

Abstract
Many patients with hyperandrogenemia are overweight or obese, which exacerbates morbidities associated with polycystic ovary syndrome (PCOS). To examine the ability of testosterone (T) to generate PCOS-like symptoms, monkeys received T or cholesterol (control) implants (n = 6/group) beginning prepubertally. As previously reported, T-treated animals had increased neuroendocrine drive to the reproductive axis [increased luteinizing hormone (LH) pulse frequency] at 5 yr, without remarkable changes in ovarian or metabolic features. To examine the combined effects of T and obesity, at 5.5 yr (human equivalent age: 17 yr), monkeys were placed on a high-calorie, high-fat diet typical of Western cultures [Western style diet (WSD)], which increased body fat from <2% (pre-WSD) to 15–19% (14 mo WSD). By 6 mo on WSD, LH pulse frequency in the controls increased to that of T-treated animals, whereas LH pulse amplitude decreased in both groups and remained low. The numbers of antral follicles present during the early follicular phase increased in both groups on the WSD, but maximal follicular size decreased by 50%. During the late follicular phase, T-treated females had greater numbers of small antral follicles than controls. T-treated monkeys also had lower progesterone during the luteal phase of the menstrual cycle. Although fasting insulin did not vary between groups, T-treated animals had decreased insulin sensitivity after 1 yr on WSD. Thus, while WSD consumption alone led to some features characteristic of PCOS, T + WSD caused a more severe phenotype with regard to insulin insensitivity, increased numbers of antral follicles at midcycle, and decreased circulating luteal phase progesterone levels.
Keywords: testosterone, obesity, amenorrhea, insulin insensitivity, ovarian follicle
polycystic ovary syndrome (PCOS) affects 4–8% of reproductive-aged women (5, 48) and is characterized by hyperandrogenemia (HA), irregular menstrual cycles, and polycystic ovaries, in addition to rapid luteinizing hormone (LH) pulse frequency, increased LH response to gonadotropin-releasing hormone (GnRH), decreased sensitivity to progesterone (P4) negative feedback, and decreased insulin sensitivity (SI) (13, 20, 90). Moreover, up to 90% of women with PCOS are overweight or obese, which is higher than the general population of the United States, where 65% of women over the age of 20 are overweight or obese (6, 33). Women with PCOS also tend to have a central distribution of adiposity, which is associated with increased rates of high blood pressure and cardiovascular disease (23, 51).
Being overweight or obese aggravates many symptoms of PCOS. Basal production of testosterone (T) and free T are reportedly elevated in obese, compared with nonobese, PCOS patients (2, 39, 45, 63). Hirsutism, a clinical marker of elevated androgens, is also more severe in obese PCOS women (45). Obese PCOS patients also have higher rates of oligo- and amenorrhea, infertility, and miscarriage compared with lean women with the disorder (8, 45, 65). Additionally, between 50 and 70% of women with PCOS suffer from some degree of insulin insensitivity, and SI scores worsen with increasing body mass index (BMI) (21, 28, 43, 62). Compensatory hyperinsulinemia follows a similar pattern, with obese PCOS women having the highest insulin secretion (20). Many PCOS patients resume menstruation and ovulation with even mild weight loss, although this strategy is not always successful at normalizing reproductive function (48, 67).
Obesity in peripubertal girls is also associated with HA (57), and this is thought to be a forerunner of adult PCOS (34, 88). Like adults with PCOS, obese girls with HA have increased levels of LH and decreased sensitivity to P4 negative feedback (22, 49). In late puberty, girls with obesity reportedly have increased frequency of pulsatile LH release compared with nonobese girls (57). In addition, obese girls are more likely to have irregular menstrual cycles during late puberty compared with nonobese controls (57). As with adults, hyperinsulinemia is also more common in adolescents with PCOS, especially in those who are obese (76).
Cumulatively, there is substantial evidence that PCOS is characterized by both HA and obesity. However, there are women with PCOS who are normal weight (6, 53), indicating that, while obesity likely aggravates symptoms of PCOS, it is probably not the sole cause of PCOS. In a previous study of female monkeys that were treated chronically with T beginning at 1 yr of age, we observed that elevated serum T levels led to an increased central drive to the reproductive axis (59), similar to what is seen in women with PCOS (3, 7, 86). However, there were no robust group differences in ovarian or metabolic function after 4 yr of continuous exposure to slightly elevated T. Thus, based on evidence that obesity aggravates PCOS symptoms in women, it was of interest to investigate whether weight gain would also lead to the development of ovarian or metabolic symptoms in these T-treated monkeys. All females from the original cohort (59) were placed on a high-calorie, high-fat Western style diet (WSD). We hypothesized that there would be a continued effect of T on neuroendocrine function, in addition to the development of an ovarian and metabolic PCOS-like phenotype in the T-treated animals.
MATERIALS AND METHODS
Animals
Twelve female rhesus macaques (Macaca mulatta) that were 5.5 yr of age (young adults) at the beginning of this study were used for this protocol and were the same animals that we reported on previously (59). All monkeys were housed individually in single cages (81 × 61 × 69 cm) in a temperature-controlled room (24 ± 2°C), with lights on for 12 h/day (0700–1900). Monkeys were trained to approach the front of their cage so menses could be detected daily throughout the study by vaginal swabbing. The first day of menses was designated day 1 of a menstrual cycle. All procedures in this study were reviewed and approved by the Oregon National Primate Research Center (ONPRC) Institutional Animal Care and Use Committee.
Western Style Diet
Until the monkeys were 5.5 yr of age, they were fed two meals of Purina LabDiet fiber-balanced monkey chow each day (no. 5000; Purina Mills, St. Louis, MO), supplemented with fresh fruits and vegetables. Thereafter, their diet was switched from fiber-balanced monkey chow (15% calories from fat, 27% from protein, 59% from carbohydrates) to a diet that was specifically formulated to mimic a typical WSD (33% calories from fat, 17% from protein, 51% from carbohydrates; 5A1F; Purina Mills) (80, 81). The animals were fed ad libitum, and each meal was supplemented with a high-sugar, high-calorie treat (e.g., a cookie, snack cake, etc.). Previous studies have shown that monkeys eating this diet gained over 10% of their body weight within the first 5–6 mo of being on this diet (80, 82).
Testosterone and Cholesterol Implants
T and cholesterol implants were prepared and placed subcutaneously as described previously (59). Blood samples were taken weekly to ensure that the implants sustained a three- to fourfold increase in T levels above those in control monkeys (1.2–1.6 ng/ml in T-treated monkeys), as is observed in obese girls who are predisposed to developing PCOS (58). All animals received their first T- or cholesterol-containing (n = 6/group) implant at 1 yr of age. Implants were made of Silastic tubing (Dow Corning, Midland, MI) and were initially 5 mm in length with an inner and outer diameter of 0.335 and 0.465 cm, respectively. Implants were filled with cholesterol (control animals), or a T/cholesterol mixture (T-treated animals), with a T-to-cholesterol ratio of 1:15 or 1:12 at the beginning of the experiment. As the animals grew, the length of the implant increased to 3 cm and the T-to-cholesterol ratio increased gradually to 1:4 to maintain the desired serum T levels. Both cholesterol and T were purchased from Sigma Aldrich (St. Louis, MO).
Catheterization
The average age at menarche in both T-treated and control animals was ∼2.5 yr (59). At 3.5 yr of age, a chronic indwelling venous catheter was implanted in the subclavian or femoral vein by the ONPRC surgical staff using isoflurane anesthesia (Hospira, Lake Forest, IL), as described previously (59). Previous studies established that female monkeys with chronic indwelling catheters display normal, regular menstrual cycles (38). Catheters were kept patent with a constant infusion of physiological saline (Baxter Healthcare, Deerfield, IL) containing heparin sodium (4 IU/ml), at a rate of ∼100 ml/day. Animals were sedated weekly with ketamine hydrochloride (Ketaset, 10 mg/kg iv; Wyeth, Madison, NJ) to inspect the catheter system and replace a sterile dressing covering the exit site. No ketamine was administered in the 24 h preceding any experiment. Following catheterization, animals were allowed a minimum of 3 wk to recover before any experiments were performed.
Blood Collection
Blood samples were collected weekly from all animals to track serum P4, estradiol (E2), and T concentrations. Small aliquots of blood were collected through the remote sampling system and processed as described previously (59). Plasma was removed, and the red blood cells were sterilely resuspended in saline and reinfused through the catheter system to the animal. Each week's samples were assayed for T, and, when an individual animal's serum T concentration fell below the threshold of 1.2 ng/ml, the implant was replaced. Cholesterol implants were also changed regularly so that the control animals received the same average number of implant surgeries as the T-treated animals. Blood samples were also collected daily during one menstrual cycle in each animal (at 6.5 yr of age) to track the patterns and levels of E2 and P4 during the follicular and luteal phases. More frequent samples were collected during the neuroendocrine and glucose tolerance testing (GTT) protocols, as reported previously (59).
Assays
T was measured by ELISA (catalog no. IB79106; IBL America, Minneapolis, MN) by the Endocrine Technology Support Core (ETSC) Laboratory at the ONPRC. The sensitivity of the T assay was 0.08 ng/m, and the intra- and interassay coefficients of variation for the assays were 8.9 and 13.5%, respectively. Both E2 and P4 were also assayed by the ETSC Laboratory using the Immulite 2000 platform as previously described (59). As with many validated clinical platforms, the Immulite 2000 runs three quality control (QC) serum pools daily, and, as such, no specific intra-assay QC data are available. The interassay coefficient of variation, reflecting variability in daily QC results over the period in which these assays were performed was 8.5% for E2 and 9.4% for P4. Sex hormone-binding globulin (SHBG) was measured by a monkey-specific SHBG ELISA validated by the ETSC (catalog no. 251544; Hölzel Diagnostika, Köln, Germany). The sensitivity of the SHBG assay was 9.6 pmol/ml, and the intra-assay coefficient of variation was 4.9%.
LH and follicle-stimulating hormone (FSH) were measured by RIA at the University of Pittsburgh assay core using recombinant cynomolgus monkey LH and FSH (National Hormone and Peptide Program, Harbor-UCLA Medical Center, Torrance, CA) as standards (59, 86). The sensitivity of the LH assay was 0.1 ng/ml, and the intra- and interassay coefficients of variation for the assays used in this study were 6.6 and 12.2%, respectively. The sensitivity of the FSH assay was 0.05 ng/ml, and the intra-assay coefficient of variation was 9.3%.
Physical Activity
Physical activity levels were assessed continuously, as described previously (41), for the final 10 mo of this study, when the animals were 6–7 yr of age. The animals were on the WSD for the entire duration that activity was monitored. Animals wore jackets that contained a pocket housing a three-way accelerometer (Actical; Respironics, Bend, OR). The monitors were programmed to record total activity counts per minute, and data were downloaded approximately one time every 3 wk when monkeys were sedated with ketamine hydrochloride.
Experimental Protocols
Reproductive and metabolic parameters were measured before (pre-WSD) and after (up to 16 mo) onset of the WSD (see Fig. 1). The animals began WSD consumption at 5.5 yr of age.
Fig. 1.

Schematic diagram indicating ages (with approximate human equivalent ages) at which experiments were performed. Shaded area indicates period of Western style diet (WSD) consumption. DEXA, dual-energy X-ray absorptiometry.
Experiment 1. Evaluation of neuroendocrine function of the reproductive axis.
Pulsatile LH secretion was measured during the early follicular phase (D1–3) of the menstrual cycle in the animals when they were 5 yr (pre-WSD, data previously reported in Ref. 59), and again after 6 and 14 mo on the WSD. Small blood samples (0.4 ml/sample) were collected as described previously (59) every 10 min from 1300 to 0100. The early follicular phase was chosen because this is the time period when differences in pulsatile LH secretion are most apparent between women with PCOS and healthy women (56), and when differences were seen between the groups in our previous studies (59).
Pulsatile LH secretion was also measured during the luteal phase of a menstrual cycle after 6 and 14 mo on the WSD. Daily blood samples were taken beginning on D8 of the menstrual cycle and were assayed to track P4 levels on a day-by-day basis. On the first day that P4 levels were ≥2.0 ng/ml, pulsatile LH secretion was measured using the methods described above. A threshold of 2.0 ng/ml was chosen because it indicates a functional corpus luteum and is a standardized functional level of P4 (i.e., prevents menstruation).
LH responsiveness to GnRH (National Hormone and Peptide Program, Harbor-UCLA Medical Center) was measured between 0900 and 1000 on D2–4 of a menstrual cycle at 5 yr of age (pre-WSD) and after 6 and 14 mo on the WSD, as previously described (59). GnRH (250 ng/kg iv) was infused at time (t) 0, and blood samples (0.4 ml/sample) were collected at −15, −8, −1, 15, 30, 60, and 90 min (19, 59). This dose was chosen so that monkeys would receive a physiological dose that caused an LH response, but a response that was submaximal to allow detection of individual differences in LH responsiveness.
Experiment 2. Ovarian structure-function.
Weekly (throughout the protocol) and daily (for one menstrual cycle, 14 mo after beginning WSD) blood samples were assayed for E2 and P4 as described above, and the presence of the follicular and luteal phases of the menstrual cycle were determined based on these data. From the daily samples, the length of the follicular phase was denoted as the number of days from onset of menses to the day of the peak E2 value (inclusive). The luteal phase length was defined as occurring from the day after the peak E2 value to the day before onset of the next menses (inclusive).
Ovarian ultrasounds were performed on D1–3 of a menstrual cycle when the animals were 5 yr old (pre-WSD, data reported previously, see Ref. 59) and 3 and 14 mo after onset of WSD. At 14 mo WSD, ultrasounds were also performed during the late follicular phase, once E2 levels reached >100 pg/ml or at 10 days postmense, whichever occurred first. A GE Medical Systems Voluson 730 Expert Doppler ultrasound instrument (GE Healthcare, Waukesha, WI) with both two-dimensional (2D, 4.5–16.5 MHz) and four-dimensional (4D, 3.3–9.1 MHz) transabdominal probes was used, as described by Bishop et al. (12). Animals were assigned random identifiers so the ultrasonographer (Bishop) remained blind to the treatment group. The 2D probe was used to orient the image field to the uterus and identify the ovaries. The 4D probe was then used to generate a data file of each individual ovary, which included a series of images collected in one scan through the entire ovary. Ovaries were analyzed for circumference, number of antral follicles (0.5 mm diameter or greater) in each ovary, and the mean, maximum, and minimum size of the antral follicles in each ovary, using previously defined methods for adult female rhesus monkeys (12), and as reported previously (59). All parameters were then decoded for comparisons between treatment groups.
Experiment 3. Metabolic parameters.
GTT was performed during the early follicular phase of a menstrual cycle when the animals were 3.5 yr of age (pre-WSD; data reported previously in Ref. 59) and 12 mo after onset of WSD. For monkeys not showing regular menstrual cycles, the GTT was performed when a blood sample indicated that E2 and P4 levels were low, and thus the absence of a dominant follicle or corpus luteum in the ovaries. Each animal was sedated initially with telazol (tiletamine hydrochloride and zolazepam hydrochloride, 2–6 mg/kg im; Fort Dodge Animal Health, Fort Dodge, IA) and subsequently with ketamine to maintain sedation. The protocol was based on that designed by Bergman et al. (11) and was described previously (59). Dextrose (300 mg/kg) was infused intravenously through the catheter system, and blood samples were taken from 15 min before to 3 h after the glucose infusion. Tolbutamide (5 mg/kg) was infused intravenously 20 min after the dextrose to stimulate the pancreas to secrete more insulin. All samples were immediately assayed for glucose using the YSI 2300 Stat Plus (YSI, Yellow Springs, OH) and subsequently for insulin by RIA (Linco Human Insulin RIA; Millipore, Billerica, MA). The sensitivity of the insulin assay was 1 μIU/ml, and the intra-assay coefficient of variation was 4.9%.
Metabolic rate was measured over a 24-h period when the animals were 4.5 yr old (pre-WSD) and 12 mo after onset of WSD, using previously published methods (81, 82). The animals were placed in a sealed Lexan and stainless steel metabolic chamber (Columbus Instruments, Columbus, OH) at ∼1000. Fresh air was pumped in and circulated with a 4-in. fan. The amounts of oxygen consumed and carbon dioxide produced were measured using a computer-controlled open-circuit calorimeter, and total energy expenditure (kcal) was calculated using the Oxymax system (Columbus Instruments). The animals did not receive their normal meals during this time but were fed a 110-g banana at 1500.
Dual-energy X-ray absorptiometry (DEXA) scanning was performed at 5 yr of age (pre-WSD; data reported previously, see Ref. 59) and 16 mo after onset of WSD. Percent body fat, percent central fat, fat mass, and lean tissue mass were determined using DEXA scanning. Monkeys were sedated with ketamine and positioned supine on the bed of a Hologic Dexa scanner (Discovery scanner; Hologic, Bedford, MA). Two to three scans were performed for each monkey in “infant whole body” mode, and averages were calculated for each measure. To delineate central fat mass from peripheral fat mass, fat in the trunk (including both the subcutaneous and visceral compartments) and fat in the extremities were calculated using standard methodology (24, 59).
Statistical Analysis
Pulsatile LH secretion.
LH pulses were identified using the Pulsar algorithm developed by Merriam and Wachter (62), and used previously to detect LH pulses in these monkeys (59). Pulse frequency was defined as the number of Pulsar-detected pulses in 12 h and the following G values were used: G(1), 50.00; G(2), 1.0; G(3), 0.40; G(4), 0.40; and G(5), 0.40. Pulse amplitude was calculated by subtracting the baseline LH level from the peak LH level of each pulse. For all analyses, LH values below the level of detectability for the assay were assigned the minimum detectable concentration of the assay. Independent Student's t-tests were used to determine group differences in the number of pulses, proportion of daytime (vs. nighttime) pulses, pulse amplitude, and LH response to GnRH. For the GnRH stimulation, LH area under the curve (AUC) was calculated, correcting for baseline LH levels.
Ovarian morphology, function, and interactions.
Changes in total numbers of antral follicles measured by ultrasound analysis on D1–3 (combining counts from both ovaries in each animal) from before and throughout WSD were analyzed by the mixed-models procedure (treatment × time). Numbers of antral follicles present on each ovary during the late follicular phase (after possible selection of the dominant follicle) and P4 AUC levels during the luteal phase were analyzed by ANOVA (main effect of treatment). Post hoc tests employed were paired t-tests with a Bonferroni correction for multiple comparisons. Pearson r correlations were used to examine relationships between neuroendocrine, antral follicle counts (by ovary), and metabolic parameters independently of T exposure to determine the main effect of WSD on ovarian structure-function.
Metabolic parameters.
The MINMOD Millennium computer program was used to determine glucose effectiveness (Sg), SI, acute insulin response to glucose (AIRg), and disposition index (DI) values (15). This program was designed to calculate these values based on the GTT protocol that was described by Bergman and colleagues (11) and used in this study. Sg indicates the capacity of glucose to mediate its own disposal, AIRg addresses the adequacy of insulin secretion, and SI quantifies the capacity of insulin to promote glucose disposal. DI is the product of AIRg and SI, and it therefore takes into account insulin concentration and action (10, 15). Independent-sample t-tests were used to determine group differences in GTT measures.
Basal metabolic rate was calculated as the average number of kilocalories expended per kilogram per hour from 2300 to 0300. This time period is when monkeys are typically asleep and their heart rate is slowest (Cameron, unpublished data). BMI (kg/m2) was calculated using the crown-rump length as height. The free androgen index was calculated as T/SHBG × 100. An independent t-test was used to assess group differences in basal metabolic rate and body composition parameters.
Mixed-measures ANOVAs were used to determine changes in all measures across time. Some data from this study were compared with data collected from the same animals before WSD treatment (59), when the monkeys were between 3.5 and 5 yr of age. Statistical analyses of ovarian function and morphology were performed using SAS (version 9.2; SAS Institute, Cary, NC). All other analyses were performed using IBM SPSS Statistics 20 (SPSS, Chicago, IL). All values are presented as means ± SE. Significance was set at P < 0.05.
RESULTS
Weight Increase and Testosterone Concentrations
The animals increased their daily caloric intake by an average of 140 ± 13% when switched from normal chow to the WSD (before WSD: 467 ± 0 kcal/day; after WSD: 1,121 ± 62 kcal/day), but there were no differences between T-treated and control animals with regard to average kilocalories of WSD consumed per day (P = 0.95). All animals increased their body weight by at least 10% while on the WSD (average percent gain for T-treated animals: 29.4 ± 7.5%, controls: 27.3 ± 4.0%). When weight gain over the entire postpubertal period was analyzed (i.e., from 3.5 to 7 yr of age), there was a significant effect of treatment [F(1,10) = 5.19, P = 0.046], with T-treated animals gaining more weight than controls, although percent weight gain on the WSD was not different between the groups (P > 0.1). Weight gain while on the WSD was positively correlated with caloric intake (r = 0.78, P = 0.003). BMI increased from 21.8 ± 0.5 kg/m2 before the WSD to 25.6 ± 1.2 kg/m2 after 16 mo on the WSD (P = 0.002), and was not different between the groups (P > 0.1). There was a strong trend toward T-treated animals being less active than controls over the 10-mo period that activity was measured [average activity counts/day: T treated, 201,852 ± 54,929; control, 423,053 ± 84,067; t(10)=2.20, P = 0.052]. However, there was no correlation between weight gain during WSD consumption and average daily activity during the last 10 mo of the study (r = −0.17, P = 0.60). There was also no correlation between postpubertal weight gain and average daily activity during the final 10 mo of the study (r = −0.001, P = 0.997).
Plasma T concentrations in T-treated animals were maintained in a narrow range regardless of weight gain; levels were 4.2 ± 0.2-fold higher than in control animals from the time of first implant at 1 yr of age through 7 yr of age (T treated, 1.71 ± 0.05 ng/ml; control, 0.46 ± 0.02 ng/ml, P = 0.001). SHBG levels did not change from before to 16 mo after onset of WSD, and were not different between the groups either before or after the WSD (before WSD: T treated, 13.1 ± 1.1 pmol/ml; control, 13.1 ± 1.0 pmol/ml; after WSD: T treated, 14.4 ± 1.5 pmol/ml; control, 12.8 ± 1.1 pmol/ml; P > 0.1). The free androgen index was higher in T-treated animals compared with controls both before and after WSD consumption (both P < 0.01), and decreased slightly after consuming the WSD in both groups (before WSD: T treated, 11.6 ± 1.6; control, 2.3 ± 0.4; after WSD: T treated, 9.3 ± 1.9; control, 1.1 ± 0.1). Implants for both groups were replaced an average of every 9.2 ± 0.4 wk throughout the study.
Experiment 1. Neuroendocrine Function of the Reproductive Axis
When assessing the change in LH pulse frequency from before to 14 mo after starting the WSD, there was a significant diet × treatment interaction [F(1,9) = 8.8, P = 0.016]. This resulted from an increase in pulse frequency in the control but not the T-treated animals from before to 6 mo after beginning the WSD [t(5) = −5.0, P = 0.004; Fig. 2A]. LH pulse number did not change in either group when comparing the data from 6 mo with 14 mo on WSD [before WSD: T treated, 9.7 ± 1.8 pulses/12 h; control, 3.7 ± 1.8; after 6 mo WSD: T treated, 11.4 ± 1.0; control, 13.3 ± 1.1; after 14 mo WSD: T treated, 13.0 ± 1.0; control, 12.3 ± 1.0; Fig. 2A]. There was a significant effect of diet on LH pulse amplitude, which decreased in both groups by 6 mo after the animals switched to the WSD and remained lower after 14 mo [before WSD: T treated, 0.73 ± 0.21 ng/ml; control, 0.70 ± 0.15 ng/ml; after 6 mo WSD: T treated, 0.16 ± 0.02 ng/ml; control, 0.38 ± 0.12 ng/ml; after 14 mo WSD: T treated, 0.27 ± 0.04 ng/ml; control, 0.34 ± 0.11 ng/ml; F(1,8)=13.1, P = 0.007; Fig. 2B]. There was no effect of time or treatment, nor was there a time × treatment interaction on LH response to exogenous GnRH (either assessed as LH AUC or peak LH, data not shown; all P > 0.1).
Fig. 2.
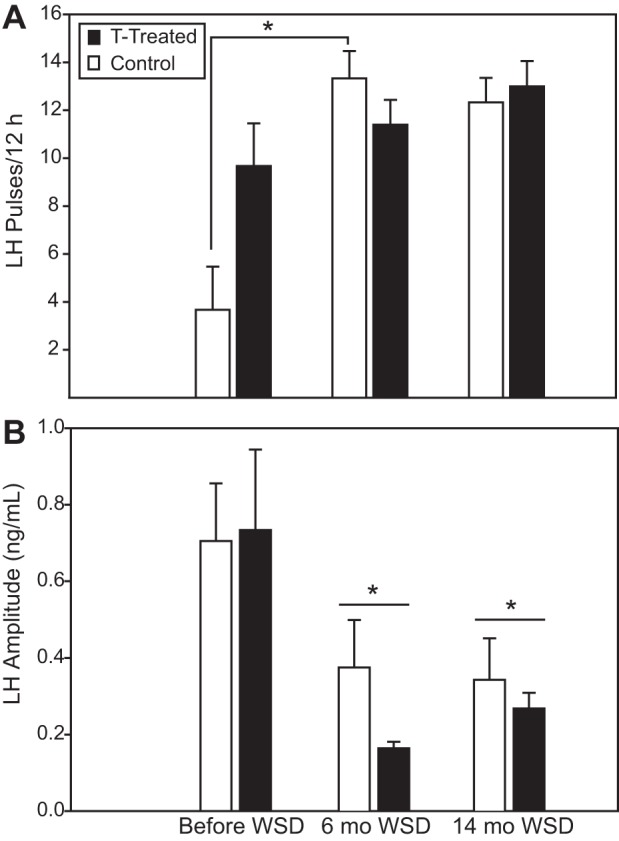
Number (A) and amplitude (B) of luteinizing hormone (LH) pulses occurring in testosterone (T)-treated and control animals during a 12-h period on days (D) 1–3 of a menstrual cycle before, 6 mo after, and 14 mo after starting the WSD. Data from before WSD were previously published (59). Closed bars, T-treated animals; open bars, control animals. *Significantly different from before WSD (P < 0.05).
Six months after starting the WSD, the effect of P4 negative feedback on pulsatile LH secretion was examined by comparing LH pulsatility measured during the early follicular phase (D1–3) with that measured during the luteal phase on the day that plasma P4 was first over 2 ng/ml (Fig. 3, top). Two control animals and one T-treated animal were not tested because of either lack of ovulation or catheter removal. The analyses were performed on the remaining four controls and five T-treated monkeys. There were no group differences in LH pulse frequency, LH pulse amplitude, or P4 level measured during the luteal phase pulse bleeds (all P > 0.1). Most animals showed a decrease in LH pulse frequency measured during the luteal phase compared with the values on D1–3, and there were no group differences in percent suppression [t(7)= −0.88, P = 0.4; Fig. 3]. This experiment was performed again after 14 mo on the WSD (Fig. 3, bottom). Two T-treated animals were not tested due to catheter removal, so analyses were performed on the remaining four T-treated and six control animals. Similar to the results after 6 mo on the WSD, most animals showed a decrease in pulse frequency during the luteal phase compared with the D1–3 data. Again, however, there were no group differences in LH pulse frequency or amplitude, or percent suppression between the follicular and luteal measures (all P > 0.1; Fig. 3). When analyzing data from the four animals from each group that were tested after both six and 14 mo of WSD, there was a weak effect of time on percent suppression of LH during the luteal phase [6 mo WSD: 29.6 ± 9.5% suppressed; 14 mo WSD: 42.0 ± 9.0% suppressed; F(1,3) = 6.16, P = 0.09], but there was no effect of T treatment (P > 0.1).
Fig. 3.

Suppression of LH pulsatility in the luteal phase compared with the early follicular phase of the menstrual cycle after 6 (A and B) and 14 (C and D) mo on the WSD. Each panel shows LH pulse frequency on D1–3 (circles) and during the luteal phase (diamonds) as a function of average progesterone values on the day of blood sampling. Data from individual animals are connected by lines. Data are shown from control animals, 6 mo WSD (A); T-treated animals, 6 mo WSD (B); control animals, 14 mo WSD (C); and T-treated animals, 14 mo WSD (D).
Experiment 2. Ovarian Structure-Function
When comparing the year before with the year after beginning the WSD, animals had an increased number of menstrual cycles [before WSD: 7.3 ± 0.6 cycles; after WSD: 9.2 ± 0.4 cycles; t(11) = −2.99, P = 0.01]. There was also an increased number and percentage of putative ovulatory cycles (as determined by elevated P4 during the luteal phase) from before to after onset of WSD [before WSD: 49.6 ± 8.7% ovulatory; after WSD: 86.2 ± 2.8% ovulatory; t(11) = −4.62, P = 0.001]. T-treated and control animals did not differ in the number of menstrual cycles (T treated, 14.2 ± 0.9 cycles; control, 13.8 ± 0.6 cycles) or in the number or percentage of ovulatory cycles (T treated, 84.7 ± 4.9% ovulatory; control, 87.7 ± 3.1% ovulatory) that occurred when they were 5.5–7 yr of age and on the WSD.
Samples were taken daily throughout one entire menstrual cycle after the animals had been on the WSD for 14 mo, and each animal experienced an ovulatory cycle as indicated by elevated P4 levels. There was no overall effect of 14 mo of WSD on peak E2 during the follicular phase, nor on peak P4 or P4 AUC during the luteal phase when all of the animals were considered (6 T treated and 6 control). This was still true when only analyzing animals that ovulated (4 T treated and 5 controls ovulated before WSD; all animals ovulated 14 mo after WSD). However, in animals that were ovulatory both before and after WSD, there was a significant decrease in D1–3 E2 values after 14 mo on the WSD [before WSD: T treated, 78.2 ± 18.0 pg/ml; control, 90.2 ± 12.0 pg/ml; 14 mo WSD: T treated, 53.4 ± 10.7 pg/ml; control, 49.8 ± 6.1 pg/ml; F(1,7) = 5.57, P = 0.05; Fig. 4], and a significant decrease in E2 surge dynamics (measured by AUC) after 14 mo on the WSD in control females, and T-treated females tended to have reduced E2 AUC [before WSD: T treated AUC, 1,879 ± 184.8; control AUC, 2,056 ± 259.4; 14 mo WSD: T treated AUC, 1,205.5 ± 220.6; control AUC 1,289.5 ± 121.3; repeated-measures ANOVA treatment × year F(2,10), P = 0.025; control (before vs. after WSD), P = 0.032; T treated (before vs. after WSD), P = 0.062]. Cycle length also decreased significantly from before to 14 mo after WSD [before WSD: T treated, 30.2 ± 2.7 days; controls, 30.6 ± 1.9 days; 14 mo WSD: T treated, 25.8 ± 0.9 days; controls, 26.8 ± 0.6 days; F(1,7) = 8.66, P = 0.022; Fig. 4]. This was due mainly to a decrease in follicular phase length [before WSD: T treated, 17.0 ± 4.1 days; controls: 15.2 ± 2.5 days; 14 mo WSD: T treated, 11.5 ± 1.2 days; controls: 10.8 ± 0.6 days; F(1,7)=4.92, P = 0.062; Fig. 4], which mimicked E2 AUC pattern, since there was no significant change in luteal phase length. Luteal phase length was positively correlated with P4 AUC (r = 0.604, P = 0.037).
Fig. 4.

Characteristics of the menstrual cycle based on daily blood samples and checks of menses. Only animals that ovulated were included (n = 4 T treated and 5 controls before WSD; n = 6/group 14 mo after WSD). A: total menstrual cycle length. B: follicular phase length. C: luteal phase length. D: peak estrogen. E: average estrogen from D1 to 3. F: peak progesterone. Closed bars, T-treated animals. Open bars, controls. *Significant difference between groups, P = 0.02; †significant difference from before WSD, P < 0.05; #trend toward difference from before WSD, P = 0.062.
After 14 mo on the WSD, there was a significant treatment effect on peak P4 value during the luteal phase, with T-treated animals having lower peak P4 compared with controls [T treated, 6.8 ± 1.1 ng/ml; control, 12.1 ± 1.6 ng/ml; t(10) = 2.0, P = 0.02; Fig. 4F]. T-treated animals also had a lower AUC when P4 values were assessed during the luteal phase [T treated, 32.7 ± 5.4 ng/luteal phase; control, 52.4 ± 6.0 ng/luteal phase; t(10) = 2.45, P = 0.034]. There were no group differences in E2 values measured on D1–3, peak E2 during the follicular phase, FSH measured on D1–2, the LH-to-FSH ratio on D1–2, follicular phase length, luteal phase length, or total cycle length (all P > 0.1). These results did not change when controlling for weight.
When analyzing the ovarian ultrasound data from the early follicular phase from before to 14 mo after beginning the WSD, there was an increase in total numbers of antral follicles [F(2,20) = 45.8, P < 0.001; Fig. 5A] and a decrease in maximum follicle size [F(2,20) = 7.92, P = 0.003; Fig. 5B] in both groups, but there was no diet × treatment interaction and no main effect of T treatment. However, after 14 mo on WSD, T-treated females displayed more antral follicles per ovary than the controls during the late follicular phase [F(1,10) = 6.38, P = 0.027; Fig. 6A]. Additionally, the size of the largest antral follicle was smaller in T-treated females compared with controls during the late follicular phase [control, 2.1 ± 0.3 mm; T treated, 1.4 ± 0.1 mm; F(1,10) = 4.84, P = 0.053; Fig. 6B]. There was also a significant correlation between percent weight gained on WSD and the total number of antral follicles present on ovaries during the late follicular phase (r = 0.425, P = 0.039; Fig. 6C). There were no effects of time or T treatment on ovarian circumference (P > 0.1).
Fig. 5.
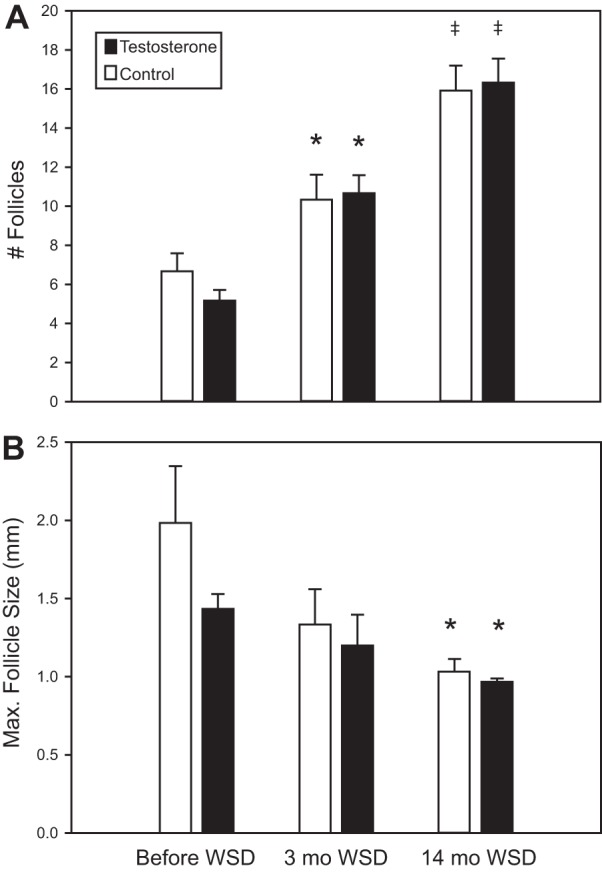
Changes in antral follicles present on ovaries during the early follicular phase as measured by ultrasound from before to 14 mo after starting WSD. Closed bars, T-treated animals. Open bars, controls. *Significant difference (P < 0.05) from before WSD; ‡significant difference (P < 0.05) from both before WSD and 3 mo WSD.
Fig. 6.

Effects of hyperandrogenemia (T) and WSD on antral follicles during the late follicular phase after 14 mo of WSD consumption. T-treated females showed greater numbers of antral follicles in the ovaries (A), and smaller follicle size (B) during the late follicular phase (after the dominant follicle is typically selected). *Significant changes due to T exposure (P < 0.05). C: effects of WSD on antral follicle count correlate positively with %body fat after 14 mo on the WSD; r = 0.42, P < 0.05.
Experiment 3. Metabolic Parameters
When GTT data were compared between before and 12 mo after onset of WSD, there was a significant increase in peak glucose [F(1,10) = 8.0, P = 0.018], baseline (fasting) insulin [F(1,10) = 27.1, P < 0.001; Fig. 7D], peak insulin [F(1,10) = 8.41, P = 0.016], and AIRg [F(1,10) = 10.2, P = 0.01]. However, there was no main effect of WSD on SI (P = 0.61) or DI (P = 0.29). Weight, percent fat, percent central fat, and changes in these variables while on the WSD were not correlated with any parameters measured using the GTT.
Fig. 7.

Insulin sensitivity, disposition index, fasting glucose, and fasting insulin measured during a glucose tolerance test before and after 12 mo of WSD consumption. Closed bars, T-treated animals. Open bars, controls. *Significant group difference (P < 0.05); #significant difference from before WSD.
After 12 mo on the WSD, there were no treatment effects on baseline or peak glucose, baseline or peak insulin, or on AIRg or Sg as calculated by MINMOD (all P > 0.1, data not shown). However, T-treated animals displayed lower SI [T treated, 5.3 ± 1.8 (mU/l)−1/min; control, 13.9 ± 3.4 (mU/l)−1/min; t(10)=2.23, P = 0.05; Fig. 7A]. There was also a significant difference in DI, the unitless product of SI and AIRg, with T-treated animals having a lower DI [T treated, 4,223 ± 791; control, 7,567 ± 1,099; t(10) = 2.47, P = 0.03; Fig. 7B]. These differences remained significant when controlling for weight.
Luteal phase length when the animals had been on the WSD for 14 mo correlated positively with SI (r = 0.909, P < 0.001). SI was also positively correlated with P4 AUC (r = 0.69, P = 0.014). There were no significant correlations between SI and follicular phase length, D1–3 E2, peak E2, or peak P4 (all P > 0.1).
There was a trend toward a main effect of time when basal metabolic rate (measured at night between 2300 and 0300) was compared from before with 1 yr after beginning WSD, with metabolic rate decreasing over this time period [F(1,10) = 4.192, P = 0.068]. There were no treatment effects on basal metabolic rate either before or after WSD consumption (before WSD: T treated, 1.50 ± 0.16 kcal·kg−1·h−1; control, 1.84 ± 0.37 kcal·kg−1·h−1; 1 yr WSD: T treated, 1.21 ± 0.08 kcal·kg−1·h−1; control, 1.23 ± 0.05 kcal·kg−1·h−1, P = 0.8). There were also no treatment effects on metabolic rate measured over the rest of the 24-h period (all P > 0.1, data not shown).
There was a significant effect of diet on body composition, since percent fat and percent central fat increased significantly and percent lean mass decreased significantly during 16 mo of WSD consumption (all P < 0.001; Table 1). There was no effect of T treatment on percent fat, percent central fat, or percent lean mass in monkeys before or 16 mo after starting the WSD.
Table 1.
Body composition parameters before and 16 mo after starting WSD
| Before WSD |
16 Months WSD |
|||||||
|---|---|---|---|---|---|---|---|---|
| Weight, kg | Fat, % | Central fat, % | Lean mass, % | Weight, kg | Fat, % | Central fat, % | Lean mass, % | |
| Control | 4.6 (0.2) | 1.5 (0.2) | 0.6 (0.1) | 95.4 (0.2) | 5.9 (0.3) | 14.9 (2.8) | 16.3 (3.9) | 81.3 (3.3) |
| T treated | 5.2 (0.1) | 1.7 (0.3) | 1.0 (0.3) | 95.2 (0.3) | 6.7 (0.4) | 19.3 (3.8) | 22.2 (4.9) | 76.0 (3.6) |
Data are presented as mean (SE). T, testosterone. There was a significant change in all parameters from before to after Western style diet (WSD), although there were no between-group differences.
DISCUSSION
The focus of this study was to examine the effects of chronic HA, with introduction of a WSD, on metabolic and reproductive parameters in female primates. The results may be relevant to considering the effects of higher-than-normal androgens in producing the PCOS phenotype in women, but not the ultimate (e.g., potentially genetic) causes of HA. The findings presented in this paper indicate that mildly elevated levels of androgen, coupled with consumption of a typical WSD, can lead to functional disturbances in the neuroendocrine, ovarian, and metabolic systems. After being on the WSD for 16 mo, all animals showed increases in body weight and percentage body fat, as expected. While on the WSD and over the course of development, the central neuroendocrine drive to the reproductive axis in the control (WSD alone) animals increased so that it was similar to that in the T-treated animals, indicating that T treatment led to earlier maturation of the hypothalamic pulse generator. However, during the period on WSD, LH pulse amplitude was diminished both with and without T treatment. The size and dynamics of the antral follicle pool in the ovaries were also altered by the WSD both with and without T exposure. T-treated monkeys also had significantly lower SI compared with controls, suggesting that T in combination with increased adiposity had a negative effect on metabolic function.
While consuming a WSD, there were no differences in early follicular phase LH pulse frequency or LH pulse amplitude between the T-treated and control groups. Compared with measurements taken before animals started the WSD (59), LH pulse frequency increased in control animals but did not change in T-treated monkeys, such that monkeys from both groups had approximately hourly LH pulses after 6 mo on the WSD, and maintained this pulse frequency after 14 mo on this diet. Women with PCOS, who have elevated androgens, consistently have about 1 LH pulse/h (86), whereas in healthy women, LH pulse frequencies during the early follicular phase have been reported to be 1 pulse/90 min (4, 86) in some studies and 1 pulse/h in other studies (44, 70). However, most studies that were performed in normally cycling, nonexperimental, female monkeys detected 1 LH pulse/h during the early follicular phase (38, 64). Thus, it appears that, although T treatment led to an earlier activation of the LH pulse generator in monkeys (59), this difference was not maintained as monkeys matured into adulthood, and was not exacerbated by consuming a WSD. Similarly, in adolescents with HA, the transition from pubertal to adult patterns of LH secretion occurs about 2.5 yr earlier than in control girls, even with no difference in age at menarche (3).
Notably, LH pulse amplitude significantly decreased in both control and T-treated monkeys within 6 mo of beginning the WSD, and remained lower after 14 mo. This finding is consistent with evidence from PCOS patients indicating that obesity is associated with a decrease in LH pulse amplitude (4, 66, 84), although not all studies have found such a change (27). Studies that found a negative correlation between BMI and LH pulse amplitude in women with PCOS did not find correlations between BMI and LH pulse frequency (66, 84). Therefore, the theory was put forth that obesity acts to modulate LH release at the pituitary rather than at the hypothalamic level (4, 66). In the T-treated animals, pulse frequency was not significantly changed even though pulse amplitude decreased while on the WSD, which would support the theory that obesity modulates LH release at the level of the pituitary. Nevertheless, we note that, in this study, there were no monkeys that continued eating normal monkey chow; thus, we cannot definitively determine if obesity was associated with a decrease in LH pulse amplitude or whether the decrease in amplitude was a maturational change occurring in early adulthood. Likewise, since data on regional fat distribution were only collected at the start and after 16 mo of WSD, as opposed to weekly weight analysis, LH pulse frequency/amplitude and other parameters were only related to weight. Ongoing studies, with control and T-treated monkeys on both normal diet and WSD throughout development, will be able to address these issues.
Neither LH responsiveness to GnRH nor the effect of P4 negative feedback on pulsatile LH secretion was different between the T-treated and control groups during consumption of the WSD. Decreased sensitivity to P4 feedback is seen in about one-half of PCOS patients and HA adolescents (22, 68), and has also been reported in adult female sheep that were treated prenatally with T (72). Visual inspection of the current data (see Fig. 3) reveals that there are several animals that did not have a decrease in pulse number between D1–3 and the luteal phase of a menstrual cycle; however, these animals are distributed between the T-treated and control groups, so decreased sensitivity to P4 negative feedback was not more prevalent in T-treated compared with control animals. It is also unlikely that there was an effect of the increased adiposity from the WSD on the sensitivity of the hypothalamic pulse generator to P4 negative feedback, since the animals that had no change in LH pulse frequency were not the animals that gained the most weight or had the highest percent fat after switching to the WSD (data not shown). It is possible that the reduced sensitivity to P4 feedback seen in PCOS patients is not due to the direct actions of T, but is instead caused by entrainment of the hypothalamic-pituitary axis during development, which could explain why we did not see group differences in this study.
Changes in ovarian structure and function were apparent in both the control and T-treated groups after 14 mo of WSD. The animals had shorter follicular phases, which were accompanied by lower levels of E2 on D1–3 of the menstrual cycle. Low E2 during the follicular phase is associated with luteal phase defects (LPD) in women (26), and the current data showing that T-treated monkeys on WSD also had lower P4 levels in the luteal phase suggest that androgen exacerbates these abnormalities. Notably, the reduced E2 levels in the early follicular phase suggest that the modest levels of exogenous T do not serve as a major substrate for estrogen production. This was tested during our initial studies (Ref. 59, unpublished data) before introducing the WSD protocol. Measurement of circulating E2 before and after T implantation indicated no differences. This does not rule out some elevation in local E2 in ovarian compartments where aromatization occurs, especially in the dominant follicle. However, no differences in E2 levels were noted between treatment groups at midcycle. Future studies will also examine the effects of both WSD and T on steroidogenesis within the ovary/dominant follicle.
Additionally, the monkeys displayed an increased number of small antral follicles and a decrease in antral follicle size during the early follicular phase after WSD consumption. Alterations in follicular dynamics were also evident during the late follicular phase, when changes were more pronounced in the T-treated group. We previously used ultrasound studies to document changes to the antral follicle cohort during the follicular phase in rhesus macaques (12). In adult rhesus females on a normal diet with normal menstrual cycles, the largest antral follicle is typically larger (4.36 ± 0.63 mm) during the late follicular phase than was found in either the control or T-treated group during the current study (see Fig. 6B). Additionally, our previous study found that the dominant follicle had a cohort of only three to four other antral follicles during the late follicular phase (12) compared with the average cohort of seven follicles found in this study (Fig. 6A). Androgen exposure was previously shown to increase antral follicle formation when given in high doses to adult rhesus monkeys for 3–10 days (85). Our study indicates that long-term, mildly increased androgen exposure compounded the effect of the WSD to alter follicular growth dynamics, either delaying or suppressing the growth of the dominant antral follicle and increasing the number of small antral follicles present.
After 14 mo on the WSD, T-treated animals had a lower peak P4 value and lower P4 AUC during the luteal phase compared with control animals, which could have resulted from defects in the dominant antral follicle, as discussed above. Low levels of P4 during the luteal phase in women are indicative of LPD (25, 77). LPD have been associated with abnormal gonadotropin secretion (25, 40, 77, 78), poor follicular quality (26), and stress during the follicular phase of the menstrual cycle (89). Women with PCOS were found to have lower levels of P4 than controls during the luteal phase after either spontaneous or induced ovulation, indicative of the presence of LPD in these women (60). Similarly, Goy and Robinson (36) reported an increase in LPD in female monkeys that were given high doses of androgens prenatally, even though ovulation rates were not different from control monkeys. Although luteal phase P4 was lower in the T-treated monkeys in the current study, it was not low enough to cause premature menstruation, and thus may have allowed implantation if a fertilized ovum were present. However, a recent report of P4 resistance in endometrial tissue in women with PCOS (74) suggests that further studies are warranted to determine if lower P4 secretion or uterine dysfunction plays a part in PCOS-associated infertility.
Metabolic changes were evident after the monkeys had been eating the WSD for 12–16 mo. As expected, weight, percent fat, and percent central fat all increased with consumption of the WSD, and percent lean mass decreased. However, there was no difference in percent central fat between T-treated and control animals. Higher T levels are associated with increased central adiposity in women (46, 55), and antiandrogen treatment reportedly decreases visceral adiposity (35, 55). Adult female rhesus monkeys that were treated prenatally with T also had increased abdominal fat mass compared with weight- and BMI-matched controls (30). Thus, androgens are likely the cause of increased central fat, as opposed to the increased central adiposity leading to increased levels of androgens. We therefore expected the T-treated monkeys in the current study to have higher levels of central adiposity compared with the control animals, but this was not the case. It is possible, however, that, if the animals remained on the WSD for a longer period of time and continued to gain weight, a group difference might have emerged.
Total postpubertal weight gain was higher in T-treated animals compared with controls. Although T-treated monkeys did not eat more calories than control animals, there was a strong trend toward the T-treated animals being less active than controls during the last 10 mo of this study. Studies in humans and animals found that individuals that are less active tend to gain more weight (17, 47, 82), which is consistent with the current data. Because activity levels were not assessed in our prior study (59) before T implants were placed, it is unknown if the T-treated animals had lower levels of activity at baseline or whether the decreased activity levels occurred later as a result of chronic T treatment.
Metabolic changes were also evident between the control and T-treated groups after 12 mo on the WSD. SI scores were significantly lower in T-treated compared with control animals, even after controlling for weight. Similarly, women with PCOS have lower SI than weight-matched controls, and this relationship is present even in lean PCOS patients (20, 28). However, we did not find any effect of T on SI when the monkeys were tested at 3.5 yr of age, before starting the WSD. This suggests that, while initial T treatment alone did not lead to decreased SI in this study, T treatment in combination with adiposity had a more detrimental effect on SI than adiposity alone. Alternatively, more prolonged T treatment until 6.5 yr of age may have led to decreased SI in the absence of WSD consumption. The trend toward increased insulin levels during WSD exposure is consistent with those observed in rhesus females with ∼20% body fat in the ONPRC colony (54). Unfortunately, there is a paucity of data from age-matched “lean” (3% fat) females as young adults. Ongoing studies using larger cohorts of female monkeys with and without WSD should clarify these data.
Studies by Abbott and colleagues (1) observed insulin resistance and hyperinsulinemia in all monkeys they studied that had a high BMI (over 41.2 kg/m2) in adulthood. However, animals that were prenatally androgenized in addition to being obese and insulin resistant were more likely to be anovulatory than animals that were obese but not exposed to T during gestation. The average BMI after 16 mo on the WSD in the current study was 25.6 kg/m2, and the highest was 34.6 kg/m2, so it is possible that some of the T-treated animals may have become anovulatory at a later time if they continued to gain weight on the WSD.
Although we did not see any effect of T treatment on ovulation rates in the current study, there was a relationship between metabolic and ovarian parameters. The animals with lower SI also had shorter luteal phases, which can be an early indicator of LPD (77). Decreased SI and anovulation are both common in women with PCOS, and, when these women ovulate, they often have LPD (60). Thus, it is interesting that the animals with decreased SI were the same animals that had shorter luteal phases in this study, and it is possible that the T-treated monkeys would develop higher rates of anovulation with continued exposure to elevated T.
There was a decrease in basal metabolic rate from before to 12 mo after the animals began eating the WSD. This was likely due to aging, since metabolic rate declines during puberty in people (16) and continues to decline with age (37, 71, 73). The decrease in metabolic rate also could have resulted from a decrease in activity, since lower activity is associated with a decreased metabolic rate (52, 69). The lack of differences in metabolic rate between T-treated and control animals indicates that the change in metabolic rate over time was likely not attributable to T.
Obesity was associated with increased levels of free T in women in some (31, 79, 83), but not all (14), studies. We did not see an increase in total T or free T, or a decrease in SHBG in the monkeys as they gained weight on the WSD. Thus, the effects of WSD that we report in control monkeys (i.e., weight gain and changes in ovarian structure) are unlikely to be a result of increased circulating androgen.
In summary, although increased adiposity as a result of the WSD did not appear to act in concert with T treatment to aggravate neuroendocrine dysfunction, T + WSD treatment reduced SI, increased the number and reduced the maximal size of antral follicles at midcycle, and reduced P4 secretion during the luteal phase of the menstrual cycle compared with controls. Thus, prolonged elevation of T combined with WSD can lead to impairment of metabolic and ovarian function. This dysfunction may have resulted from the extended length of T treatment, or the combination of T and WSD. However, since this project was a preliminary study, resources were not available to implement the optimal experimental design, which would have included both control and T-treated groups that were fed normal monkey chow, as well as control and T-treated groups that received WSD. Without these non-WSD groups, it is difficult to definitively state the role of the WSD (vs. age and length of T treatment) with regard to changes in SI and ovarian function in the T-treated group in the current study. It may have been that T treatment alone for a prolonged period would have led both to luteal dysfunction and reductions in SI. Ongoing studies that include groups of both T-treated and control monkeys maintained on both normal monkey chow and WSD should clarify the role of adiposity compared with the role of elevated androgen on the neuroendocrine, reproductive, and metabolic parameters that were measured in the current study, which may have relevance to the PCOS phenotype in women.
GRANTS
This study was funded by a grant from the OHSU Center for Women's Health Circle of Giving, pilot funds from National Institutes of Health (NIH) Grant P51-OD-11092, and Eunice Kennedy Shriver National Institute of Child Health and Human Development/NIH through cooperative agreements U54-HD-18185, U54-HD-28934, and U54-HD-12303 as part of the Specialized Cooperative Centers Program in Reproduction and Infertility Research, as well as by an R21 grant from NIH (RR-030276). W. K. McGee was supported by a fellowship from the OHSU ARCS Foundation and NIH Grant T32-HD-07133.
DISCLOSURES
The authors have nothing to disclose.
AUTHOR CONTRIBUTIONS
Author contributions: W.M., J.C., J.C.M., R.L.S., and J.L.C. conception and design of research; W.M. and C.B. performed experiments; W.M., C.B., C.P., and F.K.Y.P. analyzed data; W.M., C.B., C.P., J.C., J.C.M., R.L.S., and J.L.C. interpreted results of experiments; W.M. and C.B. prepared figures; W.M. drafted manuscript; W.M., C.B., J.C., J.C.M., F.K.Y.P., R.L.S., and J.L.C. edited and revised manuscript; W.M., C.B., C.P., J.C., J.C.M., F.K.Y.P., R.L.S., and J.L.C. approved final version of manuscript.
ACKNOWLEDGMENTS
We thank Mandi Bulechowsky for outstanding technical assistance and express appreciation to the ONPRC Department of Comparative Medicine, veterinary and surgical staff for the excellent care of the animals in this study. Gratitude is expressed to the ONPRC Endocrine Technology Support Core Laboratory and the Assay Core of the Center for Reproductive Physiology at the University of Pittsburgh for assistance with hormone assays. We also thank Dr. Elinor Sullivan for consultations regarding metabolic measurements made in this study.
REFERENCES
- 1.Abbott DH, Dumesic DA, Eisner JR, Kemnitz JW, Goy RW. The prenatally androgenized female rhesus monkey as a model for PCOS. In: Androgen Excess Disorders in Women, edited by Azziz R, Nestler JE, Dewailly D. Philadelphia, PA: Lippincott-Raven, 1997 [Google Scholar]
- 2.Acien P, Quereda F, Matallin P, Villarroya E, Lopez-Fernandez JA, Acien M, Mauri M, Alfayate R. Insulin, androgens, and obesity in women with and without polycystic ovary syndrome: a heterogeneous group of disorders. Fertil Steril 72: 32–40, 1999 [DOI] [PubMed] [Google Scholar]
- 3.Apter D, Butzow T, Laughlin GA, Yen SS. Accelerated 24-hour luteinizing hormone pulsatile activity in adolescent girls with ovarian hyperandrogenism: relevance to the developmental phase of polycystic ovarian syndrome. J Clin Endocrinol Metab 79: 119–125, 1994 [DOI] [PubMed] [Google Scholar]
- 4.Arroyo A, Laughlin GA, Morales AJ, Yen SS. Inappropriate gonadotropin secretion in polycystic ovary syndrome: influence of adiposity. J Clin Endocrinol Metab 82: 3728–3733, 1997 [DOI] [PubMed] [Google Scholar]
- 5.Asuncion M, Calvo RM, San Millan JL, Sancho J, Avila S, Escobar-Morreale HF. A prospective study of the prevalence of the polycystic ovary syndrome in unselected Caucasian women from Spain. J Clin Endocrinol Metab 85: 2434–2438, 2000 [DOI] [PubMed] [Google Scholar]
- 6.Azziz R, Sanchez LA, Knochenhauer ES, Moran C, Lazenby J, Stephens KC, Taylor K, Boots LR. Androgen excess in women: experience with over 1000 consecutive patients. J Clin Endocrinol Metab 89: 453–462, 2004 [DOI] [PubMed] [Google Scholar]
- 7.Bachelot A, Laborde K, Bresson JL, Plu-Bureau G, Raynaud A, Bertagna X, Mogenet A, Mansour M, Lucas-Jouy V, Gayno JP, Reznik Y, Kuhn JM, Billaud L, Vacher-Lavenu MC, Putterman M, Mowszowicz I, Touraine P, Kuttenn F. Luteinizing hormone pulsatility in patients with major ovarian hyperandrogenism. J Endocrinol Invest 30: 636–646, 2007 [DOI] [PubMed] [Google Scholar]
- 8.Balen AH, Conway GS, Kaltsas G, Techatrasak K, Manning PJ, West C, Jacobs HS. Polycystic ovary syndrome: the spectrum of the disorder in 1741 patients. Hum Reprod 10: 2107–2111, 1995 [DOI] [PubMed] [Google Scholar]
- 9.Barber TM, Alvey C, Greenslade T, Gooding M, Barber D, Smith R, Marland A, Wass JA, Child T, McCarthy MI, Franks S, Golding SJ. Patterns of ovarian morphology in polycystic ovary syndrome: a study utilising magnetic resonance imaging. Eur Radiol 20: 1207–1213, 2010 [DOI] [PubMed] [Google Scholar]
- 10.Bergman RN. Toward physiological understanding of glucose tolerance (Abstract). Diabetes 38: 16, 1989 [DOI] [PubMed] [Google Scholar]
- 11.Bergman RN, Ider YZ, Bowden CR, Cobelli C. Quantitative estimation of insulin sensitivity. Am J Physiol Endocrinol Metab Gastrointest Physiol 236: E667–E677, 1979 [DOI] [PubMed] [Google Scholar]
- 12.Bishop CV, Sparman ML, Stanley JE, Bahar A, Zelinski MB, Stouffer RL. Evaluation of antral follicle growth in the macaque ovary during the menstrual cycle and controlled ovarian stimulation by high-resolution ultrasonography. Am J Primatol 71: 384–392, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Blank SK, McCartney CR, Marshall JC. The origins and sequelae of abnormal neuroendocrine function in polycystic ovary syndrome. Human Repro Update 12: 351–361, 2006 [DOI] [PubMed] [Google Scholar]
- 14.Bordini B, Littlejohn E, Rosenfield RL. Blunted sleep-related luteinizing hormone rise in healthy premenarcheal pubertal girls with elevated body mass index. J Clin Endocrinol Metab 94: 1168–1175, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Boston RC, Stefanovski D, Moate PJ, Sumner AE, Watanabe RM, Bergman RN. MINMOD Millennium: a computer program to calculate glucose effectiveness and insulin sensitivity from the frequently sampled intravenous glucose tolerance test. Diabetes Technol Ther 5: 1003–1015, 2003 [DOI] [PubMed] [Google Scholar]
- 16.Brown DC, Kelnar CJ, Wu FC. Energy metabolism during male human puberty. I. Changes in energy expenditure during the onset of puberty in boys. Ann Hum Biol 23: 273–279, 1996 [DOI] [PubMed] [Google Scholar]
- 17.Brown WJ, Williams L, Ford JH, Ball K, Dobson AJ. Identifying the energy gap: magnitude and determinants of 5-year weight gain in midage women. Obes Res 13: 1431–1441, 2005 [DOI] [PubMed] [Google Scholar]
- 18.Buckett WM, Bouzayen R, Watkin KL, Tulandi T, Tan SL. Ovarian stromal echogenicity in women with normal and polycystic ovaries. Hum Reprod 14: 618–621, 1999 [DOI] [PubMed] [Google Scholar]
- 19.Cameron JL, Nosbisch C. Suppression of pulsatile luteinizing hormone and testosterone secretion during short term food restriction in the adult male rhesus monkey (Macaca mulatta). Endocrinology 128: 1532–1540, 1991 [DOI] [PubMed] [Google Scholar]
- 20.Carmina E, Bucchieri S, Esposito A, Del Puente A, Mansueto P, Orio F, Di Fede G, Rini G. Abdominal fat quantity and distribution in women with polycystic ovary syndrome and extent of its relation to insulin resistance. J Clin Endocrinol Metab 92: 2500–2505, 2007 [DOI] [PubMed] [Google Scholar]
- 21.Carmina E, Koyama T, Chang L, Stanczyk FZ, Lobo RA. Does ethnicity influence the prevalence of adrenal hyperandrogenism and insulin resistance in polycystic ovary syndrome? Am J Obstet Gynecol 167: 1807–1812, 1992 [DOI] [PubMed] [Google Scholar]
- 22.Chhabra S, McCartney CR, Yoo RY, Eagleson CA, Chang RJ, Marshall JC. Progesterone inhibition of the hypothalamic gonadotropin-releasing hormone pulse generator: evidence for varied effects in hyperandrogenemic adolescent girls. J Clin Endocrinol Metab 90: 2810–2815, 2005 [DOI] [PubMed] [Google Scholar]
- 23.Chuang SY, Chou P, Hsu PF, Cheng HM, Tsai ST, Lin IF, Chen CH. Presence and progression of abdominal obesity are predictors of future high blood pressure and hypertension. Am J Hypertens 19: 788–795, 2006 [DOI] [PubMed] [Google Scholar]
- 24.Clark MK, Dillon JS, Sowers M, Nichols S. Weight, fat mass, and central distribution of fat increase when women use depot-medroxyprogesterone acetate for contraception. Int J Obes (Lond) 29: 1252–1258, 2005 [DOI] [PubMed] [Google Scholar]
- 25.Cook CL, Rao CV, Yussman MA. Plasma gonadotropin and sex steroid hormone levels during early, midfollicular, and midluteal phases of women with luteal phase defects. Fertil Steril 40: 45–48, 1983 [DOI] [PubMed] [Google Scholar]
- 26.DiZerega GS, Hodgen GD. Luteal phase dysfunction infertility: a sequel to aberrant folliculogenesis. Fertil Steril 35: 489–499, 1981 [DOI] [PubMed] [Google Scholar]
- 27.Dunaif A, Mandeli J, Fluhr H, Dobrjansky A. The impact of obesity and chronic hyperinsulinemia on gonadotropin release and gonadal steroid secretion in the polycystic ovary syndrome. J Clin Endocrinol Metab 66: 131–139, 1988 [DOI] [PubMed] [Google Scholar]
- 28.Dunaif A, Segal KR, Futterweit W, Dobrjansky A. Profound peripheral insulin resistance, independent of obesity, in polycystic ovary syndrome. Diabetes 38: 1165–1174, 1989 [DOI] [PubMed] [Google Scholar]
- 29.Eagleson CA, Bellows AB, Hu K, Gingrich MB, Marshall JC. Obese patients with polycystic ovary syndrome: evidence that metformin does not restore sensitivity of the gonadotropin-releasing hormone pulse generator to inhibition by ovarian steroids. J Clin Endocrinol Metab 88: 5158–5162, 2003 [DOI] [PubMed] [Google Scholar]
- 30.Eisner JR, Dumesic DA, Kemnitz JW, Colman RJ, Abbott DH. Increased adiposity in female rhesus monkeys exposed to androgen excess during early gestation. Obes Res 11: 279–286, 2003 [DOI] [PubMed] [Google Scholar]
- 31.Evans DJ, Hoffmann RG, Kalkhoff RK, Kissebah AH. Relationship of androgenic activity to body fat topography, fat cell morphology, and metabolic aberrations in premenopausal women. J Clin Endocrinol Metab 57: 304–310, 1983 [DOI] [PubMed] [Google Scholar]
- 32.Faure N, Prat X, Bastide A, Lemay A. Assessment of ovaries by magnetic resonance imaging in patients presenting with polycystic ovarian syndrome. Hum Reprod 4: 468–472, 1989 [DOI] [PubMed] [Google Scholar]
- 33.Flegal KM, Carroll MD, Ogden CL, Curtin LR. Prevalence and trends in obesity among US adults, 1999–2008. J Am Med Assoc 303: 235–241, 2010 [DOI] [PubMed] [Google Scholar]
- 34.Franks S. Adult polycystic ovary syndrome begins in childhood. Best Pract Res Clin Endocrinol Metab 16: 263–272, 2002 [DOI] [PubMed] [Google Scholar]
- 35.Gambineri A, Patton L, Vaccina A, Cacciari M, Morselli-Labate AM, Cavazza C, Pagotto U, Pasquali R. Treatment with flutamide, metformin, and their combination added to a hypocaloric diet in overweight-obese women with polycystic ovary syndrome: a randomized, 12-month, placebo-controlled study. J Clin Endocrinol Metab 91: 3970–3980, 2006 [DOI] [PubMed] [Google Scholar]
- 36.Goy RW, Robinson JA. Prenatal exposure of rhesus monkeys to patent androgens: morphological, behavioral, and physiological consequences. Banbury Report 11: 355–378, 1982 [Google Scholar]
- 37.Henry CJ. Mechanisms of changes in basal metabolism during ageing. Eur J Clin Nutr 54, Suppl 3: S77–S91, 2000 [DOI] [PubMed] [Google Scholar]
- 38.Herod SM, Pohl CR, Cameron JL. Treatment with a CRH-R1 antagonist prevents stress-induced suppression of the central neural drive to the reproductive axis in female macaques. Am J Physiol Endocrinol Metab 300: E19–E27, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Holte J, Bergh T, Berne C, Lithell H. Serum lipoprotein lipid profile in women with the polycystic ovary syndrome: relation to anthropometric, endocrine and metabolic variables. Clin Endocrinol (Oxf) 41: 463–471, 1994 [DOI] [PubMed] [Google Scholar]
- 40.Homburg R, Armar NA, Eshel A, Adams J, Jacobs HS. Influence of serum luteinising hormone concentrations on ovulation, conception, and early pregnancy loss in polycystic ovary syndrome. Br Med J 297: 1024–1026, 1988 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hunnell NA, Rockcastle NJ, McCormick KN, Sinko LK, Sullivan EL, Cameron JL. Physical activity of adult female rhesus monkeys (Macaca mulatta) across the menstrual cycle. Am J Physiol Endocrinol Metab 292: E1520–E1525, 2007 [DOI] [PubMed] [Google Scholar]
- 42.Johnstone EB, Rosen MP, Neril R, Trevithick D, Sternfeld B, Murphy R, Addauan-Andersen C, McConnell D, Pera RR, Cedars MI. The polycystic ovary post-Rotterdam: A common, age-dependent finding in ovulatory women without metabolic significance. JCEM 95: 4965–4972, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kauffman RP, Baker VM, Dimarino P, Gimpel T, Castracane VD. Polycystic ovarian syndrome and insulin resistance in white and Mexican American women: a comparison of two distinct populations. Am J Obstet Gynecol 187: 1362–1369, 2002 [DOI] [PubMed] [Google Scholar]
- 44.Kazer RR, Kessel B, Yen SS. Circulating luteinizing hormone pulse frequency in women with polycystic ovary syndrome. J Clin Endocrinol Metab 65: 233–236, 1987 [DOI] [PubMed] [Google Scholar]
- 45.Kiddy DS, Sharp PS, White DM, Scanlon MF, Mason HD, Bray CS, Polson DW, Reed MJ, Franks S. Differences in clinical and endocrine features between obese and non-obese subjects with polycystic ovary syndrome: an analysis of 263 consecutive cases. Clin Endocrinol (Oxf) 32: 213–220, 1990 [DOI] [PubMed] [Google Scholar]
- 46.Kirchengast S, Huber J. Body composition characteristics and body fat distribution in lean women with polycystic ovary syndrome. Hum Reprod 16: 1255–1260, 2001 [DOI] [PubMed] [Google Scholar]
- 47.Klesges RC, Klesges LM, Haddock CK, Eck LH. A longitudinal analysis of the impact of dietary intake and physical activity on weight change in adults. Am J Clin Nutr 55: 818–822, 1992 [DOI] [PubMed] [Google Scholar]
- 48.Knochenhauer ES, Key TJ, Kahsar-Miller M, Waggoner W, Boots LR, Azziz R. Prevalence of the polycystic ovary syndrome in unselected black and white women of the southeastern United States: a prospective study. J Clin Endocrinol Metab 83: 3078–3082, 1998 [DOI] [PubMed] [Google Scholar]
- 49.Knudsen KL, Blank SK, Burt Solorzano C, Patrie JT, Chang RJ, Caprio S, Marshall JC, McCartney CR. Hyperandrogenemia in obese peripubertal girls: correlates and potential etiological determinants. Obesity (Silver Spring) 18: 2118–2124, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kristensen SL, Ramlau-Hansen CH, Ernst E, Olsen SF, Bonde JP, Vested A, Toft G. A very large proportion of young Danish women have polycystic ovaries: is a revision of the Rotterdam criteria needed? Hum Reprod 25: 3117–3122, 2010 [DOI] [PubMed] [Google Scholar]
- 51.Lapidus L, Bengtsson C, Larsson B, Pennert K, Rybo E, Sjostrom L. Distribution of adipose tissue and risk of cardiovascular disease and death: a 12 year follow up of participants in the population study of women in Gothenburg, Sweden. Br Med J (Clin Res Ed) 289: 1257–1261, 1984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lawson S, Webster JD, Pacy PJ, Garrow JS. Effect of a 10-week aerobic exercise programme on metabolic rate, body composition and fitness in lean sedentary females. Br J Clin Pract 41: 684–688, 1987 [PubMed] [Google Scholar]
- 53.Legro RS. The genetics of obesity. Lessons for polycystic ovary syndrome. Ann NY Acad Sci 900: 193–202, 2000 [DOI] [PubMed] [Google Scholar]
- 54.Li S, Kievit P, Robertson A, Kolumam G, Xiumin L, von Wachenfeldt K, Valfridsson C, Bullens S, Messaoudi I, Bader L, Cowan KJ, Kamath A, van Bruggen N, Bunting S, Frendeus B, Grove KL. Targeting oxidized LDL improves insulin sensitivity and immune cell function in obese Rhesus macaques. Mol Metab 2: 256–269, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lovejoy JC, Bray GA, Bourgeois MO, Macchiavelli R, Rood JC, Greeson C, Partington C. Exogenous androgens influence body composition and regional body fat distribution in obese postmenopausal women–a clinical research center study. J Clin Endocrinol Metab 81: 2198–2203, 1996 [DOI] [PubMed] [Google Scholar]
- 56.McCartney CR, Eagleson CA, Marshall JC. Regulation of gonadotropin secretion: implications for polycystic ovary syndrome. Semin Reprod Med 20: 317–326, 2002 [DOI] [PubMed] [Google Scholar]
- 57.McCartney CR, Prendergast KA, Blank SK, Helm KD, Chhabra S, Marshall JC. Maturation of luteinizing hormone (gonadotropin-releasing hormone) secretion across puberty: evidence for altered regulation in obese peripubertal girls. J Clin Endocrinol Metab 94: 56–66, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.McCartney CR, Prendergast KA, Chhabra S, Eagleson CA, Yoo R, Chang RJ, Foster CM, Marshall JC. The association of obesity and hyperandrogenemia during the pubertal transition in girls: obesity as a potential factor in the genesis of postpubertal hyperandrogenism. J Clin Endocrinol Metab 91: 1714–1722, 2006 [DOI] [PubMed] [Google Scholar]
- 59.McGee WK, Bishop CV, Bahar A, Pohl CR, Chang RJ, Marshall JC, Pau FK, Stouffer RL, Cameron JL. Elevated androgens during puberty in female rhesus monkeys lead to increased neuronal drive to the reproductive axis: a possible component of polycystic ovary syndrome. Hum Reprod 27: 531–540, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Meenakumari KJ, Agarwal S, Krishna A, Pandey LK. Effects of metformin treatment on luteal phase progesterone concentration in polycystic ovary syndrome. Braz J Med Biol Res 37: 1637–1644, 2004 [DOI] [PubMed] [Google Scholar]
- 61.Meirow D, Yossepowitch O, Rosler A, Brzezinski A, Schenker JG, Laufer N, Raz I. Insulin resistant and non-resistant polycystic ovary syndrome represent two clinical and endocrinological subgroups. Hum Reprod 10: 1951–1956, 1995 [DOI] [PubMed] [Google Scholar]
- 62.Merriam GR, Wachter KW. Algorithms for the study of episodic hormone secretion. Am J Physiol Endocrinol Metab 243: E310–E318, 1982 [DOI] [PubMed] [Google Scholar]
- 63.Moran C, Renteria JL, Moran S, Herrera J, Gonzalez S, Bermudez JA. Obesity differentially affects serum levels of androstenedione and testosterone in polycystic ovary syndrome. Fertil Steril 90: 2310–2317, 2008 [DOI] [PubMed] [Google Scholar]
- 64.Norman RL, Lindstrom SA, Bangsberg D, Ellinwood WE, Gliessman P, Spies HG. Pulsatile secretion of luteinizing hormone during the menstrual cycle of rhesus macaques. Endocrinology 115: 261–266, 1984 [DOI] [PubMed] [Google Scholar]
- 65.Ozgun MT, Uludag S, Oner G, Batukan C, Aygen EM, Sahin Y. The influence of obesity on ICSI outcomes in women with polycystic ovary syndrome. J Obstet Gynaecol 31: 245–249, 2011 [DOI] [PubMed] [Google Scholar]
- 66.Pagan YL, Srouji SS, Jimenez Y, Emerson A, Gill S, Hall JE. Inverse relationship between luteinizing hormone and body mass index in polycystic ovarian syndrome: investigation of hypothalamic and pituitary contributions. J Clin Endocrinol Metab 91: 1309–1316, 2006 [DOI] [PubMed] [Google Scholar]
- 67.Pasquali R, Antenucci D, Casimirri F, Venturoli S, Paradisi R, Fabbri R, Balestra V, Melchionda N, Barbara L. Clinical and hormonal characteristics of obese amenorrheic hyperandrogenic women before and after weight loss. J Clin Endocrinol Metab 68: 173–179, 1989 [DOI] [PubMed] [Google Scholar]
- 68.Pastor CL, Griffin-Korf ML, Aloi JA, Evans WS, Marshall JC. Polycystic ovary syndrome: evidence for reduced sensitivity of the gonadotropin-releasing hormone pulse generator to inhibition by estradiol and progesterone. J Clin Endocrinol Metab 83: 582–590, 1998 [DOI] [PubMed] [Google Scholar]
- 69.Poehlman ET, Danforth E., Jr Endurance training increases metabolic rate and norepinephrine appearance rate in older individuals. Am J Physiol Endocrinol Metab 261: E233–E239, 1991 [DOI] [PubMed] [Google Scholar]
- 70.Reame N, Sauder SE, Kelch RP, Marshall JC. Pulsatile gonadotropin secretion during the human menstrual cycle: evidence for altered frequency of gonadotropin-releasing hormone secretion. J Clin Endocrinol Metab 59: 328–337, 1984 [DOI] [PubMed] [Google Scholar]
- 71.Roberts SB, Dallal GE. Energy requirements and aging. Public Health Nutr 8: 1028–1036, 2005 [DOI] [PubMed] [Google Scholar]
- 72.Robinson JE, Forsdike RA, Taylor JA. In utero exposure of female lambs to testosterone reduces the sensitivity of the gonadotropin-releasing hormone neuronal network to inhibition by progesterone. Endocrinology 140: 5797–5805, 1999 [DOI] [PubMed] [Google Scholar]
- 73.Ruggiero C, Metter EJ, Melenovsky V, Cherubini A, Najjar SS, Ble A, Senin U, Longo DL, Ferrucci L. High basal metabolic rate is a risk factor for mortality: the Baltimore Longitudinal Study of Aging. J Gerontol A Biol Sci Med Sci 63: 698–706, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Savaris RF, Groll JM, Young SL, DeMayo FJ, Jeong JW, Hamilton AE, Giudice Lessey BALC. Progesterone resistance in PCOS endometrium: a microarray analysis in clomiphene citrate-treated and artificial menstrual cycles. J Clin Endocrinol Metab 96: 1737–1746, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Shadoan MK, Anthony MS, Rankin SE, Clarkson TB, Wagner JD. Effects of tibolone and conjugated equine estrogens with or without medroxyprogesterone acetate on body composition and fasting carbohydrate measures in surgically postmenopausal monkeys. Metabolism 52: 1085–1091, 2003 [DOI] [PubMed] [Google Scholar]
- 76.Silfen ME, Denburg MR, Manibo AM, Lobo RA, Jaffe R, Ferin M, Levine LS, Oberfield SE. Early endocrine, metabolic, and sonographic characteristics of polycystic ovary syndrome (PCOS): comparison between nonobese and obese adolescents. J Clin Endocrinol Metab 88: 4682–4688, 2003 [DOI] [PubMed] [Google Scholar]
- 77.Soules MR, McLachlan RI, Ek M, Dahl KD, Cohen NL, Bremner WJ. Luteal phase deficiency: characterization of reproductive hormones over the menstrual cycle. J Clin Endocrinol Metab 69: 804–812, 1989 [DOI] [PubMed] [Google Scholar]
- 78.Stouffer RL, Hodgen GD. Induction of luteal phase defects in rhesus monkeys by follicular fluid administration at the onset of the menstrual cycle. J Clin Endocrinol Metab 51: 669–671, 1980 [DOI] [PubMed] [Google Scholar]
- 79.Strain GW, Zumoff B, Miller LK, Rosner W. Sex difference in the effect of obesity on 24-hour mean serum gonadotropin levels. Horm Metab Res 35: 362–366, 2003 [DOI] [PubMed] [Google Scholar]
- 80.Sullivan EL, Daniels AJ, Koegler FH, Cameron JL. Evidence in female rhesus monkeys (Macaca mulatta) that nighttime caloric intake is not associated with weight gain. Obes Res 13: 2072–2080, 2005 [DOI] [PubMed] [Google Scholar]
- 81.Sullivan EL, Grayson B, Takahashi D, Robertson N, Maier A, Bethea CL, Smith MS, Coleman K, Grove KL. Chronic consumption of a high-fat diet during pregnancy causes perturbations in the serotonergic system and increased anxiety-like behavior in nonhuman primate offspring. J Neurosci 30: 3826–3830, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sullivan EL, Koegler FH, Cameron JL. Individual differences in physical activity are closely associated with changes in body weight in adult female rhesus monkeys (Macaca mulatta). Am J Physiol Regul Integr Comp Physiol 291: R633–R642, 2006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Taponen S, Martikainen H, Jarvelin MR, Laitinen J, Pouta A, Hartikainen AL, Sovio U, McCarthy MI, Franks S, Ruokonen A. Hormonal profile of women with self-reported symptoms of oligomenorrhea and/or hirsutism: Northern Finland birth cohort 1966 study. J Clin Endocrinol Metab 88: 141–147, 2003 [DOI] [PubMed] [Google Scholar]
- 84.Taylor AE, McCourt B, Martin KA, Anderson EJ, Adams JM, Schoenfeld D, Hall JE. Determinants of abnormal gonadotropin secretion in clinically defined women with polycystic ovary syndrome. J Clin Endocrinol Metab 82: 2248–2256, 1997 [DOI] [PubMed] [Google Scholar]
- 85.Vendola KA, Zhou J, Adesanya OO, Weil SJ, Bondy CA. Androgens stimulate early stages of follicular growth in the primate ovary. J Clin Invest 101: 2622–2629, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Waldstreicher J, Santoro NF, Hall JE, Filicori M, Crowley WF., Jr Hyperfunction of the hypothalamic-pituitary axis in women with polycystic ovarian disease: indirect evidence for partial gonadotroph desensitization. J Clin Endocrinol Metab 66: 165–172, 1988 [DOI] [PubMed] [Google Scholar]
- 87.Williams NI, Caston-Balderrama AL, Helmreich DL, Parfitt DB, Nosbisch C, Cameron JL. Longitudinal changes in reproductive hormones and menstrual cyclicity in cynomolgus monkeys during strenuous exercise training: abrupt transition to exercise-induced amenorrhea. Endocrinology 142: 2381–2389, 2001 [DOI] [PubMed] [Google Scholar]
- 88.Witchel SF. Puberty and polycystic ovary syndrome. Mol Cell Endocrinol 254–255: 146–153, 2006 [DOI] [PubMed] [Google Scholar]
- 89.Xiao E, Xia-Zhang L, Ferin M. Inadequate luteal function is the initial clinical cyclic defect in a 12-day stress model that includes a psychogenic component in the Rhesus monkey. J Clin Endocrinol Metab 87: 2232–2237, 2002 [DOI] [PubMed] [Google Scholar]
- 90.Zawadzki JK, Dunaif A. Diagnostic criteria for polycystic ovary syndrome: towards a rational approach. In: Polycystic Ovary Syndrome, edited by Dunaif A, Givens JR, Haseltine FP, Merriam GR. Boston, MA: Blackwell Scientific, 1992, p. 377–384 [Google Scholar]