

Abstract
WC 44 and WC 10 are phenylpiperazines with low (23 fold) to moderate (42 fold) selectivity for dopamine D3 receptors (D3Rs) over D2Rs, respectively. WC 44 is a full D3R agonist in the forskolin-stimulated adenylyl cyclase (AC) assay, whereas WC 10 has little efficacy. In contrast to their opposite effects in the AC assay, these drugs often produce similar behavioral effects, suggesting that the AC assay does not predict the efficacy of these drugs in vivo. Here we examined whether Fos protein expression induced by these drugs would be more consistent with their behavioral effects in vivo. Rats received either vehicle, WC 10 (5.6 mg/kg, i.p.), WC 44 (10.0 mg/kg, i.p), cocaine (10.0 mg/kg, i.p.), or cocaine with WC 10 (5.6 mg/kg, i.p.) or with WC 44 (10.0 mg/kg, i.p). Locomotion was monitored for 90 min and the brains were harvested for immunohistochemistry. Both WC 10 and WC 44 decreased spontaneous and cocaine-induced locomotion. Both compounds also increased Fos expression relative to saline in the dorsal striatum and nucleus accumbens core and shell, and relative to cocaine alone in the nucleus accumbens shell. The findings suggest that even though these compounds have different efficacy in the AC bioassy, they produce similar brain activation and attenuation of cocaine hyperlocomotion. Together with our previous research demonstrating that these compounds down-shift the cocaine self-administration dose-effect function, the findings support the idea that D3R-selective compounds may be useful for cocaine dependence medications development.
Keywords: c-fos, immediate early gene, striatum, nucleus accumbens
INTRODUCTION
There are five subtypes of dopamine receptors grouped into two families, the D1-like family (D1 and D5) and the D2-like family (D2, D3, and D4) (Missale, 1998; Vallone, 2000). The dopamine D3 receptor (D3R) subtype has garnered attention as a target for development of new therapeutic agents to treat psychostimulant addiction, as well as Parkinson's disease, LDOPA-induced dyskinesia, and schizophrenia (Joyce and Millan, 2005; Kumar et al., 2009; Le Foll et al., 2005; Luedtke and Mach 2003; Heidbreder and Newman 2010; Levant, 1997; Newman et al., 2005). This is partially due to the fact that up-regulation of D3Rs occurs in response to administration of many drugs of abuse, including cocaine (Le Foll et al., 2003; Spangler et al., 2003; Vengeliene et al., 2006). Human cocaine overdose fatalities exhibit increases in D3R binding and mRNA expression (Segal et al., 1997; Staley and Mash, 1996). In rodents, increases in motivation to seek cocaine correspond to up-regulation of D3Rs during the course of abstinence from cocaine (Conrad et al., 2010; Neisewander et al., 2004).
Another important feature of D3Rs is that they show differential expression across brain regions. D3Rs have higher expression in Isles of Calleja in the olfactory tubercle and the nucleus accumbens (collectively referred to as the ventral striatum) relative to other dopamine terminals (Sokoloff et al., 1990). The nucleus accumbens is thought to play a critical role in the development of addiction especially during the early stages, whereas the dorsal striatum may be critically involved during the later stages of addiction (Belin and Everitt, 2008; Di Chiara and Bassareo, 2007; Di Ciano et al., 2008; Ito et al., 2002; Koob and Volkow, 2010; Porrino et al. 2007). For instance, functional activation observed in rhesus monkeys using 2-[14C]deoxyglucose imaging shifts from the ventral striatum early during cocaine self-administration to the dorsal striatum with chronic cocaine self-administration (Porrino et al., 2007). The nucleus accumbens is critical for reward learning (Apicella et al. 1991; Ikemoto, 2007; Schultz et al., 2000), and is specifically involved in translating the value of a reward into appropriate goal-directed behaviors (Roesch et al., 2009).
We have recently developed 2-methoxy substituted phenylpiperazine compounds with varying degrees of selectivity and intrinsic activity in the forskolin-stimulated adenylyl cyclase assay including: WC 10, a low efficacy partial agonist (which can function like an antagonist) with 42-fold selectivity for D3Rs vs. D2Rs; and WC 44, a full agonist with 23-fold selectivity for D3Rs vs. D2Rs (Chu et al., 2005; Kumar et al., 2009; Xu et al., 2009). The structures of these compounds are shown in Figure 1. WC 10 has a log P value of 3.09 and WC 44 has a log P value of 2.94 (Chu et al., 2005), which predicts that they should readily cross the blood-brain barrier (Dishino et al., 1983). These compounds represent the first series in an ongoing effort to develop useful D3R-selective drugs with varying intrinsic activity.
Figure 1.
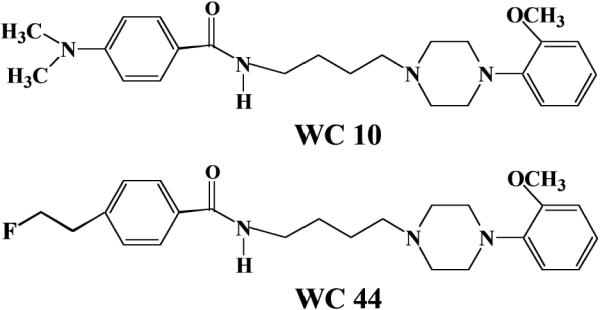
Chemical structures of WC 10 and WC 44.
Surprisingly, our research on behavioral effects of WC 10 and WC 44 has revealed only subtle differences in their effects despite their opposite effects in the forskolin-stimulated adenylyl cyclase assay. We have found that both compounds have therapeutic potential for the treatment of L-DOPA-induced dyskinesia (Kumar et al., 2009) and are effective in attenuating deficits in pre-pulse inhibition of acoustic startle, a model of sensory integration deficits observed in schizophrenia (Weber et al., 2009). We have also found that WC 10 and WC 44 decrease cocaine and sucrose intake and that WC 10 increases the latency to perform an operant response for both reinforcers, while WC 44 selectively increases the latency to respond for cocaine only (Cheung et al., 2012). These results suggest that both WC compounds may reduce the motivation to seek cocaine and that the forskolin-stimulated adenylyl cyclase assay may not be predictive of the effects of the drugs in vivo (Cheung et al., 2012).
To further examine the potential differences in the mechanism of action of these drugs, the present study used Fos protein expression as a marker of the brain functional activation induced by WC 10 and WC 44 given either alone or with cocaine. Fos is the protein product of the immediately early gene c-fos, and its expression has been used as a tool for mapping functional activity in the brain (Chaudhuri 1997; Herdegen and Leah 1998). We measured Fos in the nucleus accumbens core (NAcC) and shell (NAcS), dorsal striatum (dSt), and anterior cingulate cortex (ACC) with the latter included as a control region where we did not expect the compounds to alter Fos expression. Additionally, we examined the effects of WC 10 and WC 44 on spontaneous and cocaine-induced locomotor activity.
MATERIALS AND METHODS
Animals
Adult male Sprague-Dawley rats, weighing 250 ± 25 g at the beginning of the experiment, were housed individually in a climate-controlled colony room with a reversed 12-h light/dark cycle. Housing conditions were in accordance with the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources on Life Sciences, National Research Council, 1996). Animals were acclimated to handling daily for 3 days immediately prior to starting the experiments.
Drugs
Cocaine hydrochloride (RTI International, Research Triangle Park, NC) was dissolved in saline. WC 10 and WC 44 were dissolved a small amount of 1.0 N hydrochloric acid and then brought to proper concentration with 5% dimethyl sulfoxide in saline (0.9% NaCl). An identical solution without drug was used as vehicle. All injections were given intraperitoneally (i.p.) at a volume of 1.0 ml/kg.
Behavioral Testing
Animals were habituated to the procedure by placing them into the test chambers for 30 min after receiving an acute injection of saline on each of 3 consecutive days immediately preceding the test day. The test chambers were clear Plexiglas chambers (48.3 × 26.7 × 15.6 cm) with a thin layer of pine bedding on the floor and stainless steel wire lids.
Rats were randomly assigned to 1 of 4 test conditions for each compound, using a between-subject design. For WC 10, the four conditions were tested in two experiments that were conducted with separate cohorts of rats run at different times. Experiment 1a tested the effect of vehicle vs. WC 10 (5.6 mg/kg, i.p., n=10 for both groups). Experiment 1b tested the effect of cocaine (10.0 mg/kg, i.p.) alone or given with WC 10 (5.6 mg/kg, i.p., n=8 for both groups). Experiment 2 examined the effects of WC 44 on spontaneous and cocaine-induced locomotion using a single cohort of rats in a 2×2 factorial design with the resulting four groups (n=10 per group): vehicle (i.e., vehicle + saline), WC 44 (i.e., 10.0 mg/kg WC 44 + saline), Coc (i.e., vehicle + 10.0 mg/kg cocaine) and Coc+WC 44 (10.0 mg/kg of both WC 44 and cocaine). The doses of WC 10 and WC 44 were chosen based on our previous findings that they flatten the dose-effect curve for cocaine intake (Cheung et al., 2012). All drug injections were given 5 min before the start of the test session. Locomotor activity was measured using a computer-automated system (Cleversys, Inc., Restyon,VA) that continuously tracked the animals throughout the 90-min test period. The conclusion of the 90-min test session coincides with previous observations of peak Fos expression after an acute stimulus (Herdegen and Leah, 1998; Nestler, 2004).
Tissue Preparation
Immediately after the test for locomotion, rats were deeply anesthetized with 50–100 mg/kg, i.p., Euthasol and intracardially perfused with ice cold phosphate buffered saline (PBS, 0.1M, pH 7.4) followed by ice cold paraformaldehyde (PFA, 4% in 0.1M PBS, pH 7.4). Brains were then removed and allowed to post-fix in PFA for 24 h at 4°C. Next, brains were cryoprotected in a 10% sucrose solution dissolved in 0.1M phosphate-buffered saline (PBS) for 24 h followed by 30% sucrose dissolved in 0.1M PBS for 48 h. A freezing microtome was then utilized to cut sections (40 μm) approximately ± 0.2 mm from our target of +1.6 mm from bregma (Paxinos and Watson, 1998). Serial sections were stored at −20°C in cryoprotectant solution consisting of 30% ethylene glycol, 30% sucrose, 10% polyvinyl pyrrolidone, and 0.1M PBS until labeled using immunohistochemistry.
Immunohistochemistry
Sections from rats in each group were processed simultaneously for Fos protein expression using methods adapted from Kufalh et al. (2009). Sections were first thawed for 20 min at room temperature followed by extensive washes in 0.1 M phosphate buffer (PB, pH 7.3, 3 times for 30-min). This was followed by incubation in hydrogen peroxide (3% for 30 min) and then another incubation in normal goat serum (3% NGS, Vector Labs, Burlingame, CA). Sections were then incubated in primary antibody (rabbit polyclonal anti-Fos antibody, Santa Cruz Biotech., Santa Cruz, CA: sc-52; 1:5000) dissolved in phosphate buffered goat serum (0.1% bovine serum albumin, 0.2% Triton X-100, 2% NGS and 0.1M PB 0.1M) for 72 h at 4°C with agitation. The tissue was then washed 3 times for 10 min in 0.1M PB and allowed to incubate for 1 h in secondary antibody (biotinylated goat anti-rabbit IgG antibody, Vector Laboratories, Burlingame, CA; 1:500), dissolved in 0.1 % bovine serum albumin (Sigma, St. Louis, MO), 0.2% Triton-X (Sigma, St. Louis, MO), 2% NGS and 0.1 M PB. This was followed by 3 washes for 10 min in 0.1M PB and subsequent incubation in extravidin peroxidase (1:1000) dissolved in 0.1 % bovine serum albumin, 0.2% Triton-X, 2% NGS and 0.1 M PB for 90 min. Once again the sections were washed 3 times for 30 min in 0.1M PB. They were then incubated in 3,3'-diaminobenzidine tetrahydrochloride (Sigma, St. Louis, MO) containing ammonium chloride (0.4%), D-Glucose (20%) and nickel ammonium sulphate (2%) for 20 min. The reaction was visualized using glucose oxidase for 10 min and immediately terminated by 2 washes for 30 min in 0.1M PB. Finally, sections were stored in 0.1M PB at 4°C until they were mounted onto slides coated with gelatin chrom-alum, dried, dehydrated, and cover-slipped.
Microscopy and Analysis
Regions sampled at +1.6 ± 0.2 mm from bregma (Paxinos and Watson, 1998) were the nucleus accumbens core (NAcC) and shell (NAcS), the dorsal striatum (dSt), and anterior cingulate cortex (ACC, Figure 2). The ACC served as a control region, as we did not expect Fos expression to be affected in this region by changes in D3R transduction (Southam et al. 2007). These regions were sampled bilaterally in 2 different brain sections for each subject at 20x magnification using a Nikon Eclipse E600 microscope (Nikon Instruments, Melville, NY, USA). Digital images were stored as .tiff files for later analysis. The photomicrographs were captured by individuals blind to group assignment. The software Image J (public domain imaging software developed at the National Institutes of Health) was used to analysize the unmodified images. Qualitative judgment was used to set a threshold (in Image J) for detecting Fos based on size and darkness. Quantification was then performed automatically using the Image J software. Fos-positive nuclei in each region were counted by individuals blind to group assignment. For each region in each subject, the counts were averaged across samples (i.e., 1 sample per hemisphere per duplicate sections), and these averages were analyzed statistically as described below.
Figure 2.

Diagram indicating the regions sampled: dSt = dorsal striatum; NAcC = nucleus accumbens core; NAcS = nucleus accumbens shell; ACC = cingulate cortex. Drawing is based on Paxinos and Watson (1998).
Statistical Analysis
For WC 10, locomotion and Fos-labeled nuclei were analyzed separately for Experiments 1a and 1b (vehicle vs. WC 10 and cocaine vs. cocaine + WC 10, respectively) because these experiments were conducted at different times. Independent sample t-tests were used to test differences between groups. For WC 44 (experiment 2), these dependent variables were analyzed using a 2 × 2 factorial ANOVA (vehicle/WC 44 × vehicle/cocaine). Newman-Keuls tests were used for post-hoc analyses.
RESULTS
Effects of WC 10
WC 10 (5.6 mg/kg, i.p.) decreased spontaneous and cocaine-induced locomotor activity (Figure 3) and increased Fos expression in all brain regions examined except the ACC (Figure 4A). Independent sample t-tests comparing the effect of vehicle vs. WC 10 alone in Experiment 1a revealed that WC 10 reduced spontaneous locomotor activity (t(18)= 3.87, p<.01) and increased Fos expression in the dSt, NAcC and NAcS (t(18)=4.57, p<.01, t(18)=7.30, p<.01, and t(18)=9.34, p<.01 respectively). Fos expression in the ACC did not differ between groups (t(18)=0.77, n.s.), demonstrating that the effect of WC 10 was region-specific. Similarly, comparing the effect of cocaine (10 mg/kg, i.p.) alone vs. cocaine + WC 10 (5.6 mg/kg, i.p.) in Experiment 1b revealed that WC 10 reduced cocaine-induced locomotor activity (t(14)= 2.92, p<.02). WC 10 further increased Fos expression relative to cocaine alone in the NAcS (t(14)=2.91, p<.02; Figure 4, B). Fos expression in the other 3 regions did not differ between groups.
Figure 3.
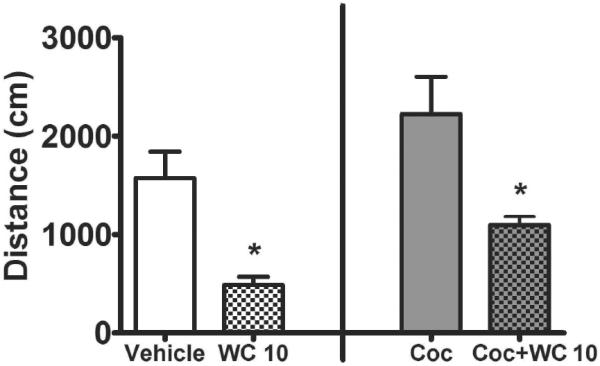
Mean distance traveled (cm + SEM) during a 90-min tracking session for rats following acute administration of either vehicle or WC 10 (5.6 mg/kg, i.p.) in experiment 1a (left; n = 10/group), or following administration of cocaine (Coc, 10 mg/kg, IP) or cocaine + WC 10 (5.6 mg/kg, i.p.) in experiment 1b (right; n = 8/group). Injections occurred 5 min prior to the beginning of the session. * indicates a difference from vehicle control (p<0.01).
Figure 4.

Mean number of Fos stained nuclei (+ SEM) across regions: dSt = dorsal striatum; NAcC = nucleus accumbens core; NAcS = nucleus accumbens shell; ACC = anterior cingulate cortex. A: Rats were given acute administration of either vehicle or WC 10 (5.6 mg/kg, i.p.) 95 min prior to perfusion (n = 10/group). B: Rats were given acute administration of either cocaine (Coc, 10.0 mg/kg, i.p.) or cocaine + WC 10 (5.6 mg/kg, i.p.) 95 min prior to perfusion (n = 8/group). * indicates a difference from respective vehicle (A) or Coc (B) control group (p<0.05).
Effects of WC 44
There was an interaction between cocaine and the full agonist WC 44 whereby cocaine increased locomotion but only in rats pretreated with vehicle and not those pretreated with WC 44, and WC 44 reduced locomotion regardless of vehicle versus cocaine pretreatment (Figure 5). Due to a technical error, locomotor activity data from 2 rats in each group were lost. The final group size for the behavioral measure was therefore 8 rats per group. The ANOVA of distance travelled revealed a significant interaction between WC 44 (10 mg/kg, i.p.) and cocaine (10 mg/kg, i.p.; F(1,28)=6.11, p<0.05). Subsequent post-hoc analysis indicated that animals given cocaine alone exhibited an increase in locomotion relative to all other groups (p<0.05, Newman-Keuls tests). WC 44 decreased locomotor activity relative to vehicle-treated controls regardless of whether animals received co-treatment with vehicle or cocaine (p<0.05, Newman-Keuls tests).
Figure 5.
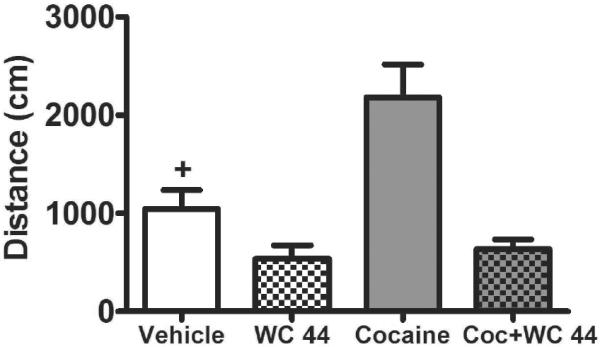
Mean distance traveled (cm + SEM) during a 90-min tracking session following acute administration of either vehicle, WC 44 (10 mg/kg, i.p.), cocaine (Coc, 10 mg/kg, i.p.), or cocaine with WC 44 (both drug doses were 10 mg/kg, i.p.). Rats (n=8/group) were given the drug injections 5 min prior to the beginning of the tracking session. + indicates a difference from all other groups (Newman-Keuls, p<0.05).
Region-specific increases in Fos protein expression by both cocaine and WC 44 were observed in the striatum but not the ACC (Figure 6). For the dSt, ANOVA found a significant interaction between the effect of WC 44 and cocaine (F(1,36)=4.97, p<0.05). Post-hoc analysis revealed that all three experimental groups showed significant increases in Fos expression relative to vehicle in the dSt (p<0.05, Newman-Keuls tests). For the NAcC, ANOVA found significant main effects of WC 44 (F(1,36)=10.07, p<0.01) and cocaine (F(1,36)=4.62, p<0.05), indicating that both drugs increased Fos expression; the WC 44×cocaine interaction was not significant. For the NAcS, ANOVA found a main effect of WC 44 (F(1,36)=26.04, p<0.01), with increased Fos expression in animals treated with WC 44 regardless of co-treatment with vehicle vs. cocaine. Although ANOVA failed to detect a main effect of cocaine (F(1,36)=1.98, n.s.) or a WC 44×cocaine interaction in the NAcS (F(1,36)=2.14, n.s.) a previous study has found that cocaine alone increases Fos expression in this region (Uslander, et al. 2001). Thus it is worth noting that a two-tailed t-test suggested that Fos expression was indeed increased in the cocaine alone group compared with vehicle (t(18)=3.86, p<.01). For the ACC, there were neither main effects nor an interaction, suggesting that Fos expression in this area was not affected by any of the treatments.
Figure 6.
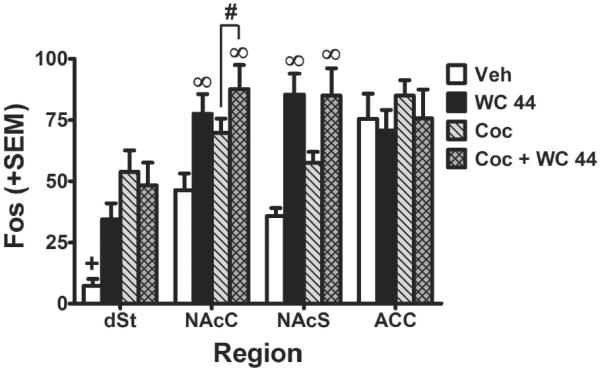
Mean number of Fos stained nuclei (+ SEM) across counting regions: dSt = dorsal striatum; NAcC = nucleus accumbens core; NAcS = nucleus accumbens shell; ACC = cingulate cortex. Rats (n=10 / group) were given an acute injection of vehicle, WC 44 (10 mg/kg, i.p.), cocaine (Coc, 10 mg/kg, i.p.) or cocaine with WC 44 (10 mg/kg, i.p. for both drugs). + indicates a difference from all groups (Newman-Keuls, p<0.05). ∞ indicates a main effect of WC 44 (ANOVA, p<0.01). # indicates main effect of cocaine (ANOVA, p<0.05).
DISCUSSION
The purpose of the present study was to determine whether WC 10 and WC 44 produce differential changes in functional neuronal activity as predicted from their differential effects on D3R transduction through the adenylate cyclase pathway measured in a D3R cell line in vitro (Chu et al. 2005). Functional activity in this study was measured by Fos immunoreactivity in striatal brain regions that are associated with the motor stimulant effects of cocaine. In contrast to that predicted by their in vitro effects on adenylate cyclase, the pattern of Fos expression in dorsal and ventral striatal subregions was similar across the two compounds when given either alone or with cocaine. The present results thus parallel previous findings that several behavioral effects of WC 10 and WC 44 are similar, such as the ability to decrease spontaneous and cocaine-induced locomotion, and to reduce operant responding for cocaine and sucrose (Cheung et al., 2012; Kumar et al., 2009).
The decrease in spontaneous and cocaine-induced locomotion observed with acute administration of WC 10 (5.6 mg/kg, i.p.) and WC 44 (10 mg/kg, i.p.) suggests that even though these drugs suppress cocaine self-administration, they also produce motor side-effects (Cheung et al., 2012; Kumar et al., 2009). This differs from the D3R-selective antagonist SB-277011-A, which does not alter psychostimulant-induced locomotor activity at doses up to 40 mg/kg (Southam et al. 2007) but is effective in reducing cocaine abuse-related behaviors (Di Ciano et al., 2003; Le Foll et al., 2002; Vorel et al., 2002). A likely explanation for the differences is that WC 10 and WC 44, at the doses tested here, may also block D2Rs since both drugs have lower selectivity for D3Rs vs. D2Rs than SB-277011-A (43 fold for WC 10, 23 fold for WC 44, 100 fold for SB-277011-A) (Kumar et al. 2009; Reavill et al. 2000) and act as low efficacy partial agonists or antagonists at the D2R according to the adenylyl cyclase assay (Kumar et al. 2009). Similar to WC 10, other D3R-preferring antagonists, such as U99194A, attenuate locomotor activity at high doses (Rodriguez-Arias et al., 1999; Carr et al. 2002). In contrast, non-selective and D3R-preferring agonists have biphasic effects on locomotion where low doses decrease locomotion and higher doses first decrease then increase locomotion (Fuchs et al., 2002; Khroyan et al., 1995; Khroyan et al., 1999; Jackson et al. 1995; Li et al., 2010). It is controversial whether or not the hypolocomotion produced by D3R-preferring agonists is due to a post-synaptic D3R effect or to preferential stimulation of D2/D3 autoreceptors resulting in a decrease in dopamine release (Ackerman and White, 1990; Pierce et al., 1995). Thus, either or both mechanisms may have contributed to the effects of WC 44 and WC 10 on locomotion in the present study.
Both WC compounds, when given alone, increased Fos expression relative to vehicle in the NAcS, NAcC, and dSt, but not in the ACC. The regions affected by the drugs correspond to those that have moderate to high D3R expression (Diaz et al. 2000; Sokoloff et al., 1990; Surmeier et al., 1996; Xu et al. 2010). A significant enhancement of cocaine-induced Fos expression was also observed with both WC 10 and WC 44 in the NAcS, which has the highest density of D3Rs of all regions examined. Our results are consistent with a previous report that the D3R-preferring antagonist U99194A increased Fos expression in the NAcS, the NAcC, and the dSt (Carr, 2002). However, another study found that the D3R-preferring antagonist GR103691 failed to alter c-fos mRNA levels in either the dorsal or ventral striatum (Hurley et al. 1996). With the more the selective D3R antagonist SB-277011-A, Southam and colleagues (2007) found that Fos expression is selectively increased in the NAcS and NAcC, but not in the dSt. The increase in dSt Fos expression with the WC compounds in contrast to SB-277011 suggests that this effect involves an action at D2Rs, which also likely accounts for the motor side-effects of the WC compounds in comparison to SB-277011-A. Indeed, D2R antagonists such as haloperidol (> 15 fold selectivity for D2 vs. D3) are capable of inducing Fos in the medial and lateral regions of the dSt, as well as the NAc (Robertson, 1994).
The effects of D3R-preferring agonists on c-fos induction have been mixed. For example, the D3/D4R agonist cis-8-OH-PBZI (90-fold D3R selectivity vs. D2R; 10-fold selectivity vs. D4) increases Fos expression in vivo in the rat dorsal and ventral striatum and medial prefrontal cortex (Scheideler et al. 1997). The increase in Fos in the medial prefrontal cortex suggests an action of cis-8-OH-PBZI at D4Rs since there are very few D3Rs in this region. The D3R-preferring agonists 7-OH-DPAT (16-fold D3R selectivity vs. D2R) and PD 128907 (56-fold D3R selectivity vs. D2R) both increase c-fos mRNA expression in in vitro cultures of dorsal and ventral striatal neurons (Morris et al. 2000). However, several other studies report that in vivo administration of D3R-preferring agonists does not increase c-fos mRNA in the dorsal striatum or nucleus accumbens (Guo et al., 1995; Ishibashi et al., 2002; Ridray et al., 1998) but blocks Fos expression induced by haloperidol or by the atypical antipsychotic clozapine in both regions (Guo et al., 1995; Vahid-Ansari and Robertson, 1996). Furthermore, clozapine-induced Fos expression in the nucleus accumbens can be blocked by a low dose of 7-OH-DPAT that is thought to act mainly at D3Rs (Vahid-Ansari and Robertson, 1996). These results suggest that D2/D3R agonists do not increase Fos expression in the nucleus accumbens and can attenuate the induction of Fos expression by other drugs in this area. This differs from the present finding that WC 44 increases Fos expression in both nucleus accumbens and dorsal striatum, which is more similar to the Fos expression pattern induced by D2/D3R antagonists.
Enhancement of cocaine-induced Fos expression has been associated with lower D3R activity. For instance, Zhang et al. (2004) found that mutant mice lacking D3Rs showed increased cocaine-induced Fos expression in the NAc and dSt. Furthermore, Carta et al. (2000) found that D3R knockout mice showed enhanced behavioral responses and increased Fos expression in the dorsal and ventral striatum in response to acute administration of cocaine compared to wildtype mice. In contrast to D3R knockout mice, Welter et al. (2007) reported that D2R knockout mice show a lack of cocaine-induced c-fos response and decreased cocaine-induced motor activity. These findings suggest that cocaine-induced Fos expression involves stimulation of D2Rs whereas stimulation of D3Rs may inhibit mechanisms involved in cocaine-induced Fos expression. Thus, the increased Fos expression in the NAc in rats given cocaine plus WC 10 or WC 44 relative to those given cocaine alone may be due to antagonism of D3Rs. Importantly, the similar patterns of Fos expression induced by WC 44 and WC 10 suggests that the action of these drug in vivo is likely the same in contrast to their characterization as an agonist and an antagonist, respectively, in D3R-expressing cell in vitro.
There are several possible explanations for why WC 44 and WC 10 produced similar effects in vivo despite their different efficacies in modulating adenylyl cyclase in vitro. First, WC 44 may become defluorinated in vivo and the resulting metabolite may act as an antagonist at the D3R (Tu et al. 2011). Second, the drugs may produce opposite effects in regions other than the ventral striatum. D3Rs are found in high levels in the cerebellum (Stanwood et al., 2000) where local infusion of the D3R-preferring agonist 7-OH-DPAT reduces locomotor activity, an effect that is reversed by the D3R antagonist S33084 (Kolasiewicz, 2008), which has 100-fold D3R selectivity over D2R (Millan, 2000). Consistent with this idea, systemic treatments with 7-OH-DPAT have been found to increase c-fos mRNA in the rat cerebellum, whereas changes in the dorsal striatum or the nucleus accumbens were negligible (Ishibashi, 2002). Third, the D3R-related behavioral effects of WC 10 and WC 44 in vivo may not be mediated via an adenylyl cyclase pathway. D3R-selective phenylpiperazines exhibit functional selectivity such that the intrinsic efficacy differs for the adenylyl cyclase versus the mitogenic signaling pathways (Taylor et al., 2010). Therefore, the intrinsic activity of WC 10 and WC 44 may differ for adenylyl cyclase inhibition, but may be similar for another pathway. Fourth, some of the in vivo effects of WC 10 and WC 44 may be due to an action at the 5-HT1AR since both compounds have affinity for these receptors (Chu et al., 2005). 5-HT1AR agonists do not increase Fos expression in the dorsal striatum or the nucleus accumbens, but attenuate the induction of Fos expression in both areas (Ohno et al., 2008, 2009; Tilakaratne and Friedman, 1996; Tremblay et al., 1998; Tsuchida et al., 2009). In contrast, the 5-HT1AR antagonist WAY-100635, which also acts as an agonist at D4Rs (Chemel et al., 2006; Marona-Lewicka and Nichols, 2009), increases Fos expression in the dorsal striatum and nucleus accumbens (Jongsma et al., 2002). WAY-100635 has also been reported to attenuate cocaine-induced locomotion (Carey et al., 2000, 2001; Müller et al., 2002) and cocaine seeking behavior (Burmeister et al., 2004; Schenk, 2000). It is therefore possible that blockade of 5-HT1ARs contributes to some of the effects of WC 10 and WC 44.
Although we have highlighted the similarities in Fos expression and behavioral effects of WC 10 and WC 44, it is important to note that some differences exist including: 1) their ability to reverse prepulse inhibition (PPI) of startle deficits induced by the non-selective dopamine agonist apomorphine, for which only WC 10 is effective even though both drugs reverse PPI deficits produced by the D3R-preferring agonist pramipexole (Weber et al. 2009), and 2) a more selective effect of WC 44 to increase latency to respond for cocaine but not for sucrose, whereas WC 10 increases latency to respond for both reinforcers (Cheung et al. 2012). An important goal of future research is to determine the dopamine signaling pathways involved in the above behavioral effects in an effort to establish in vitro assays with validity for predicting potential therapeutic and side effects of novel D3R drugs.
In conclusion, the results provide further information regarding effects of the D3R-preferring phenylpiperazines WC 10 and WC 44 on cocaine-induced behaviors. Moreover, the similarity in the pattern of Fos immunoreactivity and changes in spontaneous and cocaine-induced locomotion following WC 10 and WC 44 administration suggests these compounds may have the same action in vivo despite being classified as an antagonist and an agonist, respectively, in the in vitro adenlyate cyclase inhibition assay (Chu et al. 2005; Kumar et la. 2009). Further research aimed at understanding the relationship between efficacy and selectivity of drugs that act at D2-like receptors is essential for developing medications for psychostimulant dependence. At present, the literature suggests that selective D3R antagonists may be useful for promoting abstinence without producing unwanted side-effects; however, they may not be effective in decreasing cocaine intake should relapse occur (Heidbreder and Newman, 2010). Less selective D3/D2R partial agonists and antagonists appear to be more effective in reducing psychostimulant intake in preclinical self-administration models than selective D3R antagonists, but they also produce more side-effects (Millan et al., 2004). It is possible that some degree of co-occupancy of D2 and D3Rs is needed to reduce psychostimulant intake and perhaps what is needed are compounds with a ratio of D3R:D2R occupancy levels that can inhibit cocaine SA while minimizing the potential side effects.
Acknowledgments
Grants: This work is supported by NIDA grant DA023957.
REFERENCES
- Ackerman JM, White FJ. A 10 somatodendritic dopamine autoreceptor sensitivity following withdrawal from repeated cocaine treatment. Neuroscie Lett. 1990;117:181–187. doi: 10.1016/0304-3940(90)90141-u. [DOI] [PubMed] [Google Scholar]
- Apicella P, Ljungberg T, Scarnarti E, Schultz W. Responses to reward in monkey dorsal and ventral striatum. Exp Brain Res. 1991;85:491–500. doi: 10.1007/BF00231732. [DOI] [PubMed] [Google Scholar]
- Belin D, Everitt BJ. Cocaine seeking habits depend upon dopamine-dependent serial connectivity linking the ventral with the dorsal striatum. Neuron. 2008;57:432–441. doi: 10.1016/j.neuron.2007.12.019. [DOI] [PubMed] [Google Scholar]
- Burmeister JJ, Lungren EM, Kirschner KF, Neisewander JL. Differential roles of 5-HT receptor subtypes in cue and cocaine reinstatement of cocaine-seeking behavior in rats. Neuropsychopharmacology. 2004;29:660–8. doi: 10.1038/sj.npp.1300346. [DOI] [PubMed] [Google Scholar]
- Carey R, Damianopoulos E, DePalma G. The 5-HT1A antagonist WAY 100635 can block the low-dose locomotor stimulant effects of cocaine. Brain Res. 2000;862:242–6. doi: 10.1016/s0006-8993(00)02084-9. [DOI] [PubMed] [Google Scholar]
- Carey RJ, DePalma G, Damianopoulos E. Cocaine and serotonin: a role for the 5-HT1A receptor site in the mediation of cocaine stimulant effects. Behav Brain Res. 2001;126:127–33. doi: 10.1016/s0166-4328(01)00253-4. [DOI] [PubMed] [Google Scholar]
- Carr KD, Yamamoto N, Omura M, Cabeza de Vaca S, Krahne L. Effects of the D3 dopamine receptor antagonist, U99194A, on brain stimulation and d-amphetamine reward, motor activity, and c-fos expression in ad libitum fed and food-restricted rats. Psychopharmacology. 2002;163:76–84. doi: 10.1007/s00213-002-1132-0. [DOI] [PubMed] [Google Scholar]
- Carta AR, Gerfen CR, Steiner H. Cocaine effects on gene regulation in the striatum and behavior: increased sensitivity in D3 dopamine receptor-deficient mice. Neuroreport. 2000;11:2395–2399. doi: 10.1097/00001756-200008030-00012. [DOI] [PubMed] [Google Scholar]
- Chaudhuri A. Neural activity mapping with inducible transcription factors. Neuroreport. 1997;8:iii–vii. [PubMed] [Google Scholar]
- Chemel BR, Roth BL, Armbruster B, Watts VJ, Nichols DE. WAY-100635 is a potent dopamine D4 receptor agonist. Psychopharmacology. 2006;188:244–51. doi: 10.1007/s00213-006-0490-4. [DOI] [PubMed] [Google Scholar]
- Cheung TH, Nolan BC, Hammerslag LR, Weber SM, Durbin JP, Peartree NA, Mach RH, Luedtke RR, Neisewander JL. Phenylpiperazine derivatives with selectivity for dopamine D3 receptors modulate cocaine self-administration in rats. Neuropharm. 2012;63:1346–1359. doi: 10.1016/j.neuropharm.2012.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu W, Tu Z, McElveen E, Xu J, Taylor M, Luedtke RR, Mach RH. Synthesis and in vitro binding of N-phenyl piperazine analogs as potential dopamine D3 receptor ligands. Bioorg Med Chem. 2005;13:77–87. doi: 10.1016/j.bmc.2004.09.054. [DOI] [PubMed] [Google Scholar]
- Conrad KL, Ford K, Marinelli M, Wolf ME. Dopamine receptor expression and distribution dynamically change in the rat nucleus accumbens after withdrawal from cocaine self-administration. Neurosci. 2010;169:182–194. doi: 10.1016/j.neuroscience.2010.04.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz J, Pilon C, Le Foll B, Gros C, Triller A, Schwartz JC, Sokoloff P. Dopamine D3 receptors expressed by all mesencephalic dopamine neurons. J Neurosci. 2000;20:8677–8684. doi: 10.1523/JNEUROSCI.20-23-08677.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Ciano P, Underwood RJ, Hagan JJ, Everitt BJ. Attenuation of cue-controlled cocaine seeking by a selective D3 dopamine receptor antagonist SB-277011-A. Neuropsychopharmacology. 2003;28:329–338. doi: 10.1038/sj.npp.1300148. [DOI] [PubMed] [Google Scholar]
- Di Ciano P, Robbins TW, Everitt BJ. Differential effects of nucleus accumbens core, shell, or dorsal striatal inactivations on the persistence, reacquisition, or reinstatement of responding for a drug-paired conditioned reinforcer. Neuropsychopharmacology. 2008;33:1413–1435. doi: 10.1038/sj.npp.1301522. [DOI] [PubMed] [Google Scholar]
- Di Chiara G, Bassareo V. Reward system and addiction: what dopamine does and doesn't do. Pharmacology. 2007;7:1–6. doi: 10.1016/j.coph.2006.11.003. [DOI] [PubMed] [Google Scholar]
- Dishino DD, Welch MJ, Kilbourn MR, Raichle ME. Relationship between lipophilicity and brain extraction of C-11-labeled radiopharmaceuticals. J Nucl Med. 1983;24:1030–8. [PubMed] [Google Scholar]
- Fuchs RA, Tran-Nguyen LT, Weber SM, Khroyan TV, Neisewander JL. Effects of 7-OH-DPAT on cocaine seeking behavior and on re-establishment of cocaine self-administration. 2002. [DOI] [PubMed] [Google Scholar]
- Heidbreder CA, Newman AH. Current perspectives on selective dopamine D(3) receptor antagonists as pharmacotherapeutics for addictions and related disorders. Ann N Y Acad Sci. 2010;1187:4–34. doi: 10.1111/j.1749-6632.2009.05149.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herdegen T, Leah J. Inducible and constitutive transcription factors in the mammalian nervous system: control of gene expression by Jun, Fos and Krox, and CREB/ATF proteins. Brain Res Rev. 1998;28:370–490. doi: 10.1016/s0165-0173(98)00018-6. [DOI] [PubMed] [Google Scholar]
- Hurley MJ, Stubbs CM, Jenner P, Marsden CD. Dopamine D3 receptors are not involved in the induction of c-fos mRNA by neuroleptic drugs: comparison of the dopamine D3 receptor antagonist GR103691 with typical and atypical neuroleptics. Eur J Pharmacol. 1996;318:283–293. doi: 10.1016/s0014-2999(96)00798-4. [DOI] [PubMed] [Google Scholar]
- Ikemoto S. Dopamine reward circuitry: Two projection systems from the ventral midbrain to the nucleus accumbens-olfactory tubercle complex. Brain Res Rev. 2007;56:27–78. doi: 10.1016/j.brainresrev.2007.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishibashi T, Wakabayashi J, Ohno Y. 7-Hydroxy-N,N'-di-n-propyl-2-aminotetraline, a preferential dopamine D3 agonist, induces c-fos mRNA expression in the rat cerebellum. Jpn J Pharmacol. 2002;89:309–315. doi: 10.1254/jjp.89.309. [DOI] [PubMed] [Google Scholar]
- Ito R, Dalley JW, Robbins TW, Everitt BJ. Dopamine release in the dorsal striatum during cocaine-seeking behavior under the control of a drug-associated cue. J Neurosci. 2002;22:6247–6253. doi: 10.1523/JNEUROSCI.22-14-06247.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson DM, Mohell N, Georgiev J, Bengtsson A, Larsson LG, Magnusson O, Ross SB. Time course of bromocriptine induced excitation in the rat: behavioral and biochemical studies. Naunyn-Schmiedeberg's archives of pharmacology. 1995;351:146–155. doi: 10.1007/BF00169328. [DOI] [PubMed] [Google Scholar]
- Jongsma ME, Sebens JB, Bosker FJ, Korf J. Effect of 5-HT1A receptor-mediated serotonin augmentation on Fos immunoreactivity in rat brain. Eur J Pharmacol. 2002;455:109–15. doi: 10.1016/s0014-2999(02)02583-9. [DOI] [PubMed] [Google Scholar]
- Joyce JN, Millan MJ. Dopamine D3 receptor antagonists as therapeutic agents. Drug Discovery Today. 2005;10:917–925. doi: 10.1016/S1359-6446(05)03491-4. [DOI] [PubMed] [Google Scholar]
- Khroyan TV, Baker DA, Neisewander JL. Dose-dependent effects of the D-3-preferring agonist 7-OH-DPAT on motor behaviors and place conditioning. Psychopharmacology. 1995;122:351–357. doi: 10.1007/BF02246265. [DOI] [PubMed] [Google Scholar]
- Khroyan T, Fuchs R, Beck A, Groff R, Neisewander J. Behavioral interactions produced by co-administration of 7-OH-DPAT with cocaine or apomorphine in the rat. Psychopharmacology. 1999;142:383–392. doi: 10.1007/s002130050903. [DOI] [PubMed] [Google Scholar]
- Kolasiewicz W, Maj J, Ossawska K. The involvement of cerebellar dopamine D3 receptors in locomotor activity of rats. Journal of Neural Transm. 2008;115:677–681. doi: 10.1007/s00702-007-0009-0. [DOI] [PubMed] [Google Scholar]
- Koob G, Volkow N. Neurocircuitry of Addiction. Neuropsychopharmacology. 2010;35:217–238. doi: 10.1038/npp.2009.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kufahl PR, Pentkowski NS, Heintzelman K, Neisewander JL. Cocaine-induced Fos expression is detectable in the frontal cortex and striatum of rats under isoflurane but not alpha-chloralose anesthesia: implications for FMRI. J Neurosci Methods. 2009;181:241–248. doi: 10.1016/j.jneumeth.2009.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar R, Riddle L, Griffin S, Chu W, Vangveravong S, Neisewander J, Mach R, Luedtke R. Evaluation of D2 and D3 dopamine receptor selective compounds on L-dopa-dependent abnormal involuntary movements in rats. Neuropharmacology. 2009;56:956–969. doi: 10.1016/j.neuropharm.2009.01.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Foll B, Francés H, Diaz J, Schwartz JC, Solokoff P. Role of the dopamine D3 receptor in reactivity to cocaine-associated cues in mice. Eur. J. Neurosci. 2002;15:2016–2026. doi: 10.1046/j.1460-9568.2002.02049.x. [DOI] [PubMed] [Google Scholar]
- Le Foll B, Diaz J, Sokoloff P. Increased dopamine D3 receptor expression accompanying behavioral sensitization to nicotine in rats. Synapse. 2003;47:176–183. doi: 10.1002/syn.10170. [DOI] [PubMed] [Google Scholar]
- Le Foll B, Goldberg SR, Sokoloff P. The dopamine D3 receptor and drug dependence: effects on reward or beyond? Neuropharmacology. 2005;49:525–541. doi: 10.1016/j.neuropharm.2005.04.022. [DOI] [PubMed] [Google Scholar]
- Levant B. The D3 dopamine receptor: neurobiology and potential clinical relevance. Pharmacol Rev. 1997;49:231–252. [PubMed] [Google Scholar]
- Li SM, Collins GT, Paul NM, Grundt P, Newman AH, Xu M, Grandy DK, Woods JH, Katz JL. Yawning and locomotor behavior induced by dopamine receptor agonists in mice and rats. Behav Pharmacol. 2010;21:171–181. doi: 10.1097/FBP.0b013e32833a5c68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luedtke RR, Mach RH. Progress in developing D3 dopamine receptor ligands as potential therapeutic agents for neurological and neuropsychiatric disorders. Curr Pharm Des. 2003;9:643–671. doi: 10.2174/1381612033391199. [DOI] [PubMed] [Google Scholar]
- Marona-Lewicka D, Nichols DE. WAY 100635 produces discriminative stimulus effects in rats mediated by dopamine D4 receptor activation. Behav Pharmacol. 2009;20:114–8. doi: 10.1097/FBP.0b013e3283242f1a. [DOI] [PubMed] [Google Scholar]
- Mead A, Li M, Kapur S. Clozapine and olanzapine exhibit an intrinsic anxiolytic property in two conditioned fear paradigms: contrasts with haloperidol and chlordiazepoxide. Pharmacology, biochemistry, and behavior. 2008;90:551–62. doi: 10.1016/j.pbb.2008.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Missale C, Nash SR, Robinson SW, Jaber M, Caron M. Dopamine Receptors: From Structure to Function. Physiologicla Reviews. 1998;78:189–225. doi: 10.1152/physrev.1998.78.1.189. [DOI] [PubMed] [Google Scholar]
- Millan MJ, Gobert A, Newman-Tancredi A, Lejeune F, Cussac D, Rivet JM, Audinot V, Dubuffet T, Lavielle G. S33084, a novel, potent, selective, and competitive antagonist at dopamine D3-receptors: I. Receptorial, electrophysiological and neurochemical profile compared with GR218,231 and L741,626. J Pharmacol Exp Ther. 2000;293:1048–1062. [PubMed] [Google Scholar]
- Millan MJ, Seguin L, Gobert A, Cussac D, Brocco M. The role of dopamine D3 compared with D2 receptors in the control of locomotor activity: a combined behavioural and neurochemical analysis with novel, selective antagonists in rats. Psychopharmacology. 2004;174:341–57. doi: 10.1007/s00213-003-1770-x. [DOI] [PubMed] [Google Scholar]
- Morris B, Newman-Tancredi J, Audinot V, Simpson C, Millan M. Activation of Dopamine D3 Receptors Induces c-fos Expression in Primary Cultures of Rat Striatal Neurons. J Neurosci Res. 2000;59:740–749. doi: 10.1002/(SICI)1097-4547(20000315)59:6<740::AID-JNR6>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- Müller CP, De Souza Silva MA, DePalma G, Tomaz C, Carey RJ, Huston JP. The selective serotonin 1A-receptor antagonist WAY 100635 blocks behavioral stimulating effects of cocaine but not ventral striatal dopamine increase. Behav Brain Res. 2002;134:337–46. doi: 10.1016/s0166-4328(02)00042-6. [DOI] [PubMed] [Google Scholar]
- Neisewander JL, Fuchs RA, Tran-Nguyen LT, Weber SM, Coffey GP, Joyce JN. Increases in dopamine D3 receptor binding in rats receiving a cocaine challenge at various time points after cocaine self-administration: implications for cocaine-seeking behavior. Neuropsychopharmacology. 2004;29:1479–1487. doi: 10.1038/sj.npp.1300456. [DOI] [PubMed] [Google Scholar]
- Nestler E. Molecular mechanisms of drug addiction. Neuropharmacology. 2004;47:24–32. doi: 10.1016/j.neuropharm.2004.06.031. [DOI] [PubMed] [Google Scholar]
- Newman AH, Grundt P, Nader MA. Dopamine D3 receptor partial agonists and antagonists as potential drug abuse therapeutic agents. J Med Chem. 2005;48:3663–3679. doi: 10.1021/jm040190e. [DOI] [PubMed] [Google Scholar]
- Ohno Y, Shimizu S, Imaki J. Effects of tandospirone, a 5-HT1A agonistic anxiolytic agent, on haloperidol-induced catalepsy and forebrain Fos expression in mice. J Pharmacol Sci. 2009;109:593–9. doi: 10.1254/jphs.08313fp. [DOI] [PubMed] [Google Scholar]
- Ohno Y, Shimizu S, Imaki J, Ishihara S, Sofue N, Sasa M, Kawai Y. Anticataleptic 8-OHDPAT preferentially counteracts with haloperidol-induced Fos expression in the dorsolateral striatum and the core region of the nucleus accumbens. Neuropharmacology. 2008;55:717–23. doi: 10.1016/j.neuropharm.2008.06.005. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. Academic Press; San Diego: 1998. [Google Scholar]
- Pierce RC, Duffy RW, Kalivas P. Sensitization to cocaine and dopamine autoreceptor subsensitivity in the nucleus accumbens. Synapse. 1995;20:33–36. doi: 10.1002/syn.890200106. [DOI] [PubMed] [Google Scholar]
- Porrino LJ, Smith HR, Nader MA, Beveridge TJ. The effects of cocaine: a shifting target over the course of addiction. Progress in neuro-psychophamracology and biological psychiatry. 2007;31:1593–1600. doi: 10.1016/j.pnpbp.2007.08.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reavill C, Taylor S, Wood M, Ashmeade T, Austin N, Avenell K, Boyfield I, Branch C, Cilia J, Coldwell M, Hadley M, Medhurst A, Middlemiss D, Nash D, Riley G, Routledge C, Stempt G, Thewlis K, Trail B, Vong A, Hagan J. Pharmacological Actions of a Novel, High-Affinity, and Selective Human Dopamine D3 Receptor Antagonist, SB-277011-A. J Pharmacol Exp Ther. 2000;294:1154–1165. [PubMed] [Google Scholar]
- Robertson GS, Matsumura H, Fibiger HC. Induction patterns of Fos-like immunoreactivity in the forebrain as predictors of atypical antipsychotic activity. J Pharmacol Exp Ther. 1994;271:1058–1066. [PubMed] [Google Scholar]
- Rodriguez-Arias M, Felip C, Broseta I, Minarro The dopamine D3 antagonist U-99194A maleate increases social behaviors of isolation-induced aggressive male mice. Psychopharmacology. 1999;144:90–94. doi: 10.1007/s002130050981. [DOI] [PubMed] [Google Scholar]
- Roesch MR, Singh T, Brown PL, Mullins SE, Schoenbaum G. Ventral striatal neurons encode the value of the chosen action in rats deciding between differently delayed or sized rewards. J Neurosci. 2009;29:13365–13376. doi: 10.1523/JNEUROSCI.2572-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schenk S. Effects of the serotonin 5-HT2 antagonist, ritanserin, and the serotonin 5-HT1A antagonist, WAY 100635, on cocaine-seeking in rats. Pharmacol Biochem Behav. 2000;67:363–9. doi: 10.1016/s0091-3057(00)00377-4. [DOI] [PubMed] [Google Scholar]
- Schultz W, Tremblay L, Hollerman J. Reward processing in primate orbitofrontal cortex and basal ganglia. Cerebral Cortex. 2000;10:272–283. doi: 10.1093/cercor/10.3.272. [DOI] [PubMed] [Google Scholar]
- Segal DM, Moraes CT, Mash DC. Up-regulation of D3 dopamine receptor mRNA in the nucleus accumbens of human cocaine fatalities. Mol Brain Res. 1997;45:335–339. doi: 10.1016/s0169-328x(97)00025-9. [DOI] [PubMed] [Google Scholar]
- Staley JK, Mash DC. Adaptive increase in D3 dopamine receptors in the brain reward circuits of human cocaine fatalities. J Neurosci. 1996;16:6100–6106. doi: 10.1523/JNEUROSCI.16-19-06100.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheideler MA, Martin J, Hohlweg R, Rasmussen JS, Naerum L, Ludvigsen TS, Larsen PJ, Korsgaard N, Crider AM, Ghosh D, Cruse SF, Fink-Jensen A. The preferential dopamine D3 receptor agonist cis-8-OH-PBZI induces limbic Fos expression in rat brain. Eur J Pharmacol. 1997;339:261–270. doi: 10.1016/s0014-2999(97)01372-1. [DOI] [PubMed] [Google Scholar]
- Sokoloff P, Giros B, Martres M, Bouthenet M, Schwartz J. Molecular cloning and characterization of a novel dopamine receptor D3 as a target for neuroleptics. Nature. 1990;347:146–151. doi: 10.1038/347146a0. [DOI] [PubMed] [Google Scholar]
- Southam E, Lloyd A, Jennings C, Cluderay J, Cilia J, Gartlon J, Jones D. Effect of the selective dopamine D3 receptor antagonist SB-277011-A on regional c-fos-like expression in rat forebrain. Brain Research. 2007;1149:50–57. doi: 10.1016/j.brainres.2007.02.051. [DOI] [PubMed] [Google Scholar]
- Spangler R, Goddard NL, Avena NM, Hoebel BG, Leibowitz SF. Elevated D3 dopamine receptor mRNA in dopaminergic and dopaminoceptive regions of the rat brain in response to morphine. Brain Res Mol Brain Res. 2003;111:74–83. doi: 10.1016/s0169-328x(02)00671-x. [DOI] [PubMed] [Google Scholar]
- Stanwood G, Artymyshyn R, Kung M, Kung H, Lucki I, Mcgonigle P. Quantitative Autoradiographic Mapping of Rat Brain Dopamine D3 Binding with [125I]7-OH-PIPAT: Evidence for the Presence of D3 Receptors on Dopaminergic and Nondopaminergic Cell Bodies and Terminals. J Pharmacol Exp Ther. 2000;295:1223–1231. [PubMed] [Google Scholar]
- Surmeier DJ, Song WJ, Yan Z. Coordinated expression of dopamine receptors in neostriatal medium spiny neurons. J Neurosci. 1996;16:6579–6591. doi: 10.1523/JNEUROSCI.16-20-06579.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tu Z, Li S, Cui J, Xu J, Taylor M, Ho D, Luedtke RR, Mach RH. Synthesis and pharmacological evaluation of fluorine-containing D3 dopamine receptor ligands. J Med Chem. 2011;54:1555–1564. doi: 10.1021/jm101323b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor M, Grundt P, Griffin SA, Newman AH, Luedtke RR. Dopamine D3 receptor selective ligands with varying intrinsic efficacies at adenylyl cyclase inhibition and mitogenic signaling pathways. Synapse. 2010;64:251–266. doi: 10.1002/syn.20725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tilakaratne N, Friedman E. Genomic responses to 5-HT1A or 5-HT2A/2C receptor activation is differentially regulated in four regions of rat brain. Eur J Pharmacol. 1996;307:211–7. doi: 10.1016/0014-2999(96)00233-6. [DOI] [PubMed] [Google Scholar]
- Tremblay PO, Gervais J, Rouillard C. Modification of haloperidol-induced pattern of c-fos expression by serotonin agonists. Eur J Neurosci. 1998;10:3546–55. doi: 10.1046/j.1460-9568.1998.00372.x. [DOI] [PubMed] [Google Scholar]
- Tsuchida R, Kubo M, Shintani N, Abe M, Köves K, Uetsuki K, Kuroda M, Hashimoto H, Baba A. Inhibitory effects of osemozotan, a serotonin 1A-receptor agonist, on methamphetamine-induced c-Fos expression in prefrontal cortical neurons. Biol Pharm Bull. 2009;324:728–31. doi: 10.1248/bpb.32.728. [DOI] [PubMed] [Google Scholar]
- Uslaner J, Badiani A, Day HEW, Watson SJ, Akil H, Robinson TE. Environmental context modulates the ability of cocaine and amphetamine to induce c-fos mRNA expression in the neocortex, caudate nucleus, and nucleus accumbens. Brain Res. 2001;920:106–116. doi: 10.1016/s0006-8993(01)03040-2. [DOI] [PubMed] [Google Scholar]
- Vallone D, Picetti R, Borrelli E. Structure and function of dopamine receptors. Neuroscience and Biobehavioral Reviews. 2000;24:125–132. doi: 10.1016/s0149-7634(99)00063-9. [DOI] [PubMed] [Google Scholar]
- Vangveravong S, McElveen E, Taylor M, Xu J, Tu Z, Luedtke RR, Mach RH. Synthesis and characterization of selective dopamine D2 receptor antagonists. Bioorg Med Chem. 2006;14:815–825. doi: 10.1016/j.bmc.2005.09.008. [DOI] [PubMed] [Google Scholar]
- Vengeliene V, Leonardi-Essmann F, Perreau-Lenz S, Gebicke-Haerter P, Drescher K, Gross G, Spanagel R. The dopamine D3 receptor plays an essential role in alcohol-seeking and relapse. FASEB J. 2006;20:2223–2233. doi: 10.1096/fj.06-6110com. [DOI] [PubMed] [Google Scholar]
- Vorel SR, Ashby CR, Jr, Paul M, Liu X, Hayes R, Hagan JJ, Middlemiss DN, Stemp G, Gardner EL. Dopamine D3 receptor antagonism inhibits cocaine-seeking and cocaine-enhanced brain reward in rats. J. Neurosci. 2002;22:9595–9603. doi: 10.1523/JNEUROSCI.22-21-09595.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber M, Chang WL, Durbin JP, Park PE, Luedtke RR, Mach RH, Swerdlow NR. Using prepulse inhibition to detect functional D3 receptor antagonism: effects of WC 10 and WC 44. Pharmacol Biochem Behav. 2009;93:141–147. doi: 10.1016/j.pbb.2009.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welter M, Vallone D, Samad TA, Meziane H, Usiello A, Borrelli E. Absence of dopamine D2 receptors unmasks an inhibitory control over the brain circuitries activated by cocaine. Proc Natl Acad Sci USA. 2007;104:6840–6845. doi: 10.1073/pnas.0610790104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu J, Chu W, Tu Z, Jones L, Luedtke R, Perlmutter J, Mintun M, Mach R. [3H]4-(Dimethylamino)-N-[4-(4-(2-Methoxyphenyl)Piperazin-1-yl)Butyl]Benzamide, a Selective Radioligand for Dopamine D3 Receptors. I. In Vitro Characterization. Synapse. 2009;63:717–728. doi: 10.1002/syn.20652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu J, Hassanzadeh B, Chu W, Tu Z, Jones L, Luedtke R, Perlmutter J, Minton M, Mach R. [3H]4-(Dimethylamino)-N-[4-(4-(2-Methoxyphenyl)Piperazin-1-yl)Butyl]Benzamide: A Selective Radioligand or Dopamine D3 receptors. II. Quantitative Analysis of Dopamine D3 and D2 Receptor Density Ratio in the Caudate-Putamen. Synapse. 2010;64:449–459. doi: 10.1002/syn.20748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang L, Lou D, Jiao H, Zhang D, Wang X, Xia Y, Zhang J, Xu M. Cocaine-Induced Intracellular Signaling and Gene Expression Are Oppositely Regulated by the Dopamine D1 and D3 Receptos. J Neurosci. 2004;24:3344–3354. doi: 10.1523/JNEUROSCI.0060-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]