

Abstract
Although a correlation exists between obesity and insulin resistance, roughly 25 % of obese individuals are insulin sensitive. AMP-activated protein kinase (AMPK) is a cellular energy sensor that among its many actions, integrates diverse physiological signals to restore energy balance. In addition, in many situations it also increases insulin sensitivity. In this context, AMPK activity is decreased in very obese individuals undergoing bariatric surgery who are insulin resistant compared to equally obese patients who are insulin sensitive. In this review, we will both explore what distinguishes these individuals, and evaluate the evidence that diminished AMPK is associated with insulin resistance and metabolic syndrome-associated disorders in other circumstances.
Keywords: Obesity, AMPK, Insulin resistant, Bariatric surgery, Fetuin-A, Insulin sensitive
Introduction
The ‘obesity epidemic’ is thought to be responsible for the rising prevalence of metabolic syndrome-associated diseases including type 2 diabetes, cardiovascular and nonalcoholic fatty liver disease (NAFLD), and certain forms of cancer [1–3]. Although the relationship between obesity and insulin resistance is a hallmark of the metabolic syndrome, it has been long recognized that some obese individuals (~25 %) are insulin sensitive [4•, 5]. In general, they have less abdominal fat (both visceral and subcutaneous) than their insulin resistant counterparts [4•, 5, 6]. In addition, they are less likely to develop atherosclerotic cardiovascular disease [7••], and presumably other obesity-associated comorbidities although the latter remain to be proven [8].
AMP-activated protein kinase (AMPK) is a conserved eukaryotic protein serine/threonine kinase that senses the energy status of the cell and coordinates a global metabolic response to restore energy homeostasis [9, 10]. It can also be activated by agents that increase intracellular Ca2+ [11, 12]. AMPK appears to be an attractive therapeutic target for metabolic syndrome-associated diseases. For example, popular anti-diabetic drugs such as metformin [13] and thiazolidinediones (TZDs) [14], the endogenous insulin sensitizing adipokine adiponectin [15], and exercise [16] all have been shown both to activate AMPK and be therapeutic for metabolic syndrome-associated diseases. Conversely, decreases in AMPK activity have been associated with an increase in such disorders in experimental animals and more recently, in humans. Table 1 provides a list of physiological and pharmacological regulators of AMPK; and Fig. 1 indicates some of the metabolic and other biological actions of AMPK in mammalian tissues.
Table 1.
Physiologic and pharmacologic regulators of AMPK activity
| Physiological activators | Physiological inhibitors | Pharmacological activators |
|---|---|---|
| Exercise | High glucose | Biguanides (metformin) |
| Caloric restriction | Branch-chain amino acids | Thiazolidinediones (TZDs) |
| SIRT1 | Insulin | Salicylates |
| Adiponectin | TNF-α | Statins |
| GLP-1 | Microbiota | Fenofibrate |
| Leptin | Protein phosphatases | Resveratrol |
| IL-6 | Glucocorticoids | α-Lipoic acid |
| IL-10 | Leptin (CNS) | Berberine |
| Estrogen | Ghrelin (periphery) | Curcumin |
| Catecholamines | Palmitate | |
| Leptin (periphery) | ||
| Ghrelin (CNS) |
Fig. 1.
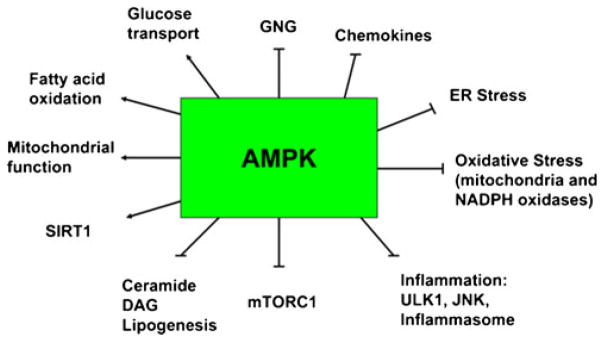
Effects of AMPK activation. In addition to the items listed in the figure, AMPK has been shown to increase eNOS, angiogenesis, autophagy, and the synthesis of anti-inflammatory cytokines such as IL-10. Furthermore, it phosphorylates the FOX Os and almost certainly many other regulatory molecules. Where studied, SIRT1 can produce many of the same effects as AMPK by activating transcriptional activators and co-activators and very likely by other mechanisms. GNG: gluconeogenesis, ULK1: UNC-51 like kinase 1, JNK: JUN activated kinase, DAG: diacylglycerol. The actions of AMPK, as listed above, have been extensively reviewed in [103–105] (Figure adapted from [22]
In this review, we will describe how AMPK becomes dysregulated in obesity, and why such dysregulation is associated with insulin resistance and metabolic syndrome-associated disorders. We will focus predominantly on how these events take place in adipose tissue and liver. In addition, we will discuss the relationship of the pro-inflammatory molecule fetuin-A to insulin resistance, and the involvement of AMPK in fetuin-A regulation. Since data from humans are limited and causality is often difficult to establish, we will refer to rodent studies where applicable.
AMPK: A Multifaceted Molecule with Actions Beyond Energy Balance
An increasing body of work indicates that AMPK is a central regulator of a host of events including inflammation, oxidative/ER stress, autophagy, mitochondrial function and fatty acid oxidation, all of which when dysregulated, could be pathogenic for insulin resistance and ultimately, metabolic syndrome-associated diseases (Fig. 1). Furthermore, it has been shown that AMPK has a prominent role in the central nervous system as well as peripheral tissues. For example, leptin released by adipose tissue decreases both AMPK activity in the hypothalamus and secondarily food intake [17, 18], whereas ghrelin increases both hypothalamic AMPK and food intake [19].
Excess nutrients can down-regulate AMPK activity in peripheral tissues. For example, glucose infused at a high rate in vivo has been shown to diminish AMPK activity and cause insulin resistance in rat muscle and liver [20]; and a high level of glucose or leucine has similar effects on incubated rat muscle [21]. Similarly, rodents with diet- or genetic-induced obesity have unequivocally shown a diminished AMPK activity and insulin resistance in multiple tissues, as well as a predilection to metabolic syndrome-associated diseases [22]. As already noted, studies of human subjects also have revealed a correlation between low AMPK activity in adipose tissue and metabolic disorders associated with insulin resistance and obesity [23••, 24, 25]. However, efforts to find such a correlation in human skeletal muscle have yielded mixed results [16].
AMPK and Adipose Tissue
Rodents and in Vitro Studies
Multiple lines of evidence suggest a link between dysregulation of AMPK activity and oxidative stress, inflammation, and insulin resistance in adipose tissue. They also suggest that exercise (which increases insulin sensitivity) activates AMPK [26]. Conversely, AMPK activity has been reported to be less in the adipose tissue of obese and insulin resistant rodent models [27, 28]. In cultured 3 T3-L1 adipocytes, Gauthier et al. showed that inhibiting AMPK during lipolysis is associated with elevated oxidative stress [29]. The antagonizing effect of AMPK on oxidative stress also has been reported in other tissues [30•, 31]. As for inflammation, genetic deletion of either the α1 or β subunit of AMPK leads to adipose tissue inflammation, insulin resistance, and increased obesity in mice fed a high fat diet [32].
Humans
Decreased AMPK activity attributable to metabolic and hormonal abnormalities in humans was first reported by Kola et al. [24] in visceral adipose tissue of patients with Cushing’s syndrome, a disorder marked by abnormally high plasma cortisol levels [24]. Individuals with Cushing’s syndrome typically present with metabolic abnormalities including insulin resistance and a predisposition to type 2 diabetes, hypertension, and premature coronary artery disease [33]. In a separate study, the same group found that infusion of glucocorticoids into rats led to decreased AMPK activity in adipose and several other tissues [34]. Interestingly, glucocorticoids also have been demonstrated to increase oxidative stress in cultured cells [35].
More recently, studies in severely obese patients undergoing bariatric surgery have revealed an association between diminished AMPK activity, increased visceral adiposity and oxidative stress, and inflammation in multiple adipose tissue depots of insulin resistant individuals compare to a BMI-matched insulin sensitive group [23••, 25]. It is unclear from existing data whether AMPK dysregulation, oxidative stress, or inflammation occurs first. ER stress was not investigated in these studies; however, it should be emphasized that it can down-regulate AMPK activity and also affect both oxidative stress and inflammation [22]. Gregor et al. have reported that ER stress is reduced in adipose and liver of morbidly obese subjects following gastric bypass surgery [36], although they did not divide the study subjects into insulin sensitive and resistant groups. Evaluating fat biopsies post-operatively would be of great interest since it would allow investigators to determine temporally when such pathogenic factors as decreased AMPK activity, oxidative (and ER) stress and inflammation are corrected by the weight loss surgery. In addition, although bariatric surgery is generally associated with a durable remission of type 2 diabetes, about one third of severely obese diabetic patients experience a relapse within 5 years [37, 38]. Such measurements in post-operative biopsies of adipose tissue and in plasma might provide insights why such remissions and relapses occur; and what might be done to prevent the latter. For instance, if evidence of decreased AMPK activity is found, could AMPK-based therapy (i.e., metformin, TZDs, GLP-1 analogs) be useful for patients in relapse? It will also be of interest to follow non-obese offspring of these severely obese individuals to search for early manifestation of the metabolic syndrome (such as insulin resistance, hypertriglycemia [39]) and genetic or other abnormalities in their adipose tissue. Presumably diet and exercise would warrant a trial in such individuals.
AMPK and Insulin Resistance in Liver
Similar to adipose tissue, the α1 subunit is the predominant AMPK isoform in human liver [40], and it is activated by many of the same mechanisms, including exercise [26, 41], starvation [42], IL-6 [43], and adiponectin [44]. Also it is diminished by obesity and nutrient excess [20, 45–47]. In liver, AMPK coordinates fatty acid metabolism [42] by reciprocally regulating triglyceride (TG) synthesis and fatty acid oxidation [48]. The absence or a marked decrease in AMPK activity results in increased hepatic gluconeogenesis, steatosis, and oxidative stress, as well as reductions in mitochondrial biogenesis and fatty acid oxidation [40]. Importantly, hepatic mitochondrial dysfunction (perhaps caused by reduced levels of AMPK) may precede and contribute to insulin resistance and hepatic steatosis, leading to NAFLD [49]. In contrast, activation of AMPK through a variety of means has been shown to restore normoglycemia [50, 51], and lower hepatic glucose production [50–52] and plasma TG levels in both animal models of obesity [53, 54] and type 2 diabetic patients [55]. It remains to be determined whether AMPK activity in liver differs between insulin sensitive and resistant humans. However, based on studies in rodents [50, 56, 57], AMPK activity positively correlates with insulin sensitivity.
Fetuin-A and Insulin Resistance
Similar to adipokine secretion from visceral adipose tissue, the liver can produce hepatokines. The most well-studied pro-inflammatory hepatokine is fetuin-A, a glycoprotein produced primarily by hepatocytes [58], and as reported recently, also by adipocytes [59]. Obese rodents demonstrate elevated fetuin-A mRNA and protein in liver [60–62] and serum [63, 64]. Likewise, elevated fetuin-A levels have been observed in humans with NAFLD and NASH [62, 65, 66]. Mechanistically, fetuin-A is thought to act as an inhibitor of insulin receptor signaling [67], and mice lacking fetuin-A are protected against the development of insulin resistance [68]. Both hyperglycemia [69] and the saturated fatty acid palmitate [63] stimulate fetuin-A production and secretion by hepatocytes (Fig. 2). Excess concentrations of fetuin-A in turn promote inflammatory cytokine production in at least adipose tissue and monocytes [63, 70, 71•] apparently by acting as an endogenous ligand for toll-like receptor 4 (TLR4) that allows palmitate to trigger inflammation and insulin resistance [71•] (Fig. 2). Work from our group has shown that fetuin-A also can induce inflammation in cultured endothelial cells by a similar mechanism and that AMPK can prevent this (R. Valentine et al., unpublished observations).
Fig. 2.

Metabolic consequences of fetuin-A production. Hepatic production of the hepatokine fetuin-A can be induced by both increased glucose and palmitate. Fetuin-A is released into the circulation and inhibits insulin signaling by binding to the insulin receptor in insulin-responsive tissues, thereby inhibiting tyrosine autophosphorylation and inducing insulin resistance. Fetuin-A also serves as an adaptor protein for saturated fatty acids, allowing them to activate Toll-like receptor 4 (TLR4) and consequently induce inflammatory signaling and insulin resistance. AMPK can act to 1) suppress fetuin-A production and secretion; 2) diminish fetuin-A induced inflammation; and 3) restore insulin signaling inhibited by fetuin-A
Clinical data have linked fetuin-A to obesity, insulin resistance [62, 72, 73], and an increased risk of developing type 2 diabetes [74–76]. In addition, fetuin-A levels have been correlated with cardiovascular disease (CVD) although only in older adults [77–79]. More specifically, it has been associated with carotid artery intima-media thickness and stiffness [80–82], endothelial dysfunction [83], and a risk of myocardial infarction and stroke [84–86]. Interestingly, the link between fetuin-A and CVD appears to be most evident in patients with established type 2 diabetes [79, 87, 88].
AMPK Inhibits Fetuin-A
Currently, clinical trials to lower hepatic and circulating fetuin-A are limited. Weight loss produced by both bariatric surgery [89] and caloric restriction [60, 90] diminishes serum fetuin-A levels, as does short-term exercise independent of weight loss [91]. Likewise, physically active men exhibit lower serum fetuin-A levels than their inactive counterparts [92]. Similar findings have been observed using treatment with metformin [93] or rosiglitazone [94] in small human studies. Interestingly, all of these treatments can activate AMPK. As already noted, our own work has shown that AMPK can inhibit fetuin-A-induced pro-inflammatory responses in cultured endothelial cells (R. Valentine et al., unpublished data). In summary, accumulating evidence indicate that treatments that activate AMPK, including metformin, caloric restriction, curcumin, adiponectin, and salicylate, diminish fetuin-A in cultured cells and in vivo [61, 64, 93].
The novel relationship between AMPK and fetuin-A provides an intriguing framework by which AMPK may be involved in the prevention/treatment of a host of inflammatory diseases/conditions, including diabetes, metabolic syndrome and fatty liver disease. Very recently, work by Chatterjee et al. [59] revealed an adipocyte source of fetuin-A that can initiate macrophage migration and polarization in the adipose tissue in the setting of obesity. It would be interesting to investigate whether AMPK can diminish fetuin-A generation and action in adipose tissue and whether this in turn helps prevent the adipose tissue inflammation associated with obesity.
AMPK and the Microbiome
The gut microbiota affects host energy expenditure and metabolic function [95], and an altered gut microbiota has been associated with several diseases including obesity and diabetes [96]. Studies in germ-free mice by Backhed et al. revealed that the gut microbiota enhances adiposity mainly by increasing energy extraction from food and by regulating fat storage [97]. The same group subsequently demonstrated that germ-free mice are protected from diet-induced obesity in part due to an increased rate of fatty acid oxidation as a result of increases in AMPK activity in their liver and skeletal muscle [98]. More recently, a study in Caenorhabditis elegans showed that an alteration of microbial metabolism caused by the antidiabetic drug metformin can also have a positive effect on the host’s health [99], suggesting a possible relationship between AMPK and microbial metabolism. Whether AMPK is a key target that modulates the gut microbiota remains to be elucidated. Equally unknown is whether the composition of the gut microbiome differs between insulin sensitive and resistant populations.
The Sirtuins
To date, seven sirtuins have been identified in mammalian cells. Of these, the most studied is SIRT1, a NAD+-dependent protein deacetylase. Like AMPK, SIRT1 plays a pivotal role in mediating a wide variety of events including fuel metabolism, mitochondrial function, senescence, the growth of cancer cells, and possibly longevity. SIRT1 expression and activity are controlled by a regulatory network that functions at several levels including transcriptional, post-transcriptional and post-translational [100]. In addition, an increased NAD+ bioavailability has been shown to be a major regulator of SIRT1 activity [101]. A substantial body of work suggests that SIRT1, like AMPK, is activated by caloric restriction and an increase in energy expenditure [102, 103], and is down-regulated by energy oversupply [104]. Beyond this, SIRT1 has been shown to activate and be activated by AMPK [105, 106], and the two molecules share many downstream targets including but not limited to PGC1α, FOXO1, p65/NFκB [107•] (Fig. 3). For instance, AMPK and SIRT1 jointly act on the master regulator of mitochondrial biogenesis PGC-1α to enhance the synthesis of many mitochondrial proteins [107•]. Evidence that the mitochondrial SIRT3 may interact with AMPK and PGC1α in a similar fashion has been proposed very recently [108].
Fig. 3.

The putative AMPK/SIRT1 cycle. Activation of AMPK by means such as decreased energy state leads to activation of SIRT1 (via increasing NAD+ and/or activity of Nampt). SIRT1 then deacetylates and activates LKB1, which in turn activates AMPK. Conversely, these events could be initiated by factors that act on SIRT1. The joint activation of AMPK and SIRT1 concurrently phosphorylate and deacetylate the listed target molecules and possibly others. The predicted result would be a decreased susceptibility to metabolic syndrome-associated disorders
In the setting of metabolic syndrome, downregulation of SIRT1 in adipose tissue has been shown to increase obesity and macrophage accumulation/inflammation in rodents [109, 110]. Likewise, decreased SIRT1 expression has been observed by several investigators in adipose tissue of obese humans who are insulin resistant [111]. In our own studies, we found decreased SIRT3 and Nampt (a key enzyme involved in the NAD+ biosynthesis) expression in the adipose tissue of insulin resistant obese individuals compared to their insulin sensitive counterpart, although we did not find a decrease in SIRT1 (Xu et al., unpublished data). Finally, in a rodent model with diet-induced obesity and insulin resistance, decreases in AMPK and SIRT1 were observed in liver (compared to control mice) and both of these parameters returned to control level after gastric bypass surgery [112]. Not surprisingly, just like AMPK, SIRT1 is viewed as an extremely attractive target to improve oxidative metabolism and mitochondrial function, and the possibility of jointly using an agent or agents that activate both molecules for treating metabolic syndrome-associated disorders has been entertained [107•]. In this context, it is noteworthy that resveratrol, a pharmacological agent at low concentration activates cellular SIRT1 while at a high dose stimulates AMPK activity in a SIRT1-independent manner in rodents [113].
Conclusion
Since the initial discovery of the role of AMPK in restoring cellular energy balance, there has been an exponential increase in the number of studies examining its effects on various physiological and pathophysiological events. It is now clear that not all obesity is the same, and that at least in white adipose tissue, a lower AMPK activity can distinguish insulin resistant from insulin sensitive obese populations. In light of the ongoing epidemic of obesity and metabolic syndrome-associated diseases, evaluating AMPK for the prevention and therapy of these disorders is certainly worthy of further exploration.
Acknowledgments
Neil B. Ruderman has received grant support and support for travel to meetings for the study or otherwise from NIH.
Footnotes
Compliance with Ethics Guidelines
Human and Animal Rights and Informed Consent This article does not contain any studies with human or animal subjects performed by any of the authors.
Conflict of Interest X. Julia Xu declares that she has no conflict of interest.
Rudy J. Valentine declares that he has no conflict of interest.
Neil B. Ruderman has received grant support from Takeda for studies of GLIP-1 activation of AMPK in endothelial cells.
Contributor Information
X. Julia Xu, Email: juliaxu@bu.edu.
Rudy J. Valentine, Email: rvalenti@bu.edu.
Neil B. Ruderman, Email: nrude@bu.edu.
References
Papers of particular interest, published recently, have been highlighted as:
• Of importance
•• Of major importance
- 1.Grundy SM, Brewer HB, Jr, Cleeman JI, et al. Definition of metabolic syndrome: report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientific issues related to definition. Arterioscler Thromb Vasc Biol. 2004;24:e13–8. doi: 10.1161/01.ATV.0000111245.75752.C6. [DOI] [PubMed] [Google Scholar]
- 2.Morris MJ. Cardiovascular and metabolic effects of obesity. Clin Exp Pharmacol Physiol. 2008;35:416–9. doi: 10.1111/j.1440-1681.2008.04912.x. [DOI] [PubMed] [Google Scholar]
- 3.van Kruijsdijk RC, van der Wall E, Visseren FL. Obesity and cancer: the role of dysfunctional adipose tissue. Cancer Epidemiol Biomarkers Prev. 2009;18:2569–78. doi: 10.1158/1055-9965.EPI-09-0372. [DOI] [PubMed] [Google Scholar]
- 4•.Samocha-Bonet D, Chisholm DJ, Tonks K, et al. Insulin-sensitive obesity in humans - a ‘favorable fat’ phenotype? Trends Endocrinol Metab. 2012;23:116–24. doi: 10.1016/j.tem.2011.12.005. An extensive review that compare cardiovascular disease, type 2 diabetes, and all-cause mortality in obese humans who are insulin sensitive and insulin resistant. [DOI] [PubMed] [Google Scholar]
- 5.Karelis AD. Metabolically healthy but obese individuals. Lancet. 2008;372:1281–3. doi: 10.1016/S0140-6736(08)61531-7. [DOI] [PubMed] [Google Scholar]
- 6.Sims EA. Are there persons who are obese, but metabolically healthy? Metabolism. 2001;50:1499–504. doi: 10.1053/meta.2001.27213. [DOI] [PubMed] [Google Scholar]
- 7••.Sjostrom L, Peltonen M, Jacobson P, et al. Bariatric surgery and long-term cardiovascular events. JAMA. 2012;307:56–65. doi: 10.1001/jama.2011.1914. This paper also showed greater treatment benefits occur in patients with initially higher insulin levels. [DOI] [PubMed] [Google Scholar]
- 8.Fogarty S, Hardie DG. Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim Biophys Acta. 1804;2010:581–91. doi: 10.1016/j.bbapap.2009.09.012. [DOI] [PubMed] [Google Scholar]
- 9.Hardie DG. AMP-activated/SNF1 protein kinases: conserved guardians of cellular energy. Nat Rev Mol Cell Biol. 2007;8:774–85. doi: 10.1038/nrm2249. [DOI] [PubMed] [Google Scholar]
- 10.Hue L, Rider MH. The AMP-activated protein kinase: more than an energy sensor. Essays Biochem. 2007;43:121–37. doi: 10.1042/BSE0430121. [DOI] [PubMed] [Google Scholar]
- 11.Woods A, Dickerson K, Heath R, et al. Ca2+/calmodulin-dependent protein kinase kinase-beta acts upstream of AMP-activated protein kinase in mammalian cells. Cell Metab. 2005;2:21–33. doi: 10.1016/j.cmet.2005.06.005. [DOI] [PubMed] [Google Scholar]
- 12.Hurley RL, Anderson KA, Franzone JM, et al. The Ca2+/calmod-ulin-dependent protein kinase kinases are AMP-activated protein kinase kinases. J Biol Chem. 2005;280:29060–6. doi: 10.1074/jbc.M503824200. [DOI] [PubMed] [Google Scholar]
- 13.Hawley SA, Gadalla AE, Olsen GS, et al. The antidiabetic drug metformin activates the AMP-activated protein kinase cascade via an adenine nucleotide-independent mechanism. Diabetes. 2002;51:2420–5. doi: 10.2337/diabetes.51.8.2420. [DOI] [PubMed] [Google Scholar]
- 14.LeBrasseur NK, Kelly M, Tsao TS, et al. Thiazolidinediones can rapidly activate AMP-activated protein kinase in mammalian tissues. Am J Physiol Endocrinol Metab. 2006;291:E175–81. doi: 10.1152/ajpendo.00453.2005. [DOI] [PubMed] [Google Scholar]
- 15.Zhou L, Deepa SS, Etzler JC, et al. Adiponectin activates AMP-activated protein kinase in muscle cells via APPL1/LKB1-dependent and phospholipase C/Ca2+/Ca2+/calmodulin-dependent protein kinase kinase-dependent pathways. J Biol Chem. 2009;284:22426–35. doi: 10.1074/jbc.M109.028357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Richter EA, Ruderman NB. AMPK and the biochemistry of exercise: implications for human health and disease. Biochem J. 2009;418:261–75. doi: 10.1042/BJ20082055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dagon Y, Hur E, Zheng B, et al. p70S6 kinase phosphorylates AMPK on serine 491 to mediate leptin’s effect on food intake. Cell Metab. 2012;16:104–12. doi: 10.1016/j.cmet.2012.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Minokoshi Y, Alquier T, Furukawa N, et al. AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature. 2004;428:569–74. doi: 10.1038/nature02440. [DOI] [PubMed] [Google Scholar]
- 19.Yang Y, Atasoy D, Su HH, et al. Hunger states switch a flip-flop memory circuit via a synaptic AMPK-dependent positive feedback loop. Cell. 2011;146:992–1003. doi: 10.1016/j.cell.2011.07.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kraegen EW, Saha AK, Preston E, et al. Increased malonyl-CoA and diacylglycerol content and reduced AMPK activity accompany insulin resistance induced by glucose infusion in muscle and liver of rats. Am J Physiol Endocrinol Metab. 2006;290:E471–9. doi: 10.1152/ajpendo.00316.2005. [DOI] [PubMed] [Google Scholar]
- 21.Saha AK, Xu XJ, Lawson E, et al. Downregulation of AMPK accompanies leucine- and glucose-induced increases in protein synthesis and insulin resistance in rat skeletal muscle. Diabetes. 2010;59:2426–34. doi: 10.2337/db09-1870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ruderman NB, Carling D, Prentki M, et al. AMPK, insulin resistance, and the metabolic syndrome. J Clin Invest. 2012;123:2764–72. doi: 10.1172/JCI67227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23••.Xu XJ, Gauthier MS, Hess DT, et al. Insulin sensitive and resistant obesity in humans: AMPK activity, oxidative stress, and depot-specific changes in gene expression in adipose tissue. J Lipid Res. 2012;53:792–801. doi: 10.1194/jlr.P022905. The first study to characterize the differences between subcutaneous abdominal and two types of visceral fat in BMI-matched insulin sensitive and insulin resistant obese subjects. Results indicate that AMPK activity is diminished, and protein carbonylation is increased in all three depots of insulin resistant patients, whereas the expression of inflammation and other genes varied between depots. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kola B, Christ-Crain M, Lolli F, et al. Changes in adenosine 5′-monophosphate-activated protein kinase as a mechanism of visceral obesity in Cushing’s syndrome. J Clin Endocrinol Metab. 2008;93:4969–73. doi: 10.1210/jc.2008-1297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gauthier MS, O’Brien EL, Bigornia S, et al. Decreased AMP-activated protein kinase activity is associated with increased inflammation in visceral adipose tissue and with whole-body insulin resistance in morbidly obese humans. Biochem Biophys Res Commun. 2011;404:382–7. doi: 10.1016/j.bbrc.2010.11.127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Park H, Kaushik VK, Constant S, et al. Coordinate regulation of malonyl-CoA decarboxylase, sn-glycerol-3-phosphate acyltransferase, and acetyl-CoA carboxylase by AMP-activated protein kinase in rat tissues in response to exercise. J Biol Chem. 2002;277:32571–7. doi: 10.1074/jbc.M201692200. [DOI] [PubMed] [Google Scholar]
- 27.Lindholm CR, Ertel RL, Bauwens JD, et al. A high-fat diet decreases AMPK activity in multiple tissues in the absence of hyperglycemia or systemic inflammation in rats. J Physiol Biochem. 2013;69:165–75. doi: 10.1007/s13105-012-0199-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Caton PW, Kieswich J, Yaqoob MM, et al. Metformin opposes impaired AMPK and SIRT1 function and deleterious changes in core clock protein expression in white adipose tissue of genetically-obese db/db mice. Diabetes Obes Metab. 2011;13:1097–104. doi: 10.1111/j.1463-1326.2011.01466.x. [DOI] [PubMed] [Google Scholar]
- 29.Gauthier MS, Miyoshi H, Souza SC, et al. AMP-activated protein kinase is activated as a consequence of lipolysis in the adipocyte: potential mechanism and physiological relevance. J Biol Chem. 2008;283:16514–24. doi: 10.1074/jbc.M708177200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30•.Salminen A, Kaarniranta K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res Rev. 2012;11:230–41. doi: 10.1016/j.arr.2011.12.005. A recent review of various links between AMPK and inflammation and their physiological relevance. [DOI] [PubMed] [Google Scholar]
- 31.Zhang W, Zhang X, Wang H, et al. AMP-activated protein kinase alpha1 protects against diet-induced insulin resistance and obesity. Diabetes. 2012;61:3114–25. doi: 10.2337/db11-1373. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 32.Galic S, Fullerton MD, Schertzer JD, et al. Hematopoietic AMPK beta1 reduces mouse adipose tissue macrophage inflammation and insulin resistance in obesity. J Clin Invest. 2011;121:4903–15. doi: 10.1172/JCI58577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.De Leo M, Pivonello R, Auriemma RS, et al. Cardiovascular disease in Cushing’s syndrome: heart versus vasculature. Neuroendocrinology. 2010;92 (Suppl 1):50–4. doi: 10.1159/000318566.. [DOI] [PubMed] [Google Scholar]
- 34.Christ-Crain M, Kola B, Lolli F, et al. AMP-activated protein kinase mediates glucocorticoid-induced metabolic changes: a novel mechanism in Cushing’s syndrome. FASEB J. 2008;22:1672–83. doi: 10.1096/fj.07-094144. [DOI] [PubMed] [Google Scholar]
- 35.Houstis N, Rosen ED, Lander ES. Reactive oxygen species have a causal role in multiple forms of insulin resistance. Nature. 2006;440:944–8. doi: 10.1038/nature04634. [DOI] [PubMed] [Google Scholar]
- 36.Gregor MF, Yang L, Fabbrini E, et al. Endoplasmic reticulum stress is reduced in tissues of obese subjects after weight loss. Diabetes. 2009;58:693–700. doi: 10.2337/db08-1220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Schauer PR, Kashyap SR, Wolski K, et al. Bariatric surgery versus intensive medical therapy in obese patients with diabetes. N Engl J Med. 2012;366:1567–76. doi: 10.1056/NEJMoa1200225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Arterburn DE, Bogart A, Sherwood NE, et al. A multisite study of long-term remission and relapse of type 2 diabetes mellitus following gastric bypass. Obes Surg. 2013;23:93–102. doi: 10.1007/s11695-012-0802-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ruderman N, Chisholm D, Pi-Sunyer X, et al. The metabolically obese, normal-weight individual revisited. Diabetes. 1998;47:699–713. doi: 10.2337/diabetes.47.5.699. [DOI] [PubMed] [Google Scholar]
- 40.Viollet B, Guigas B, Leclerc J, et al. AMP-activated protein kinase in the regulation of hepatic energy metabolism: from physiology to therapeutic perspectives. Acta Physiol (Oxf) 2009;196(1):81–98. doi: 10.1111/j.1748-1716.2009.01970.x.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Carlson CL, Winder WW. Liver AMP-activated protein kinase and acetyl-CoA carboxylase during and after exercise. J Appl Physiol. 1999;86(2):669–74. doi: 10.1152/jappl.1999.86.2.669. [DOI] [PubMed] [Google Scholar]
- 42.Assifi MM, Suchankova G, Constant S, et al. AMP-activated protein kinase and coordination of hepatic fatty acid metabolism of starved/carbohydrate-refed rats. Am J Physiol Endocrinol Metab. 2005;289(5):E794–800. doi: 10.1152/ajpendo.00144.2005. [DOI] [PubMed] [Google Scholar]
- 43.Kelly M, Keller C, Avilucea PR, et al. AMPK activity is diminished in tissues of IL-6 knockout mice: the effect of exercise. Biochem Biophys Res Commun. 2004;320:449–54. doi: 10.1016/j.bbrc.2004.05.188. [DOI] [PubMed] [Google Scholar]
- 44.Yamauchi T, Kamon J, Minokoshi Y, et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med. 2002;8:1288–95. doi: 10.1038/nm788. [DOI] [PubMed] [Google Scholar]
- 45.Ha SK, Kim J, Chae C. Role of AMP-activated protein kinase and adiponectin during development of hepatic steatosis in high-fat diet-induced obesity in rats. J Comp Pathol. 2011;145(1):88–94. doi: 10.1016/j.jcpa.2010.11.011.. [DOI] [PubMed] [Google Scholar]
- 46.Henriksen BS, Curtis ME, Fillmore N, et al. The effects of chronic AMPK activation on hepatic triglyceride accumulation and glycerol 3-phosphate acyltransferase activity with high fat feeding. Diabetol Metab Syndr. 2013;5(1):29. doi: 10.1186/1758-5996-5-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zang M, Zuccollo A, Hou X, et al. AMP-activated protein kinase is required for the lipid-lowering effect of metformin in insulin-resistant human HepG2 cells. J Biol Chem. 2004;279(46):47898–905. doi: 10.1074/jbc.M408149200. [DOI] [PubMed] [Google Scholar]
- 48.Muoio DM, Seefeld K, Witters LA, et al. AMP-activated kinase reciprocally regulates triacylglycerol synthesis and fatty acid oxidation in liver and muscle: evidence that sn-glycerol-3-phosphate acyltransferase is a novel target. Biochem J. 1999;338(Pt 3):783–91. [PMC free article] [PubMed] [Google Scholar]
- 49.Rector RS, Thyfault JP, Uptergrove GM, et al. Mitochondrial dysfunction precedes insulin resistance and hepatic steatosis and contributes to the natural history of non-alcoholic fatty liver disease in an obese rodent model. J Hepatol. 2010;52(5):727–36. doi: 10.1016/j.jhep.2009.11.030.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Foretz M, Ancellin N, Andreelli F, et al. Short-term overexpression of a constitutively active form of AMP-activated protein kinase in the liver leads to mild hypoglycemia and fatty liver. Diabetes. 2005;54(5):1331–9. doi: 10.2337/diabetes.54.5.1331. [DOI] [PubMed] [Google Scholar]
- 51.Viana AY, Sakoda H, Anai M, et al. Role of hepatic AMPK activation in glucose metabolism and dexamethasone-induced regulation of AMPK expression. Diabetes Res Clin Pract. 2006;73(2):135–42. doi: 10.1016/j.diabres.2005.12.011. [DOI] [PubMed] [Google Scholar]
- 52.Lochhead PA, Salt IP, Walker KS, et al. 5-aminoimidazole-4-carboxamide riboside mimics the effects of insulin on the expression of the 2 key gluconeogenic genes PEPCK and glucose-6-phosphatase. Diabetes. 2000;49:896–903. doi: 10.2337/diabetes.49.6.896. [DOI] [PubMed] [Google Scholar]
- 53.Cool B, Zinker B, Chiou W, et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab. 2006;3(6):403–16. doi: 10.1016/j.cmet.2006.05.005. [DOI] [PubMed] [Google Scholar]
- 54.Yang J, Maika S, Craddock L, et al. Chronic activation of AMP-activated protein kinase-alpha1 in liver leads to decreased adiposity in mice. Biochem Biophys Res Commun. 2008;370(2):248–53. doi: 10.1016/j.bbrc.2008.03.094.. [DOI] [PubMed] [Google Scholar]
- 55.Boon H, Bosselaar M, Praet SF, et al. Intravenous AICAR administration reduces hepatic glucose output and inhibits whole body lipolysis in type 2 diabetic patients. Diabetologia. 2008;51(10):1893–900. doi: 10.1007/s00125-008-1108-7.. [DOI] [PubMed] [Google Scholar]
- 56.Bergeron R, Previs SF, Cline GW, et al. Effect of 5-aminoimidazole-4-carboxamide-1-beta-D-ribofuranoside infusion on in vivo glucose and lipid metabolism in lean and obese Zucker rats. Diabetes. 2001;50:1076–82. doi: 10.2337/diabetes.50.5.1076. [DOI] [PubMed] [Google Scholar]
- 57.Andreelli F, Foretz M, Knauf C, et al. Liver adenosine monophosphate-activated kinase-alpha2 catalytic subunit is a key target for the control of hepatic glucose production by adiponectin and leptin but not insulin. Endocrinology. 2006;147:2432–41. doi: 10.1210/en.2005-0898. [DOI] [PubMed] [Google Scholar]
- 58.Ix JH, Sharma K. Mechanisms linking obesity, chronic kidney disease, and fatty liver disease: the roles of fetuin-A, adiponectin, and AMPK. J Am Soc Nephrol. 2010;21(3):406–12. doi: 10.1681/ASN.2009080820.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Chatterjee P, Seal S, Mukherjee S, et al. Adipocyte fetuin-a contributes to macrophage migration into adipose tissue and polarization of macrophages. J Biol Chem. 2013;288(39):28324–30. doi: 10.1074/jbc.C113.495473.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Choi KM, Han KA, Ahn HJ, et al. The effects of caloric restriction on fetuin-A and cardiovascular risk factors in rats and humans: a randomized controlled trial. Clin Endocrinol (Oxf) 2013;79(3):356–63. doi: 10.1111/cen.12076.. [DOI] [PubMed] [Google Scholar]
- 61.Jung TW, Youn BS, Choi HY, et al. Salsalate and adiponectin ameliorate hepatic steatosis by inhibition of the hepatokine fetuin-A. Biochem Pharmacol. 2013;86(7):960–9. doi: 10.1016/j.bcp.2013.07.034.. [DOI] [PubMed] [Google Scholar]
- 62.Stefan N, Hennige AM, Staiger H, et al. Alpha2-Heremans-Schmid glycoprotein/fetuin-A is associated with insulin resistance and fat accumulation in the liver in humans. Diabetes Care. 2006;29(4):853–7. doi: 10.2337/diacare.29.04.06.dc05-1938. [DOI] [PubMed] [Google Scholar]
- 63.Dasgupta S, Bhattacharya S, Biswas A, et al. NF-kappaB mediates lipid-induced fetuin-A expression in hepatocytes that impairs adipocyte function effecting insulin resistance. Biochem J. 2010;429(3):451–62. doi: 10.1042/BJ20100330.. [DOI] [PubMed] [Google Scholar]
- 64.Oner-Iyidogan Y, Kocak H, Seyidhanoglu M, et al. Curcumin prevents liver fat accumulation and serum fetuin-A increase in rats fed a high-fat diet. J Physiol Biochem. 2013;69(4):677–86. doi: 10.1007/s13105-013-0244-9.. [DOI] [PubMed] [Google Scholar]
- 65.Kahraman A, Sowa JP, Schlattjan M, et al. Fetuin-A mRNA expression is elevated in NASH compared with NAFL patients. Clin Sci (Lond) 2013;125(8):391–400. doi: 10.1042/CS20120542.. [DOI] [PubMed] [Google Scholar]
- 66.Kantartzis K, Machann J, Schick F, et al. The impact of liver fat vs visceral fat in determining categories of prediabetes. Diabetologia. 2010;53(5):882–9. doi: 10.1007/s00125-010-1663-6.. [DOI] [PubMed] [Google Scholar]
- 67.Srinivas PR, Wagner AS, Reddy LV, et al. Serum alpha 2-HS-glycoprotein is an inhibitor of the human insulin receptor at the tyrosine kinase level. Mol Endocrinol. 1993;7(11):1445–55. doi: 10.1210/mend.7.11.7906861. [DOI] [PubMed] [Google Scholar]
- 68.Mathews ST, Singh GP, Ranalletta M, et al. Improved insulin sensitivity and resistance to weight gain in mice null for the Ahsg gene. Diabetes. 2002;51(8):2450–8. doi: 10.2337/diabetes.51.8.2450. [DOI] [PubMed] [Google Scholar]
- 69.Takata H, Ikeda Y, Suehiro T, et al. High glucose induces transactivation of the alpha2-HS glycoprotein gene through the ERK1/2 signaling pathway. J Atheroscler Thromb. 2009;16(4):448–56. doi: 10.5551/jat.no950. [DOI] [PubMed] [Google Scholar]
- 70.Hennige AM, Staiger H, Wicke C, et al. Fetuin-A induces cytokine expression and suppresses adiponectin production. PLoS One. 2008;3(3):e1765. doi: 10.1371/journal.pone.0001765.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71•.Pal D, Dasgupta S, Kundu R, et al. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nat Med. 2012;18(8):1279–85. doi: 10.1038/nm.2851. This article described a novel finding that hepatokine fetuin-A can serve as an endogenous ligand for TLR4 under lipids-induce insulin resistance condition. Fetuin-A thus can be considered a new therapeutic target for insulin resistance and type 2 diabetes. [DOI] [PubMed] [Google Scholar]
- 72.Ix JH, Shlipak MG, Brandenburg VM, et al. Association between human fetuin-A and the metabolic syndrome: data from the Heart and Soul Study. Circulation. 2006;113(14):1760–7. doi: 10.1161/CIRCULATIONAHA.105.588723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Ix JH, Wassel CL, Chertow GM, et al. Fetuin-A and change in body composition in older persons. J Clin Endocrinol Metab. 2009;94(11):4492–8. doi: 10.1210/jc.2009-0916.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ix JH, Wassel CL, Kanaya AM, et al. Fetuin-A and incident diabetes mellitus in older persons. JAMA. 2008;300(2):182–8. doi: 10.1001/jama.300.2.182.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Sun Q, Cornelis MC, Manson JE, et al. Plasma levels of fetuin-A and hepatic enzymes and risk of type 2 diabetes in women in the U.S. Diabetes. 2013;62(1):49–55. doi: 10.2337/db12-0372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Jensen MK, Bartz TM, Djousse L, et al. Genetically elevated fetuin-A levels, fasting glucose levels, and risk of type 2 diabetes: the Cardiovascular Health Study. Diabetes Care. 2013;36(10):3121–7. doi: 10.2337/dc12-2323.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Ix JH, Biggs ML, Mukamal KJ, et al. Association of fetuin-a with incident diabetes mellitus in community-living older adults: the cardiovascular health study. Circulation. 2012;125(19):2316–22. doi: 10.1161/CIRCULATIONAHA.111.072751.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Jensen MK, Bartz TM, Mukamal KJ, et al. Fetuin-A, type 2 diabetes, and risk of cardiovascular disease in older adults: the cardiovascular health study. Diabetes Care. 2013;36(5):1222–8. doi: 10.2337/dc12-1591.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Laughlin GA, Cummins KM, Wassel CL, et al. The association of fetuin-A with cardiovascular disease mortality in older community-dwelling adults: the Rancho Bernardo study. J Am Coll Cardiol. 2012;59(19):1688–96. doi: 10.1016/j.jacc.2012.01.038.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Fiore CE, Celotta G, Politi GG, et al. Association of high alpha2-Heremans-Schmid glycoprotein/fetuin concentration in serum and intima-media thickness in patients with atherosclerotic vascular disease and low bone mass. Atherosclerosis. 2007;195(1):110–5. doi: 10.1016/j.atherosclerosis.2006.08.052. [DOI] [PubMed] [Google Scholar]
- 81.Mori K, Emoto M, Araki T, et al. Association of serum fetuin-A with carotid arterial stiffness. Clin Endocrinol (Oxf) 2007;66(2):246–50. doi: 10.1111/j.1365-2265.2006.02716.x. [DOI] [PubMed] [Google Scholar]
- 82.Rittig K, Thamer C, Haupt A, et al. High plasma fetuin-A is associated with increased carotid intima-media thickness in a middle-aged population. Atherosclerosis. 2009;207(2):341–2. doi: 10.1016/j.atherosclerosis.2009.05.018.. [DOI] [PubMed] [Google Scholar]
- 83.Dogru T, Genc H, Tapan S, et al. Plasma fetuin-A is associated with endothelial dysfunction and subclinical atherosclerosis in subjects with nonalcoholic fatty liver disease. Clin Endocrinol (Oxf) 2013;78(5):712–7. doi: 10.1111/j.1365-2265.2012.04460.x.. [DOI] [PubMed] [Google Scholar]
- 84.Tuttolomondo A, Di Raimondo D, Di Sciacca R, et al. Fetuin-A and CD40 L plasma levels in acute ischemic stroke: differences in relation to TOAST subtype and correlation with clinical and laboratory variables. Atherosclerosis. 2010;208(1):290–6. doi: 10.1016/j.atherosclerosis.2009.07.032.. [DOI] [PubMed] [Google Scholar]
- 85.Voros K, Graf L, Jr, Prohaszka Z, et al. Serum fetuin-A in metabolic and inflammatory pathways in patients with myocardial infarction. Eur J Clin Invest. 2011;41(7):703–9. doi: 10.1111/j.1365-2362.2010.02456.x.. [DOI] [PubMed] [Google Scholar]
- 86.Weikert C, Stefan N, Schulze MB, et al. Plasma fetuin-a levels and the risk of myocardial infarction and ischemic stroke. Circulation. 2008;118(24):2555–62. doi: 10.1161/CIRCULATIONAHA.108.814418.. [DOI] [PubMed] [Google Scholar]
- 87.Jung C-H, Kim B-Y, Kim C-H, et al. Associations of serum fetuin-A levels with insulin resistance and vascular complications in patients with type 2 diabetes. Diabetes Vasc Dis Res. 2013;10:459–67. doi: 10.1177/1479164113490766. [DOI] [PubMed] [Google Scholar]
- 88.Lorant DP, Grujicic M, Hoebaus C, et al. Fetuin-A levels are increased in patients with type 2 diabetes and peripheral arterial disease. Diabetes Care. 2011;34(1):156–61. doi: 10.2337/dc10-0788.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Brix JM, Stingl H, Hollerl F, et al. Elevated Fetuin-A concentrations in morbid obesity decrease after dramatic weight loss. J Clin Endocrinol Metab. 2010;95(11):4877–81. doi: 10.1210/jc.2010-0148.. [DOI] [PubMed] [Google Scholar]
- 90.Reinehr T, Roth CL. Fetuin-A and its relation to metabolic syndrome and fatty liver disease in obese children before and after weight loss. J Clin Endocrinol Metab. 2008;93:4479–85. doi: 10.1210/jc.2008-1505. [DOI] [PubMed] [Google Scholar]
- 91.Malin SK, Mulya A, Fealy CE, et al. Fetuin-A is linked to improved glucose tolerance after short-term exercise training in non-alcoholic fatty liver disease. J Appl Physiol. 2013;115(7):988–94. doi: 10.1152/japplphysiol.00237.2013.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Jenkins NT, McKenzie JA, Hagberg JM, et al. Plasma fetuin-A concentrations in young and older high- and low-active men. Metabolism. 2011;60(2):265–71. doi: 10.1016/j.metabol.2010.01.026.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Haukeland JW, Dahl TB, Yndestad A, et al. Fetuin A in nonalcoholic fatty liver disease: in vivo and in vitro studies. Eur J Endocrinol. 2012;166(3):503–10. doi: 10.1530/EJE-11-0864.. [DOI] [PubMed] [Google Scholar]
- 94.Mori K, Emoto M, Araki T, et al. Effects of pioglitazone on serum fetuin-A levels in patients with type 2 diabetes mellitus. Metabolism. 2008;57(9):1248–52. doi: 10.1016/j.metabol.2008.04.019.. [DOI] [PubMed] [Google Scholar]
- 95.Tilg H, Kaser A. Gut microbiome, obesity, and metabolic dysfunction. J Clin Invest. 2011;121:2126–32. doi: 10.1172/JCI58109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Karlsson F, Tremaroli V, Nielsen J, et al. Assessing the human gut microbiota in metabolic diseases. Diabetes. 2013;62:3341–9. doi: 10.2337/db13-0844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Backhed F, Ding H, Wang T, et al. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A. 2004;101:15718–23. doi: 10.1073/pnas.0407076101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Backhed F, Manchester JK, Semenkovich CF, et al. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci U S A. 2007;104:979–84. doi: 10.1073/pnas.0605374104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Cabreiro F, Au C, Leung KY, et al. Metformin retards aging in C. elegans by altering microbial folate and methionine metabolism. Cell. 2013;153:228–39. doi: 10.1016/j.cell.2013.02.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kwon HS, Ott M. The ups and downs of SIRT1. Trends Biochem Sci. 2008;33:517–25. doi: 10.1016/j.tibs.2008.08.001. [DOI] [PubMed] [Google Scholar]
- 101.Boutant M, Canto C. SIRT1 metabolic actions: integrating recent advances from mouse models. Mol Metab. 2014 doi: 10.1016/j.molmet.2013.10.006.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Suwa M, Nakano H, Radak Z, et al. Endurance exercise increases the SIRT1 and peroxisome proliferator-activated receptor gamma coactivator-1alpha protein expressions in rat skeletal muscle. Metabolism. 2008;57:986–98. doi: 10.1016/j.metabol.2008.02.017. [DOI] [PubMed] [Google Scholar]
- 103.Chen D, Bruno J, Easlon E, et al. Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev. 2008;22:1753–7. doi: 10.1101/gad.1650608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Pfluger PT, Herranz D, Velasco-Miguel S, et al. Sirt1 protects against high-fat diet-induced metabolic damage. Proc Natl Acad Sci U S A. 2008;105:9793–8. doi: 10.1073/pnas.0802917105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Canto C, Gerhart-Hines Z, Feige JN, et al. AMPK regulates energy expenditure by modulating NAD+metabolism and SIRT1 activity. Nature. 2009;458:1056–60. doi: 10.1038/nature07813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Lan F, Cacicedo JM, Ruderman N, et al. SIRT1 modulation of the acetylation status, cytosolic localization, and activity of LKB1. Possible role in AMP-activated protein kinase activation. J Biol Chem. 2008;283:27628–35. doi: 10.1074/jbc.M805711200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107•.Ruderman NB, Xu XJ, Nelson L, et al. AMPK and SIRT1: a longstanding partnership? Am J Physiol Endocrinol Metab. 2010;298:E751–60. doi: 10.1152/ajpendo.00745.2009. This article reviews the inter-relationship of SIRT1 and AMK in regulating each other and additional molecules that affect cellular function and the predisposition to metabolic syndrome-associated diseases. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Brenmoehl J, Hoeflich A. Dual control of mitochondrial biogenesis by sirtuin 1 and sirtuin 3. Mitochondrion. 2013;13:755–61. doi: 10.1016/j.mito.2013.04.002. [DOI] [PubMed] [Google Scholar]
- 109.Gillum MP, Kotas ME, Erion DM, et al. SirT1 regulates adipose tissue inflammation. Diabetes. 2011;60:3235–45. doi: 10.2337/db11-0616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Chalkiadaki A, Guarente L. High-fat diet triggers inflammation-induced cleavage of SIRT1 in adipose tissue to promote metabolic dysfunction. Cell Metab. 2012;16:180–8. doi: 10.1016/j.cmet.2012.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Xu XJ, Pories WJ, Dohm LG, et al. What distinguishes adipose tissue of severely obese humans who are insulin sensitive and resistant? Curr Opin Lipidol. 2013;24:49–56. doi: 10.1097/MOL.0b013e32835b465b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Peng Y, Rideout DA, Rakita SS, et al. Does LKB1 mediate activation of hepatic AMP-protein kinase (AMPK) and sirtuin1 (SIRT1) after Roux-en-Y gastric bypass in obese rats? J Gastrointest Surg. 2010;14:221–8. doi: 10.1007/s11605-009-1102-5. [DOI] [PubMed] [Google Scholar]
- 113.Price NL, Gomes AP, Ling AJ, et al. SIRT1 is required for AMPK activation and the beneficial effects of resveratrol on mitochondrial function. Cell Metab. 2012;15:675–90. doi: 10.1016/j.cmet.2012.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Hardie DG, Ross FA, Hawley SA. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol. 2012;13(4):251–62. doi: 10.1038/nrm3311.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Steinberg GR, Kemp BE. AMPK in health and disease. Physiol Rev. 2009;89(3):1025–78. doi: 10.1152/physrev.00011.2008.. [DOI] [PubMed] [Google Scholar]