

Abstract
Considerable effort has been expended to identify genes that account for myeloid lineage commitment and development. However, currently available non-invasive mouse models utilize myeloid-specific reporters that are significantly expressed in hematopoietic stem cells as well as lymphoid compartments. Here, we describe a myeloid-specific marker that is not shared by any other lineage. We show that lactotransferrin mRNA is expressed by Gr-1+/CD11b+ cells in the bone marrow, as opposed to hematopoietic stem cells or any peripheral cell population. To follow the progeny of lactotransferrin-expressing bone marrow cells, we generated a mouse model in which a reporter gene is irreversibly activated from the lactotransferrin-promoter. We found that lactotransferrin-reporter labels a majority of neutrophils, monocytes, macrophages and distinct subtypes of dendritic cells, while excluding T, B, natural killer cells, interferon-producing killer dendritic cells, plasmacytoid dendritic cells, erythrocytes and eosinophils. Lactotransferrin-reporter− bone marrow cells retain lymphoid, erythroid and long-term repopulating potential, while lactotransferrin-reporter+ bone marrow cells confer only myeloid, but not lymphoid potential. We conclude that lactotransferrin represents a late stage differentiation marker of neutrophils, macrophages and distinct subtypes of dendritic cells.
Introduction
There is general agreement that hematopoiesis is established through hierarchical stages from long-term repopulating hematopoietic stem cells (LT-HSCs) to more lineage-restricted cells.1 Segmentation of common myeloid progenitors (CMPs), giving rise to the myelo/erythroid lineage, and the common lymphoid progenitors (CLPs), developing into the lymphoid lineage, is generally accepted as the initial branching point in the classical hematopoietic lineage map.2–4
Several recently defined models have challenged this idea. Experimentally, many primordial and mature cell populations give rise to myeloid cells, suggesting a “myeloid-biased” model of hematopoiesis in which the myeloid potential is retained even after T and B cells have diverged.5–7 Accordingly, Flt3high multipotent progenitors (LMPPs) have been shown to give rise directly to myeloid and lymphoid cells.8 Similarly, fate-tracing of the transcription factors PU.1 and GATA-1 has suggested that two distinct progenitor entities, the classical (GATA-1+) CMPs and the (PU.1+) LMPPs give rise to myelo/erythroid and myelo/lymphoid cells, respectively.9 In contrast, in a recent work using the interleukin-7 receptor alpha (IL7Rα)-fate-tracing mouse model the authors showed that IL7Ra+ T-cell progenitors never develop into myeloid cells in vivo.10
Apart from myeloid potential in lymphoid progenitors, it has also been claimed that myeloid progenitors possess lymphoid potential in vitro and in vivo.2,11,12 Here, CMPs yield CD19+ B cells in vitro,2 while CMPs and granulocyte-macrophage progenitors (GMPs) display significant amounts of lymphoid genes and give rise to a substantial proportion of B and T cells when injected intravenously and intrathymically into sublethally irradiated recipients.11,12 Single cell experiments performed along with these studies excluded the possibility that the “converse” lineage had arisen from contaminating myeloid and lymphoid progenitors.
These multiple origins of myeloid and lymphoid cells are not consistent with the classical dichotomic model of hematopoiesis, assuming mutual exclusion of the other lineage in the newly generated progenitor. One possible explanation is that myeloid and lymphoid lineages are resolved more downstream of CMPs and CLPs.7 At least for the lymphoid lineage, expression of IL7Rα on progenitors seems to mark the end of the myeloid-bias.10 Hence, there is a need to evaluate whether myeloid progenitors possess lymphoid potential at any stage of fate of myeloid development.
Currently available non-invasive mouse models employ gene promoters, which are promiscuously expressed among HSCs, LMPPs, CMPs and GMPs. In mice harboring a LysM-Cre transgene together with a Rosa26 YFP reporter, transplantation of YFP+ LT-HSCs is able to induce long-term engraftment in recipient mice.13 Similarly, mice carrying a GFP-reporter in the PU.1-locus show GFP expression in LT-HSCs.14,15 More recently, in vivo tracing of C/EBPα during hematopoiesis has revealed that HSCs express C/EBPα.16 To the best of our knowledge, there is no available mouse model for tracing myeloid cells without concurrent labeling of HSCs and lymphoid cells.
Lactotransferrin (LTF, LF, CSP82) is well known as an iron-binding protein in the milk, saliva and mucosal secretions of the trachea, uterus and ovaries and has been implicated in innate immune responses against microbial infections.17,18 Ltf knockout mice have normal iron homeostasis and show no gross abnormalities with respect to terminal differentiation into hematopoietic lineages.19 However, LTF exerts various immunomodulatory effects in monocytes, macrophages and neutrophils17 and may affect myelopoiesis.20,21 A membrane-bound form of LTF (CSP82) has been implicated in regulating dendritic development in vitro and in vivo.22 In a search for myeloid-specific markers that are absent from HSCs, lymphoid and erythroid cells, we analyzed the expression pattern of Ltf during hematopoietic development.
Here, we show that Ltf is specifically expressed in Gr-1+/CD11b+ bone marrow (BM) cells. To delineate the cellular compartments derived from Ltf+ progenitors during hematopoietic development in vivo, we employed a lineage-tracing mouse model expressing Cre recombinase under the control of the Ltf-promoter, together with the irreversibly Cre-inducible tandem-dimer red fluorescent protein (tdRFP).23 We show the Ltf-reporter+ cells give rise to populations of dendritic cells (DCs), monocytes, macrophages and neutrophils, while sparing eosinophils, T cells, B cells, natural killer (NK) cells and erythrocytes.
Methods
Mice
All mice were kept under specific pathogen-free conditions at the University of Veterinary Medicine Vienna and the Research Institute of Molecular Pathology. Animal experiments were discussed and approved by the institutional ethics committees and were performed in accordance with Austrian law (BMWF-68.205/0243-II/3b/2011 and 66.009/0065-II/10b/2009).
Generation of B6;129Sv-Tg(Ltf-iCre)14 transgenic mice
A codon-improved Cre (iCre) recombinase was inserted into a bacterial artificial chromosome carrying the Ltf gene (BAC; RP24-166N8; purchased from Children’s Hospital Oakland Research Institute, Oakland, CA, USA) via homologous recombination in E. coli. A cassette containing the iCre recombinase, an artificial intron, a bovine growth hormone polyadenylation signal and an ampicillin-resistance gene flanked by Frt (Flp recombinase target) sites, was recombined into the first exon of the Ltf gene. To delete the ampicillin gene, a plasmid expressing FLP recombinase was transfected into E. coli harboring the recombined BAC. Correct insertion and ampR-gene deletion were verified by Southern blotting and by DNA sequencing. BAC was purified using a CsCl-gradient and ultracentrifugation and injected into the pro-nuclei of B6/129SvF1 oocytes. Genotyping of B6;129Sv-Tg(Ltf-iCre)14 (Ltf-Cre+) mice was performed with primers GGAAGCTGGCCTCTAAGAAC (forward) and ATCAGAGGTGGCATCCACAG (reverse), yielding a 259 base pair polymerase chain reaction product.
Fluorescence activated cell sorting analysis, staining strategy and cell sorting
The following antibodies were used: Ter119 (Ly-76, Ter119)-APC, CD3ɛ (145-2C11)-PerCP, CD8α (Ly2)-PerCP-Cy5.5, CD19 (1D3)-APC and -APC-Cy7, CD90.2 (Thy1.2, 53-2.1)-APC, Ly6G/Ly6C (Gr-1, RB6-8C5)-APC, CD34 (RAM34)-FITC and -APC, Ly6A/E (Sca-1, D7)-PE-Cy7, NK1.1 (PK136)-APC, CD11b (Mac-1, M1/70)-APC, CD71 (transferrin receptor, C2)-biotin and second step detection streptavidin-APC-Cy7 were obtained from BD Pharmingen. CD3ɛ (eBio500A2)-APC and –Pacific Blue, CD45R (B220; RA3.6B2)-PacificBlue, Ter119 (Ly-76, Ter119)-eFluor450, Ly6G (Gr-1, RB6-8C5)-PacificBlue, CD62L (L-Selectin, DREG-56)-APC-eFluor780, CD11b (Mac-1, M1/70)-PE-Cy7 and –Pacific Blue, CD127 (IL7Rα, A7R34)-biotin and –Pacific Blue, Ly6C (HK1.4)-eFluor450, CD135 (Flk-2/Flt3/Ly-72, A2F10.1)-biotin, CD16/CD32 (FcγIII/IIR, 2.4G2)-biotin, CD172a (SIRPa; PB8)-PerCP-eFluor710, CD11c (N418)-PE-Cy7, CD115 (M-CSFR/c-fms; AFS98)-FITC, MHC-II (I-A/I-E; M5/114.15.2), Ly6A/E (Sca-1, D7)-APC, F4/80 (BM8)-APC, SiglecH (eBio440c)-PE-Cy7 and CD117 (c-kit, 2B8)-PE-Cy5 were purchased from eBioscience. Ly6G (1A8)-PerCP-Cy5.5 was purchased from Biolegend. Staining strategies are summarized in Online Supplementary Table S1. Flow cytometric analysis of all organs was performed on an eight-color BD FACSCantoII (BD Biosciences) equipped with 488 nm, 633 nm and 405 nm lasers. Fluorescence activated cell sorting (FACS) was performed at 4°C immediately after biopsy using a ten-color BD FACSAriaIII device equipped with 488 nm, 561 nm, 633 nm and 395 nm lasers.
Statistical analysis
The log rank test was used to compute survival curves. One-way ANOVA and Tukey or Bonferroni multiple comparison tests assuming a 95% confidence interval were performed for all organ distributions and tdRFP frequencies. Student t-tests were used for quantitative polymerase chain reaction data and colony-forming unit (CFU) assays.
Results
Ltf mRNA and Ltf promoter-driven transgene expression is restricted to Gr-1+/CD11b+ cells in the bone marrow
To determine the availability of Ltf mRNA during distinct stages of hematopoietic development, we examined the expression of Ltf transcripts throughout stem, progenitor/precursor stages in the BM and among distinct hematopoietic lineages in peripheral blood. In the BM, abundant Ltf mRNA expression was readily detectable in Gr-1+/CD11b+ cells, but was below detection limits in HSCs, CLPs and myeloid progenitors (CMPs and MEPs, Figure 1A). GMPs showed low expression of Ltf mRNA. Surprisingly, Ltf mRNA was below the detection limit in NK1.1+, CD3ɛ+, Ter119+/FSClow and CD11c+ cells and barely detectable in Gr-1+/CD11b+ cells in the periphery (Figure 1A). These results demonstrate that Ltf is absent from stem/progenitor compartments and peripheral blood cells, while being expressed by Gr-1+/CD11b+ BM cells. Given the fact that all myeloid cells originate in the BM, we aimed to analyze myeloid lineages in the periphery that have developed from Ltf-expressers in the BM.
Figure 1.
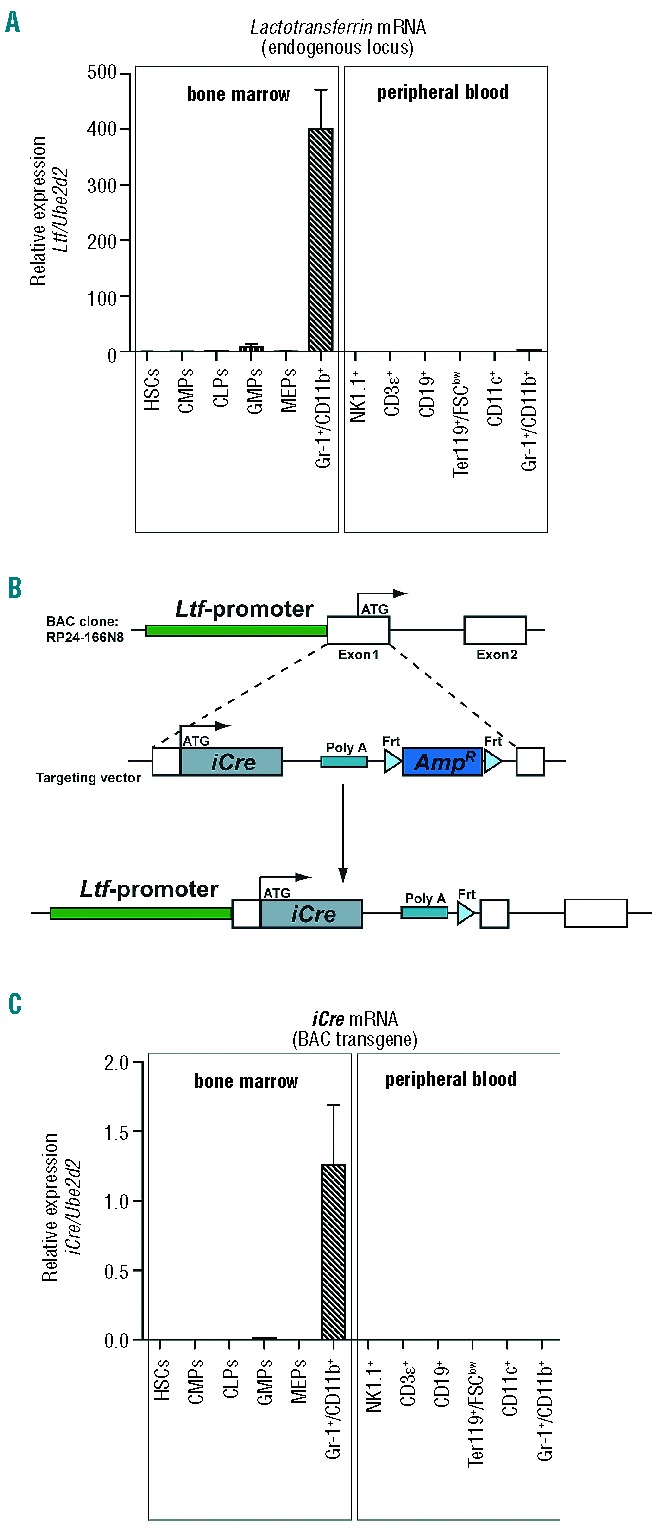
Ltf-expression is restricted to myeloid cells in the BM. (A) Summary of quantitative polymerase chain reaction (Q-PCR) analysis depicting the relative expression of Ltf mRNA from the endogenous locus for the indicated hematopoietic populations. Expression was normalized to Ube2d2 expression. Results of one representative quantification experiment out of two are depicted. Total RNA was isolated from 5000 cells from each population presented. Bars show means ± SEM. (B) Schematic representation of the BAC cloning strategy used for the generation of Ltf-Cre-transgenic mice. A cassette carrying iCre and regulatory elements was cloned into exon 1 of the Ltf-gene from the BAC-clone RP24-166N8. (C) Q-PCR analysis of iCre mRNA expression from the BAC-transgene. Values were normalized to Ube2d2 expression. Total RNA was isolated from 5000 cells/population presented. Bars show means ± SEM. Results of one representative experiment out of two are depicted.
We generated transgenic mice expressing Cre recombinase under the control of Ltf locus regulatory regions (Ltf-Cre mice). The transgene cloning strategy is summarized in Figure 1B. Injection of the recombinant engineered BAC transgene into zygotes resulted in two transgenic lines (#14 and #15). The offspring of both founder lines did not show any overt phenotypic differences in gross morphology, life expectancy or breeding efficiency, nor did we observe any organ pathologies during a breeding period of >36 months (data not shown). Line 14 was used for further analysis. We next analyzed transgene expression in non-hematopoietic organs in hemizygous Ltf-Cre+ mice. Codon-improved Cre (iCre) mRNA was not detectable in intestinal, liver or muscle tissues from various individuals. Only marginal iCre expression was observed in brain and breast tissues from one out of three individuals (Online Supplementary Figure S1A). To exclude any detrimental effects of the transgene expression, we assessed relative and total percentages of hematopoietic cells in the BM and in the periphery. Total percentages of myeloid progenitors in the BM were not different in Ltf-Cre+ mice as compared to wild-type littermates (Online Supplementary Figure S1B,C). Similarly, wild-type and transgenic animals showed equal distributions of peripheral myeloid- and lymphoid-derived cells (Online Supplementary Figure S1D).
To determine whether the iCre expression in Ltf-Cre+ mice is confined to developmental stages with detectable Ltf expression, we next investigated iCre-transcripts in distinct hematopoietic populations. Again, iCre mRNA was detectable in BM-derived Gr-1+/CD11b+ cells (Figure 1C), but lacking in HSCs and CMPs. We did not detect iCre mRNA on myeloid, lymphoid or erythroid cells in the periphery (Figure 1C).
Ltf-Cre gives myeloid cell specific reporter expression
To clarify which hematopoietic lineages display iCre-activity, we crossed Ltf-Cre+ mice with mice carrying a Cre-inducible tdRFP reporter gene in the Rosa26 locus (Rosa26flSTOPfl−tdRFP). These mice possess a transcriptional stop cassette flanked by two heterologous loxP sites, enabling the irreversible removal of the stop cassette and subsequent expression of the reporter gene in iCre-expressing cells (Figure 2A).23 All progeny of Ltf-expressing cells in Ltf-Cre+ Rosa26tdRFP mice (henceforth: Ltf-reporter+ mice) should express tdRFP. We examined the contribution of Ltf-reporter+ cells to individual hematopoietic populations in the bone marrow and peripheral blood. We found that tdRFP+ cells did not contribute to LT-HSCs, ST-HSCs, MPPs or CLP-fractions in the BM (Figure 2B). In contrast, a small proportion of CMPs and GMPs expressed tdRFP (0.4±0.2% each). The latter was confirmed using a recent staining protocol for early myeloid stages:24 we found comparable percentages of Ltf-reporter+ KLS−CD41−CD150−FcγRII/III+ GMPs (0.4± 0.1%; Online Supplementary Figure S2A). No reporter-activity was detected in lymphoid cells, while a fraction of erythroid cells (Ter119+/FSClow; 0.1±0.1%) as well as the vast majority of Gr-1+/CD11b+ cells (95±2.4%) expressed tdRFP (Figure 2C). Figure 2C summarizes data obtained from peripheral blood analyses of 12 individual mice. To determine whether any residual iCre-expression exists in hematopoietic lineages, we FACS-sorted BM-derived Gr-1+/CD11b+ cells and BM cells depleted of Gr-1+/CD11b+ cells regardless of Ltf-reporter expression. As indicated in Figure 2D, Gr-1+/CD11b+ cells expressed on average 96.8±1.5% of detectable iCre-transcripts in whole BM cells from four individual mice. In contrast, BM cells depleted of Gr-1+/CD11b+ cells contributed to 3.2±1.5% of detectable iCre mRNA, suggesting that this percentage may correspond to iCre+ myeloid and erythroid progenitor stages in the BM (CMPs, GMPs, Ter119+/FCSlow cells, etc.). We next analyzed cells of the dendritic lineage and observed that only distinct subtypes of DCs were Ltf-reporter+ (Online Supplementary Figure S2B–E). Ltf-reporter-expression varied between 10 to 60% in CD11chigh/MHCIIhigh/CD8α+ and CD11chigh/MHCIIhigh/CD8α − DCs (Online Supplementary Figure S2B,E), suggesting a heterogeneity that cannot be resolved using classical DC cell markers (CD11c, CD11b, MHCII and CD8α). The majority of CD11cint/MHCIIint/CD11b+/B220+/CD8α+ cells were Ltf-reporter+, while hardly any CD11cint/MHCIIint/CD11b−/B220+/CD8α+ and CD11cint/MHCIIint/CD8α − cells expressed tdRFP (Online Supplementary Figure S2B,E). Similarly, no reporter activity was detected in CD11cint/CD11b−/B220+/SiglecH+ plasmacytoid DCs (Online Supplementary Figure S2C) and NK1.1+/DX5+/CD3ɛ−/CD11cint/B220+ interferon-producing killer DCs (iKDCs) (Online Supplementary Figure S2D), except for a tiny population of CD11cint/CD11b+/B220+/SiglecH+ plasmacytoid DCs (Online Supplementary Figure S2C). We further investigated the succession of the dendritic lineage in the BM using markers for the recently identified earliest clonogenic progenitor of DCs, the myeloid progenitors (MPs), the monocyte/macrophage and DC precursor (MDP), and its successor stages, common DC-precursors (CDPs) and pre-classical dendritic cells (pre-cDCs).4 Pre-cDCs expressed significant amounts of tdRFP (60±12%, Online Supplementary Figure S2C).
Figure 2.
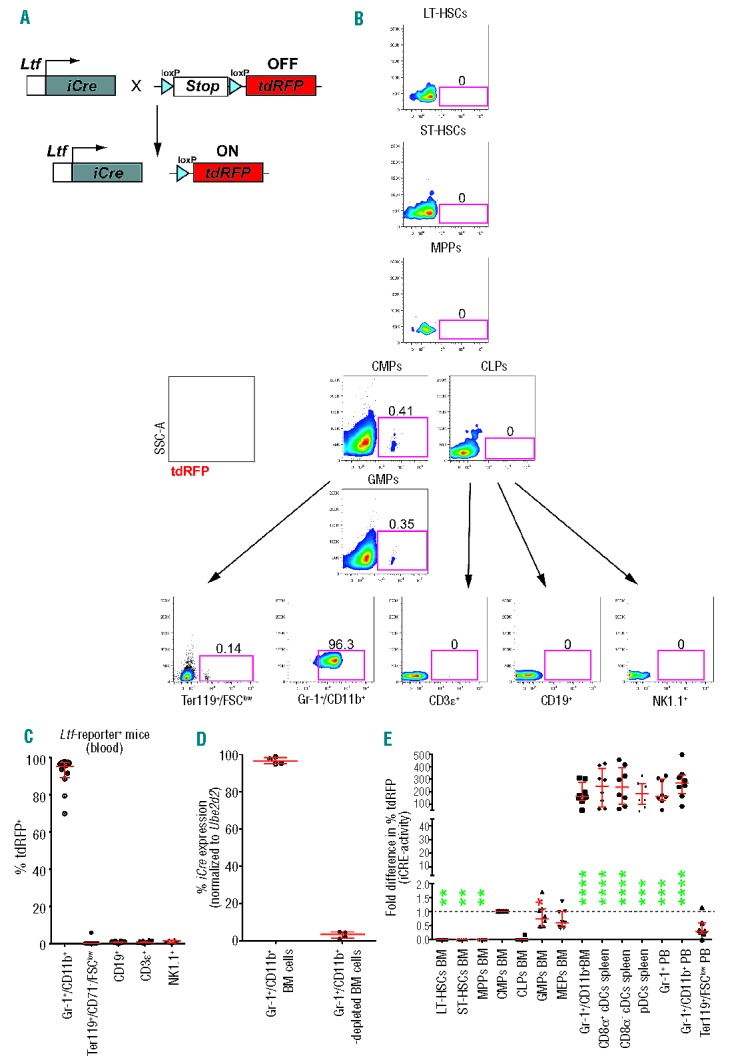
Fate-tracing reveals contributions to granulocytic and dendritic lineages. (A) To visualize the history of Ltf-expression, Ltf-Cre transgenic mice were crossed with inducible Rosa26 tdRFP mice. Only upon usage of Ltf-promoter does iCre transcription and activity result in excision of the Stop-cassette and continuous expression of the reporter. (B) Representative FACS data of tdRFP-expression in multiple cell populations from Ltf-reporter mice are depicted in a hierarchical model. (C) Summary of data obtained from peripheral blood analyses of 12 individual mice. Reporter labeling reveals exclusive myeloid reporter expression in the peripheral blood of Ltf-reporter mice. Means ± SEM are indicated by red error bars. (D) Quantitative polymerase chain reaction analysis of iCre mRNA expression in BM-derived Gr-1+/CD11b+ versus Gr-1+/CD11b+-depleted BM cells. Values were normalized to Ube2d2 expression. The expression was further normalized to total iCre mRNA detected in both samples (100%). Bars show means ± SEM of four biological replicates. (E) Fold difference in the percentage of tdRFP+ cells compared to tdRFP+ CMPs from the same mouse. HSCs and CLPs do not contain any floxed alleles. Mature Ltf-reporter+ classic DCs (cDCs), plasmacytoid (pDCs), granulocytes and erythrocytes display a strong increase in iCRE-activity compared to Ltf-reporter+ CMPs. No comparisons between mature cell populations were statistically significant (n=12). Error bars indicate medians ± interquartile range. *P<0.05, **P<0.01, ***P<0.001 and ****P<0.0001.
Variations in expression of the reporter gene were recently observed in a different mouse model and were attributed to different iCRE-activities at the floxed-stop cassette.25,26 To determine whether the distinct myeloid lineages in the periphery become reporter+ due to differentiation through an Ltf+ progenitor stage in the BM or due to aberrant recombination, we tested the biological variation of iCRE-activity by analyzing the end product of recombination within a large cohort of mice. The proportion of reporter+ cells in all progeny derived from the developmentally first reporter+ progenitor must be at least equal or higher to that of the reporter+ progenitor (Jakubzick et al., 2008). Repeated utilization of the Ltf-promoter during differentiation will result in a further increase in the proportion of Ltf-reporter+ cells. To this end, we normalized the percentage of tdRFP+ cells in myeloid populations to the percentage of the developmentally first progenitor expressing the Ltf-reporter (i.e. CMPs; see Figure 2B) in each individual mouse.25,27 As expected, iCRE-activities in LT-HSCs, ST-HSCs, MPPs and CLPs were not detectable (Figure 2E). GMPs exerted comparable iCRE-activity to CMPs. Percentages of Ltf-reporter+ Ter119+/FSClow erythrocytes did not differ significantly from those of MEPs (P=ns), confirming that no additional Ltf-reporter expression had taken place in the erythroid lineage. In contrast, BM-derived Gr-1+/CD11b+ cells displayed a profound increase in iCRE-activity which was comparable to that of the myeloid cells in the periphery. Importantly, we did not detect any statistically significant differences among monocytes/macrophages/granulocytes and DCs. These data point to a myeloid-restricted expression of tdRFP in Ltf-reporter+ mice.
Low level of Ltf-reporter expression during erythropoiesis
As a proportion of erythroid cells express tdRFP (Figure 2C), we examined reporter gene expression throughout distinct developmental stages of erythropoiesis in the BM. We found that percentages of MEPs (0.3± 0.2%) were not significantly increased as compared to CMPs (P>0.5, ns; Figure 3A). We did not observe any contribution of tdRFP+ cells to pre-Meg/E, MkP or pre-CFU-E populations24 (Figure 3A). To identify Ltf-reporter expression in subsequent stages of erythropoiesis, we used combined staining of CD71 and Ter119.28 FACS plots from one representative mouse are depicted in Figure 3B. We observed that percentages of Ltf-reporter+ cells were slightly increased in fraction R1 (0.71± 0.26%) but declined throughout the following stages of development (R2: 0.64±0.53%; R3: 0.26±0.09%; R4: 0.31±0.23%). In fraction R5, only 0.06±0.03% cells expressed tdRFP (Figure 3B). This led us to conclude that Ltf-reporter expression is sparse in the erythroid lineage.
Figure 3.
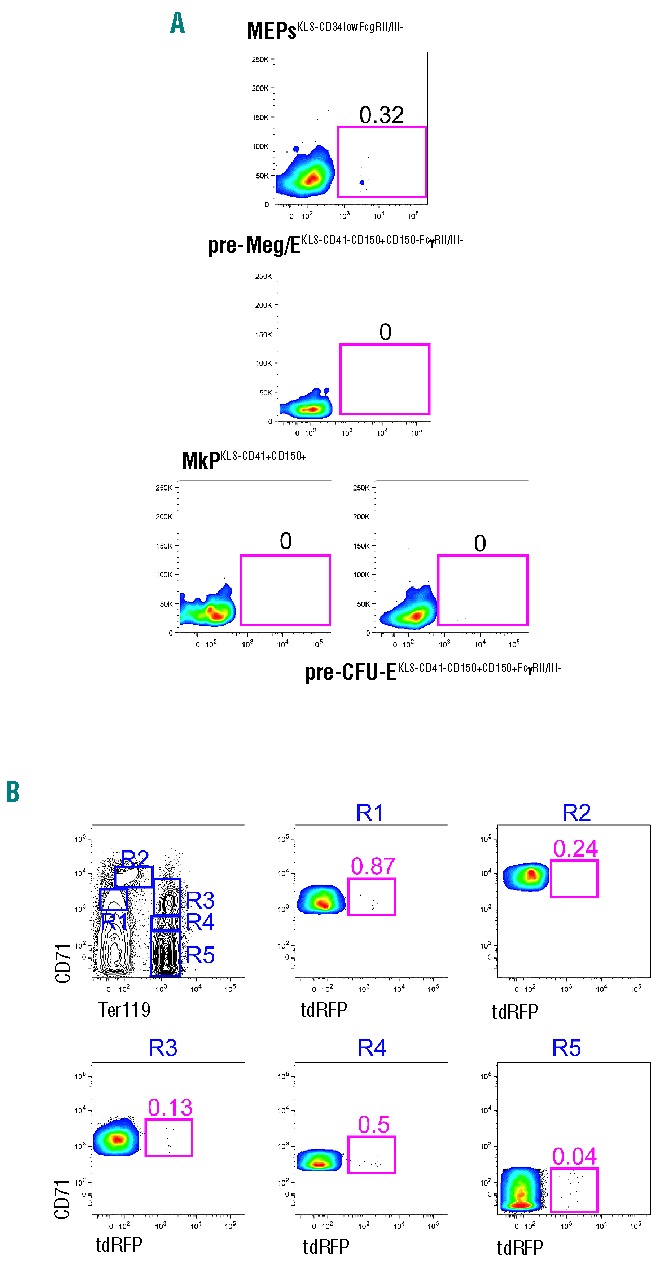
Ltf-reporter mice do not label erythroid cells. (A) (From top to bottom) proportion of tdRFP+ (I) MEPs as defined by gating on lin−/c-kit+/Sca-1−(KLS−)/CD34low/FcγRII/III−, (II) pre-Meg/Es as defined by KLS−/CD41−/CD150+/CD105−/FcγRII/III−, (III) MkPs as defined by KLS−/CD41+/CD150+ and (IV) pre-CFU-Es as defined by KLS−/CD41−/CD150+/CD105+/FcγRII/III+ cells. (B) tdRFP expression in erythoid precursors as determined by differential CD71 and Ter119 co-expression. One representative FACS plot out of 12 showing BM cells is depicted.
Ltf-reporter+ bone marrow lacks lymphoid potential in vitro
HSC promiscuously express myeloid-specific markers.9,29,30 Accordingly, transplantation of EGFP+ BM cells from LysM-Cre EGFP mice yields long-term reconstitution in irradiated recipients.13 In addition, CMPs and GMPs were found to have pronounced lymphoid potential in vitro and in vivo.2,11,12 To exclude that any Ltf-reporter+ population in the BM contained multilineage or lymphoid potential, we evaluated the potential of the entire Ltf-reporter+ BM entity to generate multilineage colonies in a semi-solid medium. We thus fractionated BM cells into Ltf-reporter− and Ltf-reporter+ cells and compared their potential to differentiate under defined cytokine-instructive conditions in vitro. We seeded BM cells in methylcellulose supplemented with stem cell factor, interleukin (IL)-3, IL-6, IL-7, Flt-3L, insulin-like growth factor-1, erythropoietin and thrombopoietin in order to induce myeloid, lymphoid and erythroid differentiation. Morphological characterization of colonies revealed that Ltf-reporter− BM cells have multilineage potential, giving rise to colonies of the myeloid, erythroid and lymphoid lineages (CFU-GEMM, CFU-GM, CFU-G, CFU-M, BFU-E, CFU-E, CFU-Meg and CFU-pro-B; Figure 4A). In contrast, Ltf-reporter+ BM cells formed colonies only corresponding to CFU-M and CFU-G. Ltf-reporter+ BM cells did not confer any lymphoid potential in vitro. Representative pictures of colonies attributed to individual lineages are provided in Online Supplementary Figure S4. These data indicate that Ltf-reporter+ BM cells lack multipotency in colony-forming assays and confer solely CFU-G and CFU-M activities.
Figure 4.
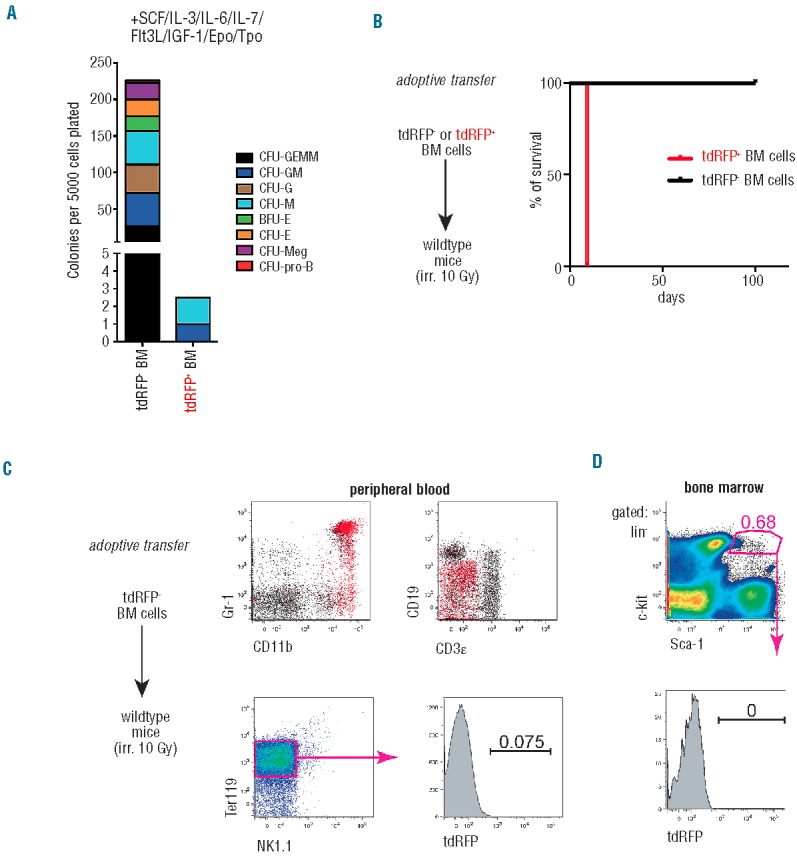
Lack of self-renewal and lymphoid capacity of Ltf-reporter+ BM cells in vitro and in vivo. (A) Total colony formation activity and clonogenic analysis of tdRFP+ and tdRFP− BM cells upon multilineage differentiation cues. See Methods section for the cytokines used in this assay. The colonies were counted on day 7. (B) Kaplan-Meier plot showing survival of lethally-irradiated wild-type mice transplanted with tdRFP+ or tdRFP− BM cells from Ltf-reporter+ mice (n=5 for each group). (C) Analysis of de novo generation of tdRFP+ cells in mice receiving Ltf-reporter−BM cells. Bi-colored FACS plots indicate myeloid, but not lymphoid lineage contribution of de novo-generated tdRFP+ cells on day 100 after transplant (red color; left panels). HSCs do not express tdRFP after long-term reconstitution (right panels). Representative FACS plots are shown.
Ltf-reporter+ bone marrow cells lack self-renewal capacity
Next, we tested for multilineage and long-term reconstitution capacity in vivo by transplanting purified Ltf-reporter+ and Ltf-reporter− BM cells into lethally-irradiated mice. We observed that Ltf-reporter+ BM cells failed to rescue mice from irradiation-induced BM depletion: recipient mice deceased coincidentally with the irradiation control (Figure 4B and data not shown) and thus did not receive long-term repopulation potential. Only the Ltf-reporter-population had the potential for multilineage reconstitution (Figure 4B). To test whether the Ltf-reporter becomes activated during transplantation-induced myeloid differentiation from the Ltf-reporter− (HSC-containing) fraction, we investigated de novo generation of Ltf-reporter+ myeloid cells in transplanted mice. Myeloid cells of Ltf-reporter− origin did indeed regain tdRFP expression, while lymphoid and erythroid cells remained Ltf-reporter− (Figure 4C, left panels). Moreover, no tdRFP expression could be observed in HSCs from these mice (Figure 4D).
Ltf-reporter expression separates monocytes, macrophages, neutrophils and dendritic cells from eosinophils, lymphocytes and erythroid cells
To determine which myeloid subtypes express the Ltf-reporter, we next FACS-sorted Ltf-reporter+ and Ltf-reporter− cell fractions from the BM. Grünwald-Giemsa staining revealed neutrophils, monocytes and macrophages in the Ltf-reporter+ fraction (Figure 5A, left panel). In the Ltf-reporter− fraction, we detected eosinophils together with lymphocytes and erythroid cells (Figure 5A, right panel). To confirm this, we analyzed BM, spleen and blood cells of Ltf-reporter+ mice for the expression of Ly6C, CD11b, CD115, F4/80, CD62L and Ly6G. The combination of these markers covers the identification of granulocytes, monocytes and macrophages.31,32 Resolution by Ly6C and CD11b revealed seven distinct populations (I–VII), which were differently distributed among BM and peripheral cells (spleen and blood; Figure 5B). As depicted in Figure 5C, individual fractions differed in their expression of CD115, F4/80, CD62L and Ly6G. Ltf-reporter+ cells could only be found within three different myeloid populations: II, V and VII (Figure 5D). To characterize these fractions morphologically, cytospins of FACS-purified cells were stained with Grünwald Giemsa. Ltf-reporter+ Ly6Chigh/CD11b+ cells (population II) displayed various stages of monocytic development (Figure 5E, left panel and Online Supplementary Figure S5A). tdRFP+ Ly6C+/CD11bhigh cells (population V) consisted of a homogeneous population of neutrophils containing segmented ring-shaped nuclei (Figure 5E, middle panel). Ltf-reporter+ Ly6C−/CD11bhigh cells (population VII) morphologically correspond to macrophages (Figure 5E, right panel). In contrast, no tdRFP+ cells could be found within Ly6Cint/CD11bint cells (population IV) which are enriched for distinct developmental stages of eosinophils (Online Supplementary Figure S5B).
Figure 5.

Ltf-reporter expression in monocytes, macrophages and neutrophils. (A) Morphology of Ltf-reporter+ and Ltf-reporter− BM cells was determined by Grünwald-Giemsa staining from sorted fractions. Representative cells from individual images of the same specimen were grouped with regard to their morphology as described elsewhere.24 Notably, eosinophils were enriched specifically in the Ltf-reporter− BM fraction. (B–E) Murine BM, spleen and peripheral blood cells from Ltf-reporter+ mice were stained with antibodies against CD11b, Ly6C, Ly6G, CD115, F4/80 and CD62L. (B) Resolution of live cells with CD11b and Ly6C reveals seven distinct populations (I–VII). (C) Each population was further gated for CD115 and F4/80 (upper panels) and CD62L and Ly6G (lower panels). (D) Histograms depict the intensity of Ltf-reporter in each of the seven populations. Populations II, V and VII consist of Ltf-reporter+ cells. (E) Morphology of populations II, V and VII was determined by Grünwald-Giemsa staining on cytospin preparations from sorted cells. Ly6Chigh/CD11b+/CD115+/F4/80+/CD62L+/Ly6Glow cells resemble monocytes (population II), while Ly6C+/CD11bhigh/CD115−/F4/80−/CD62L+/Ly6Ghigh cells consist of ring-shaped neutrophilic precursors. Ly6C−/CD11bhigh/CD115+/F4/80+/CD62Llow/Ly6Glow resemble macrophages.
Discussion
The development of myeloid cells is a complex and tightly regulated process giving rise to innate immune cells that combat pathogens such as bacteria, viruses, fungi and parasites. Much effort has been expended to identify cellular pathways, genes and growth factors that mediate myeloid lineage commitment. In the past, seminal studies addressed fate-tracing of myeloid9,13,15,16,27 development in vivo. The outcomes of these studies have suggested that the precise branching site of the myeloid lineage, as postulated in the classical model of hematopoiesis, might be difficult to approximate because of promiscuous expression of putative lineage-restricted genes by HSCs. To the best of our knowledge, there is currently no mouse model available which utilizes a myeloid-specific gene whose expression does not coincide with the HSC stage.
Here, we identified Ltf as a myeloid-specific marker whose expression is absent in HSCs and in more than 99% of CMPs and GMPs. Ltf-expression is limited to Gr-1+/CD11b+ cells residing in the BM, but not to peripheral cells of the myeloid lineage. To determine the contribution of Ltf-expressing cells to the myeloid lineage during steady-state hematopoiesis in vivo, we generated Ltf-Cre mice. We found that both Ltf and iCre mRNA was specifically expressed in Gr-1+/CD11b+ cells in the BM, thus following the reported expression pattern of Ltf in BM cells (https://gexc.stanford.edu/search/3/gene/Ltf). Analysis of iCre expression also revealed that only 3.2±1.5% of all detectable iCre-transcripts in the BM do not belong to Gr-1+/CD11b+ cells.
The exclusive expression of Ltf in the BM, but not in peripheral blood cells, allowed us to mark and follow the development of peripheral myeloid cells by crossing Ltf-Cre mice with a reporter strain (Ltf-reporter mice). Ltf-reporter mice efficiently labeled peripheral cells such as ring-shaped neutrophils, monocytes, macrophages and distinct subtypes of DCs, although they did not express Ltf mRNA. In contrast, eosinophils, T cells, B cells, interferon-producing killer DC and NK cells did not express the Ltf-reporter.
Currently, there is intense debate on the model of hematopoietic development due to findings that certain progenitor populations (CMPs, GMPs, LMPPs, CLPs, etc.) have the potential to develop into cells of more than one lineage (lineage-bias). Thus, in the myeloid lineage, classical myeloid progenitors (e.g. CMPs and GMPs) still possess some lymphoid potential.2,11,12 Similar observations have been made in the lymphoid lineage.5,6 These findings are contradictory to the dichotomy of the classical model of hematopoiesis describing a hierarchical and non-overlapping branching of lymphoid and myeloid lineages from multipotent stem cells. To reconcile the models, it has been postulated that lymphoid and myeloid lineages might be resolved at later stages.7,33 Support for this hypothesis comes from the finding that IL7Ra+ T-cell progenitors (CLPs) never develop into myeloid cells in vivo.10 Here we show that Ltf-expression is established late during myelopoiesis, subsequent to the erythroid and lymphoid branching and beyond the CMP/GMP stages. Ltf-reporter− BM cells retain multilineage potential in vitro and transplantation of Ltf-reporter− BM cells into lethally irradiated mice fully reconstitutes the hematopoietic system including de-novo generation of peripheral Ltf-reporter+ cells. One explanation is that that all labeled cell types express Ltf during terminal differentiation in the BM. This is in line with our finding that BM-derived Ltf-reporter+ cells have the potential to become CFU-G and CFU-M in vitro while lacking the potential to develop into lymphoid and erythroid lineages. The lack of cell surface markers indicative of stages of late myeloid development (downstream of GMPs) limits our efforts and represents a major drawback to understanding the consecutive events during late myeloid development. In the light of our results, it is tempting to speculate that Ltf-expression occurs independently in distinct subtypes of myeloid lineages (i.e. as a result of microenvironmental cues in the BM niche). Alternatively, a single hypothetic common neutrophil/monocyte/macrophage/dendritic progenitor cell might exist, which expresses Ltf that is subsequently found in its progeny. However, the verification of such a specific Ltf+ common progenitor requires the identification of additional independent markers.
The Ltf-reporter mouse model showed in vivo labeling of neutrophils, monocytes, macrophages and CD11cint/MHCIIint/CD11b+/B220+/CD8a+ DCs, as opposed to eosinophils, interferon-producing killer DCs, CD11cint/MHCIIint/CD11b−/B220+/CD8α+ and CD11cint/MHCIIint/CD8α− classical DCs and SiglecH+ pla-macytoid DCs. In the case of eosinophils, this might point to different origins of certain granulocytic subtypes. This is supported by a recent study showing that in human hematopoiesis, granulocyte subtypes originate from distinct hematopoietic lineages.34 Remarkably, the existence of a common erythro/myeloid progenitor - giving rise to human eosinophils, basophils and megakaryocytes - has already been proposed previously.35,36 Alongside with common erythro/myeloid progenitors, it has been proposed that there is a progenitor for neutrophils and macrophages downstream of LMPPs and GMPs.34 Interestingly, EPRO cells - an Ltf-expressing promyelocytic cell line37 - can be differentiated into either monocytes/macrophages or neutrophils, but not into eosinophils.38 It remains to be determined whether these progenitors express Ltf in human hematopoiesis and - if so - whether Ltf+ myeloid progenitors may give rise to neutrophils, monocytes, macrophages and subpopulations of DCs, while lacking the potential to develop into eosinophils, basophils or megakaryocytes.
In summary, our findings highlight the expression of Ltf in late myelopoiesis. Future experiments will be necessary to phenotypically characterize and prospectively isolate Ltf-reporter+ myeloid progenitors using yet undefined cell surface markers, thereby circumventing the current unavailability of anti-murine antibodies to recognize the membrane-bound LTF. In addition, Ltf-Cre mice will provide a powerful in vivo tool to study myeloid-specific gene functions using loss-of-function and gain-of-function approaches.
Acknowledgments
In memoriam of Hartmut Beug (1945–2011) under whose supervision this study was initiated. We thank Hans-Joerg Fehling for the generous gift of Rosa26flSTOPfl-tdRFP mice. The work was financially supported by the Austrian Science Fund (FWF SFB-F28) with funds to VS and MM and the GEN-AU program (project Austromouse) of the Austrian Federal Ministry of Science and Research. We thank Hans-Christian Theussl and Jacek Rodegar Wojciechowski for the help with zygote micro-injections and Miriam Kleiter and Siegfried Kosik for the help with mice irradiation. We also thank Graham Tebb for critically reading the manuscript and for helpful scientific suggestions.
Footnotes
The online version of this article has a Supplementary Appendix.
Authorship and Disclosures
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Orkin SH, Zon LI. Hematopoiesis: an evolving paradigm for stem cell biology. Cell 2008;132(4):631–44 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Akashi K, Traver D, Miyamoto T, Weissman IL. A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 2000;404(6774):193–7 [DOI] [PubMed] [Google Scholar]
- 3.Kondo M, Weissman IL, Akashi K. Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell 1997;91(5):661–72 [DOI] [PubMed] [Google Scholar]
- 4.Liu K, Victora GD, Schwickert TA, Guermonprez P, Meredith MM, Yao K, et al. In vivo analysis of dendritic cell development and homeostasis. Science 2009;324(5925):392–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bell JJ, Bhandoola A. The earliest thymic progenitors for T cells possess myeloid lineage potential. Nature 2008;452(7188):764–7 [DOI] [PubMed] [Google Scholar]
- 6.Wada H, Masuda K, Satoh R, Kakugawa K, Ikawa T, Katsura Y, et al. Adult T-cell progenitors retain myeloid potential. Nature 2008;452(7188):768–72 [DOI] [PubMed] [Google Scholar]
- 7.Kawamoto H, Ikawa T, Masuda K, Wada H, Katsura Y. A map for lineage restriction of progenitors during hematopoiesis: the essence of the myeloid-based model. Immunol Rev 2010;238(1):23–36 [DOI] [PubMed] [Google Scholar]
- 8.Adolfsson J, Mansson R, Buza-Vidas N, Hultquist A, Liuba K, Jensen CT, et al. Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revised road map for adult blood lineage commitment. Cell 2005;121(2):295–306 [DOI] [PubMed] [Google Scholar]
- 9.Arinobu Y, Mizuno S, Chong Y, Shigematsu H, Iino T, Iwasaki H, et al. Reciprocal activation of GATA-1 and PU.1 marks initial specification of hematopoietic stem cells into myeloerythroid and myelolymphoid lineages. Cell Stem Cell 2007;1(4):416–27 [DOI] [PubMed] [Google Scholar]
- 10.Schlenner SM, Madan V, Busch K, Tietz A, Laufle C, Costa C, et al. Fate mapping reveals separate origins of T cells and myeloid lineages in the thymus. Immunity 2010;32(3):426–36 [DOI] [PubMed] [Google Scholar]
- 11.Ng SY, Yoshida T, Zhang J, Georgopoulos K. Genome-wide lineage-specific transcriptional networks underscore Ikaros-dependent lymphoid priming in hematopoietic stem cells. Immunity 2009;30(4):493–507 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yang GX, Lian ZX, Chuang YH, Shu SA, Moritoki Y, Lan R, et al. Generation of functionally distinct B lymphocytes from common myeloid progenitors. Clin Exp Immunol 2007;150(2):349–57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ye M, Iwasaki H, Laiosa CV, Stadtfeld M, Xie H, Heck S, et al. Hematopoietic stem cells expressing the myeloid lysozyme gene retain long-term, multilineage repopulation potential. Immunity 2003;19(5):689–99 [DOI] [PubMed] [Google Scholar]
- 14.Iwasaki H, Somoza C, Shigematsu H, Duprez EA, Iwasaki-Arai J, Mizuno S, et al. Distinctive and indispensable roles of PU.1 in maintenance of hematopoietic stem cells and their differentiation. Blood 2005;106(5): 1590–600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nutt SL, Metcalf D, D’Amico A, Polli M, Wu L. Dynamic regulation of PU.1 expression in multipotent hematopoietic progenitors. J Exp Med 2005;201(2):221–31 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wolfler A, Danen-van Oorschot AA, Haanstra JR, Valkhof M, Bodner C, Vroegindeweij E, et al. Lineage-instructive function of C/EBPalpha in multipotent hematopoietic cells and early thymic progenitors. Blood 2010;116(20):4116–25 [DOI] [PubMed] [Google Scholar]
- 17.Garcia-Montoya IA, Cendon TS, Arevalo-Gallegos S, Rascon-Cruz Q. Lactoferrin a multiple bioactive protein: an overview. Biochim Biophys Acta 2012;1820(3):226–36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Arnold RR, Cole MF, McGhee JR. A bactericidal effect for human lactoferrin. Science 1977;197(4300):263–5 [DOI] [PubMed] [Google Scholar]
- 19.Ward PP, Mendoza-Meneses M, Park PW, Conneely OM. Stimulus-dependent impairment of the neutrophil oxidative burst response in lactoferrin-deficient mice. Am J Pathol 2008;172(4):1019–29 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sawatzki G, Rich IN. Lactoferrin stimulates colony stimulating factor production in vitro and in vivo. Blood Cells 1989;15(2):371–85 [PubMed] [Google Scholar]
- 21.Broxmeyer HE, Smithyman A, Eger RR, Meyers PA, de Sousa M. Identification of lactoferrin as the granulocyte-derived inhibitor of colony-stimulating activity production. J Exp Med 1978;148(4):1052–67 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Al-Shaibi N, Ghosh SK. A novel cell-surface protein CSP82 on bone marrow stem cells and a cytosolic phosphoprotein DP58 (ankyrinRD 34B) are involved in promyeloid progenitor induction. Cell Immunol 2009; 258(2):172–80 [DOI] [PubMed] [Google Scholar]
- 23.Luche H, Weber O, Nageswara Rao T, Blum C, Fehling HJ. Faithful activation of an extra-bright red fluorescent protein in “knock-in” Cre-reporter mice ideally suited for lineage tracing studies. Eur J Immunol 2007;37(1): 43–53 [DOI] [PubMed] [Google Scholar]
- 24.Pronk CJ, Rossi DJ, Mansson R, Attema JL, Norddahl GL, Chan CK, et al. Elucidation of the phenotypic, functional, and molecular topography of a myeloerythroid progenitor cell hierarchy. Cell Stem Cell 2007;1(4):428–42 [DOI] [PubMed] [Google Scholar]
- 25.Boyer SW, Schroeder AV, Smith-Berdan S, Forsberg EC. All hematopoietic cells develop from hematopoietic stem cells through Flk2/Flt3-positive progenitor cells. Cell Stem Cell 2011;9(1):64–73 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Benz C, Martins VC, Radtke F, Bleul CC. The stream of precursors that colonizes the thymus proceeds selectively through the early T lineage precursor stage of T cell development. J Exp Med 2008;205(5):1187–99 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jakubzick C, Bogunovic M, Bonito AJ, Kuan EL, Merad M, Randolph GJ. Lymph-migrating, tissue-derived dendritic cells are minor constituents within steady-state lymph nodes. J Exp Med 2008;205(12):2839–50 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Socolovsky M, Nam H, Fleming MD, Haase VH, Brugnara C, Lodish HF. Ineffective erythropoiesis in Stat5a(−/−)5b(−/−) mice due to decreased survival of early erythroblasts. Blood 2001;98(12):3261–73 [DOI] [PubMed] [Google Scholar]
- 29.Akashi K, He X, Chen J, Iwasaki H, Niu C, Steenhard B, et al. Transcriptional accessibility for genes of multiple tissues and hematopoietic lineages is hierarchically controlled during early hematopoiesis. Blood 2003;101(2):383–9 [DOI] [PubMed] [Google Scholar]
- 30.Iwasaki H, Akashi K. Myeloid lineage commitment from the hematopoietic stem cell. Immunity 2007;26(6):726–40 [DOI] [PubMed] [Google Scholar]
- 31.Auffray C, Sieweke MH, Geissmann F. Blood monocytes: development, heterogeneity, and relationship with dendritic cells. Annu Rev Immunol 2009;27:669–92 [DOI] [PubMed] [Google Scholar]
- 32.Yona S, Kim KW, Wolf Y, Mildner A, Varol D, Breker M, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity 2013;38(1):79–91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Schlenner SM, Rodewald HR. Early T cell development and the pitfalls of potential. Trends Immunol 2010;31(8):303–10 [DOI] [PubMed] [Google Scholar]
- 34.Gorgens A, Radtke S, Mollmann M, Cross M, Durig J, Horn PA, et al. Revision of the human hematopoietic tree: granulocyte subtypes derive from distinct hematopoietic lineages. Cell Rep 2013;3(5):1539–52 [DOI] [PubMed] [Google Scholar]
- 35.Leary AG, Ogawa M. Identification of pure and mixed basophil colonies in culture of human peripheral blood and marrow cells. Blood 1984;64(1):78–83 [PubMed] [Google Scholar]
- 36.Nakahata T, Spicer SS, Ogawa M. Clonal origin of human erythro eosinophilic colonies in culture. Blood 1982;59(4):857–64 [PubMed] [Google Scholar]
- 37.Tsai S, Bartelmez S, Sitnicka E, Collins S. Lymphohematopoietic progenitors immortalized by a retroviral vector harboring a dominant-negative retinoic acid receptor can recapitulate lymphoid, myeloid, and erythroid development. Genes Dev 1994;8(23):2831–41 [DOI] [PubMed] [Google Scholar]
- 38.Lawson ND, Berliner N. Representational difference analysis of a committed myeloid progenitor cell line reveals evidence for bilineage potential. Proc Natl Acad Sci USA 1998;95(17):10129–33 [DOI] [PMC free article] [PubMed] [Google Scholar]