthritis Rheum. 1993;36:1103–10. doi: 10.1002/art.1780360811. [DOI] [PubMed] [Google Scholar]
- 21.Giancotti FG. Integrin Signaling. Science. 1999;285:1028–33. doi: 10.1126/science.285.5430.1028. [DOI] [PubMed] [Google Scholar]
- 22.Loeser RF. Chondrocyte integrin expression and function. Biorheology. 2000;37:109–16. [PubMed] [Google Scholar]
- 23.Reynolds LE, Conti FJ, Lucas M, Grose R, Robinson S, Stone M, et al. Accelerated re-epithelialization in β3-integrin-deficient- mice is associated with enhanced TGF-β1 signaling. Nat Med. 2005;11:167–74. doi: 10.1038/nm1165. [DOI] [PubMed] [Google Scholar]
- 24.Wang AZ, Wang JC, Fisher GW, Diamond HS. Interleukin-1β-stimulated invasion of articular cartilage by rheumatoid synovial fibroblasts is inhibited by antibodies to specific integrin receptors and by collagenase inhibitors. Arthritis Rheum. 1997;40:1298–307. doi: 10.1002/1529-0131(199707)40:7<1298::AID-ART15>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 25.Chen X, Abair TD, Ibanez MR, Su Y, Frey MR, Dise RS, et al. Integrin α1β1 controls reactive oxygen species synthesis by negatively regulating epidermal growth factor receptor-mediated Rac activation. Mol Cell Biol. 2007;27:3313–26. doi: 10.1128/MCB.01476-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Arner EC, Tortorella MD. Signal transduction through chondrocyte integrin receptors induces matrix metalloproteinase synthesis and synergizes with interleukin-1. Arthritis Rheum. 1995;38:1304–14. doi: 10.1002/art.1780380919. [DOI] [PubMed] [Google Scholar]
- 27.Shakibaei M, Csaki C, Mobasheri A. Diverse roles of integrin receptors in articular cartilage. Adv Anat Embryol Cell Biol. 2008;197:1–60. doi: 10.1007/978-3-540-78771-6. [DOI] [PubMed] [Google Scholar]
- 28.Heino J. The collagen receptor integrins have distinct ligand recognition and signaling functions. Matrix Biol. 2000;19:319–23. doi: 10.1016/s0945-053x(00)00076-7. [DOI] [PubMed] [Google Scholar]
- 29.Zemmyo M, Meharra EJ, Kühn K, Creighton-Achermann L, Lotz M. Accelerated, aging-dependent development of osteoarthritis in alpha1 integrin-deficient mice. Arthritis Rheum. 2003;48:2873–80. doi: 10.1002/art.11246. [DOI] [PubMed] [Google Scholar]
- 30.Salter DM, Godolphin JL, Gourlay MS. Chondrocyte Heterogeneity: Immunohistologically Defined Variation of Integrin Expression at Different Sites in Human Fetal Knees. J Histochem Cytochem. 1995;43:447–57. doi: 10.1177/43.4.7897185. [DOI] [PubMed] [Google Scholar]
- 31.Ostergaard K, Salter DM, Petersen J, Bendtzen K, Hvolris J, Andersen CB. Expression of alpha and beta subunits of the integrin superfamily in articular cartilage from macroscopically normal and osteoarthritic human femoral heads. Ann Rheum Dis. 1998;57:303–8. doi: 10.1136/ard.57.5.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Loeser RF, Carlson CS, McGee MP. Expression of beta 1 integrins by cultured articular chondrocytes and in osteoarthritic cartilage. Exp Cell Res. 1995;217:248–57. doi: 10.1006/excr.1995.1084. [DOI] [PubMed] [Google Scholar]
- 33.Gardner H, Kreidberg J, Koteliansky V, Jaenisch R. Deletion of integrin alpha 1 by homologous recombination permits normal murine development but gives rise to a specific deficit in cell adhesion. Dev Biol. 1996;175:301–13. doi: 10.1006/dbio.1996.0116. [DOI] [PubMed] [Google Scholar]
- 34.Berridge MJ. Inositol Trisphosphate and Calcium Signaling. Ann N Y Acad Sci. 2006;766:31–43. doi: 10.1111/j.1749-6632.1995.tb26646.x. [DOI] [PubMed] [Google Scholar]
- 35.Berridge MJ. Inositol trisphosphate and calcium signalling mechanisms. Biochim Biophys Acta. 2009;1793:933–40. doi: 10.1016/j.bbamcr.2008.10.005. [DOI] [PubMed] [Google Scholar]
- 36.Clark AL, Votta BJ, Kumar S, Liedtke W, Guilak F. Chondroprotective role of the osmotically-sensitive ion channel TRPV4: Age- and sex-dependent progression of osteoarthritis in Trpv4 deficient mice. Arthritis Rheum. 2010;62:2973–83. doi: 10.1002/art.27624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pritchard S, Guilak F. Effects of interleukin-1 on calcium signaling and the increase of filamentous actin in isolated and in situ articular chondrocytes. Arthritis Rheum. 2006;54:2164–74. doi: 10.1002/art.21941. [DOI] [PubMed] [Google Scholar]
- 38.Furman BD, Strand J, Hembree WC, Ward BD, Guilak F, Olson SA. Joint degeneration following closed intraarticular fracture in the mouse knee: A model of posttraumatic arthritis. J Orthop Res. 2007;25:578–92. doi: 10.1002/jor.20331. [DOI] [PubMed] [Google Scholar]
- 39.Van Beuningen HM, van der Kraan PM, Arntz OJ, van den Berg WB. In vivo protection against interleukin-1-induced articular cartilage damage by transforming growth factor-beta 1: age-related differences. Ann Rheum Dis. 1994;53:593–600. doi: 10.1136/ard.53.9.593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Luo L, Cruz T, McCulloch C. Interleukin 1-induced calcium signalling in chondrocytes requires focal adhesions. Biochem J. 1997;324:653–8. doi: 10.1042/bj3240653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Leddy HA, Guilak F. Site-specific molecular diffusion in articular cartilage measured using fluorescence recovery after photobleaching. Ann Biomed Eng. 2003;31:753–60. doi: 10.1114/1.1581879. [DOI] [PubMed] [Google Scholar]
- 42.Ottaviani E, Sassi D, Kletsas D. PDGF- and TGF-beta-induced changes in cell shape of invertebrate immunocytes: effect of calcium entry blockers. Eur J Cell Biol. 1997;74:336–41. [PubMed] [Google Scholar]
- 43.Sayeed MM. Alterations in calcium signaling and cellular responses in septic injury. New Horizons. 1996;4:72–86. [PubMed] [Google Scholar]
- 44.Worster AA, Nixon AJ, Brower-Toland BD, Williams J. Effect of transforming groth factor β1 on chondrogenic differentiation of cultured equine mesenchymal stem cells. Am J Vet Res. 2000;61:1003–10. doi: 10.2460/ajvr.2000.61.1003. [DOI] [PubMed] [Google Scholar]
- 45.Redini F, Galera P, Mauviel A, Loyau G, Pujol JP. Transforming growth factor β stimulates collagen and glycosaminoglycan biosynthesis in cultured rabbit articular chondrocytes. FEBS Lett. 1988;234:172–5. doi: 10.1016/0014-5793(88)81327-9. [DOI] [PubMed] [Google Scholar]
- 46.Shin SY, Pozzi A, Clark AL. Spontaneous osteoarthritis develops earlier and begins in the subchondral bone of the integrin α1-null mouse knee. 58th Annu Meet Orthop Res Soc. 2012:788. [Google Scholar]
- 47.Van der Kraan PM, van den Berg WB. Anabolic and destructive mediators in osteoarthritis. Curr Opin Clin Nutr Metab Care. 2000;3:205–11. doi: 10.1097/00075197-200005000-00007. [DOI] [PubMed] [Google Scholar]
- 48.Kapoor M, Martel-Pelletie RJ, Lajeunesse D, Pelletier JP, Fahmi H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat Rev Rheumatol. 2011;7:33–42. doi: 10.1038/nrrheum.2010.196. [DOI] [PubMed] [Google Scholar]
- 49.Van Beuningen HM, Glansbeek HL, van der Kraan PM, van den Berg WB. Osteoarthritis-like changes in the murine knee joint resulting from intra-articular transforming growth factor-β injections. Osteoarthr Cartil. 2000;8:25–33. doi: 10.1053/joca.1999.0267. [DOI] [PubMed] [Google Scholar]
- 50.Bakker AC, van de Loo FA, van Beuningen HM, Sime P, van Lent PL, van der Kraan PM, et al. Overexpression of active TGF-beta-1 in the murine knee joint: evidence for synovial-layer-dependent chondro-osteophyte formation. Osteoarthr Cartil. 2001;9:128–36. doi: 10.1053/joca.2000.0368. [DOI] [PubMed] [Google Scholar]