

Abstract
Chronic alcohol abuse is associated with skeletal muscle myopathy. Previously, we demonstrated that chronic binge alcohol (CBA) consumption by rhesus macaques accentuates skeletal muscle wasting at end-stage of simian immunodeficiency virus (SIV) infection. A proinflammatory, prooxidative milieu and enhanced ubiquitin proteasome activity were identified as possible mechanisms leading to loss of skeletal muscle. The possibility that impaired regenerative capacity, as reflected by the ability of myoblasts derived from satellite cell (SCs) to differentiate into myotubes has not been examined. We hypothesized that the inflammation and oxidative stress in skeletal muscle from CBA animals impair the differentiation capacity of myoblasts to form new myofibers in in vitro assays. We isolated primary myoblasts from the quadriceps femoris of rhesus macaques that were administered CBA or isocaloric sucrose (SUC) for 19 mo. Proliferation and differentiation potential of cultured myoblasts were examined in vitro. Myoblasts from the CBA group had significantly reduced PAX7, MYOD1, MYOG, MYF5, and MEF2C expression. This was associated with decreased myotube formation as evidenced by Jenner-Giemsa staining and myonuclei fusion index. No significant difference in the proliferative ability, cell cycle distribution, or autophagy was detected between myoblasts isolated from CBA and SUC groups. Together, these results reflect marked dysregulation of myoblast myogenic gene expression and myotube formation, which we interpret as evidence of impaired skeletal muscle regenerative capacity in CBA-administered macaques. The contribution of this mechanism to alcoholic myopathy warrants further investigation.
Keywords: satellite cells, chronic binge alcohol, rhesus macaques, myoblasts, myogenic differentiation, gene expression
one of the most common pathologies associated with chronic alcohol consumption is skeletal muscle myopathy, which is independent of peripheral neuropathy, malnutrition, or liver disease (32). It occurs in almost 50% of alcoholics who can exhibit up to 30% reduction of skeletal muscle mass (33). Chronic alcoholic skeletal myopathy can result either from decreased muscle protein synthesis (29, 34) or accelerated muscle protein degradation (40) or a combination of both. Little attention has been directed to the function of adult muscle stem cells, satellite cells (SCs), in alcohol-induced myopathies. Because alcohol has been shown to impair stem cell function in other tissues, including the brain, liver, and bone marrow (2, 10, 31, 38), we hypothesized that together with an impairment of anabolic and catabolic pathways, alcohol's adverse effect on muscle mass and function also involves dysregulation of SC function to form new myofibers.
The resident population of muscle stem cells, the SCs, is mononucleated and located at the periphery of adult skeletal muscle fibers in intimate association with the plasma membrane (45). These cells are quiescent during adulthood but can enter into cell cycle upon stimulation to repair damaged muscle by proliferation, differentiation, and fusion with existing myofibers (39). Our previous studies with chronic binge alcohol (CBA)-fed, Simian immunodeficiency virus (SIV)-infected rhesus macaques have shown that chronic alcohol consumption promotes a prooxidative and proinflammatory skeletal muscle milieu in SIV-infected animals, and this is associated with accentuated wasting at end-stage SIV infection (21, 24). The inflammatory environment in skeletal muscle would be expected to activate quiescent SCs to proliferate and differentiate into myotubes to compensate for enhanced skeletal muscle proteolysis (39, 45). When there is skeletal muscle injury, such as exercise-induced muscle damage, two distinct subpopulations of macrophages sequentially invade the injured muscle. The first population secretes inflammatory cytokines, such as TNF-α and IL-1. Subsequently, the proliferation and differentiation of SCs for repair of the damaged muscle are facilitated by a second set of macrophages that secrete anti-inflammatory cytokines (45). Our previous studies have demonstrated that CBA administration to SIV-infected male macaques results in accentuated skeletal muscle upregulation of inflammatory cytokine expression and depletion of antioxidant capacity (21) compared with that of sucrose-administered SIV-infected macaques. Those findings suggest that CBA-associated chronic skeletal muscle inflammatory/oxidative milieu impairs the regenerative capacity of SCs. Thus, the aim of this study was to determine whether chronic alcohol administration alone impairs the ability of myoblasts to proliferate and differentiate into myotubes in vitro.
The SC marker, paired homeobox 7 (PAX7), is a transcription factor that is essential for adult SC survival. SC differentiation into myofibers is regulated by two families of transcription factors, the basic helix-loop-helix muscle regulatory factors (MRF), including myogenic differentiation 1 (MYOD1), myogenic factor 5 (MYF5), and myogenin (MYOG) (28), and myocyte enhancer factor 2 A-D (MEF2A-D). SCs start expressing these transcription factors when they are stimulated to differentiate. These MRFs subsequently activate transcription of muscle-specific structural and contractile genes, such as actin, myosin, and troponin (6, 45). The results from this study demonstrate that CBA negatively affects myogenic gene expression and impairs the ability of myoblasts to differentiate into myotubes in vitro. These results suggest that CBA administration impairs skeletal muscle regenerative capacity, which may contribute to the development of alcoholic myopathy.
MATERIALS AND METHODS
Experimental Protocol
All experiments were approved by the Institutional Animal Care and Use Committee at both Tulane National Primate Research Center (TNPRC) in Covington, LA, and Louisiana State University Health Sciences Center in New Orleans (LSUHSC-NO), LA, and adhered to National Institutes of Health's Guidelines for the Care and Use of Laboratory Animals. Four- to six-year-old male macaques (Macaca mulatta) obtained from TNPRC breeding colonies were used for the study. Age- and body-weight-matched animals were randomized to either CBA or isocaloric sucrose (SUC) groups. Primary myoblasts were isolated from a total of six animals, three each in the SUC and CBA groups. Animals were individually housed in a biosafety level 2 (BSL2) containment building. All cell-based experiments were performed in BSL2 laboratory facilities at LSUHSC-NO.
Animals were administered alcohol or sucrose intragastrically as previously described (5, 24). Briefly, animals were administered alcohol (13–14 g of ethanol per kilogram body wt per week; 30% wt/vol water) or sucrose for 19 mo via a surgically implanted catheter. This approach of intragastric delivery was selected to reduce experimental variability and ensure chronic binge-like intoxicating blood alcohol concentrations of 50–60 mM. Calories provided by alcohol and sucrose averaged 15% of total caloric intake. Animals were provided monkey chow ad libitum (Lab Fiber Plus Primate diet DT; PMI Nutrition International, St. Louis, MO) and supplemented with fruits, vitamins, and Noyes treats (Research Diets, New Brunswick, NJ).
Myoblast isolation and culture.
Primary myoblasts were isolated and expanded, according to previously published protocols with slight modifications (17, 18, 43). Briefly, skeletal muscle (quadriceps femoris) samples were obtained at necropsy and placed in 30 ml of DMEM (HyClone, Waltham, MA) media with penicillin, streptomycin, and fungizone (Life Technologies, Carlsbad, CA) on ice for transport. Approximately 125 mg of tissue was dissected, minced, and washed with media. Tissue was enzymatically disassociated with 0.05% trypsin for two 1 h treatments. The cells were then plated for 4–5 h on tissue culture plates for fibroblast separation. Nonadhered cells were collected and centrifuged at 500 g for 5 min, and cultured in Ham's F-12 with 10% FBS (Life Technologies) and 10 ng/ml human epidermal growth factor (hEGF; Life Technologies) in 100-mm dishes and grown to 70% confluence (passage zero, P0). Myoblasts were cultured on collagen I-coated plates (BD Biosciences, San Jose, CA) and frozen at each passage in FBS + 10% DMSO. All experiments were performed with cells from P4.
The myoblasts were identified on the basis of their expression of PAX7 and Integrin α7 (ITGA7) using flow cytometry, as previously reported (7). Myoblasts were cultured in proliferation media for 48 h and trypsinized and pelleted. The PerFix-nc kit (Roche, Indianapolis, IN) was used for permeabilization and staining with PAX7 (Abcam, Cambridge, MA) conjugated to PE-Cy7, and ITGA7 (Abcam) to APC-Cy7. The antibodies were conjugated to the respective fluorophores using commercially available conjugation kits (Abcam). Briefly, 5 μl of fixative was added to each sample, vortexed and incubated for 15 min at room temperature. PAX7 and ITGA7 antibodies were mixed with the permeabilizing buffer and incubated for 30 min at RT. The cells were then washed with 2 ml of wash buffer and resuspended in 0.5 ml of wash buffer for analysis within 24 h. Cells were acquired on a BD LSRII flow cytometer (BD Biosciences, San Jose, CA), and analysis was performed using FACSDIVA version 6.1.3 software (BD Biosciences).
Proliferation and differentiation of myoblasts.
For experiments performed during the proliferation phase, cells were cultured in Ham's-F12 media with 10% FBS and 10 ng/ml hEGF. Cells were seeded such that they would attain a 70% confluent state by days 6 and 7 of culture. Cells were harvested at 3, 5, and 7 days for cell cycle analysis, assessment of cell death, and gene expression analysis. On day 7, media were changed to differentiation media containing Ham's-F12 and 2% horse serum (Lonza, Allendale, NJ), and myoblasts were cultured an additional 7 days to assess differentiation potential.
Analytical Procedures
Total RNA isolation and real-time quantitative PCR.
Total RNA was extracted using the RNeasy mini universal kit (Qiagen, Valencia, CA), as per the manufacturer's instructions. cDNA was synthesized from 1 μg of the resulting total RNA using the Quantitect reverse transcriptase kit (Qiagen), in accordance with the manufacturer's instructions. Primers were designed to span exon-exon junctions and purchased from Life Technologies or IDT (Table 1) and used at a concentration of 500 nmol. The final reactions were made to a total volume of 20 μl with Quantitect SyBr Green PCR kit and DNase RNase-free water (Qiagen). All reactions were carried out in duplicate on a CFX96 system (Bio-Rad Laboratories, Hercules, CA) for quantitative PCR (qPCR) detection. qPCR data were analyzed using the comparative Ct (delta-delta-Ct, ΔΔCT) method. Target genes were compared with the endogenous control, ribosomal protein S13 (RPS13), and CBA values were normalized to SUC values.
Table 1.
qPCR primers (Macaca mulatta)
| Target gene | 5′ to 3′ | Primer Sequences |
|---|---|---|
| Autophagy related 5 (ATG5) | Forward | ACCAGAAACACTTCGCTGCT |
| Reverse | ATGATGGCAGTGGAGGAAAG | |
| Beclin 1 | Forward | AGGTTGAGAAAGGCGAGACA |
| Reverse | GCTTTTGTCCACTGCTCCTC | |
| Myocyte enhancer factor 2C (MEF2C) | Forward | GTCAATTGGGAGCTTGCACTAGCA |
| Reverse | TGGTACGGTCTCTAGGAGGAGAAA | |
| Myogenic differentiation 1 (MYOD1) | Forward | ACAGTGGGTGGTCATTCACA |
| Reverse | ACGGGTGTCATAGAAGTCG | |
| Myogenic factor 5 (MYF5) | Forward | AGCCCTACCTCCAACTGTTCTGAT |
| Reverse | AGGTTGCTCTGAGGAGGTGATCC | |
| Myogenin (MYOG) | Forward | TCAGCTCCCTCAACCAGGAGGA |
| Reverse | TGTGAGAGCTGCATTCGCTGG | |
| Myosin Heavy Chain 3 (MHC3) | Forward | AGCATGGTGAACCTGATCCACAGT |
| Reverse | TGAAGTTCAAGCAGAAGCAGCGAG | |
| Paired homeobox 7 (PAX7) | Forward | TCCTGGAAGAAGGTGGTTGAATGC |
| Reverse | CGTGTGCAGGTCTGGTTCAGTAA | |
| Ribosomal protein S13 (RPS13) | Forward | TCTGACGACGTGAAGGAGCAGATT |
| Reverse | TCTCTCAGGATCACACCGATTTGT |
Analysis of Cell Proliferation, Cell Cycle Progression, and Cell Death
Cell proliferation.
Cell proliferation was assessed using the Cyquant assay kit (Life Technologies), as per manufacturer's instructions (36). Briefly, cells were cultured in 96-well plates at a density of 3,000 cells/mm3 and at days 3 (d3) and 5 of proliferation were lysed with CyQUANT GR dye/lysis buffer and incubated for 5 min at room temperature in the dark. The fluorescence of each sample was measured with Infinite 200 Nanoquant microplate reader (Tecan, Durham, NC), with 485 nm excitation and 530-nm emission filters. For each experiment, a standard calibration curve was generated, and growth curves were plotted as fluorescence vs. time.
Cell cycle progression.
Cells were trypsinized and washed by centrifugation in PBS. The cells were then fixed in 70% ethanol at −20°C for at least 24 h. On the day of flow cytometric analysis, the ethanol-suspended cells were centrifuged for 5 min at 500 g, and ethanol was decanted. The cells were then suspended in PBS with 0.1% Triton X 100 for ∼1 min and then washed by centrifugation twice at 500 g for 5 min. The cells were then suspended in propidium iodide (PI) staining solution (10 μg/ml; Molecular Probes, Eugene, OR), and 100 μg/ml DNase-free RNase A in PBS and incubated for at least 30 min at 37°C and subjected to flow cytometry using a FACS Canto II system. The percentage of G1, S, and G2/M phases of the singlet cell population was calculated using MODFIT-LT software.
Cell death—apoptosis and necrosis.
Cells were trypsinized and washed with 1 ml annexin binding buffer (ABB; Invitrogen). They were centrifuged for 5 min at 500 g at 25°C, and the supernatant was removed and resuspended in 100 μl of ABB. To this suspension, 5 μl of annexin V-FITC (Life Technologies) and 1 μl of 100 μg/ml PI were added, and the cells were incubated for 15 min at room temperature in the dark. The cells were then washed in ABB and fixed for 10 min in 1% paraformaldehyde, washed with PBS, and immediately subjected to flow cytometric analysis. PI and annexin V single color cells were used as controls. All data were acquired with a Becton-Dickinson FACS Canto II system. Live statistics were used to align the X and Y mean values of the annexin V-FITC- or PI-stained quadrant populations by compensation. Quadrant markers were assigned using cell samples subjected to single color staining.
Immunocytochemistry
Cells were fixed in 2% paraformaldehyde, and immunocytochemical staining was performed using rabbit polyclonal anti-microtubule-associated protein 1A/1B-light chain 3 (LC3B) (1:1,000 for 1 h at room temperature; Life Technologies) as primary antibody and horseradish peroxidase-conjugated goat anti-rabbit IgG (1:200 for 1 h at room temperature; Life Technologies) as secondary antibody. 3,3′-diaminobenzidine (enhanced liquid substrate system; Sigma-Aldrich) was used as the chromogen substrate. Photomicrographs were taken using an Olympus BX51 microscope (Olympus, Center Valley, PA) under bright-field illumination. Control slides were prepared by replacing the primary antibody with the same concentration of normal serum.
Jenner Giemsa Staining for Assessment of Myotube Formation and Fusion Index
Cells were stained during the differentiation period for assessment of myotube formation and determination of the fusion index (42). Briefly, methanol-fixed cells were stained with Jenner staining solution (Electron Microscopy Sciences, Hatfield, PA) followed by Giemsa stain. Photomicrographs were taken using an Olympus BX51 microscope. Protein-rich myotubes have a darker purple color, and the nuclei stain pink. The fusion index was calculated as the % of nuclei that incorporated in myotubes over the total number of nuclei. For quantification of myotube density, images were taken at a resolution of 1360×1024 pixels in the red-green-blue (RGB) color mode. The images were converted into grayscale and analyzed using NISElements software (Nikon Instruments, Melville, NY). The mean number of pixels in each image attributed to tones 0–75 (darker end of spectrum) was calculated between replicates, and this is presented as the myotube density. The experiments were performed in triplicate.
Statistical Analysis
All data are presented as the means ± SE in each experimental group. For gene expression analysis, ANOVA using repeated measures was performed between the specific genes in the SUC and CBA groups. For the cell cycle analysis, Chi-squared analysis was performed. Statistical significance was set at two-sided P ≤ 0.05. Statistical analyses were performed using Prism 5 software (GraphPad Software, San Diego, CA).
RESULTS
Characterization of Myoblast Cultures
To determine the purity of myoblasts cultures, cells at passage 4 were subjected to flow cytometric analysis 48 h after plating. The percentage of cells that were positive for both PAX7 and ITGA7 were 90% in both CBA and SUC groups. Visually, cell cultures of myoblasts obtained from the CBA- and SUC-administered macaques appeared similar and reached 80% confluence at ∼6–7 days in proliferation media. However, during the differentiation phase, it appeared that the myotubes formed by myoblasts in the CBA group were not well organized and were shorter compared with the SUC group.
Myogenic Gene Expression in Myoblasts From CBA-Administered Macaques
The mRNA expression of myogenic genes was examined at days 3 (d3), 5 (d5), and 7 (d7) of the proliferation and differentiation phases. Myoblasts from CBA-administered macaques showed significant downregulation of PAX7, MYOD1, MYF5, MYOG, and MHC3 (Fig. 1).
Fig. 1.

Myogenic gene expression in myoblast cultures. mRNA expression of myogenic genes in myoblasts at day 3 (d3), d5, and d7 of proliferation (d3, d5, d7) and differentiation (d3d, d5d, d7d). PAX7 (A), MYOD1 (B), MYOG (C), MYF5 (D), and MHC3 (F) were significantly decreased in the chronic binge alcohol (CBA) vs. sucrose (SUC) group. PAX7 is a satellite cell marker. MYOD1, MYOG, MYF5 and MEF2C (E) are muscle regulatory factors, and MHC3 is a marker of maturation. $Significant (P < 0.05) effect of CBA on the respective mRNA expression. #Significant (P < 0.05) effect of time on the respective mRNA expression. *Significant interaction. Data are expressed as means ± SE.
Differentiation Potential of Myoblasts Isolated From CBA-Administered Macaques
The ultimate functional test of myoblasts is their ability to differentiate into myotubes in vitro rich in contractile protein. At d7 of culture, under conditions that promote differentiation, myotube density was markedly reduced in cultures of myoblasts isolated from CBA-administered group compared with those isolated from the SUC-administered group (Fig. 2A). Similar reduction in the number of nuclei that fused into multinucleated myofibers was observed at d7 in cultures of myoblasts isolated from CBA-administered animals (Fig. 2B).
Fig. 2.

In vitro differentiation potential of myoblasts. Images of Jenner-Giemsa-stained cultures at days 1, 4, and 7 of differentiation were analyzed. A: myotube density was calculated as the sum of pixels attributed to tones 0–75 (darker tones). There was a significant reduction in the myotube density in the CBA compared with the SUC group B: fusion index was calculated as the fraction of nuclei that incorporated into myotubes over the total nuclei. There was a significant decrease in the fusion index in the CBA group compared with the SUC group. Representative images of Jenner-Giemsa-stained cultures (C) the SUC group (D) the CBA group. *P < 0.05 relative to SUC group. Scale bar = 10 μm. Data are expressed as means ± SE.
Cell Proliferation and Cell Cycle Progression of Myoblasts
Myoblast proliferation and the distribution of cells in different phases of the cell cycle did not statistically differ between cultures of myoblasts isolated from CBA- or SUC-administered macaques during the proliferation phase (Fig. 3, A–C).
Fig. 3.

Indices of proliferation, cell cycle, and cell death in myoblasts. A: there was no difference in cell proliferation between the CBA and SUC groups. B: percent cells in the G1, S, and G2M phases of the cell cycle during proliferation. There was no significant difference in the distribution of cells in the different phases of the cell cycle between the CBA and SUC groups. C: percentage of cells in the G1 and S phase of the cell cycle during differentiation. Cell death was quantified on the basis of the apoptotic and necrotic indices of myoblasts during proliferation and differentiation. There was no significant difference in the apoptotic index (D) and the necrotic index (E) at d3 and d5 of proliferation (F) Representative images of flow cytometric analysis of dual staining for assessing cell death. The lower right quadrant (Annexin V+/PI−) is early-stage apoptotic cells, the upper right quadrant (Annexin V+/PI+) is late-stage apoptotic cells, the upper left quadrant (Annexin V−/PI+) is necrotic cells and the lower left quadrant are the live cells. Data are expressed as means ± SE.
Apoptosis, Necrosis, and Autophagy of Myoblasts
Our results show that decreased myotube formation in myoblast cultures of CBA-administered animals was not due to cell death. During the proliferation phase, the apoptotic index at d3 was 12% in the CBA group compared with 4% in the SUC group. At d5 it was 11% compared with 8% in the SUC group (Fig. 3D). The necrotic index was 1.33 and 0.56% at d3 and 9.2 and 1.6% at d5 of proliferation in the CBA and SUC groups, respectively (Fig. 3E).
To determine the possible contribution of autophagy to CBA-induced loss in muscle mass, the expression of genes associated with autophagy and the protein expression of a critical autophagosomal protein, LC3B, were assessed. There was no significant upregulation of gene expression of beclin-1 and Autophagy related 5 (ATG5) (Fig. 4A) and no significant differences in the labeling index of LC3B protein (Fig. 4B) in myoblast cultures of CBA- and SUC-administered macaques were noted during the proliferation or differentiation phase.
Fig. 4.
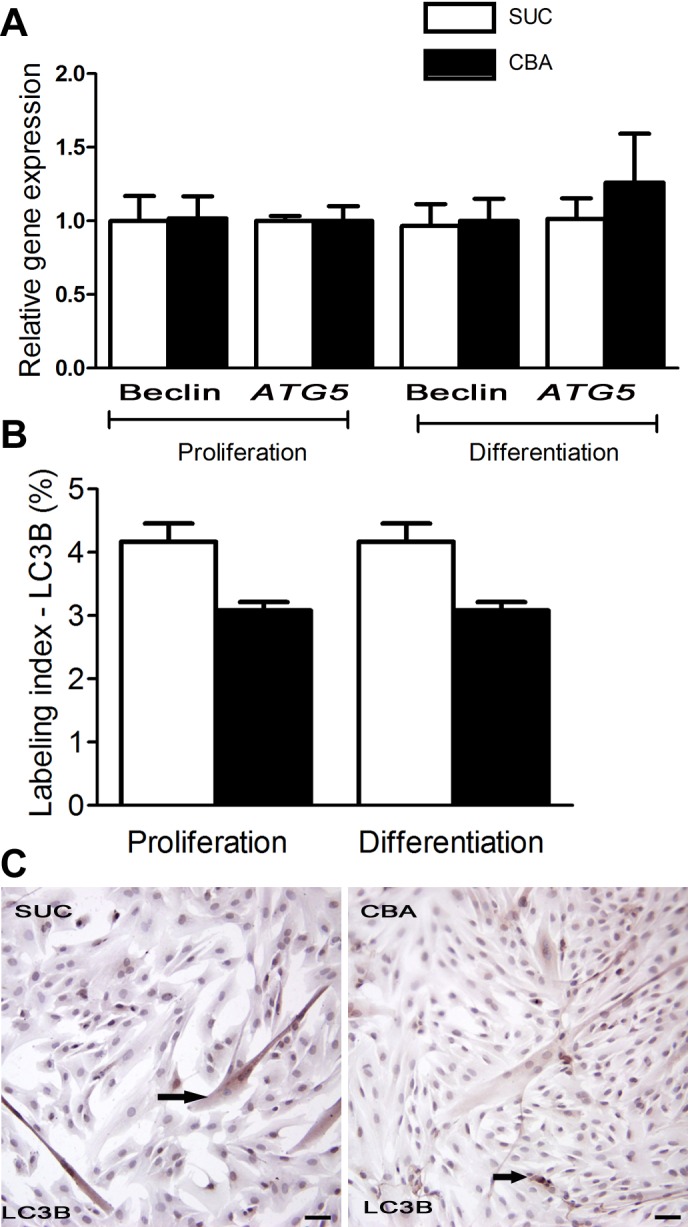
Expression of autophagic markers in myoblasts. A: there was no change in the mRNA expression of two apoptotic genes, Beclin-1 and ATG5 at d5 of proliferation and differentiation between the CBA and SUC groups. B: there was no significant change in the labeling index for LC3B, an autophagosomal protein, during d5 of proliferation and differentiation between the two groups. C: representative images of LC3B immunostaining in myoblasts of SUC and CBA groups. Arrows indicate LC3B immunostaining. Scale bar = 10 μm. Data are expressed as means ± SE.
DISCUSSION
In this study, we determined the effect of CBA on the ability of skeletal muscle myoblasts to proliferate and differentiate into myotubes in vitro. Our results indicate that myoblasts from CBA macaques had significantly reduced expression of genes critical for myogenic differentiation, and this was reflected in a marked decrease in the ability of myoblasts to form myotubes in vitro. These findings support the possibility that the oxidative stress and inflammatory state seen during excessive alcohol consumption could adversely affect the regenerative capacity of SCs, which could mechanistically contribute to the well-known myopathy reported in animal models of chronic alcohol consumption and in humans with alcohol use disorders (20, 27, 32). This decrease occurred despite minimal changes in proliferation and patterns of cell cycling.
Chronic alcohol consumption causes atrophy of type II fibers (22) and necrosis of myofibers in humans (14, 15). The mechanisms identified include direct effects on skeletal muscle leading to increased proteolysis and decreased protein synthesis, as well as secondary effects, including liver disease, malnutrition, or peripheral neuropathy (35, 41, 44). Our previous studies demonstrated that CBA administration to SIV-infected macaques results in significantly accentuated skeletal muscle inflammation preceding overt manifestations of alcoholic myopathy (25). In addition, we showed accentuated skeletal muscle wasting at end-stage SIV infection (24), which we later reported is characterized by marked dysregulation of ubiquitin proteasome components, increased skeletal muscle proteasomal activity, and attenuation of anabolic pathways (21). However, the possibility that the inflammatory/oxidative milieu could impact SC function and contribute to the observed accentuated wasting in CBA-administered SIV-infected macaques had not been previously examined. This study measured myogenic gene expression in myoblasts isolated from CBA and sucrose controls. Our results demonstrate that the mRNA expression of the SC marker, PAX7, and myogenic genes, such as MYOD1, MYOG, and MHC3, were significantly decreased in the CBA group during both the proliferation and differentiation phases of culture. Quiescent SCs express PAX7, which is later reduced as cells undergo cycles of proliferation, and myogenic transcription factors such as MYOD1 (8, 9, 37) and MYF5 (9) are induced. MEF2C is essential for muscle differentiation and establishes skeletal muscle commitment by amplifying and synergizing with MYOD1. The majority of the skeletal muscle genes requires both MYOD1 and MEF2C to activate their transcription (11). Thus, the reduced gene expression of MYOD, MYOG, MYF5 and MEF2C during differentiation in CBA myoblasts indicates impaired myogenic gene expression.
The functional relevance of the decreased myogenic gene expression observed in myoblast cultures from CBA animals was reflected in their impaired ability to form myotubes in vitro. Our results showed a reduction in both the fusion index and myotube density in the myoblast cultures from the CBA animals compared with those of the sucrose group throughout the course of the differentiation period. Garriga et al. (13) reported that in vitro ethanol administration delayed differentiation of primary myoblasts but did not reduce it. These findings are not directly comparable for various reasons. No alcohol was added to our cultures, so that the changes observed reflected the impact of CBA exposure in vivo. In addition, the studies by Garriga et al. (13) assessed myogenic differentiation by measuring the expression of creatine kinase isozymes in the cultures. For our studies, we adapted a staining technique optimized for C2C12 cells (42) that provides visual images of myotube formation and also allows for quantification of the fusion index and myotube density in an unbiased manner. Thus, the observed decreased myogenic differentiation reflects a conserved CBA-induced phenotype of SCs that remains even after removal from the in vivo alcohol environment and multiple cell culture passages.
Recent studies by Arya et al. have demonstrated a significant reduction in the length and breadth of myotubes formed and the fusion index of C2C12 cells when exposed to 100 mM alcohol over a period of 3 days (4). Similarly, Garriga et al. (13) have demonstrated that the differentiation ability of muscle cells is delayed when exposed to alcohol at a concentration of 10 or 100 mM on d5 of in vitro cultures. Our study is the first to demonstrate that in vivo exposure to CBA impairs the myogenic differentiation potential of myoblasts in vitro, even when removed from an alcohol environment. The myoblasts isolated from rhesus macaques exposed to CBA were expanded in vitro and studied during proliferation and differentiation periods in the absence of alcohol. Thus, although alcohol-induced skeletal muscle myopathy is reversible (30) and can be improved even following a short period of abstinence; our results suggest that the impact of CBA on satellite cell phenotype can be long-lasting and could affect its ability to regenerate muscle fibers in response to injury or in conditions associated with muscle wasting such as HIV disease.
Alcohol has been shown to increase stem cell death (16, 26). However, our results do not support a significant role for cell death in the impaired ability of myoblasts isolated from CBA-administered macaques to form myotubes. Autophagy and cell death regulate each other, and autophagy is considered to be a prosurvival mechanism used by cells to remove either degraded proteins or damaged organelles (12, 19). Our findings also indicate that there was no change in expression of key autophagic proteins in cultures of myoblasts obtained from CBA macaques. Hence, our results do not provide evidence for a significant role for autophagy or cell death in the CBA-associated decreased myoblast differentiation potential. Moreover, our results do not provide evidence that alcohol affects myoblast cell cycle progression. Reports on the effects of alcohol on cell cycle progression are inconclusive. McClain et al. (23) demonstrated that binge alcohol exposure reduced the proportion of neural progenitors in the hippocampus in S-phase but had no effect on G1 and G2/M phase. Anthony et al. (3) established that alcohol induces neural stem cells into premature S-phase entry, disjointed DNA synthesis, and increased apoptosis. In contrast, Ahluwalia et al. (1) demonstrated that alcohol significantly reduced proliferation in lymphocytes by inhibiting the G2-M phase and not affecting the G1-S phase. We did not observe significant differences in myoblast proliferation in cultures from CBA compared with those of SUC animals. Again, it is important to note that no alcohol was added to the cultures and that could explain the differences in observations with those of other groups. It is possible that proliferation of CBA SCs may be impaired in response to a challenge, such as exposure to proinflammatory factors, toxins, or in other types of injury. This warrants further study.
Perspectives and Significance
The results from this study suggest that CBA consumption by rhesus macaques markedly impairs skeletal muscle satellite cell regenerative capacity. Using an in vitro approach, we show significant downregulation of genes that are essential for induction of differentiation, and this was reflected in a marked decrease in myotube formation and fusion in vitro. These results are of great importance, as they indicate a certain memory of the myoblasts isolated from CBA-administered animals, detected in vitro during the proliferation and differentiation phases, despite the absence of alcohol in the cultures. One may speculate that these changes in gene expression may be the result of epigenetic modifications resulting from chronic alcohol consumption. Ongoing studies will examine the alterations in myoblast microRNA profiles and methylation patterns to elucidate the mechanisms responsible for persistent changes in these myoblasts. In addition, future studies will investigate myotube functional integrity, in particular, their metabolic response to anabolic stimulation. Our working hypothesis is that the CBA-associated proinflammatory/oxidative skeletal muscle environment results in dysregulation of satellite cell function, decreasing their ability to participate in the regeneration of myofibers. We predict this is an additional unexplored mechanism that may contribute to alcoholic myopathy and warrants further study.
GRANTS
This work was supported by the National Institute on Alcohol Abuse and Alcoholism, National Institutes of Health (Grants AA07577, AA09803, and AA11290).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
Author contributions: L.S., N.L., J.D., S.N., G.J.B., W.C., and P.E.M. conception and design of research; L.S., N.L., P.B., C.V.S., J.C.M., T.D.A., S.D.P., and J.D. performed experiments; L.S., N.L., P.B., C.V.S., J.C.M., and T.D.A. analyzed data; L.S., G.J.B., and P.E.M. interpreted results of experiments; L.S. prepared figures; L.S. and P.E.M. drafted manuscript; L.S., N.L., P.B., C.V.S., J.C.M., J.D., S.N., G.J.B., W.C., and P.E.M. edited and revised manuscript; L.S., N.L., P.B., C.V.S., J.C.M., T.D.A., S.D.P., J.D., S.N., G.J.B., W.C., and P.E.M. approved final version of manuscript.
ACKNOWLEDGMENTS
The authors wish to acknowledge the technical expertise and support received from the LSUHSC Comprehensive Alcohol Research Center Analytical Core, especially Constance Porretta; and the scientific discussions and contributions of Drs. Robert Siggins and Andrew Hollenbach.
REFERENCES
- 1.Ahluwalia BS, Westney LS, Rajguru SU. Alcohol inhibits cell mitosis in G2-M phase in cell cycle in a human lymphocytes in vitro study. Alcohol 12: 589–592, 1995 [DOI] [PubMed] [Google Scholar]
- 2.Anderson ML, Nokia MS, Govindaraju KP, Shors TJ. Moderate drinking? Alcohol consumption significantly decreases neurogenesis in the adult hippocampus. Neuroscience 224: 202–209, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Anthony B, Zhou FC, Ogawa T, Goodlett CR, Ruiz J. Alcohol exposure alters cell cycle and apoptotic events during early neurulation. Alcohol Alcohol 43: 261–273, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Arya MA, Tai AK, Wooten EC, Parkin CD, Kudryavtseva E, Huggins GS. Notch Pathway activation contributes to inhibition of C2C12 myoblast differentiation by ethanol. PLos One 8: e71632, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bagby GJ, Stoltz DA, Zhang P, Kolls JK, Brown J, Bohm RP, Jr, Rockar R, Purcell J, Murphey-Corb M, Nelson S. The effect of chronic binge ethanol consumption on the primary stage of SIV infection in rhesus macaques. Alcoholism Clin Exp Res 27: 495–502, 2003 [DOI] [PubMed] [Google Scholar]
- 6.Benchaouir R, Meregalli M, Farini A, D'Antona G, Belicchi M, Goyenvalle A, Battistelli M, Bresolin N, Bottinelli R, Garcia L, Torrente Y. Restoration of human dystrophin following transplantation of exon-skipping-engineered DMD patient stem cells into dystrophic mice. Cell Stem Cell 1: 646–657, 2007 [DOI] [PubMed] [Google Scholar]
- 7.Conboy MJ, Cerletti M, Wagers AJ, Conboy IM. Immuno-analysis and FACS sorting of adult muscle fiber-associated stem/precursor cells. Methods Mol Biol 621: 165–173, 2010 [DOI] [PubMed] [Google Scholar]
- 8.Cornelison DD, Olwin BB, Rudnicki MA, Wold BJ. MyoD(-/-) satellite cells in single-fiber culture are differentiation defective and MRF4 deficient. Dev Biol 224: 122–137, 2000 [DOI] [PubMed] [Google Scholar]
- 9.Cornelison DD, Wold BJ. Single-cell analysis of regulatory gene expression in quiescent and activated mouse skeletal muscle satellite cells. Dev Biol 191: 270–283, 1997 [DOI] [PubMed] [Google Scholar]
- 10.Cui Q, Wang Y, Saleh KJ, Wang GJ, Balian G. Alcohol-induced adipogenesis in a cloned bone-marrow stem cell. J Bone Joint Surg Am 88 Suppl 3: 148–154, 2006 [DOI] [PubMed] [Google Scholar]
- 11.Dodou E, Xu SM, Black BL. mef2c is activated directly by myogenic basic helix-loop-helix proteins during skeletal muscle development in vivo. Mech Dev 120: 1021–1032, 2003 [DOI] [PubMed] [Google Scholar]
- 12.Dolganiuc A, Thomes PG, Ding WX, Lemasters JJ, Donohue TM., Jr Autophagy in alcohol-induced liver diseases. Alcoholism Clin Exp Res 36: 1301–1308, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Garriga J, Adanero E, Fernandez-Sola J, Urbano-Marquez A, Cusso R. Ethanol inhibits skeletal muscle cell proliferation and delays its differentiation in cell culture. Alcohol Alcohol 35: 236–241, 2000 [DOI] [PubMed] [Google Scholar]
- 14.Haller RG. Experimental acute alcoholic myopathy—a histochemical study. Muscle Nerve 8: 195–203, 1985 [DOI] [PubMed] [Google Scholar]
- 15.Haller RG, Drachman DB. Alcoholic rhabdomyolysis: an experimental model in the rat. Science 208: 412–415, 1980 [DOI] [PubMed] [Google Scholar]
- 16.Hicks SD, Miller MW. Effects of ethanol on transforming growth factor β1-dependent and -independent mechanisms of neural stem cell apoptosis. Exp Neurol 229: 372–380, 2011 [DOI] [PubMed] [Google Scholar]
- 17.Kao GW, Lamb EK, Kao RL. Skeletal muscle stem cells. Methods Mol Biol 1036: 19–32, 2013 [DOI] [PubMed] [Google Scholar]
- 18.Kheterpal I, Coleman L, Ku G, Wang ZQ, Ribnicky D, Cefalu WT. Regulation of insulin action by an extract of Artemisia dracunculus L. in primary human skeletal muscle culture: a proteomics approach. Phytother Res 24: 1278–1284, 2010 [DOI] [PubMed] [Google Scholar]
- 19.Kim I, Lemasters JJ. Mitophagy selectively degrades individual damaged mitochondria after photoirradiation. Antioxid Redox Signal 14: 1919–1928, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Lang CH, Frost RA, Summer AD, Vary TC. Molecular mechanisms responsible for alcohol-induced myopathy in skeletal muscle and heart. Int J Biochem Cell Biol 37: 2180–2195, 2005 [DOI] [PubMed] [Google Scholar]
- 21.LeCapitaine NJ, Wang ZQ, Dufour JP, Potter BJ, Bagby GJ, Nelson S, Cefalu WT, Molina PE. Disrupted anabolic and catabolic processes may contribute to alcohol-accentuated SAIDS-associated wasting. J Infect Dis 204: 1246–1255, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Martin FC, Peters TJ. Assessment in vitro and in vivo of muscle degradation in chronic skeletal muscle myopathy of alcoholism. Clin Sci (Lond) 68: 693–700, 1985 [DOI] [PubMed] [Google Scholar]
- 23.McClain JA, Hayes DM, Morris SA, Nixon K. Adolescent binge alcohol exposure alters hippocampal progenitor cell proliferation in rats: effects on cell cycle kinetics. J Comp Neurol 519: 2697–2710, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Molina PE, Lang CH, McNurlan M, Bagby GJ, Nelson S. Chronic alcohol accentuates simian acquired immunodeficiency syndrome-associated wasting. Alcoholism Clin Exp Res 32: 138–147, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Molina PE, McNurlan M, Rathmacher J, Lang CH, Zambell KL, Purcell J, Bohm RP, Zhang P, Bagby GJ, Nelson S. Chronic alcohol accentuates nutritional, metabolic, and immune alterations during asymptomatic simian immunodeficiency virus infection. Alcoholism Clin Exp Res 30: 2065–2078, 2006 [DOI] [PubMed] [Google Scholar]
- 26.Morris SA, Eaves DW, Smith AR, Nixon K. Alcohol inhibition of neurogenesis: a mechanism of hippocampal neurodegeneration in an adolescent alcohol abuse model. Hippocampus 20: 596–607, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Nguyen VA, Le T, Tong M, Silbermann E, Gundogan F, de la Monte SM. Impaired insulin/IGF signaling in experimental alcohol-related myopathy. Nutrients 4: 1058–1075, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Olguin HC, Pisconti A. Marking the tempo for myogenesis: Pax7 and the regulation of muscle stem cell fate decisions. J Cell Mol Med 16: 1013–1025, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pacy PJ, Preedy VR, Peters TJ, Read M, Halliday D. The effect of chronic alcohol ingestion on whole body and muscle protein synthesis—a stable isotope study. Alcohol Alcohol 26: 505–513, 1991 [DOI] [PubMed] [Google Scholar]
- 30.Peters TJ, Martin F, Ward K. Chronic alcoholic skeletal myopathy—common and reversible. Alcohol 2: 485–489, 1985 [DOI] [PubMed] [Google Scholar]
- 31.Ponnappa BC, Rubin E. Modeling alcohol's effects on organs in animal models. Alcohol Res Health 24: 93–104, 2000 [PMC free article] [PubMed] [Google Scholar]
- 32.Preedy VR, Adachi J, Ueno Y, Ahmed S, Mantle D, Mullatti N, Rajendram R, Peters TJ. Alcoholic skeletal muscle myopathy: definitions, features, contribution of neuropathy, impact and diagnosis. Eur J Neurol 8: 677–687, 2001 [DOI] [PubMed] [Google Scholar]
- 33.Preedy VR, Crabb DW, Farres J, Emery PW. Alcoholic myopathy and acetaldehyde. Novartis Found Symp 285: 158–177; discussion 177–182, 198–159, 2007 [DOI] [PubMed] [Google Scholar]
- 34.Reilly ME, Mantle D, Richardson PJ, Salisbury J, Jones J, Peters TJ, Preedy VR. Studies on the time-course of ethanol's acute effects on skeletal muscle protein synthesis: comparison with acute changes in proteolytic activity. Alcoholism Clin Exp Res 21: 792–798, 1997 [PubMed] [Google Scholar]
- 35.Romero JC, Santolaria F, Gonzalez-Reimers E, Diaz-Flores L, Conde A, Rodriguez-Moreno F, Batista N. Chronic alcoholic myopathy and nutritional status. Alcohol 11: 549–555, 1994 [DOI] [PubMed] [Google Scholar]
- 36.Shah R, Sinanan AC, Knowles JC, Hunt NP, Lewis MP. Craniofacial muscle engineering using a 3-dimensional phosphate glass fibre construct. Biomaterials 26: 1497–1505, 2005 [DOI] [PubMed] [Google Scholar]
- 37.Smith CK, 2nd, Janney MJ, Allen RE. Temporal expression of myogenic regulatory genes during activation, proliferation, and differentiation of rat skeletal muscle satellite cells. J Cell Physiol 159: 379–385, 1994 [DOI] [PubMed] [Google Scholar]
- 38.Smith KJ, Butler TR, Prendergast MA. Ethanol impairs microtubule formation via interactions at a microtubule associated protein-sensitive site. Alcohol 47: 539–543, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Snow MH. Myogenic cell formation in regenerating rat skeletal muscle injured by mincing. II. An autoradiographic study. Anat Rec 188: 201–217, 1977 [DOI] [PubMed] [Google Scholar]
- 40.Teschner M, Schaefer RM, Weissinger F, Kulzer P, Duelk MJ, Peter G, Heidland A. Chronic ethanol ingestion enhances catabolism and muscle protease activity in acutely uremic rats. Nephron 50: 338–344, 1988 [DOI] [PubMed] [Google Scholar]
- 41.Thuluvath PJ, Triger DR. Autonomic neuropathy and chronic liver disease. Q J Med 72: 737–747, 1989 [PubMed] [Google Scholar]
- 42.Velica P, Bunce CM. A quick, simple and unbiased method to quantify C2C12 myogenic differentiation. Muscle Nerve 44: 366–370, 2011 [DOI] [PubMed] [Google Scholar]
- 43.Wang ZQ, Ribnicky D, Zhang XH, Zuberi A, Raskin I, Yu Y, Cefalu WT. An extract of Artemisia dracunculus L. enhances insulin receptor signaling and modulates gene expression in skeletal muscle in KK-A(y) mice. J Nutr Biochem 22: 71–78, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wijnia JW, Wielders JP, Lips P, van de Wiel A, Mulder CL, Nieuwenhuis KG. Is vitamin D deficiency a confounder in alcoholic skeletal muscle myopathy? Alcoholism Clin Exp Research 37 Suppl 1: E209–215, 2013 [DOI] [PubMed] [Google Scholar]
- 45.Yin H, Price F, Rudnicki MA. Satellite cells and the muscle stem cell niche. Physiol Rev 93: 23–67, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]