

Abstract
Dopamine plays an important role in the pathogenesis of hypertension by regulating epithelial sodium transport and by interacting with vasoactive hormones/humoral factors, such as aldosterone, angiotensin, catecholamines, endothelin, oxytocin, prolactin pro-opiomelancortin, reactive oxygen species, renin, and vasopressin. Dopamine receptors are classified into D1-like (D1 and D5) and D2-like (D2, D3, and D4) subtypes based on their structure and pharmacology. In recent years, mice deficient in one or more of the five dopamine receptor subtypes have been generated, leading to a better understanding of the physiological role of each of the dopamine receptor subtypes. This review summarizes the results from studies of various dopamine receptor mutant mice on the role of individual dopamine receptor subtypes and their interactions with other G protein-coupled receptors in the regulation of blood pressure.
Keywords: knockout mice, dopamine receptor
Essential hypertension is one of the most common risk factors in developed and developing countries, affecting ∼25% of the middle-aged adult population. Essential hypertension is heterogeneous and is probably caused by an interaction of genetic and environmental factors. The long-term regulation of blood pressure rests on renal and nonrenal mechanisms (53, 84, 87, 104, 161, 189, 234, 268), and abnormalities in the renal regulation of ion transport, intrinsic and extrinsic to the Kidney, have been proposed to cause essential hypertension. Hormones and humoral factors, such as those involved in the renin-angiotensin and sympathetic nervous systems, are preemininent in promoting elevated blood pressure (53, 54, 55, 76, 84, 87, 104, 161, 177, 189, 234, 268). However, hypertension may be caused not only by increased activity of prohypertensive systems but also by defects in antihypertensive systems that serve as counterregulatory mechanisms. Aberrations in these counterregulatory pathways, including the dopaminergic pathway, may be involved in the pathogenesis of essential hypertension (104, 268).
Dopamine receptors expressed in mammals belong to the α-group of the rhodopsin family of G protein-coupled receptors (GPCRs; class A, family A, or family 1) (55, 177, 186, 215). The five mammalian dopamine receptor subtypes, identified by molecular cloning, differ in their primary structures and have distinct affinities for dopamine receptor agonists and antagonists. The D1-like receptors, comprising the D1 and D5 receptor subtypes, couple to the stimulatory G proteins Gs and Golf and activate adenylyl cyclases. The D2-like receptors, comprising the D2, D3, and D4 receptor subtypes, couple to the inhibitory G proteins Gi and Go and inhibit adenylyl cyclases and modulate ion channels (104, 268).
In general, D1-like receptors are expressed postsynaptically/postjunctionally, whereas D2-like receptors can be expressed presynaptically/prejunctionally as well as postsynaptically/postjunctionally (19, 81, 95, 115, 217, 223, 261, 265, 268). Although the dopamine receptor subtypes are expressed in patterns unique to the subtype, they may be coexpressed within individual cells or in distinct cells in the same organ, including those of the Kidney, intestines, and blood Vessels. This situation often precludes the assignment of one individual receptor subtype to a particular system. Moreover, studies on the distinct functional properties of dopamine receptor subtypes expressed in vivo are limited by the lack of agonists and antagonists with selectivity for the individual receptor subtypes. In recent years, a number of laboratories have used gene targeting, via homologous recombination, to generate mice deficient in one or more of the dopamine receptor subtypes (2, 5, 17, 19, 22, 32, 56, 58, 81, 93, 95, 115, 138, 201, 217, 223, 235, 261).
Several recent studies have shown that in addition to specific intramolecular interactions that could define the activation states of GPCRs, intermolecular interactions may also be important (37). Specifically, GPCRs can regulate the function of other receptors by altering expression and/or via direct protein-protein interactions (187).
A number of direct interactions between dopamine receptors and other GPCRs, as well as between dopamine receptor subtypes, have been described in the central nervous system. In neostriatal slices, D1 receptor activation affects the trafficking of N-methyl-d-aspartate (NMDA) receptors between subcellular compartments (59). Conversely, NMDA receptor activation alters D1 receptor subcellular distribution (218). The D1 receptor and subunits NR1 and NR2 of the NMDA receptor physically interact in cell lines and hippocampal neurons, resulting in D1 receptor regulation of NMDA-elicited currents (77, 131, 181). There is a mutually inhibitory modulation between D5 and GABAA receptors that results from a protein-protein interaction between the D5 receptor and the α2-subunit of the GABAA receptor (140). The A1 adenosine receptor heterodimerizes with D1 receptors (80), whereas the closely related A2A adenosine receptor heterodimerizes with D2 receptors (40, 75, 92). These interactions result in cointernalization, cross-desensitization of signaling activities, and functional antagonism between the receptors. D2 receptors interact physically through heterooligomerization with somatostatin receptor 5 and create a novel receptor with enhanced functional activity and pharmacological properties different than those of the individual receptors (200) that may explain the synergistic interactions between somatostatin and dopamine in the central nervous system (106).
D1 and D2 receptors physically interact forming a heteromeric protein complex and coexpress and colocalize within neurons in the human and rat brain. D1 and D2 receptor coactivation generates a novel PLC-mediated calcium signal, indicating that D1 and D2 receptors acquire a new cellular function when coactivated in the same cell (132). Recombinant human dopamine D2 and D3 receptors form functional heterodimers upon coexpression in COS-7 cells, and D2 and D3 receptor coexpression enhances the potency of D2 receptor agonists in transfected cells (149, 214). Because the ultimate effect of dopamine is the sum of the interactions of the different dopamine receptor subtypes and other GPCRs (which may be dependent on the state of sodium balance), impairment of these interactions may result in defective regulation of sodium transport and hypertension.
This review summarizes the role of the different dopamine receptor subtypes in epithelial ion transport, renal physiology, and blood pressure regulation, taking into account clues obtained from mutant mice. Moreover, we also review the role of each dopamine receptor subtype in the pathogenesis of hypertension.
D1 Receptors
Localization of the D1 Receptor Subtype
The Kidney
Earlier autoradiographic, radioligand binding, and functional studies using D1-like receptor agonists and antagonists have shown the presence of D1-like receptors in the renal cortex (proximal tubules) but not in glomeruli or the medulla in the rat Kidney (71, 72, 73, 90, 98, 230). However, glomerular mesangial cells (222) and podocytes (31), in culture, express D1-like receptors. As aforementioned, these studies are limited because the ligands cannot distinguish D1 from D5 receptors.
D1 receptor mRNA is expressed in the rat renal cortex (proximal and distal tubule, arteriole, and juxtaglomerular apparatus) but not in the glomerulus or medulla (169, 248, 250). Immortalized renal proximal tubule cells from the rat (171), mouse (unpublished observations), opossum (160), human (241), and pig (123) also express D1 receptors.
Immunohistochemical and electron microscopic studies have revealed D1 receptors in the cortex and medulla in the rat (9, 170), mouse (5), and human Kidney (178), specifically in apical and basolateral membranes of the proximal and distal convoluted tubule, medullary thick ascending limb of Henle, macula densa, and cortical collecting duct. There are more D1 receptors in the S3 segment than in S1 and S2 segments of the proximal tubule of the rat Kidney (unpublished observations). The expression of D1 receptors in the collecting duct has not been detailed, but dopamine does not stimulate cAMP production in the inner medullary collecting duct (148). The inhibition of vasopressin action by dopamine in this nephron segment has also been attributed to stimulation of α2-adrenergic receptors (62).
D1 receptors are observed throughout the renal vasculature, including juxtaglomerular cells in rats (9,170, 250), but only in large intrarenal arteries in humans (178) (Table 1). In agreement with the latter finding, selective D1-like receptor stimulation increases plasma renin activity in rats but not in humans (188); the effect in mice is not known. D1 receptor immunostaining in renal veins has not been described. Consistent with mRNA studies and adenylyl cyclase activation in response to D1-like receptor agonist (70), D1 receptors are not expressed in glomeruli in either the rat or human Kidney. Immunoblot studies have revealed the presence of several forms of D1 receptors in the rat and human renal proximal tubule from 50 to 210 kDa (260), consistent with data obtained in brain tissue (109).
Table 1. Expression of dopamine receptor subtypes.
| PT | PCT | TAL | mTAL | DT | DCT | CCD | MCD | Glom | JGA | JGC | MD | RV | Species | References | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| D1 receptors | |||||||||||||||
| mRNA Protein | + | + | + | +d | Several | 169, 248, 250 | |||||||||
| + | + | + | + | + | Rat | 9, 170 | |||||||||
| + | + | + | + | + | Mouse | 5 | |||||||||
| + | + | + | + | + | Human | 178 | |||||||||
| +a | Rat | Unpublished data | |||||||||||||
| + | + | Rat | 9, 170, 250 | ||||||||||||
| +e | Human | 178 | |||||||||||||
| D5 receptors | |||||||||||||||
| mRNA Protein | |||||||||||||||
| +a | + | + | + | Rat, mouse | Unpublished data | ||||||||||
| + | + | + | + | Rat, mouse | 7, 9, 252, 254, 255, 280 | ||||||||||
| D3 receptors | |||||||||||||||
| mRNA Protein | TF | TF | TF | TF | TF | TF | TF | TF | + | + | Rat | 79 | |||
| + | Rat | 168 | |||||||||||||
| + | + | + | + | + | + | Rat | 172 | ||||||||
| +b | + | + | + | + | Mouse | 238 | |||||||||
| D4 receptors | |||||||||||||||
| mRNA Protein | + | + | Rat | 227 | |||||||||||
| + | + | + | + | Rat | 195, 227 | ||||||||||
| +c | Rat | Unpublished data | |||||||||||||
| D2 receptors | |||||||||||||||
| mRNA Protein | T | T | T | T | T | T | T | T | Rat | 79 | |||||
| + | Opossum | 159 | |||||||||||||
PT, proximal tubule; PCT, proximal convoluted tubule; TAL, thick ascending limb of loop of Henle; mTAL, medulla TAL; DT, distal tubule; DCT, distal convoluted tubule; CCD, cortical collecting duct; Glom, glomerulus; MCD, medullary collecting duct; JGA, juxtaglomerular apparatus; JGC, juxtaglomerular cell; MD, macula densa; RV, renal vasculature; TF, tubular fraction; T, tubules. Proximal tubular segments:
S3 > S1 = S2,
S1 only, and
S1 > S2 > S3;
arterioles;
large intrarenal arteries only.
Blood Vessels (Other than in the Kidney)
In addition to the inhibition of ion transport, dopamine, via D1-like receptors, also causes vasorelaxation. In the pulmonary artery, D1 receptor immunostaining has been reported in the tunica intima and media of extrapulmonary branches and tunica media of large-sized (but not medium or small sized) intrapulmonary branches. In situ hybridization studies have also shown the expression of the D1 receptor in the rat pulmonary artery as well as the aorta, common carotid, and vertebral arteries. The superior vena cava (119) but not the portal vein expresses D1-like receptors (194). In the rat aorta, common carotid artery, and vertebral artery, D1 receptor mRNA is found only in vascular smooth muscle cells (VSMCs). In contrast, in the pulmonary artery, D1 receptor mRNA is found within the tunica intima, media, and adventitia (110, 119). D1 receptor mRNA is not found in the rat caudal artery (110), and, therefore, the apparent vasoconstrictor effect of D1-like agonists in this blood vessel segment may not be due to a direct vasoconstrictor effect of D1 receptors. Immunohistochemical studies have shown that the D1 receptor is localized on the tunica media of rat pial and mesenteric arteries (8, 192, 270).
Physiology of the D1 receptor
Renal Function
Endogenous renal dopamine is a major physiological regulator of renal ion transport (104, 268). During conditions of moderate sodium balance, >50% of renal sodium excretion is regulated by D1-like receptors (104, 268). Stimulation of the D1-like receptor by exogenous ligands results in an increase in sodium excretion that is in part due to an increase in renal blood flow and a direct inhibition of renal tubular sodium transport (104, 265,268); the glomerular filtration rate is variably affected (73, 164, 171, 231). In contrast, inhibition of renal D1-like receptors decreases sodium excretion without affecting renal blood flow or the glomerular filtration rate in sodium-replete states (42, 73, 114).
As indicated above, the D1-like receptor family is composed of two receptor subtypes: D1 and D5. The D1-like receptor subtype mediating the natriuretic effect of D1-like receptor is not known because of the lack of specific agonists for each D1-like receptor subtype. Because cAMP is involved in the inhibition of renal ion transport by D1-like receptors and the D1 receptor increases cAMP production to a greater extent than the D5 receptor in renal proximal tubule cells (206), it is likely that the natriuretic effect of dopamine is due mainly to the D1 receptor in this nephron segment. This remains to be tested, however. Direct evidence of the involvement of the D1 receptor in the inhibition of renal sodium transport comes from studies with a chronic selective intrarenal cortical infusion of D1 receptor antisense oligodeoxynucleotides, which selectively decrease D1 receptor protein without affecting D5 receptors (239). In this study, sodium excretion during a normal or high salt intake is decreased by the selective renal cortical inhibition of D1 receptor expression (239).
D1-like receptors decrease ion transport in many segments of the nephron (proximal tubule, medullary thick ascending limb of Henle, and cortical collecting duct) by inhibiting the activities of sodium/hydrogen exchanger 3 (NHE3; SLC9A3), sodium-phosphate cotransporter (NaPi-IIa/SLC34A1 and NaPi-IIc/SLC34A3), and exchanger (SLC26A6) at the apical membrane and electrogenic cotransporter (NBCe1A; SLC4A4) and Na+-K+-ATPase at the basolateral membrane (5, 11, 16, 21, 24, 86, 97, 164, 180, 247). In the medullary thick ascending limb of Henle, D1-like receptors decrease sodium transport (78) by inhibiting Na+-K+-ATPase activity, but dopamine actually increases sodium-potassium-chloride cotransporter (NKCC2; SLC12A1) activity in this nephron segment (10). We have suggested that D1-like stimulation of NKCC2 may be important in K+ recycling. However, eicosanoids [20-hydroxyeicosatetraenoic acid (20-HETE)] may synergize with D1-like receptors to inhibit NKCC2 activity. G proteindependent, cAMP/PKA-dependent, and PKC-independent mechanisms are involved in the D1-like receptor inhibition of Na+/Pi2, NHE3, cotransporter, and exchanger activity, including their translocation out of brush-border membranes (20, 23, 24, 68, 125, 180, 245). In contrast, D1-like receptors inhibit Na+-K+-ATPase activity via cAMP/PKA, certain PKC isoforms, and 20-HETE, which internalize its subunits (15, 57, 65, 86, 101, 121, 135, 167, 176, 211). The inhibitory effect of D1-like receptors on Na+-K+-ATPase is nephron segment specific: PKA is involved in D1-like receptor-mediated inhibition in the cortical collecting duct, whereas PKC is involved in the proximal convoluted tubule; eicosanoids are involved in all nephron segments studied (15, 57, 65, 101, 118, 135, 167, 211, 212).
In addition to the direct inhibition of D1-like receptors on sodium excretion, D1-like and D2-like receptors interact to enhance this effect. We have reported that the increase in sodium excretion induced by Z-1046, a dopamine receptor agonist with a rank order potency of D4 > D3 > D2 > D5 > D1, is blocked by either D1-like or D2-like receptor antagonists (127). D2-like receptors may potentiate the inhibitory effect of D1-like receptors on Na+-Pi cotransport, NHE3, and Na+-K+-ATPase activities in renal proximal tubules (see below).
Cardiovascular Function
Circulating concentrations of dopamine are too low to stimulate its own receptors (104, 268). Dopamine administered systemically to increase blood pressure exerts its effect via nondopaminergic receptors; β-adrenergic receptors increase cardiac output, whereas α-adrenergic receptors increase vascular resistance. D1 receptors are expressed in the human and rat heart (152, 178, 277). However, D1-like receptor agonists do not directly affect myocardial contractility (184).
Dopamine, at low concentrations, dilates resistance arteries via D1-like receptors (87,104, 268). The vasorelaxant effect of dopamine in the rabbit pulmonary artery has been reported to be both endothelium dependent and independent (251). However, in the mesenteric artery, the vasodilatory effect of the D1 receptor agonist is endothelium independent (269, 270). The vasorelaxant effect of the D1 receptor is enhanced by calcium channel blockade with nifedipine, indicating that calcium channels are involved in the vasodilatory effect of D1 receptors (269, 270). The vasodilatation induced by D1-like receptors is probably mediated by cAMP; inhibition of Na+-K+-ATPase activity would cause vasoconstriction. In coronary arteries, cAMP cross activation of cGMP-dependent protein kinase stimulates large-conductance calcium-activated potassium channel activity (244).
Interactions with the Renin-Angiotensin system
The D1 receptor may also regulate renal and cardiovascular function by interacting with other systems, including the renin-angiotensin-aldosterone and sympathetic nervous systems. Angiotensin type 1 (AT1) receptors stimulate all renal proximal tubular ion-transporting proteins that are inhibited by D1-like receptors (46, 64, 220). The natriuretic effect of D1-like receptors is enhanced when angiotensin II production is decreased or when AT1 receptors are blocked (46). The renal vasoconstrictor effect of angiotensin II can also be antagonized by D1-like receptor agonists. D1-like and AT1 receptors have opposing effects on the generation of second messengers: D1-like receptors stimulate adenylyl cyclases, whereas AT1 receptors inhibit them. While both D1-like and AT1 receptors stimulate PLC activity, they stimulate different PKC isoforms (18, 63, 88, 256). Dopamine, via the D1-like receptor, also decreases AT1 receptor expression and angiotensin II binding sites in renal proximal tubule cells from normotensive Wistar-Kyoto (WKY) rats (49, 267, 274). However, the D1 receptor can also inhibit AT1 receptor function by direct physical interactions (267), whereas the D5 receptor may be responsible for the D1-like receptor inhibition of AT1 receptor expression (274) (see below). In contrast, angiotensin type 2 (AT2) receptors participate in the natriuresis induced by D1-like receptors (203). There is a negative interaction between D1-like and adrenergic receptors, similar to the negative interaction between AT1 and D1-like receptors in the regulation of renal sodium transport and VSMC contractility. In opossum Kidney cells, the dopaminergic inhibition of Na+/Pi2 is potentiated by treatment with a-adrenergic receptor antagonists (129). We have preliminary data showing that the increase in proliferation of VMSCs produced by stimulation of α1-adrenergic receptors is inhibited by stimulation of D1-like receptors (Li Z, Zeng C, and Jose PA; unpublished observations).
D1 Receptors and Hypertension
Impaired Renal D1 Receptor Function
In rodents with genetic hypertension [spontaneously hypertensive rats (SHRs) and Dahl salt-sensitive rats], D1-like receptor agonist-mediated diuretic and natriuretic responses are impaired (47, 73, 104, 113, 164, 268). The impaired natriuretic response to exogenous D1-like agonists is accompanied by an impaired natriuretic effect of endogenous renal dopamine (47, 73, 127). The impaired natriuretic effect of D1-like receptor agonists has specificity, because the natriuretic effect of cholecystokinin is not impaired in SHRs (127). In hypertensive humans, the ability of D1-like receptor agonists to inhibit proximal tubular reabsorption is impaired (171); however, the overall natriuretic effect of exogenously administered D1 receptor agonists may be greater in hypertensive subjects than in normotensive subjects (163, 171). This is due to the fact that the actions of D1-like receptors on renal hemodynamics and distal renal tubule function (see below) are preserved in hypertension (163, 171).
The decreased ability of D1-like receptor agonists to inhibit ion transport in rodent genetic hypertension is due to diminished D1-like receptor inhibition of NHE3 (5), exchanger (180), cotransporter (125), and Na+-K+-ATPase activities (101, 164). The impaired inhibition of ion transport by D1-like receptors in rodent models of genetic hypertension is due, in part, to impaired production of second messengers (e.g., cAMP, diacylglycerol, eicosanoids) in renal proximal tubules (45, 69, 72, 101, 103, 120, 180) and the thick ascending limb of Henle (164) but not in the cortical collecting duct (174). The impaired ability of D1-like receptors to stimulate cAMP production in renal proximal tubules in genetic hypertension need not be due to decreased total cellular expression of D1 receptor (259) but rather to increased serine phosphorylation and decreased expression of D1 receptors at the plasma membrane (205, 259). The impaired ability of D1-like receptors to stimulate cAMP production in renal proximal tubules is specific because the ability of parathyroid hormone to stimulate cAMP production or stimulate G proteins is intact (103, 120, 205); β-adrenergic function is also intact, at least in young SHRs (153). There is organ specificity because D1 receptor action in the brain striatum is also intact (72). The impaired D1-like receptor function is probably of genetic origin, because it precedes the onset of hypertension and cosegregates with high blood pressure (5, 72, 128, 175).
Impaired Arterial D1 Receptor Function
In general, the renal and nonrenal vasodilatory effects of D1-like receptors in hypertension are not impaired (156, 171). There are, however, reports of an impaired renal vasodilatory effect of D1-like receptor agonist in humans with essential hypertension (39) and in SHRs (43). Indeed, there is an impaired ability of D1-like receptors to stimulate adenylyl cyclase in renal arteries of SHRs (43). We have also reported an impaired ability of D1-like receptor agonists to vasodilate mesenteric arteries of SHRs (269).
D1 Receptor Mutant Mice
D1 receptor-null (D1−/−) mice were generated by targeted mutagenesis. The targeting construct contained 7.0 kb of 129/Sv-derived D1 receptor genomic sequence in the pPNT vector (5, 58). Both homozygous and heterozygous mice had greater systolic, diastolic, and mean arterial pressures than wild-type mice. Renal tubules from homozygous D1 receptor knockout mice have no binding sites for 125I-labeled SCH-23982, a D1-like receptor antagonist, and do not increase cAMP accumulation in response to D1-like receptor agonist stimulation. The response to parathyroid hormone, however, is intact. These data provide reasonable correlation between defective D1 receptor/signal transduction and the development of hypertension in mice (5).
Deficiency of the D1 receptor could be involved in human essential hypertension. The human D1 gene locus at chromosome 5 at q35.1 is linked to human essential hypertension (124). A polymorphism, A-48G, identified at 248 bp of the 5′-untranslated region, is associated with essential hypertension in Japanese subjects (210) but not in Caucasian subjects (30) and has also been reported to be associated with albuminuria (191). There are no reports of the association of polymorphisms of the coding region of the D1 receptor and essential hypertension.
D1 Receptors and Blood Pressure Regulation Summary
D1 receptors regulate blood pressure in the long term by decreasing renal sodium transport and may interact directly and negatively to regulate AT1 receptor function and increase AT2 receptor expression. While D1 receptors can also increase renin secretion, this may only be manifest in rats on a low-sodium diet or during hypovolemia (250); a role of D1 receptors on renin secretion in humans has not been proved. D1 receptors may also exert antihypertensive effects by preventing oxidative stress (see below). It remains to be proved, however, if D1−/− mice are salt sensitive and whether D1 receptor-mediated ion transport is impaired in the renal proximal tubule of D1−/− mice.
D2 Receptors
Localization of the D2 Receptor Subtype
The Kidney
The D2 receptor is expressed as D2short and D2long (154). It has been suggested that the D2short receptor functions as the presynaptic receptor, whereas the D2long receptor functions as the postsynaptic receptor; the D2short receptor is expressed to a greater extent in presynaptic receptors than in postsynaptic receptors (111, 139). D2long mRNA is expressed in renal tubules and glomeruli (79). Most of D2-like receptors in the rat Kidney are prejunctional (25). However, D2 receptor protein has been described in the opossum Kidney cell, a proximal tubule cell line that has some distal tubular cell characteristics (159). The expression of D2 receptors in the intact Kidney is not well documented (Table 1).
Blood Vessels
D2 receptor protein has been described in the heart and coronary artery (44). Dopamine at low and high concentrations constricts isolated porcine pial veins via postsynaptic α2-adrenoceptors and dopamine D1 and D2 receptors (228). However, in the cat and other species, D1 receptors are vasodilatory; the D2 receptor agonist LY-141865 is vasodilatory only at high concentrations (61). The reason for this apparent species difference is not readily apparent. As stated above, D1 receptor-mediated inhibition of Na+-K+-ATPase, per se, should lead to vasoconstriction because of an increase in intracellular sodium and, subsequently, intracellular calcium by the activation of the sodium/calcium exchanger. The D2 receptor could cause vasoconstriction by inhibition of cAMP production but could also cause vasodilation by inhibition of norepinephrine release.
Physiological Role of the D2 Receptor
Renal Function
D2 receptors have variable effects on sodium transport that could not be entirely related to the lack of selectivity of D2 receptor ligands (213). D2 receptors can affect renal function by regulating dopamine transporter activity (130) and renal dopamine production (179). There are studies showing vasodilatory and natriuretic effects of D2-like receptors (104, 179, 268). However, in rat renal proximal tubules, bromocriptine, a D2-like receptor agonist with a 10-fold affinity for D2 over D3 receptors, stimulates Na+-K+-ATPase activity by increasing its α-subunit in the plasma membrane (100, 158). Bromocriptine also increases chloride transport in the medullary thick ascending limb of the rat (85). LY-171555, a D2-like receptor agonist with some selectivity to the D2 receptor, stimulates Na+-K+-ATPase activity in murine fibroblasts heterologously expressing D2Long receptors (249). Interestingly, in sodium-depleted women, dopamine produces an antinatriuretic effect (3). Sulpiride, a D2-like receptor antagonist with an equal affinity for all D2-like receptors, impairs the natriuretic effect of dopamine in volume-expanded women (4). Thus, it is possible that the antinatriuretic effect of the D2 receptor may become manifest during volume depletion, whereas the natriuretic effect becomes manifest during volume-expanded states. Whether this is a direct effect or whether it is due to an interaction with other dopamine receptors (e.g., the D1 receptor) remains to be determined.
As stated above, D1 and D2 receptors can heterodimerize (60) and interact with each other (183, 185). In LTK2 cells transfected with either rat D1 or D2Long cDNA, D1-like receptor stimulation decreases Na+-K+-ATPase activity, whereas D2-like receptor stimulation produces the opposite effect; these effects are transduced by increases or decreases in cAMP production, respectively (85, 96). Bromocriptine and LY-171555 inhibit adenylyl cyclase activities; this has been thought to be the mechanism of the D2 receptor-mediated stimulation of Na+-K+-ATPase activity (100, 249). Unlike the D3 receptor, the D2 receptor can inhibit adenylyl cyclase activity even in the absence of adenylyl cyclase V (199), which is not expressed in the Kidney (33). However, in Chinese hamster ovary (CHO) cells heterologously expressing 10 times more D2 than D1 receptors, stimulation of either receptor results in a potentiation of arachidonic acid release compared with those cells expressing only one receptor (117, 183). Arachidonic acid cytochrome P-450 products have been shown to inhibit renal sodium transport (12, 101, 209). Thus, D1-D2 synergism in the production of cAMP, PLC, and arachidonic acid products might account for the D2 receptor-mediated natriuretic effect. The D1 receptor-mediated activation of PKC could lead to a D2long receptor-mediated sensitization of adenylyl cyclase VI (29). D1 and D2 receptors also synergistically interact to increase c-fos (50) and inhibit Na+-K+-ATPase activity (35). We have speculated that the D2-like receptor stimulates sodium transport under conditions of “low” sodium intake; in contrast, under conditions of “moderate” sodium excess, D2-like receptors, in concert with D1-like receptors, inhibit Na+-K+-ATPase activity in renal proximal tubules cells and synergistically increase sodium excretion (3, 4, 34, 66, 112).
D2 Receptors and Ros
D2 receptors, like D1 and D5 receptors, may also have an antioxidant function. D2 receptor-null (D2−/−) mice, which are hypertensive, have increased urinary excretion of 8-isoprostane, increased NADPH oxidase activity, and increased renal expression of NADPH oxidase subunits Nox1, Nox2, and Nox4 as well as decreased expression of the antioxidant enzyme heme oxygenase-2 in the Kidneys. Apocynin, which impairs NADPH oxidase subunit assembly and activity, or hemin, an inducer of heme oxygenase, normalizes blood pressure in D2−/− mice (14).
D2 Receptors and Hypertension
Abnormalities of D2-like receptor function have been reported in hypertension. One of the polymorphisms of the D2 receptor is associated with hypertension (232, 233). This polymorphism is also associated with decreased D2 receptor expression (165). Transfer of a segment of chromosome 8 containing the D2 receptor gene from the normotensive Brown-Norway rat onto an SHR background decreases blood pressure (122).
D2 Receptor Mutant Mice
The D2 receptor gene was mutated by homologous recombination in embryonic stem cells with a targeting vector to delete all of exon 7 and the 5′-half of exon 8, the region encoding the majority of the putative third intracellular loop, the last two transmembrane domains, and the carboxy terminus. Blastocyst injection was used to generate chimeric mice on a mixed 129/Sv × C57BL/6J background. The original F2 hybrid strain (129/Sv × C57BL/6J) that contained the mutated D2 receptor allele was backcrossed to wild-type C57BL/6J for five generations and genotyped (138). These mice have normal basic motor skills without tremor, ataxia, or abnormal stance or posture but had decreased initiation of movement. We have reported that systolic and diastolic blood pressures are higher in D2 homozygous and heterozygous mutant mice than in control (D2+/+) mice. D2−/− and D2+/+ mice have a similar ability to excrete an acute saline load. α-Adrenergic blockade decreases blood pressure to a greater extent in D2−/− mice than in D2+/+ mice. Epinephrine excretion is greater in D2−/− mice than in D2+/+ mice, and acute adrenalectomy decreases blood pressure to a similar level in D2−/− and D2+/+ mice. An endothelin type B (ETB) receptor blocker for both ETB1 and ETB2 receptors decreases, whereas a selective ETB1 blocker increases, blood pressure in D2−/− mice but not D2+/+ mice. ETB receptor expression is greater in D2−/− mice than in D2+/+ mice. We have concluded that the enhanced vascular reactivity in D2−/− mice may be caused by increased sympathetic and ETB receptor activities (138) and increased ROS production (14). We have also found that D2−/− mice have increased production of aldosterone and that treatment with a mineralocorticoid receptor blocker ameliorates hypertension in these mice (14).
In another strain of D2−/− mice, blood pressure is normal on a normal salt diet but is increased on a high-salt diet with a decrease in renal dopamine production (235). Sympathetic activity is not different between these D2−/− mice and their wild-type littermates. However, renal aromatic amino acid decarboxylase activity and dopamine synthesis are reduced in these D2−/− mice. Basal urine flow and sodium excretion are lower in D2−/− mice than in D2+/+ mice, but dopamine increases urine volume and sodium excretion in D2−/− mice to levels similar to those in D2+/+ mice (179). As with the differences in two different strains of D3−/− mice, the differences between the two strains of D2−/− mice could be related to genetic background.
D2 Receptors and Blood Pressure Regulation Summary
D2 receptors affect renal function by regulating dopamine production, dopamine transporter activity, and sympathetic nerve activity. D2 receptors, by themselves, increase renal sodium transport, but coactivation of D1 receptors produces the opposite effect. D2 receptors regulate blood pressure by influencing sodium transport and by inhibiting ROS production. D2 receptors may also have anti-inflammatory actions (13).
D3 Receptors
Localization of D3 Receptors
The Kidney
Specific radioligand binding of (±)-7-hydroxy-N,N-di-n-propyl-2-aminotetralin (7-OH-DPAT), which has a high affinity for both D2 and D3 receptors, is reported in cortical tubules and mesangial cells of the rat Kidney (26). D3 receptor mRNA is expressed in glomerular, tubular, and vascular fractions of the rat Kidney (79). D3 receptor protein is expressed in the apical/subapical area but not in the basolateral areas of the rat renal proximal tubule (168) (Table 1). The expression of the D3 receptor in other nephron segments has not been consistent. Nurnberger et al. (194) could not detect any D3 receptor protein in the distal tubule, cortical collecting duct, glomeruli, and renal Vessels. This is in contrast to the report of O'Connell et al. (172), in which D3 receptor protein was found not only in the renal proximal tubule but also in the apical membrane of the distal convoluted tubule and cortical collecting duct (intercalated cells); D3 receptor protein was also observed in glomeruli and renal blood Vessels. In mice, D3 receptor protein, detected by immunocytochemistry, is mainly found in apical vesicles, subjacent to the microvillous brush borders of the S1 segment of the proximal convoluted tubule, the macula densa, cytoplasm of the thick ascending limb of loop of Henle, distal convoluted tubule, and glomeruli, but not in the collecting duct (238). All three studies are in agreement in the absence of specific staining in the medulla. O'Connell et al. (172) could not find D3 receptors in juxtaglomerular cells but found them expressed in the macula densa of the rat Kidney. However, D3 receptor mRNA and functional D3 receptors have been described in rat juxtaglomerular cells in primary culture (207). The discrepancy between the findings in the Kidney in situ and juxtaglomerular cells in culture suggests that the expression of D3 receptors is conditional (e.g., culture dependent). Two species of D3 receptor (45 and 90 kDa) are expressed in renal brush-border membranes (127) and proximal tubule cells (264), as detected by immunoblot studies.
Blood Vessels
D2-like receptors are located mainly in the intima and adventitia of blood Vessels. In the rat renal artery, the D3 receptor is located at both prejunctional (adventitia) and post-/extrajunctional sites (tunica media) (27). In contrast, in the rat mesenteric artery, we found that the D3 receptor is expressed in both the tunicae intima and media (270). Rat pulmonary arteries do not express D3 receptors (196), and, therefore, it is possible that the localization of the D3 receptor is blood vessel specific.
Physiology of the D3 Receptor
Renal Function
The D3 receptor agonists 7-OH-DPAT and PD-128907 (134), which have preferential selectivity for D3 receptors over D2 and D4 receptors, decrease renal sodium transport (144). Acute intravenous administration of 7-OH-DPAT in salt-resistant Dahl rats increases the glomerular filtration rate and sodium and water excretion without affecting blood pressure (144). Quinerolane, a D2-like receptor agonist with 21-fold selectivity for the D3 receptor over the D2 receptor, opens K+ channels, resulting in hyperpolarization and inhibition of Na+-K+-ATPase activity (82). PD-128907, a selective D3 receptor agonist with a 120-fold selectivity over the D2 receptor, infused directly into the renal artery, dose dependently increases fractional sodium excretion in WKY rats (but not in SHRs) fed a normal salt diet or high-salt diet (263). That D3 receptors can mediate natriuresis is supported by the decreased ability of D3 receptor-deficient mice to excrete an acute saline load (17). The renal effects of D3 receptor activation are mediated by actions on postjunctional glomerular and tubular receptors and presynaptic modulation of norepinephrine release because renal denervation attenuates the effects of 7-OH-DPAT (155).
Arteries
Systemic administration of D3 receptor agonists decreases systemic blood pressure (144, 146). The postjunctional D3 receptor-mediated vasodilation is more manifest when renal vascular resistance is increased. Our studies in the isolated relaxed mesenteric artery have shown that two different D3 receptor agonists, PD-128907 and 7-OH-DPAT, have no effects on basal vascular contractility (269, 270). However, when mesenteric artery rings are preconstricted by high-potassium or norepinephrine, both PD-128907 and 7-OH-DPAT induce vasorelaxation. In norepinephrine-preconstricted mesenteric artery rings, a calcium channel blocker increases the vasorelaxant effect of PD-128907, indicating that the vasorelaxant effect of the D3 receptor is, in part, caused by a decrease in intracellular calcium. The vasodilatory effect of D3 receptors may also involve small- and/or large-conductance calcium-activated potassium channels (50, 269), a mechanism that also occurs in renal tubules (82). The hyperpolarization caused by a decrease in intracellular potassium probably prevents the vasoconstriction that occurs with inhibition of Na+-K+-ATPase (82). As with the inhibitory effect of the D3 receptor on sodium transport, the vasodilatory effect of the D3 receptor is probably not related to its ability to inhibit adenylyl cyclase activity. Indeed, the D3 receptor may not inhibit renal cAMP production because this action necessitates the presence of adenylyl cyclase V (199), which is not expressed in the Kidney (33). Thus, the D3 receptor induces a natriuresis and vasodilation via mechanisms that are different from but complement the D1 receptor, which induces a natriuresis and vasodilation by activation of Gαs and/or adenylyl cyclase activity.
D3 Receptor-Mediated Antioxidative Effects
The effect of D3 receptors on ROS is controversial. One report (67) has shown that the D3 receptor stimulates PLD activity in human embryonic Kidney (HEK)-293 cells heterologously expressing the human D3 receptor. However, the D3 receptor has been reported to increase a dopamine autotrophic factor that has an antioxidant action, and, thus, the D3 receptor may have an antioxidant effect, albeit indirectly (41). The D3 receptor is expressed in the tunica intima (269, 270), and activation of the D3 receptor inhibits superoxide production in human brain endothelium cells as measured by a cytochrome c reduction assay (Yang Z, Zeng C, and Jose PA; unpublished observations). The hypertension in D3 receptor-null (D3−/−) mice is not associated with increased production of ROS, but this may be due to increased expression of D5 receptors in these mice (unpublished data). As discussed above, the D5 receptor exhibits significant antioxidant activity.
Interactions with the Renin-Rngiotensin System
D3−/− mice have renin-dependent hypertension (17), and renal AT1 receptor expression (266, 276) is higher in D3−/− mice than in their wild-type (D3+/+) littermates, supporting the notion discussed earlier that the dopaminergic and renin-angiotensin systems interact to regulate renal function (46, 64, 99, 220, 266). Activation of the D3 receptor decreases AT1 receptor expression in renal proximal tubule cells from normotensive rats (266). The D3 receptor probably also inhibits renin release; plasma renin levels are elevated in D3−/− mice (17). Because renal proximal tubule cells can generate angiotensin II (48), a D3 receptor-mediated decrease in angiotensin II formation could also be responsible for the decrease in AT1 receptor expression since angiotensin II has been reported to increase AT1 receptor mRNA in rats (48). However, long-term infusion of angiotensin II in vivo in rats has no effect on AT1 receptor expression in renal proximal tubules (89). Angiotensin II does not increase AT1 receptor expression in immortalized renal proximal tubule cells from normotensive rats (264). Therefore, the D3 receptor, independent of angiotensin II, can regulate AT1 receptor expression.
D3 Receptors and Hypertension
D3 receptor deficiency may be important in the development of salt-sensitive hypertension. As discussed above, D3−/− mice are hypertensive (17, 266, 276). Salt-resistant Dahl rats on a high-sodium diet and chronically treated with a highly selective D3 receptor antagonist (BSF-135170) have increased blood pressure (146). D3 Ser9Gly, heterologously expressed in CHO cells, has been reported to impair D3 receptor-mediated inhibition of cAMP production but acquires the ability to decrease PGE2 production (91). However, there is no association of D3 Ser9Gly, or other D3 receptor gene variants, with hypertension (224), although the chromosome locus of the D3 receptor gene (3q13.3) has been linked to human essential hypertension (108, 197).
Impaired Renal D3 Receptor Function
Activation of D3 receptors induces a natriuresis in salt-sensitive Dahl rats on a normal sodium diet but not in hypertensive salt-sensitive Dahl rats on a high-sodium diet. The diminished functional response in the hypertensive salt-sensitive Dahl rats rat is associated with a decreased [3H]7-OH-DPAT binding to renal membrane protein (146). However, the same group (145) did not find a strain-dependent natriuretic effect of 7-OH-DAT in WKY rats and SHRs. The interpretation of this study is confounded by the systemic administration of 7-OH-DPAT; the activation of D3 receptors outside of the Kidney may have obfuscated any differential effect on sodium excretion between WKY rats and SHRs. To overcome any confounding systemic effects of administered ligands, we studied the renal effects of another selective D3 receptor agonist, PD-128907, infused directly into the renal artery in WKY rats and SHRs fed a high-salt diet. PD-128907 dose dependently increases fractional sodium excretion in WKY rats. No such effect is noted in SHRs (263). The mechanisms causing the impaired D3 receptor in the Kidney in hypertension are not completely known. The stimulatory effect of the D3 receptor agonist PD-128907 on D3 receptor expression is no longer evident in renal proximal tubule cells from SHRs (266). It is possible that mechanisms that impair D1 receptor function in SHRs also impair D3 receptor, e.g., increased GPCR-related kinase-4 (GRK4) activity (208) or the impaired synergistic interaction between D1 and D3 receptors occurs because of an impaired D1 receptor function, per se. The impaired natriuretic function of D3 receptors in SHRs may, in part, be related to the aberrant D3 and AT1 receptor interaction in renal proximal tubule cells (266). The D3 receptor decreases AT1 receptor expression in renal proximal tubule cells from WKY rats, whereas D3 receptor stimulation increases AT1 receptor expression in SHRs (266).
Impaired Arterial D3 Receptor Function
In isolated mesenteric arterial rings, D3 agonist-induced vasorelaxation is similar in WKY rats and SHRs except at very high concentrations; this effect is via the D3 receptor, because it can be blocked by the D3 receptor antagonist U99194A. There is an additive vasodilatory effect between D1 and D3 receptors in WKY rats, which is lost in SHRs (269, 270).
D1/D3 Receptor Interactions
D1 and D3 receptors have been shown to colocalize inside and outside the central nervous system (198, 272, 279). In the central nervous system, these receptors are coexpressed in the same neurons, particularly in the nucleus accumbens and caudate-putamen, and elicit opposite effects on target gene expression by regulating ERK activation and c-fos induction (279). D3−/− mice have blunted dorsal striatal and extrastriatal c-fos responses to D1 receptor stimulation, indicating cooperativity between these receptors in the modulation of c-fos responses (116). Moreover, in neocortical neurons, constitutive inactivation of D3 receptors leads to a decrease in agonist-promoted D1 receptor activity (216). In neurons of the island of Calleja, basal c-fos expression is maintained by endogenous dopamine acting tonically on D1 and D3 receptors, subserving opposite effects on the same cell while having a synergistic effect in the nucleus accumbens (198). These indicate that the two receptor subtypes may affect cells in synergy or in opposition depending on the cell type or signal generated.
In denuded preconstricted mesenteric rings, D1 receptor stimulation augments the vasodilatory effects of D3 agonists (270). In the same way, D3 receptor stimulation augments the vasodilatory effects of the D1-like agonist fenoldopam (269), supporting a role of cooperative D1/D3 interaction in the regulation of blood pressure.
Stimulation of D1 receptors increases D3 receptor expression in the medulloblastoma TE671 cell line (133), embryonic thoracic aortic smooth muscle cells (A10) (270), human coronary artery VSMCs, and immortalized renal proximal tubular cells from normotensive WKY rats (269). Conversely, in renal proximal tubular cells from WKY rats, stimulation of D3 receptors increases D1 receptor protein expression (269, 275).
D1 and D3 receptors coimmunoprecipitate in VSMCs (270) and renal proximal tubular cells from WKY rats (269). The coimmunoprecipitation is increased in renal proximal tubular cells from WKY rats after long-term treatment with the D1-like agonist fenoldopam (269) as well as after treatment with the D3 agonist PD-128907. These effects are probably mediated by increased receptor protein expression (272) and indicate a physical interaction between D1 and D3 receptors. Furthermore, in renal proximal tubular cells from WKY rats, treatment with a D3 agonist rapidly and transiently increases the amount of D1 receptors at the cell surface membrane (272). D1 receptors can be recruited to the cell surface membrane from the cytosol within minutes after D1 receptor stimulation (38). The mechanisms for the synergism between D3 and D1 receptors may be time related. The D3 receptor-mediated increase in cell surface membrane expression of D1 receptors occurs in the short term, whereas the D3 receptor-mediated increase in total D1 receptor expression is a long-term effect (272).
The D1/D3 receptor interaction is impaired in SHRs. Preconstricted mesenteric artery rings are less sensitive to the vasorelaxant effects of fenoldopam in SHRs than in WKY rats. Pretreatment with the D3 agonist PD-128907 does not produce any additional vasorelaxant effect in SHR rings as it does in WKY rings (269). There is a lack of responsiveness of SHRs to infusion of the preferential D2-like agonist Z-1046. SHRs, unlike WKY rats, do not react to the infusion of Z-1046 by increasing glomerular filtration and water and sodium excretion (127). In renal proximal tubular cells from SHRs, the D1-like agonist fenoldopam decreases rather than increases the expression of D3 receptor protein, and the D1-like agonist does not increase the coimmunoprecipitation of D3 and D1 receptors (269). Similarly, treatment with a D3 agonist does not increase the expression of D1 receptors in renal proximal tubular cells from SHRs and does not affect the coimmunoprecipitation of the two receptors (272). Furthermore, the basal level of cell surface membrane expression of D1 receptors in renal proximal tubular cells is lower in SHRs than in WKY rats and is decreased further after treatment with a D3 agonist (272). Taken together, these data indicate that the interaction between D1 and D3 receptors is absent or lessened in hypertension, and results in defective inhibition of sodium transport and relaxation of vascular smooth muscles and ultimately in the development and/or maintenance of high blood pressure.
D3 Receptor Mutant Mice
D3−/− mice were generated by targeted mutagenesis. The targeting construct contained 7 kb of 129/sv-derived D3 receptor genomic sequence in the GKNeo cassette in the antisense orientation at the SalI site in exon 2 (2). Homologous recombination resulted in a mutant allele in which sequences downstream of Arg148 in the second intracellular loop of the D3 receptor were replaced by sequences derived from the Neo gene. Despite the similarity of the primary peptide sequences of the three members of the D2-like receptors, the locomotor phenotype of D3 mutants does not resemble that of D2 mutants. D3−/− mice develop normally, are fertile, and, at most, show a transient and rapidly habituating locomotor hyperactivity in a novel environment. Heterozygous and homozygous D3 mutant mice on a mixed C57/BL6 and B129 background (17) as well as those in a congenic C57BL/6 background (238) have higher systolic and diastolic blood pressures than their wild-type (D3+/+) littermates. In a report by Staudacher et al. (225), D3−/− mice, also in a congenic C57BL/6 background, have blood pressures similar to their D3+/+ littermate on a low, normal, or high salt intake. This report has to be interpreted with caution because C57BL/6 mice fed a high-salt diet may or may not have increased blood pressure depending on the source of these mice. Thus, C57BL/6 mice from Jackson Laboratories develop hypertension when fed a high-salt diet (226), whereas those from Taconic (253) do not. Nevertheless, these two strains of D3−/− mice cannot increase sodium excretion after an acute or a chronic salt load (17, 225). In our study (17), renal renin activity and AT1 receptor expression are greater in homozygous mice than in wild-type mice; values for heterozygous mice are intermediate. Blockade of AT1 receptors decreases systolic blood pressure for a longer duration in mutant mice than in wild-type mice. Thus, disruption of the D3 receptor increases renal renin production and produces renal sodium retention and renin-dependent hypertension (17).
D3 Receptors and Blood Pressure Regulation Summary
D3 receptors may regulate blood pressure directly by inhibition of renal sodium transport or indirectly by interaction with D1 receptors or by inhibition of renin secretion and AT1 receptor expression. These effects are impaired in hypertensive states.
D4 Receptors
Localization of the D4 Receptor Subtype
The Kidney
D4 receptor mRNA is expressed in the Kidney (151), especially in the cortical and medullary collecting ducts (227). D4 receptor mRNA has also been reported in rat juxtaglomerular cells in culture (207). D4 receptor protein expression is highest in the cortical and medullary collecting ducts (227) (luminal > basolateral), followed by the proximal tubule (S1 > S2 > S3; unpublished data) and distal convoluted tubule (195). D4 receptors are not expressed in the glomerulus or loop of Henle (Table 1).
Blood Vessels
The D4 receptor is expressed in human aortic and umbilical endothelial cells and modulates von Willebrand factor secretion (262). D4 receptor protein has been reported to be expressed in the adventitia and adventitia-media border of pulmonary (196) as well as pial and mesenteric arteries (8). D4 receptors may be expressed pre- and postjunctionally (8). In the renal artery, D4 receptor protein is observed perivascularly in the adventitia and adventitia-media border, especially in the afferent and efferent arterioles. D4 receptor immunostaining disappears in the blood vessel (but not in the tubule) after renal denervation, indicating that vascular D4 receptors are prejunctional, whereas tubular D4 receptors are postjunctional (195). D4 receptors are expressed in the atria but not ventricles of rats and humans (193). In contrast, the right and left ventricles of the guinea pig express D4 receptors. The D4 receptor agonist PD-168077 exerts a negative chronotropic and inotropic effect associated with a decrease in cAMP production in the isolated guinea pig heart preparation (83). However, D4 receptors normally have minimal effects on the rat cardiovascular system (184).
Physiology of the D4 Receptor
Renal Function
D4 receptors antagonize vasopressin- and aldosterone-dependent water and sodium reabsorption in the cortical collecting duct (137, 202). In the rabbit cortical collecting duct, the D4 receptor-mediated decrease in sodium transport is exerted mainly at the basolateral membrane despite a greater expression of D4 receptors in luminal membranes (202). However, renal cortical and medullary Na+-K+-ATPase activities are similar in D4 receptor-null (D4−/−) and control (D4+/+) mice. D4−/− mice also do not have an impaired ability to excrete an acute saline load. It is possible that high blood pressure in D4−/− mice, which elicits a pressure natriuresis, obfuscates any deficit in the renal handling of sodium in D4−/− mice (32).
D4 Receptors and Ros
D4 receptors may not have inhibitory effects on the production of ROS (28).
D4 Receptors and Hypertension
A locus near the D4 receptor gene (11p15.5) has been linked to hypertension. The most intensively studied D4 receptor polymorphism is a 48-bp repeat located in exon 3 of the D4 receptor gene. This variant codes for a 16-amino acid sequence located in the third intracellular loop of D4 receptor protein, a region that is thought to interact with G proteins and influence intracellular levels of cAMP (236). The number of repeats at the D4 site varies from 2 to 10, but in Caucasian subjects the 4- and 7-repeat lengths are the most common. In this population, the long variant of the D4 gene is associated with a 3.0-mmHg higher systolic blood pressure and 2.0-mmHg higher diastolic blood pressure (219).
D4 receptor protein is increased in the renal cortex of SHRs relative to WKY rats, but D4 protein in the inner medulla is similar in those two rat strains (221). The effects of D4 agonists and antagonists on cardiovascular and renal function in genetically hypertensive rats have not been reported.
D4 Receptor Mutant Mice
A 129SvEv mouse genomic library was screened with a human D4.4 receptor probe. Positive phages were mapped and partially sequenced. The CsCl banded targeting vector was linearized (NotI),electroporated into ∼2 × 107 129/Ola Hsd E14TG2A embryonic stem cells, and maintained under double selection. The original F2 hybrid strain (129/Sv × C57BL/6) carrying a mutant form of the dopamine D4 receptor was backcrossed to C57BL/6J mice. We have reported that in conscious or pentobarbital-anesthetized mice, systolic and diastolic blood pressures are elevated in D4−/− mice compared with D4+/+ littermates. Juxtaglomerular cells in culture also express D4 receptors (207), but D4−/− mice do not have altered circulating or renal renin levels (32). The protein expression of the AT1 receptor is increased in homogenates of the Kidney and brain of D4−/− mice relative to D4+/+ mice, although AT1 receptor expression in the heart is similar in the two strains. Bolus intravenous injection of the AT1 receptor antagonist losartan initially decreases mean arterial pressure to a similar degree in D4−/− and D4+/+ littermates. However, the hypotensive effect of losartan dissipates after 10 min in D4+/+ mice, whereas the effect persists for >45 min in D4−/− mice. The absence of the D4 receptor increases blood pressure, possibly via increased AT1 receptor expression (32).
D4 Receptors and Blood Pressure Regulation Summary
D4 receptors may serve to antagonize vasopressin and aldosterone effects during conditions of normal salt intake. A role of D4 receptors in cardiovascular and renal physiology remains to be determined. However, disruption of the D4 receptor results in increased blood pressure that may be related to the activation of AT1 receptors in the brain. D4 receptors may negatively regulate the expression of AT1 receptors involved in the central regulation of blood pressure. However, areas in the brain where D4 receptors interact with AT1 receptors remain to be described. AT1 receptors are also increased in the Kidneys of D4−/− mice, but these mice do not have an impaired ability to excrete an acute sodium load. It is possible that D4−/− mice may not be able to excrete a chronic sodium load, but that remains to be determined.
D5 Receptors
The D5 receptor has generated significant interest because of its relatively high affinity for dopamine compared with the other dopamine receptors (229). Moreover, the D5 receptor can be activated in the absence or presence of low concentrations of endogenous agonist. The physiological role of the D5 receptor in the regulation of renal and cardiovascular function has been difficult to determine with certainty. This is due, in large part, to the fact that the D1 and D5 receptors are pharmacologically indistinguishable. As mentioned above, the lack of selective ligands has made it virtually impossible to selectively activate or block D1 or D5 receptors in vivo. Genetic approaches to this problem have been employed by investigators who used gene silencing techniques to inhibit D1 or D5 receptor expression and generate D1 or D5 receptor-deficient mice (5, 58, 93, 95, 223).
Localization of the D5 Receptor Subtype
The Kidney
In rats and mice, the D5 receptor is expressed in the proximal (S3 > S1 = S2) (unpublished observations) and distal convoluted tubules, cortical collecting duct, medullary ascending limb of Henle, and arterioles, but not in the glomeruli, juxtaglomerular cells, or macula densa (7, 9, 252, 254, 255, 280) (Table 1). The thick ascending limb of Henle and cortical collecting duct may also preferentially express the D5 receptor over the D1 receptor (7, 255). The opossum Kidney also expresses the D5 receptor, but its expression is lost in an opossum Kidney cell line (160). Immortalized rat and human proximal tubule cells express D5 receptors; the molecular sizes are similar to those described for the D1 receptor (259, 274).
Blood Vessels
The D5 receptor is expressed in VSMCs outside the Kidney, e.g., the coronary artery (7, 162). The vascular distribution of the D1 receptor is similar to the D5 receptor. As with the D1 receptor, the D5 receptor is expressed in pulmonary, mesenteric, and pial arteries (8, 192). In the pulmonary artery, D5 receptor immunostaining has been reported in the tunica intima and media of extrapulmonary branches and tunica media of large-sized (but not medium or small sized) intrapulmonary branches, similar to those described for the D1 receptor (196).
Physiology of the D5 Receptor
Renal Function
As indicated above, D1-like receptors induce a diuresis and natriuresis in WKY rats (104, 268). Due to the lack of selective D1 and D5 receptor agonists or antagonists, the relative contribution of D1 and D5 receptors to the natriuretic effect caused by D1-like receptor stimulation is not known. We have presumed that both D1 and D5 receptors are involved because both receptors increase cAMP production and cAMP mediates the D1-like receptor-mediated inhibition of ion transport (206). As indicated earlier, the D1 receptor increases cAMP production to a greater extent than the D5 receptor in renal proximal tubule cells (206); therefore, it is possible that the natriuretic effect of dopamine is mainly due to the D1 receptor. As also indicated above, the effect of a high-salt diet on blood pressure in D1−/− mice has not been determined. However, in D5 receptor-null (D5−/−) mice, a high-salt diet further increases blood pressure, suggesting that the renal D5 receptor plays an important role in the control of blood pressure by regulating renal salt transport (see below). However, the nephron segments in which the D5 receptor regulates ion transport remain to be determined. Because D5 receptor expression may be greater than D1 receptor expression in distal nephron segments (255), the D1-like receptor regulating sodium excretion in those nephron segments may be the D5 receptor.
D5 Receptor-Mediated Antioxidative Effects
Dopamine has contrasting, concentration-dependent effects on ROS production. High concentrations of dopamine and D1-like receptors agonists (2–300 μM) increase the generation of ROS (36, 147, 173, 237, 242). Excessive stimulation of D2-like receptors (e.g., D2 and D3) can also increase ROS production (246). However, D1-like and D2-like receptors act as antioxidants at physiologically relevant concentrations of dopamine and low concentrations of their respective agonists (51, 105, 204). It should also be noted that renal tubules do not normally synthesize dopamine at the high concentrations shown to induce oxidative stress (240).
Low concentrations of dopamine, acting on D1 and D5 receptors, decrease ROS in human lymphocytes (51), brain cortical cells (166), and renal VSMCs (258). Increased oxidative stress enhances vascular VSMC contractility and proliferation (190). The activation of both D1 and D5 receptors inhibits oxidative stress in VSMCs stimulated by PDGF-BB (258). We have reported that the D5 receptor inhibits the production of ROS by inhibition of PLD and NADPH oxidase expression and activity in HEK-293 cells heterologously expressing the human D5 receptor (252, 253). NADPH oxidase protein expression and activity in the Kidney and brain are higher in D5−/− mice than in control (D5+/+) mice. Apocynin, a drug that impairs the assembly and activity of NADPH oxidase subunits, normalizes blood pressure and NADPH oxidase activity and subunit expression in D5−/− mice. In addition, the D5 receptor increases the expression and activity of antioxidant enzymes. For example, in the same HEK-293 cells heterologously expressing the human D5 receptor, D5 receptor stimulation increases the activity and expression of heme oxygenase-1. Heme oxygenase-1 protein expression and activity in the Kidney are lower in D5−/−mice than in D5+/+ mice. Furthermore, hemin, a heme oxygenase inducer, normalizes blood pressure and renal heme oxygenase activity in D5−/− mice (143). Thus, the ability of the D5 receptor to decrease ROS production may explain, in part, its antihypertensive action.
D5 Receptors and Proliferation of VSMC
Proliferation of VSMCs is believed to play a key role in hypertension. Vasodilator hormones such as natriuretic peptides and β-adrenergic receptor agonists have been shown to act as antigrowth factors (1, 157). As aforementioned, D1-like receptors have also been shown to inhibit the proliferative effect of some hormones, such as PDGF-BB (258). This inhibition is reversed by a D1-like receptor antagonist and by D1 or D5 receptor antisense oligonucleotides, indicating that both D1 and D5 receptors have an antiproliferative effect in VSMCs. Vascular D1-like receptor agonists inhibit the proliferation of VSMCs, possibly through PKA activation and suppression of PLD, PKC, and MAPK activity (257). Inhibition of PLD and PKC is probably mediated by the D5 receptor (102, 243, 252). D1 receptors can stimulate PLC (69) and, therefore, PKC activity (63, 88, 101, 118, 167, 212, 256, 257), which can in turn increase D1 receptor function. In contrast, D5 receptors inhibit PKC activation (243); PKC can also inhibit D5 function (107).
D5 Receptors and Hypertension
D5 receptor gene locus (chromosome 4p15.1–16.1) is linked to essential hypertension (6); the human D5 receptor gene has polymorphisms that code for receptors with abnormal coupling to adenylyl cyclase (52).
D5 Receptor Mutant Mice
D5−/− mice were generated by injecting into C57BL/6 mouse blastocysts 129/SV embryonic stem cells containing the targeting construct generated by ligating the neomycin resistance gene in reverse orientation at the unique SfiI site in the second intracellular loop of the D5 receptor. D5−/− mice are viable and develop normally (93, 94). Disruption of the D5 receptor gene is not associated with an altered expression of the other dopamine receptors, including the D1 receptor (93, 94).
Hollon et al. (93) reported that D5−/− mice (>F6) are hypertensive, with an elevated epinephrine-to-norepinephrine ratio and a greater reduction in mean arterial pressure after adrenalectomy or treatment with an α-adrenergic blocker compared with D5+/+ mice. This study indicates that the hypertension is caused by increased sympathetic activity. However, because the percentage decrease in systolic blood pressure after adrenalectomy is similar in both D5−/− and D5+/+ mice, the hypertension in D5−/− mice is ascribed to central nervous system mechanisms (93, 273).
D5 receptors, present in the prefrontal cortex, project to several brain areas involved with cardiovascular regulation. Sympathetic responses from the prefrontal cortex, transmitted to the lateral hypothalamic area, stimulate non-NMDA glutamate receptors in the ventrolateral medulla (41, 150, 278). A centrally, but not peripherally, acting non-NMDA glutamatergic antagonist decreases blood pressure in D5−/− mice, suggesting that the increased blood pressure in D5−/− mice may be caused by the activation of a sympathetic/non-NMDA glutamatergic axis (93). V1 vasopressin and oxytocin antagonists that cross the blood-brain barrier also decrease blood pressure in D5−/− mice but not in D5+/+ mice. Interestingly, the hypotensive effect of the oxytocin antagonist occurs only 24 h after its administration and negates any further reduction in blood pressure by vasopressin or glutamatergic blockade. These results are consistent with the observation that oxytocin sensitizes V1 vasopressin receptors and further suggest that the decrease in blood pressure in D5−/− mice engendered by these various antagonists occurs via a central nervous system pathway involving glutaminergic, oxytocin, vasopressin, and adrenergic receptors (Fig. 1) (93, 273).
Fig. 1.

D2 and D5 receptors affect blood pressure by inhibition of the central sympathetic nervous system. D5 receptors are present in the prefrontal cortex, which projects to several brain areas involved with cardiovascular regulation, including the lateral hypothalamic area and ventrolateral medulla. D5 and D2 receptors affect blood pressure by decreasing the central sympathetic nervous system, although the detailed mechanisms remain to be determined. The dotted lines indicate inhibitory effects, whereas the solid lines indicate stimulatory effects.
The acute responses to GPCR-blocking agents are contrasted to the chronic regulation of blood pressure by the D5 receptor. Long-term (5–7 days) blockade of AT1 receptors decreases blood pressure in D5−/− mice but not in D5+/+ mice (136). This finding is in agreement with our observations of a counterregulatory interaction between D5 and AT1 receptors (274). The activation of D1-like receptors inhibits AT1 receptor expression (274); indirect evidence suggests that the D5 receptor may be involved in the inhibitory effects of a D1-like receptor agonist on the AT1 receptor (267). This is consistent with our finding in human renal proximal tubule cells showing that the D5 receptor but not the D1 receptor decreases AT1 receptor expression (74). The ability of the D5 receptor to negatively regulate AT1 receptor expression may have a significant impact on the regulation of blood pressure. Indeed, renal D5 receptor protein is increased in AT1A-deficient mice relative to their wild-type littermates (274). This effect is reciprocal: the activation of the AT1 receptor in renal proximal tubule cells also inhibits D5 receptor protein expression; in D5−/− mice, AT1 receptor expression in the renal cortical membrane is increased relative to wild-type littermates (274). Additional preliminary studies have shown that D5 receptors increase the degradation of AT1 receptors (74, 136). Because the D1 receptor may also regulate the function of the AT1 receptor by direct D1 and AT1 receptor interaction (267), an impaired regulation of AT1 receptors by both D1 and D5 receptors may be one of the mechanisms involved in the increased AT1 receptor function in SHRs.
D5 Receptors and Blood Pressure Regulation Summary
D5 receptors, being constitutively active, may play a greater role than D1 receptors in the basal regulation of blood pressure. The D5 receptor may acutely regulate blood pressure by inhibition of the central adrenergic nervous system, via the NMDAoxytocin-V1 receptor-adrenergic axis (Fig. 1). Long-term regulation of blood pressure may involve sodium transport and AT1 receptors and ROS; D5−/− mice are salt sensitive. Because D5 receptors are more numerous than D1 receptors in more distal nephron segments, the D5 receptor-mediated regulation of sodium transport may be exerted in the distal nephron.
Overall Summary
Dopamine regulates blood pressure by renal and nonrenal mechanisms, some of which are specific to the five dopamine receptor subtypes (104, 268). All dopamine receptor subtypes participate in the regulation of sodium balance, individually and by interacting with each other to increase sodium excretion under conditions of moderate sodium balance. All dopamine receptors, except the D4 receptor, have antioxidant functions. A deficiency in dopamine production and/or a dysfunction in dopamine receptors contribute to various forms of hypertension in both humans and animal models.
D1−/− mice have greater systolic, diastolic, and mean arterial pressure and do not increase cAMP production in response to D1-like receptor agonist stimulation (5). D5−/− mice develop hypertension via increased central sympathetic activity through activation of glutaminergic, oxytocin, vasopressin, and adrenergic receptors (93). Besides these central nervous system mechanisms, renal AT1 receptors and ROS are also involved (252, 253, 265, 274).
D4−/− mice have increased blood pressure, in part by increased AT1 receptor activity in the brain and Kidney (32). The increased blood pressure of D2−/− mice seems to be related to impaired renal dopamine production, impaired ability to excrete a chronic salt load, increased sympathetic activity, and increased ETB receptor expression in VSMCs (138, 179, 235). The increased blood pressure in D3−/− mice may be related to the activation of the renin-angiotensin-aldosterone system and a decreased ability to excrete a chronic sodium load (5, 225). Some discrepancies in the characteristics of different strains of D2−/− and D3−/− mice could be related to differences in genetic background. C57BL/6 mice may or may not increase their blood pressures in response to sodium intake depending on their genetic background (226, 253).
Finally, it seems that under conditions of salt depletion, dopamine, via D2 receptors, induces sodium retention, probably by stimulating sodium transport. In addition, the stimulatory effect of D1 receptors on the renin-angiotensin system may become manifest. During states of moderate sodium excess, dopamine receptors, individually or by interacting with each other, increase sodium excretion (Fig. 2). The inhibitory effect of D3 receptors also becomes manifest. Therefore, dopamine receptors may increase or decrease sodium transport to maintain sodium balance and blood pressure. Abnormalities in dopamine receptor subtypes, per se or caused by constitutively active variants of GRK4, result in hypertension. A major limitation of the studies in dopamine receptor subtype-null mice is the fact that the mutations are neither tissue specific nor inducible knockout mice. Studies in mice with inducible and tissue-specific deletion of dopamine receptor subtype genes are needed to decipher the exact role and mechanism(s) by which dopamine receptor subtypes regulate renal function and blood pressure.
Fig. 2.
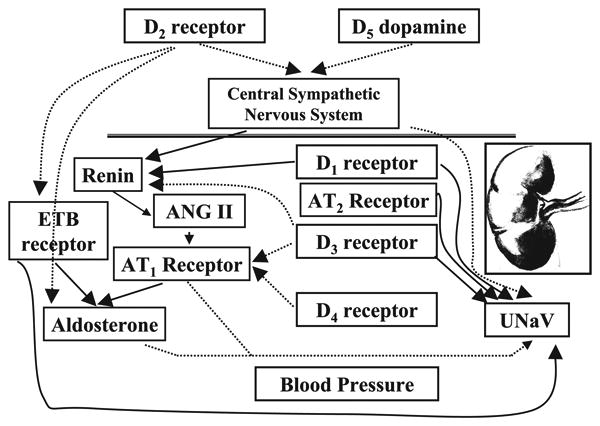
Dopamine receptors and cardiovascular function. Each of the dopamine receptor subtypes participates in the regulation of blood pressure by mechanisms specific for the subtype. The major dopamine receptor that regulates blood pressure may be the D1 receptor, which synergies with the D3 receptor to regulate sodium transport in the Kidney and intestines directly or indirectly by the inhibitory effect of D1 and D3 receptors on angiotensin II (ANG II) type 1 (AT1) receptor expression and/or interactions during conditions of modest sodium excess. D4 receptors help in this process. Furthermore, D3 receptors can inhibit renin secretion; D2 receptors aid in the excretion of sodium by decreasing aldosterone secretion and by inhibiting the vasconstrictor effect of endothelin type B (ETB) receptors. D2 and D5 receptors negatively regulate the sympathetic nervous system. UNaV, urinary excretion of sodium.
Acknowledgments
This work was supported in part by National Institutes of Health Grants HL-23081, DK-39308, HL-68686, DK-52612, and HL-41618 and by National Natural Science Foundation of China Grants 30470728 and 30672199.
References
- 1.Abell TJ, Richard AM, Ikram H, Espiner EA, Yandle Y. Atrial natriuretic factor inhibits proliferation of vascular smooth muscle cells stimulated by platelet-derived growth factor. Biochem Biophys Res Commun. 1989;160:1392–1396. doi: 10.1016/s0006-291x(89)80158-5. [DOI] [PubMed] [Google Scholar]
- 2.Accili D, Fishburn CS, Drago J, Steiner H, Lachowicz JE, Park BH, Gauda EB, Lee EJ, Cool MH, Sibley DR, Gerfen CR, Westphal H, Fuchs S. A targeted mutation of the D3 dopamine receptor gene is associated with hyperactivity in mice. Proc Natl Acad Sci USA. 1996;93:1945–1949. doi: 10.1073/pnas.93.5.1945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Agnoli GC, Borgatti R, Cacciari M, Garutti C, Ikonomu E, Lenzi P. Antagonistic effects of sulpiride–racemic and enantiomers–on renal response to low-dose dopamine infusion in normal women. Nephron. 1989;51:491–498. doi: 10.1159/000185382. [DOI] [PubMed] [Google Scholar]
- 4.Agnoli GC, Cacciari M, Garutti C, Ikonomu E, Lenzi P, Marchetti G. Effects of extracellular fluid volume changes on renal response to low-dose dopamine infusion in normal women. Clin Physiol. 1987;7:465–479. doi: 10.1111/j.1475-097x.1987.tb00189.x. [DOI] [PubMed] [Google Scholar]
- 5.Albrecht FE, Drago J, Felder RA, Printz MP, Eisner GM, Robillard JE, Sibley DR, Westphal HJ, Jose PA. Role of the D1A dopamine receptor in the pathogenesis of genetic hypertension. J Clin Invest. 1996;97:2283–2288. doi: 10.1172/JCI118670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Allayee H, de Bruin TW, Michelle Dominguez K, Cheng LS, Ipp E, Cantor RM, Krass KL, Keulen ET, Aouizerat BE, Lusis AJ, Rotter JI. Genome scan for blood pressure in Dutch dyslipidemic families reveals linkage to a locus on chromosome 4p. Hypertension. 2001;38:773–778. doi: 10.1161/hy1001.092617. [DOI] [PubMed] [Google Scholar]
- 7.Amenta F. Light microscope autoradiography of peripheral dopamine receptor subtypes. Clin Exp Hypertens. 1997;19:27–41. doi: 10.3109/10641969709080802. [DOI] [PubMed] [Google Scholar]
- 8.Amenta F, Barili P, Bronzetti E, Felici L, Mignini F, Ricci A. Localization of dopamine receptor subtypes in systemic arteries. Clin Exp Hypertens. 2000;22:277–288. doi: 10.1081/ceh-100100077. [DOI] [PubMed] [Google Scholar]
- 9.Amenta F, Barili P, Bronzetti E, Ricci A. Dopamine D1-like receptor subtypes in the rat Kidney: a microanatomical study. Clin Exp Hypertens. 1999;21:17–23. doi: 10.3109/10641969909068645. [DOI] [PubMed] [Google Scholar]
- 10.Aoki Y, Albrecht FE, Bergman KR, Jose PA. Stimulation of Na+-K+-2Cl- cotransport in rat medullary thick ascending limb by dopamine. Am J Physiol Regul Integr Comp Physiol. 1996;271:R1561–R1567. doi: 10.1152/ajpregu.1996.271.6.R1561. [DOI] [PubMed] [Google Scholar]
- 11.Aperia A, Bertorello A, Seri I. Dopamine causes inhibition of Na+-K+-ATPase activity in rat proximal convoluted tubule segments. Am J Physiol Renal Fluid Electrolyte Physiol. 1987;252:F39–F45. doi: 10.1152/ajprenal.1987.252.1.F39. [DOI] [PubMed] [Google Scholar]
- 12.Aperia A, Eklof AC, Holtback U, Nowicki S, Sundelof M, Greengard P. The renal dopamine system. Adv Pharmacol. 1998;42:870–873. doi: 10.1016/s1054-3589(08)60885-6. [DOI] [PubMed] [Google Scholar]
- 13.Armando I, Wang X, Pascua AM, Villar VM, Luo Y, Jones JE, Asico L, Escano C, Jose PA. Regulation of Renal NADPH Oxidase and Inflammation by Dopamine D2 Receptors. Orlando, FL: American Heart Association Scientific Sessions; Nov, 2007. [Google Scholar]
- 14.Armando I, Wang X, Villar VA, Jones JE, Asico LD, Escano C, Jose PA. Reactive oxygen species-dependent hypertension in dopamine D2 receptor-deficient mice. Hypertension. 2007;49:672–678. doi: 10.1161/01.HYP.0000254486.00883.3d. [DOI] [PubMed] [Google Scholar]
- 15.Asghar M, Hussain T, Lokhandwala MF. Activation of dopamine D1-like receptor causes phosphorylation of alpha1-subunit of Na+,K+-ATPase in rat renal proximal tubules. Eur J Pharmacol. 2001;411:61–66. doi: 10.1016/s0014-2999(00)00896-7. [DOI] [PubMed] [Google Scholar]
- 16.Asghar M, Hussain T, Lokhandwala MF. Overexpression of PKC-βI and -δ contributes to higher PKC activity in the proximal tubules of old Fischer 344 rats. Am J Physiol Renal Physiol. 2003;285:F1100–F1107. doi: 10.1152/ajprenal.00198.2003. [DOI] [PubMed] [Google Scholar]
- 17.Asico LD, Ladines C, Fuchs S, Accili D, Carey RM, Semeraro C, Pocchiari F, Felder RA, Eisner GM, Jose PA. Disruption of the dopamine D3 receptor gene produces renin-dependent hypertension. J Clin Invest. 1998;102:493–498. doi: 10.1172/JCI3685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Assuncao-Miranda I, Guilherme AL, Reis-Silva C, Costa-Sarmento G, Oliveira MM, Vieyra A. Protein kinase C-mediated inhibition of renal Ca2+ ATPase by physiological concentrations of angiotensin II is reversed by AT1- and AT2-receptor antagonists. Regul Pept. 2005;127:151–157. doi: 10.1016/j.regpep.2004.11.004. [DOI] [PubMed] [Google Scholar]
- 19.Azdad K, Piet R, Poulain DA, Oliet SH. Dopamine D4 receptor-mediated presynaptic inhibition of GABAergic transmission in the rat supraoptic nucleus. J Neurophysiol. 2003;90:559–565. doi: 10.1152/jn.00226.2003. [DOI] [PubMed] [Google Scholar]
- 20.Bacic D, Capuano P, Baum M, Zhang J, Stange G, Biber J, Kaissling B, Moe OW, Wagner CA, Murer H. Activation of dopamine D1-like receptors induces acute internalization of the renal Na+/phosphate co-transporter NaPi-IIa in mouse Kidney and OK cells. Am J Physiol Renal Physiol. 2005;288:F740–F747. doi: 10.1152/ajprenal.00380.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bacic D, Kaissling B, McLeroy P, Zou L, Baum M, Moe OW. Dopamine acutely decreases apical membrane Na/H exchanger NHE3 protein in mouse renal proximal tubule. Kidney Int. 2003;64:2133–2141. doi: 10.1046/j.1523-1755.2003.00308.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Baik JH, Picetti R, Saiardi A, Thiriet G, Dierich A, Depaulis A, Le Meur M, Borrelli E. Parkinsonian-like locomotor impairment in mice lacking dopamine D2 receptors. Nature. 1995;377:424–428. doi: 10.1038/377424a0. [DOI] [PubMed] [Google Scholar]
- 23.Baines AD, Drangova R. Does dopamine use several signal pathways to inhibit Na-Pi transport in OK cells? J Am Soc Nephrol. 1998;9:1604–1612. doi: 10.1681/ASN.V991604. [DOI] [PubMed] [Google Scholar]
- 24.Banday AA, Hussain T, Lokhandwala MF. Renal dopamine D1 receptor dysfunction is acquired and not inherited in obese Zucker rats. Am J Physiol Renal Physiol. 2004;287:F109–F116. doi: 10.1152/ajprenal.00396.2003. [DOI] [PubMed] [Google Scholar]
- 25.Barili P, Fringuelli C, Baldoni E, Mignini F, Zaccheo D, Amenta F. Dopamine D2-like receptors in the Kidney of spontaneously hypertensive rats: a radioligand binding assay and light microscope autoradiography study. J Auton Pharmacol. 1998;18:89–97. doi: 10.1046/j.1365-2680.1998.1820089.x. [DOI] [PubMed] [Google Scholar]
- 26.Barili P, Ricci A, Baldoni E, Mignini F, Amenta F. Pharmacological characterizsation and autoradiographic localisation of a putative dopamine D3 receptor in the rat Kidney. Eur J Pharmacol. 1997;338:89–95. doi: 10.1016/s0014-2999(97)01303-4. [DOI] [PubMed] [Google Scholar]
- 27.Barili P, Sabbatini M, Soares-da-Silva P, Amenta F. The peripheral dopaminergic system: morphological analysis, functional and clinical applications. Ital J Anat Embryol. 2002;107:145–167. [PubMed] [Google Scholar]
- 28.Bastianetto S, Danik M, Mennicken F, Williams S, Quirion R. Prototypical antipsychotic drugs protect hippocampal neuronal cultures against cell death induced by growth medium deprivation. BMC Neurosci. 2006;7:28. doi: 10.1186/1471-2202-7-28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Beazely MA, Watts VJ. Activation of a novel PKC isoform synergistically enhances D2L dopamine receptor-mediated sensitization of adenylate cyclase type 6. Cell Signal. 2005;17:647–653. doi: 10.1016/j.cellsig.2004.10.003. [DOI] [PubMed] [Google Scholar]
- 30.Beige J, Bellmann A, Sharma AM, Gessner R. Ethnic origin determines the impact of genetic variants in dopamine receptor gene (DRD1) concerning essential hypertension. Am J Hypertens. 2004;17:1184–1187. doi: 10.1016/j.amjhyper.2004.07.013. [DOI] [PubMed] [Google Scholar]
- 31.Bek M, Fischer KG, Greiber S, Hupfer C, Mundel P, Pavenstadt H. Dopamine depolarizes podocytes via a D1-like receptor. Nephrol Dial Transplant. 1999;14:581–587. doi: 10.1093/ndt/14.3.581. [DOI] [PubMed] [Google Scholar]
- 32.Bek MJ, Wang X, Asico LD, Jones JE, Zheng S, Li X, Eisner GM, Grandy DK, Carey RM, Soares-da-Silva P, Jose PA. Angiotensin-II type 1 receptor-mediated hypertension in D4 dopamine receptor-deficient mice. Hypertension. 2006;47:288–295. doi: 10.1161/01.HYP.0000198427.96225.36. [DOI] [PubMed] [Google Scholar]
- 33.Bek MJ, Zheng S, Xu J, Yamaguchi I, Asico LD, Sun XG, Jose PA. Differential expression of adenylyl cyclases in the rat nephron. Kidney Int. 2001;60:890–899. doi: 10.1046/j.1523-1755.2001.060003890.x. [DOI] [PubMed] [Google Scholar]
- 34.Bertorello A, Aperia A. Inhibition of proximal tubule Na+-K+-ATPase activity requires simultaneous activation of DA1 and DA2 receptors. Am J Physiol Renal Fluid Electrolyte Physiol. 1990;259:F924–F928. doi: 10.1152/ajprenal.1990.259.6.F924. [DOI] [PubMed] [Google Scholar]
- 35.Bertorello AM, Hopfield JF, Aperia A, Greengard P. Inhibition by dopamine of (Na+-K+)ATPase activity in neostriatal neurons through D1 and D2 dopamine receptor synergism. Nature. 1990;347:386–388. doi: 10.1038/347386a0. [DOI] [PubMed] [Google Scholar]
- 36.Bianchi P, Seguelas MH, Parini A, Cambon C. Activation of proapoptotic cascade by dopamine in renal epithelial cells is fully dependent on hydrogen peroxide generation by monoamine oxidases. J Am Soc Nephrol. 2003;14:855–862. doi: 10.1097/01.asn.0000058909.00567.5c. [DOI] [PubMed] [Google Scholar]
- 37.Bouvier M. Oligomerization of G-protein-coupled transmitter receptors. Nat Rev Neurosci. 2001;2:274–286. doi: 10.1038/35067575. [DOI] [PubMed] [Google Scholar]
- 38.Brismar H, Asghar M, Carey RM, Greengard P, Aperia A. Dopamine-induced recruitment of dopamine D1 receptors to the plasma membrane. Proc Natl Acad Sci USA. 1998;95:5573–5578. doi: 10.1073/pnas.95.10.5573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bughi S, Horton R, Antonipillai I, Manoogian C, Ehrlich L, Nadler J. Comparison of dopamine and fenoldopam effects on renal blood flow and prostacyclin excretion in normal and essential hypertensive subjects. J Clin Endocrinol Metab. 1989;69:1116–1121. doi: 10.1210/jcem-69-6-1116. [DOI] [PubMed] [Google Scholar]
- 40.Canals M, Marcellino D, Fanelli F, Ciruela F, de Benedetti P, Goldberg SR, Neve K, Fuxe K, Agnati LF, Woods AS, Ferré S, Lluis C, Bouvier M, Franco R. Adenosine A2A-dopamine D2 receptor-receptor heteromerization: qualitative and quantitative assessment by fluorescence and bioluminescence energy transfer. J Biol Chem. 2003;278:46741–46749. doi: 10.1074/jbc.M306451200. [DOI] [PubMed] [Google Scholar]
- 41.Carvey PM, McGuire SO, Ling ZD. Neuroprotective effects of D3 dopamine receptor agonists. Parkinsonism Relat Disord. 2001;7:213–223. doi: 10.1016/s1353-8020(00)00061-4. [DOI] [PubMed] [Google Scholar]
- 42.Carey RM, Siragy HM, Ragsdale NV, Howell NL, Felder RA, Peach MJ, Chevalier RL. Dopamine-1 and dopamine-2 mechanisms in the control of renal function. Am J Hypertens. 1990;3:59S–63S. doi: 10.1093/ajh/3.6.59s. [DOI] [PubMed] [Google Scholar]
- 43.Chatziantoniou C, Ruan X, Arendshorst WJ. Interactions of cAMP-mediated vasodilators with angiotensin II in rat Kidney during hypertension. Am J Physiol Renal Fluid Electrolyte Physiol. 1993;265:F845–F852. doi: 10.1152/ajprenal.1993.265.6.F845. [DOI] [PubMed] [Google Scholar]
- 44.Cavallotti C, Nuti F, Bruzzone P, Mancone M. Age-related changes in dopamine D2 receptors in rat heart and coronary Vessels. Clin Exp Pharmacol Physiol. 2002;29:412–418. doi: 10.1046/j.1440-1681.2002.03677.x. [DOI] [PubMed] [Google Scholar]
- 45.Chen C, Beach RE, Lokhandwala MF. Dopamine fails to inhibit renal tubular sodium pump in hypertensive rats. Hypertension. 1993;21:364–372. doi: 10.1161/01.hyp.21.3.364. [DOI] [PubMed] [Google Scholar]
- 46.Chen C, Lokhandwala MF. Potentiation by enalaprilat of fenoldopamevoked natriuresis is due to blockade of intrarenal production of angiotensin-II in rats. Naunyn Schmiedebergs Arch Pharmacol. 1995;352:194–200. doi: 10.1007/BF00176774. [DOI] [PubMed] [Google Scholar]
- 47.Chen CJ, Lokhandwala MF. An impairment of renal tubular DA-1 receptor function as the causative factor for diminished natriuresis to volume expansion in spontaneously hypertensive rats. Clin Exp Hypertens. 1992;14:615–628. doi: 10.3109/10641969209036211. [DOI] [PubMed] [Google Scholar]
- 48.Cheng HF, Becker BN, Burns KD, Harris RC. Angiotensin II upregulates type-1 angiotensin II receptors in renal proximal tubule. J Clin Invest. 1995;95:2012–2019. doi: 10.1172/JCI117886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Cheng HF, Becker BN, Harris RC. Dopamine decreases expression of type-1 angiotensin II receptors in renal proximal tubule. J Clin Invest. 1996;97:2745–2752. doi: 10.1172/JCI118729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cho DI, Quan W, Oak MH, Choi HJ, Lee KY, Kim KM. Functional interaction between dopamine receptor subtypes for the regulation of c-fos expression. Biochem Biophys Res Commun. 2007;357:1113–1138. doi: 10.1016/j.bbrc.2007.04.066. [DOI] [PubMed] [Google Scholar]
- 51.Cosentino M, Rasini E, Colombo C, Marino F, Blandini F, Ferrari M, Samuele A, Lecchini S, Nappi G, Frigo G. Dopaminergic modulation of oxidative stress and apoptosis in human peripheral blood lymphocytes: evidence for a D1-like receptor-dependent protective effect. Free Radic Biol Med. 2004;36:1233–12340. doi: 10.1016/j.freeradbiomed.2004.02.065. [DOI] [PubMed] [Google Scholar]
- 52.Cravchik A, Gejman PV. Functional analysis of the human D5 dopamine receptor missense and nonsense variants: differences in dopamine binding affinities. Pharmacogenetics. 1999;9:199–206. [PubMed] [Google Scholar]
- 53.Crowley SD, Gurley SB, Herrera MJ, Ruiz P, Griffiths R, Kumar AP, Kim HS, Smithies O, Le TH, Coffman TM. Angiotensin II causes hypertension and cardiac hypertrophy through its receptors in the Kidney. Proc Natl Acad Sci USA. 2006;103:17985–17990. doi: 10.1073/pnas.0605545103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.DiBona GF. Sympathetic nervous system and the Kidney in hypertension. Curr Opin Nephrol Hypertens. 2002;11:197–200. doi: 10.1097/00041552-200203000-00011. [DOI] [PubMed] [Google Scholar]
- 55.Dickson ME, Sigmund CD. Genetic basis of hypertension: revisiting angiotensinogen. Hypertension. 2006;48:14–20. doi: 10.1161/01.HYP.0000227932.13687.60. [DOI] [PubMed] [Google Scholar]
- 56.Dickinson SD, Sabeti J, Larson GA, Giardina K, Rubinstein M, Kelly MA, Grandy DK, Low MJ, Gerhardt GA, Zahniser NR. Dopamine D2 receptor-deficient mice exhibit decreased dopamine transporter function but no changes in dopamine release in dorsal striatum. J Neurochem. 1999;72:148–156. doi: 10.1046/j.1471-4159.1999.0720148.x. [DOI] [PubMed] [Google Scholar]
- 57.Done SC, Leibiger IB, Efendiev R, Katz AI, Leibiger B, Berggren PO, Pedemonte CH, Bertorello AM. Tyrosine 537 within the Na+,K+-ATPase alpha-subunit is essential for AP-2 binding and clathrin-dependent endocytosis. J Biol Chem. 2002;277:17108–17111. doi: 10.1074/jbc.M201326200. [DOI] [PubMed] [Google Scholar]
- 58.Drago J, Gerfen CR, Lachowicz JE, Steiner H, Hollon TR, Love PE, Ooi GT, Grinberg A, Lee EJ, Huang SP, Bartlett PF, Jose PA, Sibley DR, Westphal H. Altered striatal function in a mutant mouse lacking D1A dopamine receptors. Proc Natl Acad Sci USA. 1994;91:12564–12568. doi: 10.1073/pnas.91.26.12564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Dunah AW, Standaert DG. Dopamine D1 receptor-dependent trafficking of striatal NMDA glutamate receptors to the postsynaptic membrane. J Neurosci. 2001;21:5546–5558. doi: 10.1523/JNEUROSCI.21-15-05546.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Dziedzicka-Wasylewska M, Faron-Gorecka A, Andrecka J, Polit A, Kusmider M, Wasylewski Z. Fluorescence studies reveal heterodimerization of dopamine D1 and D2 receptors in the plasma membrane. Biochemistry. 2006;45:8751–8759. doi: 10.1021/bi060702m. [DOI] [PubMed] [Google Scholar]
- 61.Edvinsson L, McCulloch J, Sharkey J. Vasomotor responses of cerebral arterioles in situ to putative dopamine receptor agonists. Br J Pharmacol. 1985;85:403–410. doi: 10.1111/j.1476-5381.1985.tb08875.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Edwards RM, Brooks DP. Dopamine inhibits vasopressin action in the rat inner medullary collecting duct via α2-adrenoceptors. J Pharmacol Exp Ther. 2001;298:1001–1006. [PubMed] [Google Scholar]
- 63.Efendiev R, Bertorello AM, Pedemonte CH. PKC-beta and PKC-zeta mediate opposing effects on proximal tubule Na+,K+-ATPase activity. FEBS Lett. 1999;456:45–48. doi: 10.1016/s0014-5793(99)00925-4. [DOI] [PubMed] [Google Scholar]
- 64.Efendiev R, Budu CE, Cinelli AR, Bertorello AM, Pedemonte CH. Intracellular Na+ regulates dopamine and angiotensin II receptors availability at the plasma membrane and their cellular responses in renal epithelia. J Biol Chem. 2003;278:28719–28726. doi: 10.1074/jbc.M303741200. [DOI] [PubMed] [Google Scholar]
- 65.Efendiev R, Cinelli AR, Leibiger IB, Bertorello AM, Pedemonte CH. FRET analysis reveals a critical conformational change within the Na,K-ATPase alpha1 subunit N-terminus during GPCR-dependent endocytosis. FEBS Lett. 2006;580:5067–5070. doi: 10.1016/j.febslet.2006.08.035. [DOI] [PubMed] [Google Scholar]
- 66.Eklof AC. The natriuretic response to a dopamine DA1 agonist requires endogenous activation of dopamine DA2 receptors. Acta Physiol Scand. 1997;160:311–314. doi: 10.1046/j.1365-201X.1997.00166.x. [DOI] [PubMed] [Google Scholar]
- 67.Everett PB, Senogles SE. D3 dopamine receptor activates phospholipase D through a pertussis toxin-insensitive pathway. Neurosci Lett. 2004;371:34–39. doi: 10.1016/j.neulet.2004.08.033. [DOI] [PubMed] [Google Scholar]
- 68.Felder CC, Campbell T, Albrecht F, Jose PA. Dopamine inhibits Na+-H+ exchanger activity in renal BBMV by stimulation of adenylate cyclase. Am J Physiol Renal Fluid Electrolyte Physiol. 1990;259:F297–F303. doi: 10.1152/ajprenal.1990.259.2.F297. [DOI] [PubMed] [Google Scholar]
- 69.Felder CC, Jose PA, Axelrod J. The dopamine-1 agonist, SKF 82526, stimulates phospholipase-C activity independent of adenylate cyclase. J Pharmacol Exp Ther. 1989;248:171–175. [PubMed] [Google Scholar]
- 70.Felder RA, Blecher M, Eisner GM, Jose PA. Cortical tubular and glomerular dopamine receptors in the rat Kidney. Am J Physiol Renal Fluid Electrolyte Physiol. 1984;246:F557–F568. doi: 10.1152/ajprenal.1984.246.5.F557. [DOI] [PubMed] [Google Scholar]
- 71.Felder RA, Jose PA. Dopamine1 receptors in rat Kidneys identified with 125I-Sch 23982. Am J Physiol Renal Fluid Electrolyte Physiol. 1988;255:F970–F976. doi: 10.1152/ajprenal.1988.255.5.F970. [DOI] [PubMed] [Google Scholar]
- 72.Felder RA, Kinoshita S, Ohbu K, Mouradian MM, Sibley DR, Monsma FJ, Jr, Minowa T, Minowa MT, Canessa LM, Jose PA. Organ specificity of the dopamine1 receptor/adenylyl cyclase coupling defect in spontaneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol. 1993;264:R726–R732. doi: 10.1152/ajpregu.1993.264.4.R726. [DOI] [PubMed] [Google Scholar]
- 73.Felder RA, Seikaly MG, Cody P, Eisner GM, Jose PA. Attenuated renal response to dopaminergic drugs in spontaneously hypertensive rats. Hypertension. 1990;15:560–569. doi: 10.1161/01.hyp.15.6.560. [DOI] [PubMed] [Google Scholar]
- 74.Felder RA, Wang X, Gildea J, Bengra C, Sasaki M, Zeng C, Jones JE, Zheng W, Asico LD, Jose PA. Human renal angiotensin type 1 receptor regulation by the D1 dopamine receptor (Abstract) Hypertension. 2003;42:438A. [Google Scholar]
- 75.Ferré S, Ciruela F, Canals M, Marcellino D, Burgueno J, Casadó V, Hillion J, Torvinen M, Fanelli F, de Benedetti P, Goldberg SR, Bouvier M, Fuxe K, Agnati LF, Lluis C, Franco R, Woods A. Adenosine A2A-dopamine D2 receptor-receptor heteromers. Targets for neuro-psychiatric disorders. Parkinsonism Relat Disord. 2004;10:265–271. doi: 10.1016/j.parkreldis.2004.02.014. [DOI] [PubMed] [Google Scholar]
- 76.Fink GD. Hypothesis: the systemic circulation as a regulated free-market economy. A new approach for understanding the long-term control of blood pressure. Clin Exp Pharmacol Physiol. 2005;32:377–383. doi: 10.1111/j.1440-1681.2005.04219.x. [DOI] [PubMed] [Google Scholar]
- 77.Fiorentini C, Gardoni F, Spano P, Di Luca M, Missale C. Regulation of dopamine D1 receptor trafficking and desensitization by oligomerization with glutamate N -methyl-d-aspartate receptors. J Biol Chem. 2003;278:20196–20202. doi: 10.1074/jbc.M213140200. [DOI] [PubMed] [Google Scholar]
- 78.Fryckstedt J, Meister B, Aperia A. Control of electrolyte transport in the Kidney through a dopamine- and cAMP-regulated phosphoprotein, DARPP-32. J Auton Pharmacol. 1992;12:183–189. doi: 10.1111/j.1474-8673.1992.tb00376.x. [DOI] [PubMed] [Google Scholar]
- 79.Gao DQ, Canessa LM, Mouradian MM, Jose PA. Expression of the D2 subfamily of dopamine receptor genes in Kidney. Am J Physiol Renal Fluid Electrolyte Physiol. 1994;266:F646–F650. doi: 10.1152/ajprenal.1994.266.4.F646. [DOI] [PubMed] [Google Scholar]
- 80.Gines S, Hillion J, Torvinen M, Le Crom S, Casado V, Canela EI, Rondin S, Lew JY, Watson S, Zoli M, Agnati LF, Verniera P, Lluis C, Ferre S, Fuxe K, Franco R. Dopamine and adenosine A1 receptors form functionally interacting heterotrimeric complexes. Proc Natl Acad Sci USA. 2000;97:8606–8611. doi: 10.1073/pnas.150241097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Glickstein SB, Schmauss C. Dopamine receptor functions: lessons from knockout mice [corrected] Pharmacol Ther. 2001;91:63–83. doi: 10.1016/s0163-7258(01)00145-0. [DOI] [PubMed] [Google Scholar]
- 82.Gomes P, Soares-Da-Silva P. D2-like receptor-mediated inhibition of Na+-K+-ATPase activity is dependent on the opening of K+ channels. Am J Physiol Renal Physiol. 2002;283:F114–F123. doi: 10.1152/ajprenal.00244.2001. [DOI] [PubMed] [Google Scholar]
- 83.Gomez Mde J, Rousseau G, Nadeau R, Berra R, Flores G, Suarez J. Functional and autoradiographic characterization of dopamine D2-like receptors in the guinea pig heart. Can J Physiol Pharmacol. 2002;80:578–587. doi: 10.1139/y02-081. [DOI] [PubMed] [Google Scholar]
- 84.Granger JP, Schnackenberg CG. Renal mechanisms of angiotensin II-induced hypertension. Semin Nephrol. 2000;20:417–425. [PubMed] [Google Scholar]
- 85.Grider J, Kilpatrick E, Ott C, Jackson B. Effect of dopamine on NaCl transport in the medullary thick ascending limb of the rat. Eur J Pharmacol. 1998;342:281–284. doi: 10.1016/s0014-2999(97)01564-1. [DOI] [PubMed] [Google Scholar]
- 86.Grider JS, Ott CE, Jackson BA. Dopamine D1 receptor-dependent inhibition of NaCl transport in the rat thick ascending limb: mechanism of action. Eur J Pharmacol. 2003;473:185–190. doi: 10.1016/s0014-2999(03)01965-4. [DOI] [PubMed] [Google Scholar]
- 87.Hall JE. The Kidney, hypertension, and obesity. Hypertension. 2003;41:625–633. doi: 10.1161/01.HYP.0000052314.95497.78. [DOI] [PubMed] [Google Scholar]
- 88.Han JY, Heo JS, Lee YJ, Lee JH, Taub M, Han HJ. Dopamine stimulates 45Ca2+ uptake through cAMP, PLC/PKC, and MAPKs in renal proximal tubule cells. J Cell Physiol. 2007;211:486–494. doi: 10.1002/jcp.20956. [DOI] [PubMed] [Google Scholar]
- 89.Harrison-Bernard LM, Zhuo J, Kobori H, Ohishi M, Navar LG. Intrarenal AT1 receptor and ACE binding in ANG II-induced hypertensive rats. Am J Physiol Renal Physiol. 2002;282:F19–F25. doi: 10.1152/ajprenal.00335.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Hedge SS, Ricci A, Amenta F, Lokhandwala MF. Evidence from functional and autoradiographic studies for the presence of tubular dopamine-1 receptors and their involvement in the renal effects of fenoldopam. J Pharmacol Exp Ther. 1989;251:1237–1245. [PubMed] [Google Scholar]
- 91.Hellstrand M, Danielsen EA, Steen VM, Ekman A, Eriksson E, Nilsson CL. The ser9gly SNP in the dopamine D3 receptor causes a shift from cAMP related to PGE2 related signal transduction mechanisms in transfected CHO cells. J Med Genet. 2004;41:867–871. doi: 10.1136/jmg.2004.020941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Hillion J, Canals M, Torvinen M, Casado V, Scott R, Terasmaa A, Hansson A, Watson S, Olah ME, Mallol J. Coaggregation, cointernalization and codesensitization of adenosine A2A receptors and dopamine D2 receptors. J Biol Chem. 2002;277:18091–18097. doi: 10.1074/jbc.M107731200. [DOI] [PubMed] [Google Scholar]
- 93.Hollon TR, Bek MJ, Lachowicz JE, Ariano MA, Mezey E, Ramachandran R, Wersinger SR, Soares-da-Silva P, Liu ZF, Grinberg A, Drago J, Young WS, 3rd, Westphal H, Jose PA, Sibley DR. Mice lacking D5 dopamine receptors have increased sympathetic tone and are hypertensive. J Neurosci. 2002;22:10801–10810. doi: 10.1523/JNEUROSCI.22-24-10801.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Holmes A, Hollon TR, Gleason TC, Liu Z, Dreiling J, Sibley DR, Crawley JN. Behavioral characterization of dopamine D5 receptor null mutant mice. Behav Neurosci. 2001;115:1129–1144. [PubMed] [Google Scholar]
- 95.Holmes A, Lachowicz JE, Sibley DR. Phenotypic analysis of dopamine receptor knockout mice; recent insights into the functional specificity of dopamine receptor subtypes. Neuropharmacology. 2004;47:1117–1134. doi: 10.1016/j.neuropharm.2004.07.034. [DOI] [PubMed] [Google Scholar]
- 96.Horiuchi A, Takeyasu K, Mouradian MM, Jose PA, Felder RA. D1A dopamine receptor stimulation inhibits Na+-K+ ATPase activity through protein kinase A. Mol Pharmacol. 1993;43:281–285. [PubMed] [Google Scholar]
- 97.Hu MC, Fan L, Crowder LA, Karim-Jimenez Z, Murer H, Moe OW. Dopamine acutely stimulates Na+/H+ exchanger (NHE3) endocytosis via clathrin-coated vesicles: dependence on protein kinase A-mediated NHE3 phosphorylation. J Biol Chem. 2001;276:26906–26915. doi: 10.1074/jbc.M011338200. [DOI] [PubMed] [Google Scholar]
- 98.Huo T, Healy DP. Autoradiographic localization of dopamine DA1 receptors in rat Kidney with [3H]Sch 23390. Am J Physiol Renal Fluid Electrolyte Physiol. 1989;257:F414–F423. doi: 10.1152/ajprenal.1989.257.3.F414. [DOI] [PubMed] [Google Scholar]
- 99.Hussain T, Abdul-Wahab R, Kotak DK, Lokhandwala MF. Bromocriptine regulates angiotensin II response on sodium pump in proximal tubules. Hypertension. 1998;32:1054–1059. doi: 10.1161/01.hyp.32.6.1054. [DOI] [PubMed] [Google Scholar]
- 100.Hussain T, Abdul-Wahab R, Lokhandwala MF. Bromocriptine stimulates Na+, K+-ATPase in renal proximal tubules via the cAMP pathway. Eur J Pharmacol. 1997;321:259–263. doi: 10.1016/s0014-2999(97)00039-3. [DOI] [PubMed] [Google Scholar]
- 101.Hussain T, Lokhandwala MF. Altered arachidonic acid metabolism contributes to the failure of dopamine to inhibit Na+,K+-ATPase in Kidney of spontaneously hypertensive rats. Clin Exp Hypertens. 1996;18:963–974. doi: 10.3109/10641969609097911. [DOI] [PubMed] [Google Scholar]
- 102.Hussain T, Lokhandwala MF. Dopamine-1 receptor G-protein coupling and the involvement of phospholipase A2 in dopamine-1 receptor mediated cellular signaling mechanisms in the proximal tubules of SHR. Clin Exp Hypertens. 1997;19:131–140. doi: 10.3109/10641969709080810. [DOI] [PubMed] [Google Scholar]
- 103.Hussain T, Lokhandwala MF. Renal dopamine D1A receptor coupling with GS and Gq/11 proteins in spontaneously hypertensive rats. Am J Physiol Renal Physiol. 1997;272:F339–F346. doi: 10.1152/ajprenal.1997.272.3.F339. [DOI] [PubMed] [Google Scholar]
- 104.Hussain T, Lokhandwala MF. Renal dopamine receptors and hypertension. Exp Biol Med (Maywood) 2003;228:134–142. doi: 10.1177/153537020322800202. [DOI] [PubMed] [Google Scholar]
- 105.Ishige K, Chen Q, Sagara Y, Schubert D. The activation of dopamine D4 receptors inhibits oxidative stress-induced nerve cell death. J Neurosci. 2001;21:6069–6076. doi: 10.1523/JNEUROSCI.21-16-06069.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Izquierdo-Claros RM, Boyano-Adanez MC, Larsson C, Gustavsson L, Arilla E. Acute effects of D1- and D2-receptor agonist and antagonist drugs on somatostatin binding, inhibition of adenylyl cyclase activity and accumulation of inositol 1,4,5-trisphosphate in the rat striatum. Brain Res Mol Brain Res. 1997;47:99–107. doi: 10.1016/s0169-328x(97)00063-6. [DOI] [PubMed] [Google Scholar]
- 107.Jackson A, Sedaghat K, Minerds K, James C, Tiberi M. Opposing effects of phorbol-12-myristate-13-acetate, an activator of protein kinase C, on the signaling of structurally related human dopamine D1 and D5 receptors. J Neurochem. 2005;95:1387–1400. doi: 10.1111/j.1471-4159.2005.03476.x. [DOI] [PubMed] [Google Scholar]
- 108.James K, Weitzel LR, Engelman CD, Zerbe G, Norris JM Framing-ham Heart Study. Genome scan linkage results for longitudinal systolic blood pressure phenotypes in subjects from the Framingham Heart Study. BMC Genet. 2003;4(Suppl 1):S83. doi: 10.1186/1471-2156-4-S1-S83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Jarvie KR, Booth G, Brown EM, Niznik HB. Glycoprotein nature of dopamine D1 receptors in the brain and parathyroid gland. Mol Pharmacol. 1989;36:566–574. [PubMed] [Google Scholar]
- 110.Jin XH, Zhang JZ, Zhao YJ, Zhao RR. Localization of dopamine1A receptor mRNA in different vascular beds in rat: a nonradioactive in situ hybridization study. Methods Find Exp Clin Pharmacol. 1997;19:657–663. [PubMed] [Google Scholar]
- 111.Jomphe C, Tiberi M, Trudeau LE. Expression of D2 receptor isoforms in cultured neurons reveals equipotent autoreceptor function. Neuropharmacology. 2006;50:595–605. doi: 10.1016/j.neuropharm.2005.11.010. [DOI] [PubMed] [Google Scholar]
- 112.Jose PA, Asico LD, Eisner GM, Pocchiari F, Semeraro C, Felder RA. Effects of costimulation of dopamine D1- and D2-like receptors on renal function. Am J Physiol Regul Integr Comp Physiol. 1998;275:R986–R994. doi: 10.1152/ajpregu.1998.275.4.R986. [DOI] [PubMed] [Google Scholar]
- 113.Jose PA, Eisner GM, Drago J, Carey RM, Felder RA. Dopamine receptor signaling defects in spontaneous hypertension. Am J Hypertens. 1996;9:400–405. doi: 10.1016/0895-7061(95)00351-7. [DOI] [PubMed] [Google Scholar]
- 114.Jose PA, Eisner GM, Felder RA. Regulation of blood pressure by dopamine receptors. Nephron Physiol. 2003;95:19–27. doi: 10.1159/000073676. [DOI] [PubMed] [Google Scholar]
- 115.Joseph JD, Wang YM, Miles PR, Budygin EA, Picetti R, Gainetdinov RR, Caron MG, Wightman RM. Dopamine autoreceptor regulation of release and uptake in mouse brain slices in the absence of D3 receptors. Neuroscience. 2002;112:39–49. doi: 10.1016/s0306-4522(02)00067-2. [DOI] [PubMed] [Google Scholar]
- 116.Jung MY, Schmauss C. Decreased c-fos responses to dopamine D1 receptor agonist stimulation in mice deficient for D3 receptors. J Biol Chem. 1999;274:29406–29412. doi: 10.1074/jbc.274.41.29406. [DOI] [PubMed] [Google Scholar]
- 117.Kanterman RY, Mahan LC, Briley EM, Monsma FJ, Jr, Sibley DR, Axelrod J, Felder CC. Transfected D2 dopamine receptors mediate the potentiation of arachidonic acid release in Chinese hamster ovary cells. Mol Pharmacol. 1991;39:364–369. [PubMed] [Google Scholar]
- 118.Khundmiri SJ, Lederer E. PTH and DA regulate Na-K ATPase through divergent pathways. Am J Physiol Renal Physiol. 2002;282:F512–F522. doi: 10.1152/ajprenal.00111.2000. [DOI] [PubMed] [Google Scholar]
- 119.Kim MO, Koh PO, Kim JH, Kim JS, Kang SS, Cho GJ, Kim K, Choi WS. Localization of dopamine D1 and D2 receptor mRNAs in the rat systemic and pulmonary vasculatures. Mol Cell. 1999;9:417–421. [PubMed] [Google Scholar]
- 120.Kinoshita S, Sidhu A, Felder RA. Defective dopamine-1 receptor adenylate cyclase coupling in the proximal convoluted tubule from the spontaneously hypertensive rat. J Clin Invest. 1989;84:1849–1856. doi: 10.1172/JCI114371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kirchheimer C, Mendez CF, Acquier A, Nowicki S. Role of 20-HETE in D1/D2 dopamine receptor synergism resulting in the inhibition of Na+-K+-ATPase activity in the proximal tubule. Am J Physiol Renal Physiol. 2007;292:F1435–F1442. doi: 10.1152/ajprenal.00176.2006. [DOI] [PubMed] [Google Scholar]
- 122.Kren V, Pravenec M, Lu S, Krenova D, Wang JM, Wang N, Merriouns T, Wong A, St Lezin E, Lau D, Szpirer C, Szpirer J, Kurtz TW. Genetic isolation of a region of chromosome 8 that exerts major effects on blood pressure and cardiac mass in the spontaneously hypertensive rat. J Clin Invest. 1997;99:577–581. doi: 10.1172/JCI119198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Kruse MS, Adachi S, Scott L, Holtback U, Greengard P, Aperia A, Brismar H. Recruitment of renal dopamine 1 receptors requires an intact microtubulin network. Pflügers Arch. 2003;445:534–539. doi: 10.1007/s00424-002-0899-5. [DOI] [PubMed] [Google Scholar]
- 124.Krushkal J, Xiong M, Ferrell R, Sing CF, Turner ST, Boerwinkle E. Linkage and association of adrenergic and dopamine receptor genes in the distal portion of the long arm of chromosome 5 with systolic blood pressure variation. Hum Mol Genet. 1998;7:1379–1383. doi: 10.1093/hmg/7.9.1379. [DOI] [PubMed] [Google Scholar]
- 125.Kunimi M, Seki G, Hara C, Taniguchi S, Uwatoko S, Goto A, Kimura S, Fujita T. Dopamine inhibits renal cotransporter in rabbits and normotensive rats but not in spontaneously hypertensive rats. Kidney Int. 2000;57:534–543. doi: 10.1046/j.1523-1755.2000.00873.x. [DOI] [PubMed] [Google Scholar]
- 126.Kuzhikandathil EV, Oxford GS. Dominant-negative mutants identify a role for GIRK channels in D3 dopamine receptor-mediated regulation of spontaneous secretory activity. J Gen Physiol. 2000;115:697–706. doi: 10.1085/jgp.115.6.697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Ladines CA, Zeng C, Asico LD, Sun X, Pocchiari F, Semeraro C, Pisegna J, Wank S, Yamaguchi I, Eisner GM, Jose PA. Impaired renal D1-like and D2-like dopamine receptor interaction in the spontaneously hypertensive rat. Am J Physiol Regul Integr Comp Physiol. 2001;281:R1071–R1078. doi: 10.1152/ajpregu.2001.281.4.R1071. [DOI] [PubMed] [Google Scholar]
- 128.Lao YS, Hendley ED, Felder RA, Jose PA. Elevated renal cortical calmodulin- dependent protein kinase activity and blood pressure. Clin Exp Hypertens. 2002;24:289–300. doi: 10.1081/ceh-120004232. [DOI] [PubMed] [Google Scholar]
- 129.Lederer ED, Sohi SS, McLeish KR. Dopamine regulates phosphate uptake by opossum Kidney cells through multiple counter-regulatory receptors. J Am Soc Nephrol. 1998;9:975–985. doi: 10.1681/ASN.V96975. [DOI] [PubMed] [Google Scholar]
- 130.Lee FJ, Pei L, Moszczynska A, Vukusic B, Fletcher PJ, Liu F. Dopamine transporter cell surface localization facilitated by a direct interaction with the dopamine D2 receptor. EMBO J. 2007;26:2127–2136. doi: 10.1038/sj.emboj.7601656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Lee FJ, Xue S, Pei L, Vukusic B, Chery N, Wang Y, Wang YT, Niznik HB, Yu XM, Liu F. Dual regulation of NMDA receptor functions by direct protein-protein interactions with the dopamine D1 receptor. Cell. 2002;111:219–230. doi: 10.1016/s0092-8674(02)00962-5. [DOI] [PubMed] [Google Scholar]
- 132.Lee SP, So CH, Rashid AJ, Varghese GV, Cheng RC, Lanca AJ, O'Dowd BF, George SR. Dopamine D1 and D2 receptor co-activation generates a novel phospholipase C-mediated calcium signal. J Biol Chem. 2004;279:35671–35678. doi: 10.1074/jbc.M401923200. [DOI] [PubMed] [Google Scholar]
- 133.Levavi-Sivan B, Park BH, Fuchs S, Fishburn CS. Human D3 dopamine receptor in the medulloblastoma TE671 cell line: cross-talk between D1 and D3 receptors. FEBS Lett. 1998;439:138–142. doi: 10.1016/s0014-5793(98)01356-8. [DOI] [PubMed] [Google Scholar]
- 134.Levesque D. Aminotetralin drugs and D3 receptor functions. What may partially selective D3 receptor ligands tell us about dopamine D3 receptor functions? Biochem Pharmacol. 1996;52:511–518. doi: 10.1016/0006-2952(96)00239-0. [DOI] [PubMed] [Google Scholar]
- 135.Li D, Aperia A, Celsi G, da Cruz e Silva EF, Greengard P, Meister B. Protein phosphatase-1 in the Kidney: evidence for a role in the regulation of medullary Na+-K+-ATPase. Am J Physiol Renal Fluid Electrolyte Physiol. 1995;269:F673–F680. doi: 10.1152/ajprenal.1995.269.5.F673. [DOI] [PubMed] [Google Scholar]
- 136.Li H, Yu P, Jose PA. Role of AT1 receptor ubiquitination in the D5 dopamine receptor-mediated AT1 receptor degradation (Abstract) FASEB J. 2006;20:A1285. [Google Scholar]
- 137.Li L, Schafer JA. Dopamine inhibits vasopressin-dependent cAMP production in the rat cortical collecting duct. Am J Physiol Renal Physiol. 1998;275:F62–F67. doi: 10.1152/ajprenal.1998.275.1.F62. [DOI] [PubMed] [Google Scholar]
- 138.Li XX, Bek M, Asico LD, Yang Z, Grandy DK, Goldstein DS, Rubinstein M, Eisner GM, Jose PA. Adrenergic and endothelin B receptor-dependent hypertension in dopamine receptor type-2 knockout mice. Hypertension. 2001;38:303–308. doi: 10.1161/01.hyp.38.3.303. [DOI] [PubMed] [Google Scholar]
- 139.Lindgren N, Usiello A, Goiny M, Haycock J, Erbs E, Greengard P, Hokfelt T, Borrelli E, Fisone G. Distinct roles of dopamine D2L and D2S receptor isoforms in the regulation of protein phosphorylation at presynaptic and postsynaptic sites. Proc Natl Acad Sci USA. 2003;100:4305–4309. doi: 10.1073/pnas.0730708100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Liu F, Wan Q, Pristupa ZB, Yu XM, Wang YT, Nisnik HB. Direct protein-protein coupling enables cross-talk between dopamine D5 and γ-aminobutyric acid A receptors. Nature. 2000;403:274–280. doi: 10.1038/35002014. [DOI] [PubMed] [Google Scholar]
- 141.Lin JC, Tsao WL, Wang Y. Cardiovascular effects of NMDA in the RVLM of spontaneously hypertensive rats. Brain Res Bull. 1995;37:289–294. doi: 10.1016/0361-9230(95)00014-6. [DOI] [PubMed] [Google Scholar]
- 142.Lotharius J, O'Malley KL. The Parkinsonism-inducing drug 1-methyl-4- phenylpyridinium triggers intracellular dopamine oxidation. A novel mechanism of toxicity. J Biol Chem. 2000;275:38581–38588. doi: 10.1074/jbc.M005385200. [DOI] [PubMed] [Google Scholar]
- 143.Lu Q, Asico LD, Jones JE, Bai R, Li H, Maines MD, Jose PA. Impaired heme oxygenase activity, increased reactive oxygen species, and high blood pressure in D5 dopamine receptor deficient mice (Abstract) J Am Soc Nephrol. 2005;16:163A. [Google Scholar]
- 144.Luippold G, Kuster E, Joos TO, Muhlbauer B. Dopamine D3 receptor activation modulates renal function in anesthetized rats. Naunyn Schmiedebergs Arch Pharmacol. 1998;358:690–693. doi: 10.1007/pl00005314. [DOI] [PubMed] [Google Scholar]
- 145.Luippold G, Piesch C, Osswald H, Muhlbauer B. Dopamine D3 receptor mRNA and renal response to D3 receptor activation in spontaneously hypertensive rats. Hypertens Res. 2003;26:855–861. doi: 10.1291/hypres.26.855. [DOI] [PubMed] [Google Scholar]
- 146.Luippold G, Zimmermann C, Mai M, Kloor D, Starck D, Gross G, Muhlbauer B. Dopamine D3 receptors and salt-dependent hypertension. J Am Soc Nephrol. 2001;12:2272–2279. doi: 10.1681/ASN.V12112272. [DOI] [PubMed] [Google Scholar]
- 147.Luo Y, Roth GS. The roles of dopamine oxidative stress and dopamine receptor signaling in aging and age-related neurodegeneration. Antioxid Redox Signal. 2000;2:449–460. doi: 10.1089/15230860050192224. [DOI] [PubMed] [Google Scholar]
- 148.Maeda Y, Terada Y, Nonoguchi H, Knepper MA. Hormone and autacoid regulation of cAMP production in rat IMCD subsegments. Am J Physiol Renal Fluid Electrolyte Physiol. 1992;263:F319–F327. doi: 10.1152/ajprenal.1992.263.2.F319. [DOI] [PubMed] [Google Scholar]
- 149.Maggio R, Scarselli M, Novi F, Millan MJ, Corsini GU. Potent activation of dopamine D3/D2 heterodimers by the antiparkinsonian agents, S32504, pramipexole and ropinirole. J Neurochem. 2003;87:631–641. doi: 10.1046/j.1471-4159.2003.02038.x. [DOI] [PubMed] [Google Scholar]
- 150.Maiorov DN, Krenz NR, Krassioukov AV, Weaver LC. Role of spinal NMDA and AMPA receptors in episodic hypertension in conscious spinal rats. Am J Physiol Heart Circ Physiol. 1997;273:H1266–H1274. doi: 10.1152/ajpheart.1997.273.3.H1266. [DOI] [PubMed] [Google Scholar]
- 151.Matsumoto M, Hidaka K, Tada S, Tasaki Y, Yamaguchi T. Full-length cDNA cloning and distribution of human dopamine D4 receptor. Mol Brain Res. 1995;29:157–162. doi: 10.1016/0169-328x(94)00245-a. [DOI] [PubMed] [Google Scholar]
- 152.Matsumoto T, Ozono R, Sasaki N, Oshima T, Matsuura H, Kajiyama G, Carey RM, Kambe M. Type 1A dopamine receptor expression in the heart is not altered in spontaneously hypertensive rats. Am J Hypertens. 2000;13:673–677. doi: 10.1016/s0895-7061(99)00270-8. [DOI] [PubMed] [Google Scholar]
- 153.Michel MC, Siepmann F, Buscher R, Philipp T, Brodde OE. Ontogenesis of sympathetic responsiveness in spontaneously hypertensive rats. I. Renal alpha 1-, alpha 2-, and beta-adrenergic receptors and their signaling. Hypertension. 1993;22:169–177. doi: 10.1161/01.hyp.22.2.169. [DOI] [PubMed] [Google Scholar]
- 154.Monsma FJ, Jr, McVittie LD, Gerfen CR, Mahan LC, Sibley DR. Multiple D2 dopamine receptors produced by alternative RNA splicing. Nature. 1989;342:926–929. doi: 10.1038/342926a0. [DOI] [PubMed] [Google Scholar]
- 155.Muhlbauer B, Kuster E, Luippold G. Dopamine D3 receptors in the rat Kidney: role in physiology and pathophysiology. Acta Physiol Scand. 2000;168:219–223. doi: 10.1046/j.1365-201x.2000.00665.x. [DOI] [PubMed] [Google Scholar]
- 156.Murphy MB, Murray C, Shorten GD. Fenoldopam: a selective peripheral dopamine-receptor agonist for the treatment of severe hypertension. N Engl J Med. 2001;345:1548–1557. doi: 10.1056/NEJMra010253. [DOI] [PubMed] [Google Scholar]
- 157.Nakaki T, Nakayama M, Yamamoto S, Kato R. Alpha 1-adrenergic stimulation and beta 2-adrenergic inhibition of DNA synthesis in vascular smooth muscle cells. Mol Pharmacol. 1989;37:30–36. [PubMed] [Google Scholar]
- 158.Narkar VA, Hussain T, Lokhandwala MF. Activation of D2-like receptors causes recruitment of tyrosine-phosphorylated NKA α1-subunits in Kidney. Am J Physiol Renal Physiol. 2002;283:F1290–F1295. doi: 10.1152/ajprenal.00039.2002. [DOI] [PubMed] [Google Scholar]
- 159.Narkar VA, Hussain T, Pedemonte C, Lokhandwala MF. Dopamine D2 receptor activation causes mitogenesis via p44/42 mitogen-activated protein kinase in opossum Kidney cells. J Am Soc Nephrol. 2001;12:1844–1852. doi: 10.1681/ASN.V1291844. [DOI] [PubMed] [Google Scholar]
- 160.Nash SR, Godinot N, Caron MG. Cloning and characterization of the opossum Kidney cell D1 dopamine receptor: expression of identical D1A and D1B dopamine receptor mRNAs in opossum Kidney and brain. Mol Pharmacol. 1993;44:918–925. [PubMed] [Google Scholar]
- 161.Nasjletti A. The role of eicosanoids in angiotensin-dependent hypertension. Hypertension. 1998;31:194–200. doi: 10.1161/01.hyp.31.1.194. [DOI] [PubMed] [Google Scholar]
- 162.Natarajan AR, Han G, White R, Jose PA. The human D5 dopamine receptor mediates big KCa channel activity in human coronary artery smooth muscle cells (Abstract) Hypertension. 2006;48:e80. [Google Scholar]
- 163.Nielsen CB, Pedersen EB. Abnormal distal tubular sodium reabsorption during dopamine infusion in patients with essential hypertension evaluated by the lithium clearance methods. Clin Nephrol. 1997;47:304–309. [PubMed] [Google Scholar]
- 164.Nishi A, Eklöf AC, Bertorello AM, Aperia A. Dopamine regulation of renal Na+,K+-ATPase activity is lacking in Dahl salt-sensitive rats. Hypertension. 1993;21:767–771. doi: 10.1161/01.hyp.21.6.767. [DOI] [PubMed] [Google Scholar]
- 165.Noble EO, Blum K, Ritchie T, Montgomery A, Sheridan PJ. Allelic association of the D2 dopamine receptor gene with the receptor-binding characteristics in alcoholism. Arch Gen Psychiatry. 1991;48:648–654. doi: 10.1001/archpsyc.1991.01810310066012. [DOI] [PubMed] [Google Scholar]
- 166.Noh JS, Gwag BJ. Attenuation of oxidative neuronal necrosis by a dopamine D1 agonist in mouse cortical cell cultures. Exp Neurol. 1997;146:604–608. doi: 10.1006/exnr.1997.6569. [DOI] [PubMed] [Google Scholar]
- 167.Nowicki S, Kruse MS, Brismar H, Aperia A. Dopamine-induced translocation of protein kinase C isoforms visualized in renal epithelial cells. Am J Physiol Cell Physiol. 2000;279:C1812–C1818. doi: 10.1152/ajpcell.2000.279.6.C1812. [DOI] [PubMed] [Google Scholar]
- 168.Nurnberger A, Rabiger M, Mack A, Diaz J, Sokoloff P, Muhlbauer B, Luippold G. Subapical localization of the dopamine D3 receptor in proximal tubules of the rat Kidney. J Histochem Cytochem. 2004;52:1647–1655. doi: 10.1369/jhc.4A6359.2004. [DOI] [PubMed] [Google Scholar]
- 169.O'Connell DP, Aherne AM, Lane E, Felder RA, Carey RM. Detection of dopamine receptor D1A subtype-specific mRNA in rat Kidney by in situ amplification. Am J Physiol Renal Physiol. 1998;274:F232–F241. doi: 10.1152/ajprenal.1998.274.1.F232. [DOI] [PubMed] [Google Scholar]
- 170.O'Connell DP, Botkin SJ, Ramos SI, Sibley DR, Ariano MA, Felder RA, Carey RM. Localization of dopamine D1A receptor protein in rat Kidneys. Am J Physiol Renal Fluid Electrolyte Physiol. 1995;268:F1185–F1197. doi: 10.1152/ajprenal.1995.268.6.F1185. [DOI] [PubMed] [Google Scholar]
- 171.O'Connell DP, Ragsdale NV, Boyd DG, Felder RA, Carey RM. Differential human renal tubular responses to dopamine type 1 receptor stimulation are determined by blood pressure status. Hypertension. 1997;29:115–122. doi: 10.1161/01.hyp.29.1.115. [DOI] [PubMed] [Google Scholar]
- 172.O'Connell DP, Vaughan CJ, Aherne AM, Botkin SJ, Wang ZQ, Felder RA, Carey RM. Expression of the dopamine D3 receptor protein in the rat Kidney. Hypertension. 1998;32:886–895. doi: 10.1161/01.hyp.32.5.886. [DOI] [PubMed] [Google Scholar]
- 173.Offen D, Ziv I, Sternin H, Melamed E, Hochman A. Prevention of dopamine-induced cell death by thiol antioxidants: possible implications for treatment of Parkinson's disease. Exp Neurol. 1996;141:32–39. doi: 10.1006/exnr.1996.0136. [DOI] [PubMed] [Google Scholar]
- 174.Ohbu K, Felder RA. Nephron specificity of dopamine receptor-adenylyl cyclase defect in spontaneous hypertension. Am J Physiol Renal Fluid Electrolyte Physiol. 1993;264:F274–F279. doi: 10.1152/ajprenal.1993.264.2.F274. [DOI] [PubMed] [Google Scholar]
- 175.Ohbu K, Hendley ED, Yamaguchi I, Felder RA. Renal dopamine-1 receptors in hypertensive inbred rat strains with and without hyperactivity. Hypertension. 1993;21:485–490. doi: 10.1161/01.hyp.21.4.485. [DOI] [PubMed] [Google Scholar]
- 176.Ominato Satoh MT, Katz AI. Regulation of Na-KATPase activity in the proximal tubule: role of the protein kinase C pathway and of eicosanoids. J Membr Biol. 1996;152:235–243. doi: 10.1007/s002329900101. [DOI] [PubMed] [Google Scholar]
- 177.Osborn JW, Jacob F, Guzman P. A neural set point for the long-term control of arterial pressure: beyond the arterial baroreceptor reflex. Am J Physiol Regul Integr Comp Physiol. 2005;288:R846–R855. doi: 10.1152/ajpregu.00474.2004. [DOI] [PubMed] [Google Scholar]
- 178.Ozono R, O'Connell DP, Wang ZQ, Moore AF, Sanada H, Felder RA, Carey RM. Localization of the dopamine D1 receptor protein in the human heart and Kidney. Hypertension. 1997;30:725–729. doi: 10.1161/01.hyp.30.3.725. [DOI] [PubMed] [Google Scholar]
- 179.Ozono R, Ueda A, Oishi Y, Yano A, Kambe M, Katsuki M, Oshima T. Dopamine D2 receptor modulates sodium handling via local production of dopamine in the Kidney. J Cardiovasc Pharmacol. 2003;42:S75–S79. doi: 10.1097/00005344-200312001-00017. [DOI] [PubMed] [Google Scholar]
- 180.Pedrosa R, Jose PA, Soares-Da-Silva P. Defective D1-like receptor-mediated inhibition of exchanger in immortalized SHR proximal tubular epithelial cells. Am J Physiol Renal Physiol. 2004;286:F1120–F1126. doi: 10.1152/ajprenal.00433.2003. [DOI] [PubMed] [Google Scholar]
- 181.Pei L, Lee FJ, Moszczynska A, Vukusic B, Liu F. Regulation of dopamine D1 receptor function by physical interaction with the NMDA receptors. J Neurosci. 2004;24:1149–1158. doi: 10.1523/JNEUROSCI.3922-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Pettersson-Fernholm KJ, Forsblom CM, Perola M, Fagerudd JA, Groop PH FinnDiane Study Group. Dopamine D3 receptor gene polymorphisms, blood pressure and nephropathy in type 1 diabetic patients. Nephrol Dial Transplant. 2004;19:1432–1436. doi: 10.1093/ndt/gfh174. [DOI] [PubMed] [Google Scholar]
- 183.Piomelli D, Pilon C, Giros B, Sokoloff P, Martres MP, Schwartz JC. Dopamine activation of the arachidonic acid cascade as a basis for D1/D2 receptor synergism. Nature. 1991;353:164–167. doi: 10.1038/353164a0. [DOI] [PubMed] [Google Scholar]
- 184.Polakowski JS, Segreti JA, Cox BF, Hsieh GC, Kolasa T, Moreland RB, Brioni JD. Effects of selective dopamine receptor subtype agonists on cardiac contractility and regional haemodynamics in rats. Clin Exp Pharmacol Physiol. 2004;31:837–841. doi: 10.1111/j.1440-1681.2004.04095.x. [DOI] [PubMed] [Google Scholar]
- 185.Pollack A. Coactivation of D1and D2 dopamine receptors: in marriage, a case of his, hers, and theirs. Sci STKE. 2004:pe50. doi: 10.1126/stke.2552004pe50. [DOI] [PubMed] [Google Scholar]
- 186.Premont RT, Gainetdinov RR. Physiological roles of G protein-coupled receptor kinases and arrestins. Annu Rev Physiol. 2007;69:511–534. doi: 10.1146/annurev.physiol.69.022405.154731. [DOI] [PubMed] [Google Scholar]
- 187.Prinster SC, Hague C, Hall RA. Heterodimerization of G-protein-coupled receptors: specificity and functional significance. Pharmacol Rev. 2005;57:289–298. doi: 10.1124/pr.57.3.1. [DOI] [PubMed] [Google Scholar]
- 188.Ragsdale NV, Lynch M, Chevalier RL, Felder RA, Peach MJ, Carey RM. Selective peripheral dopamine-1 receptor stimulation: differential responses to sodium loading and depletion in humans. Hypertension. 1990;15:914–921. doi: 10.1161/01.hyp.15.6.914. [DOI] [PubMed] [Google Scholar]
- 189.Raizada MK, Der Sarkissian S. Potential of gene therapy strategy for the treatment of hypertension. Hypertension. 2006;47:6–9. doi: 10.1161/01.HYP.0000196685.91424.01. [DOI] [PubMed] [Google Scholar]
- 190.Rao GN, Berk BC. Active oxygen species stimulate vascular smooth muscle cell growth and proto-oncogene expression. Circ Res. 1992;70:593–599. doi: 10.1161/01.res.70.3.593. [DOI] [PubMed] [Google Scholar]
- 191.Rao F, Wessel J, Wen G, Zhang L, Rana BK, Kennedy BP, Greenwood TA, Salem RM, Chen Y, Khandrika S, Hamilton BA, Smith DW, Holstein-Rathlou NH, Ziegler MG, Schork NJ, O'Connor DT. Renal albumin excretion: twin studies identify influences of heredity, environment, and adrenergic pathway polymorphism. Hypertension. 2007;49:1015–1031. doi: 10.1161/HYPERTENSIONAHA.106.081679. [DOI] [PubMed] [Google Scholar]
- 192.Ricci A, Amenta F, Bronzetti E, Felici L, Hussain T, Lokhandwala MF. Age-related changes of dopamine receptor protein immunoreactivity in the rat mesenteric vascular tree. Mech Ageing Dev. 2002;123:537–546. doi: 10.1016/s0047-6374(01)00361-x. [DOI] [PubMed] [Google Scholar]
- 193.Ricci A, Bronzetti E, Fedele F, Ferrante F, Zaccheo D, Amenta F. Pharmacological characterization and autoradiographic localization of a putative dopamine D4 receptor in the heart. J Auton Pharmacol. 1998;18:115–121. doi: 10.1046/j.1365-2680.1998.1820115.x. [DOI] [PubMed] [Google Scholar]
- 194.Ricci A, Collier WL, Amenta F. Pharmacological characterization and autoradiographic localization of dopamine receptors in the portal vein. J Auton Pharmacol. 1994;14:61–68. doi: 10.1111/j.1474-8673.1994.tb00590.x. [DOI] [PubMed] [Google Scholar]
- 195.Ricci A, Marchal-Victorion S, Bronzetti E, Parini A, Amenta F, Tayebati SK. Dopamine D4 receptor expression in rat Kidney: evidence for pre- and postjunctional localization. Histochem Cytochem. 2002;50:1091–1096. doi: 10.1177/002215540205000811. [DOI] [PubMed] [Google Scholar]
- 196.Ricci A, Mignini F, Tomassoni D, Amenta F. Dopamine receptor subtypes in the human pulmonary arterial tree. Auton Autacoid Pharmacol. 2006;26:361–369. doi: 10.1111/j.1474-8673.2006.00376.x. [DOI] [PubMed] [Google Scholar]
- 197.Rice T, Rankinen T, Province MA, Chagnon YC, Perusse L, Borecki IB, Bouchard C, Rao DC. Genome-wide linkage analysis of systolic and diastolic blood pressure: the Quebec Family Study. Circulation. 2000;102:1956–1963. doi: 10.1161/01.cir.102.16.1956. [DOI] [PubMed] [Google Scholar]
- 198.Ridray S, Griffon N, Mignon V, Souil E, Carboni S, Diaz J, Schwartz JC, Sokoloff P. Coexpression of dopamine D1 and D3 receptors in islands of Calleja and shell of nucleus accumbens of the rat: opposite and synergistic functional interactions. Eur J Neurosci. 1998;10:1676–1686. doi: 10.1046/j.1460-9568.1998.00173.x. [DOI] [PubMed] [Google Scholar]
- 199.Robinson SW, Caron MG. Selective inhibition of adenylyl cyclase type V by the dopamine D3 receptor. Mol Pharmacol. 1997;52:508–514. doi: 10.1124/mol.52.3.508. [DOI] [PubMed] [Google Scholar]
- 200.Rocheville M, Lange DC, Kumar U, Patel SC, Patel RC, Patel YC. Receptors for dopamine and somatostatin: formation of hetero-oligomers with enhanced functional activity. Science. 2000;288:154–157. doi: 10.1126/science.288.5463.154. [DOI] [PubMed] [Google Scholar]
- 201.Rubinstein M, Phillips TJ, Bunzow JR, Falzone TL, Dziewczapolski G, Zhang G, Fang Y, Larson JL, McDougall JA, Chester JA, Saez C, Pugsley TA, Gershanik O, Low MJ, Grandy DK. Mice lacking dopamine D4 receptors are supersensitive to ethanol, cocaine, and methamphetamine. Cell. 1997;90:991–1001. doi: 10.1016/s0092-8674(00)80365-7. [DOI] [PubMed] [Google Scholar]
- 202.Saito O, Ando Y, Kusano E, Asano Y. Functional characterization of basolateral and luminal dopamine receptors in rabbit CCD. Am J Physiol Renal Physiol. 2001;281:F114–F122. doi: 10.1152/ajprenal.2001.281.1.F114. [DOI] [PubMed] [Google Scholar]
- 203.Salomone LJ, Howell NL, McGrath HE, Kemp BA, Keller SR, Gildea JJ, Felder RA, Carey RM. Intrarenal dopamine D1-like receptor stimulation induces natriuresis via an angiotensin type-2 receptor mechanism. Hypertension. 2007;49:155–161. doi: 10.1161/01.HYP.0000251881.89610.ee. [DOI] [PubMed] [Google Scholar]
- 204.Sam EE, Verbeke N. Free radical scavenging properties of apomorphine enantiomers and dopamine: possible implication in their mechanism of action in Parkinsonism. J Neural Transm. 1995;10:115–127. doi: 10.1007/BF02251227. [DOI] [PubMed] [Google Scholar]
- 205.Sanada H, Jose PA, Hazen-Martin D, Yu PY, Xu J, Bruns DE, Phipps J, Carey RM, Felder RA. Dopamine-1 receptor coupling defect in renal proximal tubule cells in hypertension. Hypertension. 1999;33:1036–1042. doi: 10.1161/01.hyp.33.4.1036. [DOI] [PubMed] [Google Scholar]
- 206.Sanada H, Xu J, Watanabe H, Jose PA, Felder RA. Differential expression and regulation of dopamine-1 (D-1) and dopamine-5 (D-5) receptor function in human Kidney (Abstract) Am J Hypertens. 2000;13:156A. [Google Scholar]
- 207.Sanada H, Yao L, Jose PA, Carey RM, Felder RA. Dopamine D3 receptors in rat juxtaglomerular cells. Clin Exp Hypertens. 1997;19:93–105. doi: 10.3109/10641969709080807. [DOI] [PubMed] [Google Scholar]
- 208.Sanada H, Yatabe J, Midorikawa S, Katoh T, Hashimoto S, Watanabe T, Xu J, Luo Y, Wang X, Zeng C, Armando I, Felder RA, Jose PA. Amelioration of genetic hypertension by suppression of renal G protein-coupled receptor kinase type 4 expression. Hypertension. 2006;47:1131–1139. doi: 10.1161/01.HYP.0000222004.74872.17. [DOI] [PubMed] [Google Scholar]
- 209.Sarkis A, Lopez B, Roman RJ. Role of 20-hydroxyeicosatetraenoic acid and epoxy-eicosatrienoic acids in hypertension. Curr Opin Nephrol Hypertens. 2004;13:205–214. doi: 10.1097/00041552-200403000-00009. [DOI] [PubMed] [Google Scholar]
- 210.Sato M, Soma M, Nakayama T, Kanmatsuse K. Dopamine D1 receptor gene polymorphism is associated with essential hypertension. Hypertension. 2000;36:183–186. doi: 10.1161/01.hyp.36.2.183. [DOI] [PubMed] [Google Scholar]
- 211.Satoh T, Cohen HT, Katz AI. Intracellular signaling in the regulation of renal Na-K-ATPase. I. Role of cyclic AMP and phospholipase A2. J Clin Invest. 1992;89:1496–1500. doi: 10.1172/JCI115740. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212.Satoh T, Cohen HT, Katz AI. Different mechanisms of renal Na-K-ATPase regulation by protein kinases in proximal and distal nephron. Am J Physiol Renal Fluid Electrolyte Physiol. 1993;265:F399–F405. doi: 10.1152/ajprenal.1993.265.3.F399. [DOI] [PubMed] [Google Scholar]
- 213.Sautel F, Griffon N, Levesque D, Pilon C, Schwartz JC, Sokoloff P. A functional test identifies dopamine agonists selective for D3 versus D2 receptors. Neuroreport. 1995;6:329–332. doi: 10.1097/00001756-199501000-00026. [DOI] [PubMed] [Google Scholar]
- 214.Scarselli M, Novi F, Schallmach E, Lin R, Baragli A, Colzi A, Griffon N, Corsini GU, Sokoloff P, Levenson R, Vogel Z, Maggio R. D2/D3 dopamine receptor heterodimers exhibit unique functional properties. J Biol Chem. 2001;276:30308–30314. doi: 10.1074/jbc.M102297200. [DOI] [PubMed] [Google Scholar]
- 215.Schioth HB, Fredriksson R. The GRAFS classification system of G-protein coupled receptors in comparative perspective. Gen Comp Endocrinol. 2005;142:94–101. doi: 10.1016/j.ygcen.2004.12.018. [DOI] [PubMed] [Google Scholar]
- 216.Schmauss C. A single dose of methamphetamine leads to a long term reversal of the blunted dopamine D1 receptor-mediated neocortical c-fos responses in mice deficient for D2 and D3 receptors. J Biol Chem. 2000;275:38944–38948. doi: 10.1074/jbc.M005064200. [DOI] [PubMed] [Google Scholar]
- 217.Schmitz Y, Schmauss C, Sulzer D. Altered dopamine release and uptake kinetics in mice lacking D2 receptors. J Neurosci. 2002;22:8002–8009. doi: 10.1523/JNEUROSCI.22-18-08002.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Scott L, Kruse MS, Forssberg H, Brismar H, Greengard P, Aperia A. Selective up-regulation of dopamine D1 receptors in dendritic spines by NMDA receptor activation. Proc Natl Acad Sci USA. 2002;99:1661–1664. doi: 10.1073/pnas.032654599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Sen S, Nesse R, Sheng L, Stoltenberg SF, Gleiberman L, Burmeister M, Weder AB. Association between a dopamine-4 receptor polymorphism and blood pressure. Am J Hypertens. 2005;18:1206–1210. doi: 10.1016/j.amjhyper.2005.04.010. [DOI] [PubMed] [Google Scholar]
- 220.Sheikh-Hamad D, Wang YP, Jo OD, Yanagawa N. Dopamine antagonizes the actions of angiotensin II in renal brush-border membrane. Am J Physiol Renal Fluid Electrolyte Physiol. 1993;264:F737–F743. doi: 10.1152/ajprenal.1993.264.4.F737. [DOI] [PubMed] [Google Scholar]
- 221.Shin Y, Kumar U, Patel Y, Patel SC, Sidhu A. Differential expression of D2-like dopamine receptors in the Kidney of the spontaneously hypertensive rat. J Hypertens. 2003;21:199–207. doi: 10.1097/00004872-200301000-00030. [DOI] [PubMed] [Google Scholar]
- 222.Shultz PJ, Sedor JR, Abboud HE. Dopaminergic stimulation of cAMP accumulation in cultured rat mesangial cells. Am J Physiol Heart Circ Physiol. 1987;253:H358–H364. doi: 10.1152/ajpheart.1987.253.2.H358. [DOI] [PubMed] [Google Scholar]
- 223.Sibley DR. New insights into dopaminergic receptor function using antisense and genetically altered animals. Annu Rev Pharmacol Toxicol. 1999;39:313–341. doi: 10.1146/annurev.pharmtox.39.1.313. [DOI] [PubMed] [Google Scholar]
- 224.Soma M, Nakayama K, Rahmutula D, Uwabo J, Sato M, Kunimoto M, Aoi N, Kosuge K, Kanmatsuse K. Ser9Gly polymorphism in the dopamine D3 receptor gene is not associated with essential hypertension in the Japanese. Med Sci Monit. 2002;8:CR1–CR4. [PubMed] [Google Scholar]
- 225.Staudacher T, Pech B, Tappe M, Gross G, Muhlbauer B, Luippold G. Arterial blood pressure and renal sodium excretion in dopamine D3 receptor knockout mice. Hypertens Res. 2007;30:93–101. doi: 10.1291/hypres.30.93. [DOI] [PubMed] [Google Scholar]
- 226.Sugiyama F, Churchill GA, Higgins DC, Johns C, Makaritsis KP, Gavras H, Paigen B. Concordance of murine quantitative trait loci for salt-induced hypertension with rat and human loci. Genomics. 2001;71:70–77. doi: 10.1006/geno.2000.6401. [DOI] [PubMed] [Google Scholar]
- 227.Sun D, Wilborn TW, Schafer JA. Dopamine D4 receptor isoform mRNA and protein are expressed in the rat cortical collecting duct. Am J Physiol Renal Physiol. 1998;275:F742–F751. doi: 10.1152/ajprenal.1998.275.5.F742. [DOI] [PubMed] [Google Scholar]
- 228.Sun MH, Ishine T, Lee TJ. Dopamine constricts porcine pial veins. Eur J Pharmacol. 1997;334:165–167. doi: 10.1016/s0014-2999(97)01166-7. [DOI] [PubMed] [Google Scholar]
- 229.Sunahara RK, Guan HC, O'Dowd BF, Seeman P, Laurier LG, Ng G, George SR, Torchia J, Van Tol HH, Niznik HB. Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D1. Nature. 1991;350:614–619. doi: 10.1038/350614a0. [DOI] [PubMed] [Google Scholar]
- 230.Takemoto F, Satoh T, Cohen HT, Katz AI. Localization of dopamine-1 receptors along the microdissected rat nephron. Pflügers Arch. 1991;419:243–248. doi: 10.1007/BF00371102. [DOI] [PubMed] [Google Scholar]
- 231.Teirstein PS, Price MJ, Mathur VS, Madyoon H, Sawhney N, Baim DS. Differential effects between intravenous and targeted renal delivery of fenoldopam on renal function and blood pressure in patients undergoing cardiac catheterization. Am J Cardiol. 2006;97:1076–1081. doi: 10.1016/j.amjcard.2005.10.053. [DOI] [PubMed] [Google Scholar]
- 232.Thomas GN, Critchley JA, Tomlinson B, Cockram CS, Chan JC. Relationships between the taqI polymorphism of the dopamine D2 receptor and blood pressure in hyperglycaemic and normoglycaemic Chinese subjects. Clin Endocrinol (Oxf) 2001;55:605–611. doi: 10.1046/j.1365-2265.2001.01404.x. [DOI] [PubMed] [Google Scholar]
- 233.Thomas GN, Tomlinson B, Critchley JA. Modulation of blood pressure and obesity with the dopamine D2 receptor gene TaqI polymorphism. Hypertension. 2000;36:177–182. doi: 10.1161/01.hyp.36.2.177. [DOI] [PubMed] [Google Scholar]
- 234.Touyz RM. Intracellular mechanisms involved in vascular remodelling of resistance arteries in hypertension: role of angiotensin II. Exp Physiol. 2005;90:449–455. doi: 10.1113/expphysiol.2005.030080. [DOI] [PubMed] [Google Scholar]
- 235.Ueda A, Ozono R, Oshima T, Yano A, Kambe M, Teranishi Y, Katsuki M, Chayama K. Disruption of the type 2 dopamine receptor gene causes a sodium-dependent increase in blood pressure in mice. Am J Hypertens. 2003;16:853–858. doi: 10.1016/s0895-7061(03)01013-6. [DOI] [PubMed] [Google Scholar]
- 236.Van Tol HHM, Wu CM, Guan HC, Ohara K, Bunzow JR, Civelli O, Kennedy J, Seeman P, Niznik HB, Jovanovic V. Multiple dopamine D4 receptor variants in the human population. Nature. 1992;358:149–152. doi: 10.1038/358149a0. [DOI] [PubMed] [Google Scholar]
- 237.Wang H, Joseph JA. Quantifying cellular oxidative stress by dichlorofluorescein assay using microplate reader. Free Radic Biol Med. 1999;27:612–616. doi: 10.1016/s0891-5849(99)00107-0. [DOI] [PubMed] [Google Scholar]
- 238.Wang X, Armando I, Asico LD, Jones JE, Escano CS, Jose PA. Hypertension in dopamine receptor D3 deficient mice is associated with increased Na transporters in Kidney (Abstract) J Am Soc Nephrol. 2005;16:350A. [Google Scholar]
- 239.Wang ZQ, Felder RA, Carey RM. Selective inhibition of the renal dopamine subtype D1A receptor induces antinatriuresis in conscious rats. Hypertension. 1999;33:504–510. doi: 10.1161/01.hyp.33.1.504. [DOI] [PubMed] [Google Scholar]
- 240.Wang ZQ, Siragy HM, Felder RA, Carey RM. Intrarenal dopamine production and distribution in the rat. Physiological control of sodium excretion. Hypertension. 1997;29:228–234. doi: 10.1161/01.hyp.29.1.228. [DOI] [PubMed] [Google Scholar]
- 241.Watanabe H, Xu J, Bengra C, Jose PA, Felder RA. Desensitization of human renal D1 dopamine receptors by G protein-coupled receptor kinase 4. Kidney Int. 2002;62:790–798. doi: 10.1046/j.1523-1755.2002.00525.x. [DOI] [PubMed] [Google Scholar]
- 242.Wersinger C, Chen J, Sidhu A. Bimodal induction of dopaminemediated striatal neurotoxicity is mediated through both activation of D1 dopamine receptors and autoxidation. Mol Cell Neurosci. 2004;25:124–137. doi: 10.1016/j.mcn.2003.10.002. [DOI] [PubMed] [Google Scholar]
- 243.White BH, Kimura K, Sidhu A. Inhibition of hormonally induced inositol trisphosphate production in transfected GH4C1 cells: a novel role for the D5 subtype of the dopamine receptor. Neuroendocrinology. 1999;69:209–216. doi: 10.1159/000054421. [DOI] [PubMed] [Google Scholar]
- 244.White RE, Kryman JP, El-Mowafy AM, Han G, Carrier GO. cAMP-dependent vasodilators cross-activate the cGMP-dependent protein kinase to stimulate BKCa channel activity in coronary artery smooth muscle cells. Circ Res. 2000;86:897–905. doi: 10.1161/01.res.86.8.897. [DOI] [PubMed] [Google Scholar]
- 245.Wiederkehr MR, Di Sole F, Collazo R, Quinones H, Fan L, Murer H, Helmle-Kolb C, Moe OW. Characterization of acute inhibition of Na/H exchanger NHE-3 by dopamine in opossum Kidney cells. Kidney Int. 2001;59:197–209. doi: 10.1046/j.1523-1755.2001.00480.x. [DOI] [PubMed] [Google Scholar]
- 246.Xia XG, Schmidt N, Teismann P, Ferger B, Schulz JB. Dopamine mediates striatal malonate toxicity via dopamine transporter-dependent generation of reactive oxygen species and D2 but not D1 receptor activation. J Neurochem. 2001;79:63–70. doi: 10.1046/j.1471-4159.2001.00525.x. [DOI] [PubMed] [Google Scholar]
- 247.Xu J, Li XX, Albrecht FE, Hopfer U, Carey RM, Jose PA. D1 receptor, Gsα, and Na+/H+ exchanger interactions in the Kidney in hypertension. Hypertension. 2000;36:395–399. doi: 10.1161/01.hyp.36.3.395. [DOI] [PubMed] [Google Scholar]
- 248.Yamaguchi I, Jose PA, Mouradian MM, Canessa LM, Monsma FJ, Jr, Sibley DR, Takeyasu K, Felder RA. Expression of dopamine D1A receptor gene in proximal tubule of rat Kidneys. Am J Physiol Renal Fluid Electrolyte Physiol. 1993;264:F280–F285. doi: 10.1152/ajprenal.1993.264.2.F280. [DOI] [PubMed] [Google Scholar]
- 249.Yamaguchi I, Walk SF, Jose PA, Felder RA. Dopamine D2L receptors stimulate Na+/K+ ATPase activity in murine LTK2 cells. Mol Pharmacol. 1996;49:373–378. [PubMed] [Google Scholar]
- 250.Yamaguchi I, Yao L, Sanada H, Ozono R, Mouradian MM, Jose PA, Carey RM, Felder RA. Dopamine D1A receptors and renin release in rat juxtaglomerular cells. Hypertension. 1997;29:962–968. doi: 10.1161/01.hyp.29.4.962. [DOI] [PubMed] [Google Scholar]
- 251.Yamauchi M, Kobayashi Y, Shimoura K, Hattori K, Nakase A. Endothelium-dependent and -independent relaxation by dopamine in the rabbit pulmonary artery. Clin Exp Pharmacol Physiol. 1992;19:401–410. doi: 10.1111/j.1440-1681.1992.tb00482.x. [DOI] [PubMed] [Google Scholar]
- 252.Yang Z, Asico LD, Yu P, Wang Z, Jones JE, Bai RK, Sibley DR, Felder RA, Jose PA. D5 dopamine receptor regulation of phospholipase D. Am J Physiol Heart Circ Physiol. 2005;288:H55–H61. doi: 10.1152/ajpheart.00627.2004. [DOI] [PubMed] [Google Scholar]
- 253.Yang Z, Asico LD, Yu P, Wang Z, Jones JE, Escano CS, Wang X, Quinn MT, Sibley DR, Romero GG, Felder RA, Jose PA. D5 dopamine receptor regulation of reactive oxygen species production, NADPH oxidase, and blood pressure. Am J Physiol Regul Integr Comp Physiol. 2006;290:R96–R104. doi: 10.1152/ajpregu.00434.2005. [DOI] [PubMed] [Google Scholar]
- 254.Yao L, Ruan X, Arendshorst WJ, Jose PA. Dopamine receptor subtype (D1A and D1B) expression in rat renal microVessels (Abstract) Pediatr Res. 1995;37:374A. [Google Scholar]
- 255.Yao LP, Huque E, Baraniuk J, Carey RM, Felder RA, Jose PA. Dopamine receptor subtype expression (D1A and D1B) in rat nephron segments (Abstract) J Investig Med. 1996;44:305A. [Google Scholar]
- 256.Yao LP, Li XX, Yu PY, Xu J, Asico LD, Jose PA. Dopamine D1 receptor and protein kinase C isoforms in spontaneously hypertensive rats. Hypertension. 1998;32:1049–1053. doi: 10.1161/01.hyp.32.6.1049. [DOI] [PubMed] [Google Scholar]
- 257.Yasunari K, Kohno M, Hasuma T, Horio T, Kano H, Yokokawa K, Minami M, Yoshikawa J. Dopamine as a novel antimigration and antiproliferative factor of vascular smooth muscle cells through dopamine D1-like receptors. Arterioscler Thromb Vasc Biol. 1997;17:3164–3173. doi: 10.1161/01.atv.17.11.3164. [DOI] [PubMed] [Google Scholar]
- 258.Yasunari K, Kohno M, Kano H, Minami M, Yoshikawa J. Dopamine as a novel antioxidative agent for rat vascular smooth muscle cells through dopamine D1-like receptors. Circulation. 2000;101:2302–2308. doi: 10.1161/01.cir.101.19.2302. [DOI] [PubMed] [Google Scholar]
- 259.Yu P, Asico LD, Luo Y, Andrews P, Eisner GM, Hopfer U, Felder RA, Jose PA. D1 dopamine receptor hyperphosphorylation in renal proximal tubules in hypertension. Kidney Int. 2006;70:1072–1079. doi: 10.1038/sj.ki.5001708. [DOI] [PubMed] [Google Scholar]
- 260.Yu P, Yang Z, Jones JE, Wang Z, Owens SA, Mueller SC, Felder RA, Jose PA. D1 dopamine receptor signaling involves caveolin-2 in HEK-293 cells. Kidney Int. 2004;66:2167–2180. doi: 10.1111/j.1523-1755.2004.66007.x. [DOI] [PubMed] [Google Scholar]
- 261.Zapata A, Shippenberg TS. Lack of functional D2 receptors prevents the effects of the D3-preferring agonist (+)-PD 128907 on dialysate dopamine levels. Neuropharmacology. 2005;48:43–50. doi: 10.1016/j.neuropharm.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 262.Zarei S, Frieden M, Rubi B, Villemin P, Gauthier BR, Maechler P, Vischer UM. Dopamine modulates von Willebrand factor secretion in endothelial cells via D2-D4 receptors. J Thromb Haemost. 2006;4:1588–1595. doi: 10.1111/j.1538-7836.2006.01998.x. [DOI] [PubMed] [Google Scholar]
- 263.Zeng C, Asico LD, Zheng S, Hopfer U, Eisner GM, Felder RA, Jose PA. Role of Gα12- and Gα13-protein subunit linkage of D3 dopamine receptors in the impaired natriuretic effect of D3 dopamine receptors in SHRs (Abstract) Am J Hypertens. 2004;17:96A. [Google Scholar]
- 264.Zeng C, Asico LD, Wang X, Hopfer U, Eisner GM, Felder RA, Jose PA. Angiotensin II regulation of AT1 and D3 dopamine receptors in renal proximal tubule cells of spontaneously hypertensive rats. Hypertension. 2003;41:724–729. doi: 10.1161/01.HYP.0000047880.78462.0E. [DOI] [PubMed] [Google Scholar]
- 265.Zeng C, Felder RA, Jose PA. A new approach for treatment of hypertension: modifying D1 dopamine receptor function. Cardiovasc Hematol Agents Med Chem. 2006;4:369–377. doi: 10.2174/187152506778520727. [DOI] [PubMed] [Google Scholar]
- 266.Zeng C, Liu Y, Wang Z, He D, Huang L, Yu P, Zheng S, Jones JE, Asico LD, Hopfer U, Eisner GM, Felder RA, Jose PA. Activation of D3 dopamine receptor decreases AT1 angiotensin receptor expression in rat renal proximal tubule cells. Circ Res. 2006;99:494–500. doi: 10.1161/01.RES.0000240500.96746.ec. [DOI] [PubMed] [Google Scholar]
- 267.Zeng C, Luo Y, Asico LD, Hopfer U, Eisner GM, Felder RA, Jose PA. Perturbation of D1 dopamine and AT1 receptor interaction in spontaneously hypertensive rats. Hypertension. 2003;42:787–792. doi: 10.1161/01.HYP.0000085334.34963.4E. [DOI] [PubMed] [Google Scholar]
- 268.Zeng C, Sanada H, Watanabe H, Eisner GM, Felder RA, Jose PA. Functional genomics of the dopaminergic system in hypertension. Physiol Genomics. 2004;19:233–246. doi: 10.1152/physiolgenomics.00127.2004. [DOI] [PubMed] [Google Scholar]
- 269.Zeng C, Wang D, Asico LD, Welch WJ, Wilcox CS, Hopfer U, Eisner GM, Felder RA, Jose PA. Aberrant D1 and D3 dopamine receptor transregulation in hypertension. Hypertension. 2004;43:654–660. doi: 10.1161/01.HYP.0000114601.30306.bf. [DOI] [PubMed] [Google Scholar]
- 270.Zeng C, Wang D, Yang Z, Wang Z, Asico LD, Wilcox CS, Eisner GM, Welch WJ, Felder RA, Jose PA. Dopamine D1 receptor augmentation of D3 receptor action in rat aortic or mesenteric vascular smooth muscles. Hypertension. 2004;43:673–679. doi: 10.1161/01.HYP.0000118958.27649.6f. [DOI] [PubMed] [Google Scholar]
- 271.Zeng C, Wang Z, Hopfer U, Asico LD, Eisner GM, Felder RA, Jose PA. Rat strain effects of AT1 receptor activation on D1 dopamine receptors in immortalized renal proximal tubule cells. Hypertension. 2005;46:799–805. doi: 10.1161/01.HYP.0000184251.01159.72. [DOI] [PubMed] [Google Scholar]
- 272.Zeng C, Wang Z, Li H, Yu P, Zheng S, Wu L, Asico LD, Hopfer U, Eisner GM, Felder RA, Jose PA. D3 dopamine receptor directly interacts with D1 dopamine receptor in immortalized renal proximal tubule cells. Hypertension. 2006;47:573–579. doi: 10.1161/01.HYP.0000199983.24674.83. [DOI] [PubMed] [Google Scholar]
- 273.Zeng C, Yang Z, Asico LD, Jose PA. Regulation of blood pressure by D5 dopamine receptors. Cardiovasc Hematol Agents Med Chem. 2007;5:241–248. doi: 10.2174/187152507781058708. [DOI] [PubMed] [Google Scholar]
- 274.Zeng C, Yang Z, Wang Z, Jones J, Wang X, Altea J, Mangrum AJ, Hopfer U, Sibley DR, Eisner GM, Felder RA, Jose PA. Interaction of AT1 and D5 dopamine receptors in renal proximal tubule cells. Hypertension. 2005;45:804–810. doi: 10.1161/01.HYP.0000155212.33212.99. [DOI] [PubMed] [Google Scholar]
- 275.Zeng C, Yu P, Asico LD, Hopfer U, Eisner GM, Jose PA. D3 dopamine receptor upregulates and directly interacts with the D1 dopamine receptor in renal proximal tubule cells (Abstract) Am J Hypertens. 2002;15:11A. doi: 10.1161/01.HYP.0000199983.24674.83. [DOI] [PubMed] [Google Scholar]
- 276.Zeng CY, Yang ZW, Wu LJ, Asico LD, Felder RA, Jose PA. Hypertension in D3 dopamine receptor deficient mice. Zhonghua Xin Xue Guan Bing Za Zhi. 2005;33:1132–1136. [PubMed] [Google Scholar]
- 277.Zhang H, Qiao Z, Zhao Y, Zhao R. Transcription of dopamine D1A receptor mRNAs in rat heart. Methods Find Exp Clin Pharmacol. 1996;18:183–187. [PubMed] [Google Scholar]
- 278.Zhang J, Abdel-Rahman AA. The hypotensive action of rilmenidine is dependent on functional N-methyl-d-aspartate receptor in the rostral ventrolateral medulla of conscious spontaneously hypertensive rats. J Pharmacol Exp Ther. 2002;303:204–210. doi: 10.1124/jpet.102.037333. [DOI] [PubMed] [Google Scholar]
- 279.Zhang L, Lou D, Jiao H, Zhang D, Wang X, Xia Y, Zhang J, Xu M. Cocaine-induced intracellular signaling and gene expression are oppositely regulated by the dopamine D1 and D3 receptors. J Neurosci. 2004;24:3344–3354. doi: 10.1523/JNEUROSCI.0060-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 280.Zheng S, Yu P, Zeng C, Wang Z, Yang Z, Andrews PM, Felder RA, Jose PA. Gα12- and Gα13-protein subunit linkage of D5 dopamine receptors in the nephron. Hypertension. 2003;41:604–610. doi: 10.1161/01.HYP.0000057422.75590.D7. [DOI] [PubMed] [Google Scholar]