

Abstract Abstract
In sixteen moss, lichen and mixed (moss/lichen) samples, collected from Estonia, Latvia and Lithuania, 291 specimens, 48 simplexes, including one exuvium with 6 eggs, and 8 free-laid eggs of eutardigrades were found. In total, 17 species, together with one new to science, were identified (all are new records for the Baltic states): Astatumen bartosi, Diphascon (Adropion) prorsirostre, D. (Diphascon) bullatum, D. (D.) pingue pingue, D. (D.) recamieri, D. (D.) rugosum, Hypsibius convergens, H. dujardini, H. cf. scabropygus, Isohypsibius ronsisvallei, I. sattleri, Macrobiotus harmsworthi harmsworthi, M. hufelandi hufelandi, Milnesium asiaticum, Milnesium tardigradum tardigradum, Minibiotus formosus sp. n. and Paramacrobiotus richtersi. The new species is most similar to Minibiotus gumersindoi, but differs from it mainly by the presence of two types of cuticular pores, the absence of a triangular or pentagonal arrangement of pores above a single large pore on legs, the presence of granulation on all legs and a different macroplacoid length sequence. In this paper we also provide photographs and morphometrics of H. cf. scabropygus.
Keywords: Estonia, Europe, Hypsibius cf. scabropygus, Latvia, Lithuania, new species, Tardigrada
Introduction
The Baltic States, i.e. Estonia, Latvia and Lithuania, are located on the eastern coast of the Baltic Sea, and fall within the Palearctic ecozone (Holt et al. 2012). The topography of the three countries is dominated by lowlands with the highest peaks at ca. 300 m asl. The temperate climate is intermediate between maritime and continental. Even though the phylum Tardigrada is cosmopolitan and currently comprises ca. 1,200 species (Degma et al. 2013), so far only six tardigrade taxa have been reported from the Baltic States. Specifically, two from Estonia: Eremobiotus alicatai (Binda, 1969) and Isohypsibius cf. marcellinoi (Binda & Pilato, 1971), two from Latvia: Paramacrobiotus richtersi group and Macrobiotus hufelandi group and two from Lithuania: Macrobiotus sp. and Ramazzottius sp. (Šatkauskienė and Vosyliūtė 2010, Zawierucha and Kaźmierski 2012, Ziemelis et al. 2012).
In this study we report seventeen tardigrade species, which are all new records for the Baltic States. Moreover, one of these species is also new to science. The new species belongs to the genus Minibiotus R.O. Schuster, 1980, that until 1988 contained only a single species, Minibiotus intermedius (Plate, 1888). In 1988 Pilato and Claxton (1988) described Minibiotus maculartus, and within the last decade fourteen new Minibiotus species have been described. Several species have also been transferred to Minibiotus from the genus Macrobiotus based on characters defined by Claxton (1998) and later supplemented by Guidetti et al. (2007) (Michalczyk and Kaczmarek 2003a, Pilato et al. 2003, Michalczyk and Kaczmarek 2004, Guil and Guidetti 2005, Michalczyk et al. 2005, Pilato and Lisi 2006b, Li et al. 2008, Fontoura et al. 2009a, b, Rossi et al. 2009, Meyer and Hinton 2009, Meyer and Domingue 2011, Meyer et al. 2011 Meyer 2012). Currently, the total number of Minibiotus species amounts to as many as forty seven.
In addition to the description of the new species, we also provide morphometric data and photographs of Hypsibius cf. scabropygus, a rare species that belongs to a large group of hypsibiids with at least partially sculptured dorsal cuticle and pharynx with two macroplacoids and without the microplacoid.
Material and methods
Sixteen moss, lichen and mixed (moss/lichen) samples from trees, soil and stones were collected from 15 localities in Estonia, Latvia and Lithuania between the 29 April and the 5 May 2012 by the third author (more details below). Samples were collected and examined for tardigrades using standard methods (see Dastych 1980). After extraction, animals were mounted on microscope slides in Hoyer’s medium. All specimens were examined measured and photographed using Phase Contrast Microscopy (PCM) or Scanning Electron Microscopy (SEM). In total 358 specimens (including 47 simplexes), one exuvium with 6 eggs, and 8 free-laid eggs were examined.
All measurements are given in micrometers [μm]. Structures were measured only if their orientation was appropriate. Body length was measured from the anterior extremity to the end of the body, excluding the hind legs. Buccal tube length and the level of the stylet support insertion point were measured according to Pilato (1981). Buccal tube width was measured as the external diameter at the level of the stylet support insertion point. Lengths of the claw branches were measured from the base of the claw to the top of the branch including accessory points for Minibiotus and according to Beasley et al. (2008) for Hypsibius. The pt ratio is the ratio of the length of a given structure to the length of the buccal tube expressed as a percentage (Pilato 1981). Macroplacoid length sequence is given according to Kaczmarek et al. (2014b), i.e. macroplacoids are listed from the shortest to the longest and their relative sizes are denoted with appropriate inequality, approximation and/or equality signs (<, ≤, ≈, =). Morphometric data were handled using the ‘Macrobiotoidea’ ver. 1.1 template available from the Tardigrada Register (www.tardigrada.net/register, Michalczyk and Kaczmarek 2013).
For species identification and differentiation, keys in Claxton (1998), Fontoura and Pilato (2007), Kaczmarek et al. (2011), Michalczyk et al. 2012a, b and Ramazzotti and Maucci (1983), and original descriptions and redescriptions (Ehrenberg 1859, Ramazzotti 1959, 1962, Horning et al. 1978, Bertolani and Rebecchi 1993, Dastych 1988, 1990, Binda and Pilato 1992, Michalczyk and Kaczmarek 2004, Michalczyk et al. 2005, Miller et al. 2005, Fontoura et al. 2009a, b, Meyer and Hinton 2009, Meyer et al. 2011) as well as for insertion of the stylet muscles Pilato (2013) were used. Tardigrade taxonomy is presented according to Marley et al. (2011). Only specimens determined to species level are provided in the list of species (we omitted all specimens determined only to the species group level, e.g. the hufelandi group or the oberhaeuseri group). In the species list Roman numbers indicate sample codes (see sampling localities) and Arabic numbers indicate the number of specimens, exuvia/simplexes and eggs.
Raw data underlying the description of Minibiotus formosus sp. n. are deposited in the Tardigrada Register (Michalczyk and Kaczmarek 2013) under www.tardigrada.net/register/0012.htm.
Sampling localities
56°03'08"N, 24°24'10"E, ca. 33 m asl: Lithuania, Panevėžys county, Pasvalys district municipality, along the road E67, 0.5 km before the turning to Pasvalys, moss from tree and soil (slide code: LT 2422), date: 29.04.2012.
55°25'59"N, 24°13'32"E, ca. 59 m asl: Lithuania, Kaunas county, Kėdainiai district municipality, Truskava city, near a church, lichens from tree and moss from soil (slide code: LT 2423), date: 29.04.2012.
55°17'12"N, 23°58'57"E, ca. 30 m asl: Lithuania, Kaunas county, Kėdainiai district municipality, Kėdainiai city, Kranto II street; moss from wall (slide code: LT 2424), date: 29.04.2012.
55°17'13"N, 23°58'56"E, ca. 30 m asl: Lithuania, Kaunas county, Kėdainiai district municipality, Kėdainiai city, Paeismilgio street; moss from stone (slide code: LT 2425), date: 29.04.2012.
55°43'35"N, 24°21'30"E, ca. 62 m asl: Lithuania, Panevėžys county, Panevėžys district municipality, Panevėžys city, Garden Street near Holy Trinity Rector; moss from tree (slide code: LT 2440), date: 05.05.2012.
56°38'53"N, 23°43'18"E, ca. 7 m asl: Latvia, Zemgale region, Jelgava municipality, Jelgava city, City Park; moss from soil (slide code: ŁO 2426), date: 29.04.2012.
57°10'33"N, 24°50'32"E, ca. 45 m asl: Latvia, Vidzeme region, Sigulda municipality, Gutmana Cave in the Gauja National Park; moss from rocks (slide code: ŁO 2427), date: 30.04.2012.
56°23'55"N, 24°07'33"E, ca. 25 m asl: Latvia, Zemgale region, Bauska municipality, along Road No P103, 0.5 km from Saulaine; lichens from tree (slide code: ŁO 2428) date: 29.04.2012.
57°09'55"N, 24°51'03"E, ca. 73 m asl: Latvia, Vidzeme region, Sigulda municipality, Turaida city, Turaida Castle; moss from stone (slide code: ŁO 2430), date: 30.04.2012.
56°54'32"N, 24°08'45"E, ca. 10 m asl: Latvia, Riga Region, boundary of Ķekava municipality, along road no A2; moss from tree (slide code: ŁO 2431), date: 30.04.2012.
57°09'59"N, 24°50'59"E, ca. 91 m asl: Latvia, Vidzeme region, Sigulda municipality, Sigulda city, Sigulda Castle; moss from stone (slide code: ŁO 2432), date: 30.04.2012.
56°41'22"N, 23°47'43"E, ca. 4 m asl: Latvia, Zemgale region, Ozolnieki municipality, Ozolnieki city, about 100 m from the Ozolnieki Lake; moss from soil (slide code: ŁO 2433), date: 29.04.2012.
57°45'43"N, 24°20'59"E, ca. 3 m asl: Latvia, Vidzeme region, Salacgriva municipality, Salacgriva city; moss from soil, near the beach (slide code: ŁO 2434), date: 01.05.2012.
59°10'44"N, 24°30'06"E, ca. 59 m asl: Republic of Estonia, Harju county, Kernu Parish municipality, Road No 4, moss from tree (slide code: ES 2420), date: 04.05.2012.
59°10'44"N, 24°30'06"E, ca. 59 m asl: Republic of Estonia, Harju county, Kernu Parish municipality, Road No 4, moss from tree (slide code: ES 2421), date: 04.05.2012.
58°48'47"N, 24°24'46"E, ca. 32 m asl: Republic of Estonia, Rapla County, Märjamaa municipality, forest near Konuvere village, moss from tree (slide codes: ES 2487), date: 29.04.2012.
Results
Taxonomic accounts of species found in the study
Phylum: Tardigrada (Spallanzani, 1777)
Class: Eutardigrada Richters, 1926
Order: Apochela Schuster, Nelson, Grigarick and Christenberry, 1980
Family: Milnesiidae Ramazzotti, 1962
Genus: Milnesium Doyère, 1840
Milnesium asiaticum
Tumanov, 2006
Localities and specimen numbers.
XV: 1 specimen.
Remarks.
Our specimen corresponds perfectly to the original description. Milnesium asiaticum was originally described from Kirghizstan and subsequently found in the Svalbard archipelago (Tumanov 2006, Kaczmarek et al. 2012).
Milnesium tardigradum tardigradum
Doyère, 1840
Localities and specimen numbers.
VIII: 31 specimens (including 6 simplexes) + 1 exuvium with 6 eggs.
Remarks.
Specimens correspond perfectly with the redescription by Michalczyk et al. (2012a, b). This species was reported from many localities throughout the World, however records prior to Michalczyk et al. (2012a, b) need to be verified. So far, all confirmed localities are exclusively European (Michalczyk et al. 2012a, b).
Order: Parachela Schuster, Nelson, Grigarick & Christensen, 1980
Superfamily: Hypsibioidea Pilato, 1969 (in Marley et al. 2011)
Family: Hypsibiidae Pilato, 1969
Subfamily: Diphasconinae Dastych, 1992
Genus: Diphascon Plate, 1888
Subgenus: Diphascon (Diphascon) Pilato, 1987
Diphascon (Diphascon) bullatum
Murray, 1905
Localities and specimen numbers.
XIII: 1 specimen.
Remarks.
Specimens correspond well with the limited original description (Murray 1905) and also with later descriptions (Argue 1974 and Dastych 1980, 1988). This species is very similar to Diphascon (Diphascon) patanei (Binda & Pilato, 1971) and, as suggested by Dastych (1988), these two species could be synonymous. Thus, to clarify the taxonomic status of Diphascon (Diphascon) patanei, a re-description of Diphascon (Diphascon) bullatum is necessary based on material from locus typicus in Scotland (the type material probably does not exist) (Dastych 1988).
Diphascon (Diphascon) pingue pingue
(Marcus, 1936)
Localities and specimen numbers.
XV: 11 specimens.
Remarks.
Although we have found only 11 specimens, we were confident in identifying them to Diphascon (Diphascon) pingue because they corresponded perfectly to the partial redescriptions by Pilato and Binda (1997/1998, 1999) and we also identified them with the key by Fontoura and Pilato (2007). The species belongs to the pingue group and has been previously recorded from numerous localities throughout the World, however the majority of records should be verified based on the modern taxonomy (Pilato and Binda 1997/1998, 1999). Currently, exclusively verified localities of this species are only from Europe and North America (Pilato and Binda 1997/1998).
Diphascon (Diphascon) recamieri
Richters, 1911
Localities and specimen numbers.
XV: 1 specimen.
Remarks.
The species has previously been found in many localities, mostly in the Holarctic (McInnes 1994).
Diphascon (Diphascon) rugosum
(Bartoš, 1935)
Localities and specimen numbers.
II: 3 specimens.
Remarks.
The species has previously been found in many localities in the Holarctic (McInnes 1994).
Genus: Diphascon Pilato, 1987
Subgenus. Diphascon (Adropion) prorsirostre
Thulin, 1928
Localities and specimen numbers.
XIV: 2 specimens, XV: 2 specimens.
Remarks.
The species has been previously found in many localities, mostly in the Holarctic (McInnes 1994).
Subfamily: Hypsibiinae Pilato, 1969
Genus: Hypsibius Ehrenberg, 1848
Hypsibius convergens
(Urbanowicz, 1925)
Localities and specimen numbers.
IX: 6 specimens (including 2 simplexes).
Remarks.
Belonging to the cosmopolitan convergens-dujardini complex of species (McInnes 1994, Miller et al. 2005, Kaczmarek and Michalczyk 2009a, Kaczmarek et al. 2014a), Hypsibius convergens used to be considered cosmopolitan, but it is most likely a complex of very similar (possibly also cryptic) species found throughout the world. The original Hypsibius convergens description no longer conforms to modern standards and therefore a redescription is required. Nevertheless, the examined specimens correspond perfectly with the original description and the Hypsibius convergens characteristics reviewed by Miller et al. (2005).
Hypsibius dujardini
(Doyère, 1840)
Localities and specimen numbers.
XIV: 2 specimens (including 1 simplex), XV: 5 specimens.
Remarks.
Hypsibius dujardini belongs to the cosmopolitan convergens-dujardini complex of species (McInnes 1994, Miller et al. 2005, Kaczmarek and Michalczyk 2009a, Kaczmarek et al. 2014a), and used to be considered cosmopolitan, but it is most likely a complex of similar (possibly cryptic) species found throughout the world. Due to the limited original description, Hypsibius dujardini needs a modern redescription. Nevertheless, the examined specimens correspond perfectly with the original description and the Hypsibius dujardini characteristics reviewed by Miller et al. (2005).
Hypsibius cf. scabropygus
Cuénot, 1929
Table 1.
Measurements and pt values of selected morphological structures of Hypsibius cf. scabropygus Cuénot, 1929 mounted in Hoyer’s medium (N – number of specimens/structures measured, RANGE refers to the smallest and the largest structure among all measured specimens; SD – standard deviation).
| CHARACTER | N | RANGE | MEAN | SD | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | ||||||
| Body length | 14 | 183 | – | 293 | 808 | – | 1132 | 235 | 949 | 33 | 97 |
| Buccopharyngeal tube | |||||||||||
| Buccal tube length | 15 | 22.0 | – | 28.2 | – | 24.6 | – | 2.0 | – | ||
| Stylet support insertion point | 15 | 12.0 | – | 15.9 | 51.3 | – | 57.1 | 13.5 | 54.9 | 1.1 | 1.6 |
| Buccal tube external width | 15 | 1.5 | – | 2.0 | 6.5 | – | 8.0 | 1.8 | 7.2 | 0.2 | 0.5 |
| Buccal tube internal width | 15 | 0.6 | – | 0.9 | 2.1 | – | 3.4 | 0.7 | 2.7 | 0.1 | 0.4 |
| Placoid lengths | |||||||||||
| Macroplacoid 1 | 15 | 1.8 | – | 3.2 | 7.7 | – | 12.3 | 2.4 | 9.6 | 0.4 | 1.3 |
| Macroplacoid 2 | 15 | 1.7 | – | 2.7 | 7.2 | – | 11.3 | 2.2 | 8.9 | 0.3 | 1.1 |
| Macroplacoid row | 15 | 4.7 | – | 6.3 | 19.1 | – | 26.5 | 5.3 | 21.7 | 0.5 | 1.8 |
| Claw 1 lengths | |||||||||||
| External base | 14 | 2.2 | – | 4.5 | 9.1 | – | 17.2 | 3.5 | 14.1 | 0.7 | 2.1 |
| External primary branch | 14 | 4.0 | – | 8.8 | 18.2 | – | 36.1 | 6.9 | 28.1 | 1.5 | 5.0 |
| External secondary branch | 14 | 2.3 | – | 5.9 | 10.5 | – | 24.4 | 4.4 | 17.9 | 1.0 | 3.9 |
| Internal base | 12 | 2.1 | – | 4.1 | 9.3 | – | 15.5 | 3.3 | 13.4 | 0.6 | 1.8 |
| Internal primary branch | 12 | 3.8 | – | 5.9 | 16.0 | – | 23.5 | 4.8 | 19.3 | 0.6 | 2.0 |
| Internal secondary branch | 12 | 2.4 | – | 4.3 | 9.9 | – | 16.5 | 3.3 | 13.3 | 0.6 | 2.0 |
| Claw 2 lengths | |||||||||||
| External base | 11 | 3.0 | – | 5.2 | 12.4 | – | 19.9 | 4.1 | 16.7 | 0.7 | 2.2 |
| External primary branch | 13 | 6.7 | – | 10.4 | 29.7 | – | 43.7 | 8.5 | 34.7 | 1.3 | 4.4 |
| External secondary branch | 13 | 4.3 | – | 6.7 | 19.0 | – | 27.2 | 5.4 | 21.9 | 0.7 | 2.6 |
| Internal base | 10 | 2.4 | – | 4.5 | 10.9 | – | 18.9 | 3.6 | 14.6 | 0.7 | 2.2 |
| Internal primary branch | 12 | 4.0 | – | 6.7 | 17.7 | – | 27.2 | 5.4 | 22.0 | 0.9 | 2.9 |
| Internal secondary branch | 12 | 2.6 | – | 5.4 | 11.8 | – | 22.0 | 4.1 | 16.7 | 0.9 | 3.0 |
| Claw 3 lengths | |||||||||||
| External base | 9 | 2.7 | – | 6.2 | 11.9 | – | 23.8 | 4.3 | 17.3 | 1.0 | 3.5 |
| External primary branch | 9 | 7.2 | – | 10.4 | 29.3 | – | 43.7 | 8.8 | 35.7 | 1.1 | 4.4 |
| External secondary branch | 9 | 3.6 | – | 6.5 | 12.8 | – | 27.3 | 5.2 | 21.0 | 1.0 | 4.3 |
| Internal base | 11 | 2.3 | – | 4.1 | 10.5 | – | 17.2 | 3.4 | 13.9 | 0.6 | 1.9 |
| Internal primary branch | 13 | 3.8 | – | 6.5 | 17.3 | – | 27.3 | 5.4 | 21.8 | 0.9 | 3.1 |
| Internal secondary branch | 12 | 2.7 | – | 6.1 | 12.2 | – | 24.8 | 3.9 | 16.0 | 0.9 | 3.5 |
| Claw 4 lengths | |||||||||||
| Anterior base | 13 | 3.3 | – | 5.6 | 12.8 | – | 20.1 | 4.1 | 16.8 | 0.6 | 2.2 |
| Anterior primary branch | 13 | 4.4 | – | 7.5 | 19.5 | – | 31.1 | 5.9 | 24.2 | 1.1 | 3.9 |
| Anterior secondary branch | 11 | 3.1 | – | 13.2 | 13.0 | – | 47.3 | 4.8 | 18.9 | 2.9 | 9.7 |
| Posterior base | 12 | 2.7 | – | 5.4 | 12.3 | – | 21.5 | 4.5 | 18.2 | 0.9 | 3.1 |
| Posterior primary branch | 12 | 4.9 | – | 14.9 | 22.0 | – | 60.6 | 10.3 | 41.9 | 2.9 | 11.2 |
| Posterior secondary branch | 12 | 4.0 | – | 6.5 | 15.4 | – | 25.6 | 5.2 | 21.2 | 0.9 | 3.4 |
Figures 1–5.

Hypsibius cf. scabropygus Cuénot, 1929: 1 habitus (dorso-lateral view) 2–4 caudo-dorsal cuticle with distinct sculpturing – tubercles and tubercles merged into platelets 5 a single caudo-dorsal platelet. (1–3: PCM, 4–5: SEM).
Figures 6–7.
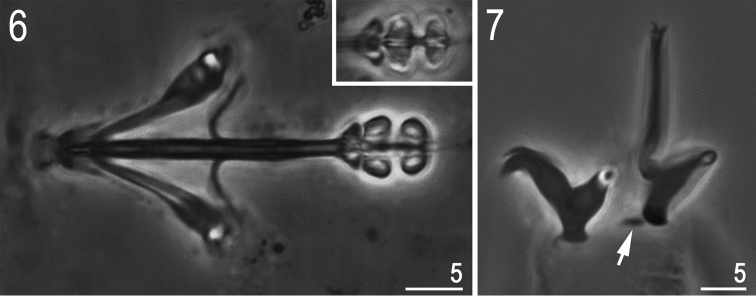
Hypsibius cf. scabropygus Cuénot, 1929: 6 bucco-pharyngeal apparatus (dorso-ventral projection, ventral placoids in the insert) 7 claws IV (arrow indicates a small cuticular bar near the posterior claw). (Both PCM).
Localities and specimen numbers.
XI: 1 simplex, XII: 1 specimen, XIV: 34 specimens (including 4 simplexes), XV: 24 specimens (including 6 simplexes).
Description
(measurements in Table 1). Adults. Body transparent/white (after preparation), eyes present in 6 of 15 (40%) specimens mounted in Hoyer’s medium (Fig. 1). Dorsal cuticle sculptured: from head to legs II without tubercles but clearly thickened, from legs II to the caudal end of the body (including legs IV) with irregular tubercles and platelets. Tubercles increasing in size from the anterior to the posterior part of the body, reaching maximum dimensions between legs III and IV, where tubercles sometimes merge and form irregular platelets (Figs 2–5). Irregular tubercles 1.0–6.0 μm in diameter. Ventral cuticle smooth (i.e. without sculpturing). Gibbosities and cuticular pores absent.
Bucco-pharyngeal apparatus of the Hypsibius type, without the ventral lamina, and with forked apophyses for stylet muscles (Fig. 6). Peribuccal lamellae absent. Teeth in the oral cavity armature absent or not visible under PCM. Pharyngeal bulb with apophyses and with two granular macroplacoids (both, without constrictions). Macroplacoid length sequence 2<1. Microplacoid and septulum absent.
Claws of the Hypsibius type, internal claws much smaller and of a different shape than the external claws (Fig. 7). All main branches with large accessory points. Smooth, indistinct areoles under claws usually visible only on posterior claws IV. Cuticular bars under claws I-III absent but a small bar is present near the posterior claw IV (Fig. 7, arrow).
Eggs. Unknown.
Remarks.
Hypsibius scabropygus has been recorded from many localities, mostly in the Holarctic (McInnes 1994). In general, our specimens correspond to the original description by Cuénot (1929, 1932) and later descriptions by Marcus (1930) (=Hypsibius callimerus spec. nov.), and by Ramazzotti and Maucci (1983). However, importantly, none of the above mentioned descriptions reported a bar between anterior and posterior claws IV, which is present in all our specimens. Given the bar is small, it is possible that it was overlooked by Cuénot and later authors. If, however, Hypsibius scabropygus does not have the bar, then our specimens should probably be classified as a new species. Thus, until Hypsibius scabropygus is redescribed, our Latvian and Estonian records should be regarded as Hypsibius cf. scabropygus. As there is a possibility of our specimens belonging to a new species, we provide standard morphometrics (Table 1) and photographs (Figs 1–7).
Subfamily: Itaquasconinae Rudescu, 1964
Genus: Astatumen Pilato, 1997
Astatumen bartosi
(Węglarska, 1959)
Localities and specimen numbers.
XIV: 1 specimen.
Remarks.
Our specimen corresponds perfectly with characters of Astatumen bartosi proposed by Dastych (1988) with the main difference between Astatumen bartosi and Astatumen trinacriae being the absence/presence of cuticular bars on legs II–III. Due to the notorious difficulties in differentiating the two species, the actual distribution of Astatumen bartosi cannot currently be described with confidence. McInnes (1994) cited this species from several localities in Europe and from single African, Asian and South American sites.
Superfamily: Isohypsibioidea Marley, McInnes & Sands, 2011
Family: Isohypsibiidae Marley, McInnes & Sands, 2011
Genus: Isohypsibius Thulin, 1928
Isohypsibius ronsisvallei
Binda & Pilato, 1969
Localities and specimen numbers.
III: 1 specimen.
Remarks.
The species has previously been reported from several, mostly European, localities in the Holarctic (McInnes 1994).
Isohypsibius sattleri
(Richters, 1902)
Localities and specimen numbers.
IX: 1 specimen, XI: 8 specimens (including 5 simplexes), XIV: 3 specimens, XV: 5 specimens, XVI: 1 specimen.
Remarks.
The species has previously been reported from many localities throughout the World, thus it is considered cosmopolitan (McInnes 1994, Kaczmarek et al. 2014a).
Superfamily: Macrobiotoidea Thulin, 1928 in Marley et al. 2011
Family: Macrobiotidae Thulin, 1928
Genus: Macrobiotus C.A.S. Schultze, 1834
Macrobiotus harmsworthi harmsworthi
Murray, 1907
Localities and specimen numbers.
XI: 1 egg, XIV: 8 specimens, 1 egg.
Remarks.
The species belongs to the harmsworthi group which is widely distributed across a broad range of ecosystems throughout the world (McInnes 1994, Kaczmarek et al. 2014a). In the last decade many new species within this group were described from a variety of localities (Michalczyk and Kaczmarek 2003b, Pilato et al. 2004, Tumanov 2005a, Pilato and Lisi 2006a, b, Pilato et al. 2006a, Kaczmarek et al. 2007, Kaczmarek and Michalczyk 2009b, Pilato and Lisi 2009a, Rossi et al. 2009, see also Kaczmarek et al. 2011 for the diagnostic key to the group). Due to many uncertain reports of Macrobiotus harmsworthi harmsworthi, especially in older literature, the distribution of the species is currently unknown. Specimens found in the present study correspond well to the characters presented in Pilato et al. (2000) and were successfully identified with the key by Kaczmarek et al. (2011).
Macrobiotus hufelandi hufelandi
C.A.S. Schultze, 1833
Localities and specimen numbers.
XIV: 4 specimens, 1 egg.
Remarks.
The species belongs to the hufelandi group which is widely distributed across a broad range of ecosystems throughout the world (McInnes 1994, Kaczmarek et al. 2014a). In the last decade new species belonging to this group have been described from various localities (Pilato et al. 2003, Kaczmarek and Michalczyk 2004, Dastych 2002, 2005, Fontoura et al. 2008, Bartels et al. 2009, Kaczmarek and Michalczyk 2009b, Pilato and Lisi 2009b, Bertolani et al. 2011, Biserov et al. 2011, Pilato et al. 2012, Guidetti et al. 2013, see also Bertolani and Rebecchi 1993 for the diagnostic key to the group). Due to many uncertain reports of Macrobiotus hufelandi hufelandi, especially in older literature, the distribution of the species is currently unknown. Specimens found in the present study correspond well with the redescription by Bertolani and Rebecchi (1993).
Genus: Minibiotus R.O. Schuster, 1980
Minibiotus formosus sp. n.
http://zoobank.org/BDBE49B7-84CF-4FE2-BE55-A399A537DE77
http://species-id.net/wiki/Minibiotus_formosus
http://www.tardigrada.net/register/0012.htm
Table 2.
Measurements and pt values of selected morphological structures of Minibiotus formosus sp. n. mounted in Hoyer’s medium (N – number of specimens/structures measured, RANGE refers to the smallest and the largest structure among all measured specimens; SD – standard deviation, ? – trait oriented unsuitably for measurement).
| CHARACTER | N | RANGE | MEAN | SD | Holotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||||||
| Body length | 7 | 113 | – | 236 | 636 | – | 1034 | 184 | 901 | 39 | 139 | 194 | 848 |
| Buccal tube | |||||||||||||
| Length | 9 | 17.7 | – | 22.9 | – | 20.3 | – | 1.7 | – | 22.9 | – | ||
| Stylet support insertion point | 9 | 9.5 | – | 12.2 | 49.5 | – | 56.2 | 10.9 | 53.8 | 0.9 | 2.0 | 12.2 | 53.3 |
| External width | 9 | 1.3 | – | 2.0 | 7.0 | – | 9.9 | 1.6 | 7.8 | 0.2 | 0.8 | 1.7 | 7.4 |
| Internal width | 9 | 0.5 | – | 0.7 | 2.4 | – | 3.4 | 0.6 | 2.8 | 0.1 | 0.3 | 0.7 | 3.1 |
| Placoid lengths | |||||||||||||
| Macroplacoid 1 | 9 | 1.3 | – | 1.9 | 6.9 | – | 8.3 | 1.5 | 7.6 | 0.2 | 0.4 | 1.8 | 7.9 |
| Macroplacoid 2 | 9 | 1.2 | – | 1.7 | 6.8 | – | 8.3 | 1.5 | 7.5 | 0.2 | 0.4 | 1.7 | 7.4 |
| Macroplacoid 3 | 9 | 1.4 | – | 2.2 | 7.4 | – | 9.6 | 1.7 | 8.2 | 0.3 | 0.6 | 1.9 | 8.3 |
| Microplacoid | 8 | 0.5 | – | 0.9 | 2.4 | – | 4.7 | 0.7 | 3.3 | 0.1 | 0.8 | 0.8 | 3.5 |
| Macroplacoid row | 9 | 4.5 | – | 6.8 | 24.3 | – | 33.7 | 5.5 | 26.9 | 0.7 | 3.0 | 5.9 | 25.8 |
| Placoid row | 8 | 5.2 | – | 7.9 | 27.1 | – | 39.1 | 6.4 | 31.0 | 0.9 | 3.9 | 6.7 | 29.3 |
| Claw 1 lengths | |||||||||||||
| External primary branch | 4 | 4.7 | – | 6.5 | 22.8 | – | 31.6 | 5.8 | 27.7 | 0.8 | 3.7 | 6.2 | 27.1 |
| External secondary branch | 3 | 3.6 | – | 4.3 | 17.5 | – | 20.9 | 3.9 | 19.4 | 0.4 | 1.7 | ? | ? |
| Internal primary branch | 6 | 4.7 | – | 6.6 | 24.3 | – | 30.4 | 5.7 | 27.4 | 0.7 | 2.4 | 6.3 | 27.5 |
| Internal secondary branch | 5 | 3.1 | – | 4.6 | 15.0 | – | 20.4 | 4.0 | 18.7 | 0.5 | 2.3 | 4.6 | 20.1 |
| Claw 2 lengths | |||||||||||||
| External primary branch | 5 | 5.2 | – | 6.9 | 25.2 | – | 34.0 | 5.9 | 28.6 | 0.6 | 3.5 | 5.9 | 25.8 |
| External secondary branch | 4 | 3.3 | – | 4.7 | 16.0 | – | 23.2 | 3.9 | 19.4 | 0.6 | 2.9 | ? | ? |
| Internal primary branch | 5 | 5.2 | – | 6.4 | 27.1 | – | 31.9 | 6.0 | 29.7 | 0.5 | 2.3 | 6.2 | 27.1 |
| Internal secondary branch | 3 | 3.5 | – | 4.1 | 18.5 | – | 20.9 | 3.9 | 19.9 | 0.3 | 1.3 | ? | ? |
| Claw 3 lengths | |||||||||||||
| External primary branch | 5 | 5.1 | – | 6.9 | 27.0 | – | 33.2 | 6.3 | 30.6 | 0.7 | 2.4 | 6.9 | 30.1 |
| External secondary branch | 6 | 3.6 | – | 4.9 | 19.0 | – | 24.1 | 4.6 | 22.1 | 0.5 | 1.8 | 4.9 | 21.4 |
| Internal primary branch | 7 | 5.1 | – | 6.6 | 25.7 | – | 32.5 | 5.9 | 29.2 | 0.6 | 2.5 | 6.4 | 27.9 |
| Internal secondary branch | 4 | 4.1 | – | 4.6 | 20.9 | – | 22.8 | 4.3 | 21.6 | 0.2 | 0.8 | ? | ? |
| Claw 4 lengths | |||||||||||||
| Anterior primary branch | 5 | 6.0 | – | 8.0 | 31.6 | – | 39.6 | 7.0 | 34.2 | 0.8 | 3.5 | ? | ? |
| Anterior secondary branch | 4 | 4.0 | – | 6.0 | 21.2 | – | 29.7 | 5.0 | 24.1 | 0.9 | 3.9 | ? | ? |
| Posterior primary branch | 6 | 6.0 | – | 7.8 | 30.7 | – | 38.4 | 7.0 | 33.9 | 0.7 | 3.1 | ? | ? |
| Posterior secondary branch | 5 | 3.9 | – | 5.5 | 20.6 | – | 27.1 | 4.8 | 23.8 | 0.6 | 2.6 | ? | ? |
Table 3.
Measurements of selected morphological structures of Minibiotus formosus sp. n. eggs mounted in Hoyer’s medium.
| CHARACTER | egg 1 | egg 2 | egg 3 |
|---|---|---|---|
| Diameter of egg without processes | 45.7 | 44.1 | ? |
| Diameter of egg with processes | 55.6 | 55.1 | ? |
| Process height | 4.5–5.2 | 4.8–5.2 | 4.6–5.3 |
| Process base width | 2.8–3.4 | 2.8–3.1 | 2.4–2.6 |
| Process base/height ratio | 57%–69% | 54%–65% | 47%–57% |
| Distance between processes | 2.0–2.5 | 1.9–3.9 | 1.8–2.0 |
| Number of processes on the egg circumference | 30 | 29 | 30 |
Figures 8–11.

Minibiotus formosus sp. n.: 8 habitus (holotype, ventral view) 9 dorsal cuticle with pores (holotype) 10–11 bucco-pharyngeal apparatus (10 dorso-ventral projection, paratype 11 lateral view, paratype). All PCM.
Figures 12–15.

Minibiotus formosus sp. n.: 12 leg II with claws, granulation (arrowhead) and a single large pore (arrow) (holotype) 13 claws IV (paratype) 14 egg (mid-section) 15 egg surface with processes. All PCM.
Type material.
Holotype and 23 paratypes, 24 specimens (including 2 simplexes) and 3 unembryonated eggs).
Type locality.
57°10'33"N, 24°50'32"E, ca. 45 m asl: Latvia, Vidzeme region, Sigulda municipality, Gutmana Cave in the Gauja National Park; moss from rock (1 sample, slide codes: ŁO 2427/*, where the asterisk can be substituted by any of the following numbers: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12).
Description
(measurements in Table 2). Body white/colourless (Fig. 8). Eyes present in 18 of 24 (75%) specimens mounted in Hoyer’s medium. Entire cuticle covered with small (0.4–1.1 μm) and large (1.9–2.5 μm) round or oval pores (Fig. 9). Pores arranged in 9–10 poorly defined transverse bands. Pores on the dorsal cuticle arranged more densely than on the ventral cuticle. A single large pore (diameter: 2.1–2.9 μm) present on external side of legs I–III (Fig. 12, arrow). A ring of pores around the mouth opening absent. Cuticle without granulation, except for legs which are all covered with fine and regular granulation (better developed on legs IV) visible only in larger specimens (Fig. 12, arrowhead).
Mouth antero-ventral. Ten peribuccal papulae present. Bucco-pharyngeal apparatus of the Minibiotus type (Figs 10–11). Oral cavity armature absent or not visible under PCM. Buccal tube with a poorly visible ventral lamina and with an anterior and a posterior bend (both visible in lateral view only, Fig. 11). Buccal tube walls thickened just below the stylet supports insertion point. Pharyngeal apophyses triangular, very near to the first macroplacoid. Three granular macroplacoids and a minute microplacoid present in the pharyngeal bulb. All macroplacoids of similar but not identical sizes, the macroplacoid length sequence: 2<1<3. Septulum absent.
Claws of the Macrobiotus type (Figs 12–13). Primary branches of claws with thin, but obvious accessory points detaching at the apogee of the primary branch curve. Smooth lunules present on all legs, distinctly larger under external and posterior claws. Bars and other cuticular thickenings on legs absent.
Eggs (measurements in Table 3). White/transparent, laid freely (Fig. 14). Spherical, without areolation. Processes in the shape of short, smooth, slightly flexible cones (Fig. 15). Processes are distributed on the surface of the egg close one to another but never in contact. Surface between processes smooth under PCM (Fig. 15).
Remarks.
Since ventral lamina is very poorly visible, the measurements of this structure are not included in Table 2. Three unembryonated eggs have been found alongside the described specimens. Given that Minibiotus formosus sp. n. was the only Minibiotus species in the sample and because no Ramazzottius Binda & Pilato, 1986 was found in the samples, we assumed that these eggs belong to the new species.
Etymology.
Given that we found the composition of small and large pores in the new species beautiful, we decided to name the animal after this impression (in Latin ‘formosus’ means ‘beautiful’).
Type depositories.
Holotype 23 paratypes and 3 eggs are deposited in the Department of Animal Taxonomy and Ecology at the Adam Mickiewicz University (Poznań, Poland).
Differential diagnosis
The new species is most similar to Macrobiotus gumersindoi Guil & Guidetti, 2005, but it differs from it by: the presence of two types of cuticular pores (small and large) in the new species vs pores of a uniform size in Macrobiotus gumersindoi, the absence of a triangular or a pentagonal arrangement of pores placed above a single large pore on legs, the presence of granulation on legs, a different macroplacoid length sequence (2<1<3 in the new species vs 1=2=3 in Macrobiotus gumersindoi), and by slightly larger macroplacoids (I: 1.3–1.9 μm; II: 1.2–1.7 μm III: 1.4–2.2 μm in the new species vs 1.0 μm in Macrobiotus gumersindoi).
Other species to which Minibiotus formosus sp. n. is similar by some characteristics of adult and/or egg morphology (e.g. pores in transverse bands, eggs with conical processes), include species listed below. The new species differs specifically from:
Macrobiotus bisoctus (Horning et al. 1978) by: the absence of trilobed and star-shaped pores (although their presence was not mentioned in the original description, they are clearly visible in Fig. 114 in Horning et al. (1978), and by stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt≈60.3 in Macrobiotus bisoctus (according to Claxton 1998)).
Macrobiotus eichhorni Michalczyk & Kaczmarek, 2004 by: a different arrangement of pores on the dorsal cuticle (9–10 transverse bands in the new species vs 8 bands in Macrobiotus eichhorni), the absence of star-shaped pores, the absence of four pores around the mouth opening, the presence of a single large pore on lateral sides of legs I-III, slightly shorter buccal tube (17.7–22.9 μm in the new species vs 24.7–34.2 μm in Macrobiotus eichhorni), stylet supports inserted in more anterior position (9.5–12.2 μm [pt=49.5–56.2] in the new species vs 16.2–23.8 μm [pt=65.4–70.6] in Macrobiotus eichhorni), a different macroplacoid sequence (2<1<3 in the new species vs 2<3<1 μm in Macrobiotus eichhorni), slightly shorter placoids, and by slightly smaller external claws I–IV (compare Table 2 below and Table 1 in Michalczyk and Kaczmarek 2004 for exact differences in dimensions of placoids and claws).
Macrobiotus furcatus (Ehrenberg, 1859) (according to Binda and Pilato 1992) by: the absence of tri- and quadrilobed cuticular pores, the presence of two types of cuticular pores (small and large in the new species vs uniformly small pores present in Macrobiotus furcatus), the presence of a single large pore on each of legs I–III, the presence of granulation on legs, the absence of the oral cavity armature, stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt≈68.4 in Macrobiotus furcatus), a different macroplacoid length sequence (2<1<3 in the new species vs 2<3<1 in Macrobiotus furcatus), and by egg processes without an obvious flexible portion (and never bifurcated).
Macrobiotus harrylewisi Meyer & Hinton, 2009 by: the absence of tri- and quadrilobed cuticular pores, the presence of two types of pores (small and large) over the entire cuticle in the new species vs small pores present only in the anterior part of the body and large pores present only in the posterior part of the body in Macrobiotus harrylewisi, the presence of a single large pore on each of legs I–III, stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt=61.4–67.6 in Macrobiotus harrylewisi), a different macroplacoid length sequence (2<1<3 in the new species vs 2≤3<1 μm in Macrobiotus harrylewisi), a different shape of egg processes (short, single-tipped cones in the new species vs elongated, tapering cones with bulbous bases in Macrobiotus harrylewisi), a smaller diameter of eggs without and with processes (44.1–45.7 μm and 55.1–55.6 μm in the new species vs 66.1–80.0 μm and 78.2–101.9 μm in Macrobiotus harrylewisi), a slightly lower number of processes on egg circumference (29–30 in the new species vs 32–41 in Macrobiotus harrylewisi), and by smaller egg processes (4.5–5.3 μm in the new species vs 7.6–12.8 μm in Macrobiotus harrylewisi).
Macrobiotus jonesorum Meyer et al., 2011 by: the absence of trilobed and polygonal pores, the presence of two types of cuticular pores (small and large) in the new species vs small pores present only in the anterior part of the body, intermediate in size in the middle of the body and large pores in the posterior part of the body in Macrobiotus jonesorum), the presence of a single large pore on each of legs I-III, the presence of granulation on all legs, a slightly shorter buccal tube (17.7–22.9 μm in the new species vs 24.4–29.6 μm in Macrobiotus jonesorum), stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt=63.0–65.6 in Macrobiotus jonesorum), a slightly smaller external diameter of the buccal tube (1.3–2.0 μm [pt=7.0-9.9] in the new species vs 2.1–2.6 μm [pt=7.4–10.7] in Macrobiotus jonesorum), a different macroplacoid length sequence (2<1<3 in the new species vs 1<2<3 in Macrobiotus jonesorum), slightly shorter macroplacoids II and III (II: 1.2–1.7 μm [pt=6.8–8.6]; III: 1.4–2.2 μm [pt=7.4–9.6] in the new species vs II: 1.9–2.3 μm [pt=7.1–8.8]; III: 2.4–2.6 μm [pt=8.4–9.9] in Macrobiotus jonesorum), a slightly shorter macroplacoid row (4.5–6.8 μm [24.3–33.7] in the new species vs 7.0–8.4 μm [pt=27.0–34.4] in Macrobiotus jonesorum), the presence of a microplacoid, and by slightly shorter primary and secondary branches of external claws I–IV (compare Table 2 below and Table 2 in Meyer et al. 2011).
Macrobiotus keppelensis Claxton, 1998 by: the lack of red pigment granules, the presence of two types of cuticular pores (small and large) in the new species vs pores uniform in size (ca. 1.0 μm) in Macrobiotus keppelensis), the presence of a single large pore on each of legs I-III, a slightly shorter buccal tube (17.7–22.9 μm in the new species vs 24.9–28.4 μm in Macrobiotus keppelensis), stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt≈60.6 in Macrobiotus keppelensis), a different macroplacoid length sequence (2<1<3 in the new species vs 2=3<1 in Macrobiotus keppelensis), a slightly shorter macroplacoid row (4.5–6.8 μm in the new species vs 7.0–7.6 μm in Macrobiotus keppelensis), the lack of a membrane around egg processes, a smaller diameter of eggs with processes (55.1–55.6 μm in the new species vs 65.0–85.0 μm in Macrobiotus keppelensis), a larger number of processes on egg circumference (29–30 in the new species vs ca. 11 in Macrobiotus keppelensis), smaller egg processes (4.5–5.3 μm in the new species vs 11.0–16.0 μm in Macrobiotus keppelensis), narrower egg processes bases (2.4–3.4 μm in the new species vs 9.0–12.0 μm in Macrobiotus keppelensis), and by slightly smaller distances between egg processes (1.8–3.9 μm in the new species vs 4.0–6.0 μm in Macrobiotus keppelensis).
Macrobiotus orthofasciatus Fontoura et al., 2009 by: cuticular pores arranged in 9–10 transverse bands (11 transverse bands present in Macrobiotus orthofasciatus), the absence of tri- and quadrilobed cuticular pores, the presence of two types of pores (small and large) in the new species vs all pores of similar size in Macrobiotus orthofasciatus, the presence of a single large pore on each of legs I–III, the presence of granulation on all legs, stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt=66.5–67.8 in Macrobiotus orthofasciatus), a different shape of egg processes (short, single tip cones without a membrane in the new species vs screw-like processes with a membrane and six areoles in Macrobiotus orthofasciatus), a slightly larger number of processes on egg circumference (29–30 in the new species vs ca. 24 in Macrobiotus orthofasciatus), and by smaller distances between egg processes (1.8–3.9 μm in the new species vs 6.4–6.9 μm in Macrobiotus orthofasciatus).
Macrobiotus poricinctus Claxton, 1998 by: cuticular pores arranged in 9–10 transverse bands (8 transverse bands in Macrobiotus poricinctus), the presence of two types of pores (small and large) in the new species vs uniform pore size in Macrobiotus poricinctus), the presence of a single large pore on each of legs I–III, stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt≈59.5 in Macrobiotus poricinctus), a different macroplacoid length sequence (2<1<3 in the new species vs 2=3<1 in Macrobiotus poricinctus), a different shape of egg processes (short, single-tipped cones without a membrane in the new species vs. screw-like processes within a membrane in Macrobiotus poricinctus), the absence of granulation on egg shell, a larger number of processes on egg circumference (29–30 in the new species vs 18–20 in Macrobiotus poricinctus), slightly smaller egg processes (4.5–5.3 μm in the new species vs 6.5–7.0 μm in Macrobiotus poricinctus), and by smaller distances between egg processes (1.8–3.9 μm in the new species vs 6.0–8.0 μm in Macrobiotus poricinctus).
Macrobiotus pustulatus (Ramazzotti, 1959) by: the absence of triangular and polygonal pores, the presence of two types of cuticular pores (small and large) in the new species vs small pores present only in the anterior part of the body, intermediate in size in the middle of the body and the large pores in the posterior part of the body in Macrobiotus pustulatus), the presence of a single large pore on each of legs I–III and, egg processes without a filiform bristle.
Macrobiotus ramazzottii Binda & Pilato, 1992 by: pores arranged in bands, the presence of two types of pores (small and large) in the new species vs universal pores size in Macrobiotus ramazzottii), the presence of a single large pore on each of legs I–III, the absence of the oral cavity armature, stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt=68.2–68.3 in Macrobiotus ramazzottii), a different macroplacoid length sequence (2<1<3 in the new species vs 3<2<1 in Macrobiotus ramazzottii), and by a lower number of processes on egg circumference (29–30 in the new species vs ca. 34–41 in Macrobiotus ramazzottii).
Macrobiotus subintermedius (Ramazzotti, 1962) by the presence of cuticular pores, the presence of granulation on all legs, and by fully developed lunules (only small open lunules present in Macrobiotus subintermedius).
Macrobiotus vinciguerrae Binda & Pilato, 1992 by: pores arranged in bands, the absence of tri- and quadrilobed pores, the presence of two types of pores (small and large) in the new species vs uniform pore size in Macrobiotus vinciguerrae), the presence of a single large pore on each of legs I–III, the absence of the oral cavity armature, a larger mean body size (184 μm in the new species vs 380 μm in Macrobiotus vinciguerrae), stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt=66.1–68.7 in Macrobiotus vinciguerrae), a different macroplacoid length sequence (2<1<3 in the new species vs 2<3<1 μm in Macrobiotus vinciguerrae), a smaller diameter of eggs without and with processes (44.1–45.7 μm and 55.1–55.6 μm in the new species vs ca. 76.4 μm and ca. 88.0 μm in Macrobiotus vinciguerrae), a slightly larger number of processes on egg circumference (29–30 in the new species vs ca. 26 in Macrobiotus vinciguerrae), egg processes without flexible filaments, smaller egg processes (4.5–5.3 μm in the new species vs ca. 8.2 μm in Macrobiotus vinciguerrae), and by narrower bases of egg processes (2.4–3.4 μm in the new species vs ca. 5.0 μm in Macrobiotus vinciguerrae).
Macrobiotus weglarskae Michalczyk et al., 2005 by: the absence of bi-, trilobed and star-shaped pores, the presence of two types of pores (small and large) in the new species vs uniform pore size in Macrobiotus weglarskae), the absence of 3–5 large triangular or irregularly shaped pores on the caudo-dorsal cuticle above hind legs, the presence of a single large pore on each of legs I-III, a different shape of egg processes (short, single tip cones without a membrane in the new species vs. screw-like processes within a membrane in Macrobiotus weglarskae), a slightly larger number of processes on egg circumference (29–30 in the new species vs ca. 24 in Macrobiotus weglarskae), and by slightly wider bases of egg processes (2.4–3.4 μm in the new species vs 1.6–2.0 μm in Macrobiotus weglarskae).
Macrobiotus xavieri Fontoura et al., 2009 by: the absence of trilobed pores, the presence of two types of pores (small and large) in the new species vs all pores of similar size in Macrobiotus xavieri), the presence of a single large pore on each of legs I–III, the presence of granulation on all legs, a smaller body size (113–236 μm in the new species vs 275–410 μm in Macrobiotus xavieri), stylet supports inserted in a more anterior position (pt=49.5–56.2 in the new species vs pt=66.1–67.9 in Macrobiotus xavieri), a different macroplacoid length sequence (2<1<3 in the new species vs 2<3<1 in Macrobiotus xavieri). shorter macroplacoids (I: 1.3–1.9 μm [pt=6.9–8.3]; II: 1.2–1.7 μm [pt=6.8–8.6] III: 1.4–2.2 μm [pt=7.4–9.6] in the new species vs I: 3.6–4.5 μm [12.7–13.8]; II: 2.9–3.6 μm [10.3–11.1] III: 3.0–3.9 μm [pt=10.9–11.9] in Macrobiotus xavieri), a shorter microplacoid (0.5–0.9 μm [pt=2.4–4.7] in the new species vs 1.5–2.0 [5.0–6.2] in Macrobiotus xavieri), a shorter macroplacoid row (4.5–6.8 μm [pt=24.3–33.7] in the new species vs 9.8–12.6 μm [pt=35.6–38.5] in Macrobiotus xavieri), a shorter placoid row (5.2–7.9 μm [pt=27.1–39.1] in the new species vs 10.9–13.9 μm [39.6–43.3] in Macrobiotus xavieri), a different shape of egg processes (short, single-tipped cones in the new species vs long cones with bi- or multi-tipped tips in Macrobiotus xavieri), egg shell and processes without granulation, a smaller diameter of eggs without and with processes (44.1–45.7 μm and 55.1–55.6 μm in the new species vs 56.0–79.0 μm and 80.0–99.2 μm in Macrobiotus xavieri), a larger number of processes on egg circumference (29–30 in the new species vs 20–23 in Macrobiotus xavieri), smaller egg processes (4.5–5.3 μm in the new species vs 10.6–19.0 μm in Macrobiotus xavieri), and by slightly narrower bases of egg processes (2.4–3.4 μm in the new species vs 3.7–6.6 μm in Macrobiotus xavieri).
Genus: Paramacrobiotus Guidetti, Schill, Bertolani, Dandekar & Wolf, 2009
Paramacrobiotus richtersi
(Murray, 1911)
Localities and specimen numbers.
XI: 2 specimens (including 1 simplex) and 1 egg.
Remarks.
Paramacrobiotus species (until recently a collection of species within Macrobiotus) can be divided into three groups: areolatus, huziori and richtersi, with respect to the combination of two traits: the presence/absence of the microplacoid in the pharynx and the type of egg areolation. Paramacrobiotus richtersi, considered cosmopolitan, is recognised as the nominal species for a group of very similar taxa that require careful taxonomic examination of adults and egg morphology for correct identification. In the last decade many new species of this group have been described from various localities (e.g. Pilato et al. 2004, Kaczmarek et al. 2005, Tumanov 2005b, Michalczyk and Kaczmarek 2006a, b, Michalczyk et al. 2006, Pilato et al. 2006a, b, Degma et al. 2008, Bartels et al. 2009, Pilato et al. 2012), with more recent additions including molecular data (Guidetti et al. 2009, Schill et al. 2010).
Supplementary Material
Acknowledgements
KZ would like to express sincere thanks to Małgorzata Kuźnik for her help in preparing figures.
Citation
Zawierucha K, Dziamięcki J, Jakubowska N, Michalczyk Ł, Kaczmarek Ł (2014) New tardigrade records for the Baltic states with a description ofMinibiotus formosus sp. n. (Eutardigrada, Macrobiotidae). ZooKeys 408: 81–105. doi: 10.3897/zookeys.408.6612
References
- Argue CW. (1974) Tardigrades from New Brunswick, Canada. 3. Canadian Journal of Zoology 52: 919-992. doi: 10.1139/z74-122 [DOI] [PubMed] [Google Scholar]
- Bartels PJ, Pilato G, Lisi O, Nelson DR. (2009) Macrobiotus (Eutardigrada, Macrobiotidae) from the Great Smoky Mountains National Park, Tennessee/North Carolina, USA (North America): two new species and six new records. Zootaxa 202: 45-57 [Google Scholar]
- Beasley CW, Kaczmarek Ł, Michalczyk Ł. (2008) Doryphoribius mexicanus, a new species of Tardigrada (Eutardigrada: Hypsibiidae) from Mexico (North America). Proceedings of the Biological Society of Washington 121(1): 34-40. doi: 10.2988/07-30.1 [Google Scholar]
- Bertolani R, Rebecchi L. (1993) A revision of the Macrobiotus hufelandi group (Tardigrada, Macrobiotidae), with some observations on the taxonomic characters of eutardigrades. Zoologica Scripta 22: 127-152. doi: 10.1111/j.1463-6409.1993.tb00347.x [Google Scholar]
- Bertolani R, Rebecchi L, Giovannini I, Cesari M. (2011) DNA barcoding and integrative taxonomy of Macrobiotus hufelandi C.A.S. Schultze 1834, the first tardigrade species to be described, and some related species. Zootaxa 2997: 19-36 [Google Scholar]
- Binda MG, Pilato G. (1992) Minibiotus furcatus, nuova posizione sistematica per Macrobiotus furcatus Ehrenberg, 1859, e descrizione di due nuove specie. Animalia 19: 111-120 [Google Scholar]
- Biserov VI, Pilato G, Lisi O. (2011) Macrobiotus turnovae sp. n., a new species of tardigrade from Russia. Invertebrate Zoology 8(1): 57-62 [Google Scholar]
- Cuénot L. (1929) Description d’un tardigrade nouveau de la faune francaise. Archives d’anatomie Microscopique 25: 121-125 [Google Scholar]
- Cuénot L. (1932) Tardigrades. In: Lechevalier P. (Ed) Faune de France 24: 1-96
- Claxton SK. (1998) A revision of the genus Minibiotus (Tardigrada: Macrobiotidae) with descriptions of eleven new species from Australia. Records of the Australian Museum 50: 125-160. doi: 10.3853/j.0067-1975.50.1998.1276 [Google Scholar]
- Dastych H. (1980) Niesporczaki (Tardigrada) Tatrzańskiego Parku Narodowego. Monografie Fauny Polski 9: 1-232 [Google Scholar]
- Dastych H. (1988) The Tardigrada of Poland. Monografie Fauny Polski 16: 1-255 [Google Scholar]
- Dastych H. (1990) Isohypsibius sattleri (Richters 1902), a valid species (Tardigrada). Senckenbergiana Biologica 71: 181-189 [Google Scholar]
- Dastych H. (2002) A new species of the genus Macrobiotus Schultze, 1834 from Iles Kerguélen, in the sub-Antarctic (Tardigrada). Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut 99: 11-27 [Google Scholar]
- Dastych H. (2005) Macrobiotus ramoli sp. n., a new tardigrade species from the nival zone of the Ötztal Alps, Austria (Tardigrada). Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut 102: 21-35 [Google Scholar]
- Degma P, Bertolani R, Guidetti R. (2013) Actual checklist of Tardigrada species. Ver. 23: 15–07–2013 http://www.tardigrada.modena.unimo.it/miscellanea/Actual%20checklist%20of%20Tardigrada.pdf
- Degma P, Michalczyk Ł, Kaczmarek Ł. (2008) Macrobiotus derkai, a new species of Tardigrada (Eutardigrada, Macrobiotidae, huzori group) from the Colombian Andes (South America). Zootaxa 1731: 1-23 [Google Scholar]
- Ehrenberg CG. (1859) Beitrag zur Bestimmung des stationären mikroscopischen Lebens in bis 20,000 Fuss Alpenhöhe. Abhandlungen der Königlichen Akademie der Wissenschaften in Berlin 1858: 429-456 [Google Scholar]
- Fontoura P, Pilato G. (2007) Diphascon (Diphascon) faialense sp. n. a new species of Tardigrada (Eutardigrada, Hypsibiidae) from Azores and a key to the species of the D. pingue group. Zootaxa 1589: 47-55 [Google Scholar]
- Fontoura P, Pilato G, Lisi O. (2008) New records of eutardigrades (Tardigrada) from Faial and Pico Islands, the Azores, with the description of two new species. Zootaxa 1778: 37-47 [Google Scholar]
- Fontoura P, Pilato G, Lisi O, Morais P. (2009a) Tardigrades from Portugal: four new records and description of two new species. Zootaxa 2030: 21-38 [Google Scholar]
- Fontoura P, Pilato G, Morais P, Lisi O. (2009b) Minibiotus xavieri, a new species of tardigrade from Parque Biológico de Gaia, Portugal (Eutardigrada: Macrobiotidae). Zootaxa 2267: 55-64 [Google Scholar]
- Guidetti R, Bertolani R, Degma P. (2007) New taxonomic position of several Macrobiotus species (Eutardigrada: Macrobiotidae). Zootaxa 1471: 61-68 [Google Scholar]
- Guidetti R, Schill RO, Bertolani R, Dandekar T, Wolf M. (2009) New molecular data for tardigrade phylogeny, with the erection of Paramacrobiotus gen. nov. Journal of Zoological Systematics and Evolutionary Research 47: 315-321. doi: 10.1111/j.1439-0469.2009.00526.x [Google Scholar]
- Guidetti R, Peluffo JR, Rocha AM, Cesari M, Moly de Peluffo MC. (2013) The morphological and molecular analyses of a new South American urban tardigrade offer new insights on the biological meaning of the Macrobiotus hufelandi group of species (Tardigrada: Macrobiotidae). Journal of Natural History. doi: 10.1080/00222933.2013.800610
- Guil N, Guidetti R. (2005) A new species of Tardigrada (Eutardigrada: Macrobiotidae) from Iberian Peninsula and Canary Islands (Spain). Zootaxa 889: 1-11 [Google Scholar]
- Holt BG, Lessard J-P, Borregaard KM, Fritz SA, Araújo MB, Dimitrov D, Fabre P-H, Graham CH, Graves GR, Jønsson KA, Nogués-Bravo D, Wang Z, Whittaker RJ, Fjeldså J, Rahbek C. (2012) An Update of Wallace’s Zoogeographic Regions of the World. Science 339(6115): 74-78. doi: 10.1126/science.1228282 [DOI] [PubMed] [Google Scholar]
- Horning DS, Schuster RO, Grigarick AA. (1978) Tardigrada of New Zealand. New Zealand Journal of Zoology 5: 185-280. doi: 10.1080/03014223.1978.10428316 [Google Scholar]
- Kaczmarek Ł, Michalczyk Ł. (2004) New records of Tardigrada from Cyprus with a description of the new species Macrobiotus marlenae (hufelandi group) (Eutardigrada: Macrobiotidae). Genus 15(1): 141-152 [Google Scholar]
- Kaczmarek Ł, Michalczyk Ł. (2009a) Redescription of Hypsibius microps Thulin, 1928 and H. pallidus Thulin, 1911 (Eutardigrada: Hypsibiidae) based on the type material from the Thulin collection. Zootaxa 2275: 60-68 [Google Scholar]
- Kaczmarek Ł, Michalczyk Ł. (2009b) Two new species of Macrobiotidae, Macrobiotus szeptyckii (harmsworthi group) and Macrobiotus kazmierskii (hufelandi group) from Argentina. Acta Zoologica Cracoviensia 52B: 87–99. doi: 10.3409/azc.52b_1-2.87-99 [Google Scholar]
- Kaczmarek Ł, Michalczyk Ł, Degma P. (2007) Description of a new tardigrade, Macrobiotus barbarae (Eutardigrada: Macrobiotidae), from the Dominican Republic. Annales Zoologici 57(3): 363-369 [Google Scholar]
- Kaczmarek Ł, Michalczyk Ł, Diduszko D. (2005) Some tardigrades from Siberia (Russia, Baikal region) with a description of Macrobiotus garynahi sp. n. (Eutardigrada: Macrobiotidae: richtersi group). Zootaxa 1053: 35-45 [Google Scholar]
- Kaczmarek Ł, Michalczyk Ł, McInnes SJ. (2014a) Annotated zoogeography of non-marine Tardigrada. Part I: Central America. Zootaxa 3763(1): 1-62. doi: 10.11646/zootaxa.3763.1.1 [DOI] [PubMed] [Google Scholar]
- Kaczmarek Ł, Cytan J, Zawierucha K, Diduszko D, Michalczyk Ł. (2014b) Tardigrades from Peru (South America), with descriptions of three new species of Parachela. Zootaxa 3790(2): 357-379. doi: 10.11646/zootaxa.3790.2.5 [DOI] [PubMed] [Google Scholar]
- Kaczmarek Ł, Gołdyn B, Prokop ZM, Michalczyk Ł. (2011) New records of Tardigrada from Bulgaria with the description of Macrobiotus binieki sp. n. (Eutardigrada: Macrobiotidae) and a key to the species of the harmsworthi group. Zootaxa 2781: 29-39 [Google Scholar]
- Kaczmarek Ł, Zawierucha K, Smykla J, Michalczyk Ł. (2012) Tardigrada of the Revdalen (Spitsbergen) with the descriptions of two new species: Bryodelphax parvuspolaris (Heterotardigrada) and Isohypsibius coulsoni (Eutardigrada). Polar Biology 35(7): 1013-1026. doi: 10.1007/s00300-011-1149-0 [Google Scholar]
- Li X, Wang D, Wang L. (2008) The Tardigrada fauna of Hainan Island (Asia: China) with descriptions of two new species. Raffles Bulletin of Zoology 56(2): 293-305 [Google Scholar]
- Marcus E. (1930) Beiträge zur Tardigradensystematik. Zoologische Jahrbuecher Systematik 59: 363-386 [Google Scholar]
- Marley NJ, McInnes SJ, Sands CJ. (2011) Phylum Tardigrada: A re-evaluation of the Parachela. Zootaxa 2819: 51-64 [Google Scholar]
- McInnes SJ. (1994) Zoogeographic distribution of terrestrial/freshwater tardigrades from current literature. Journal of Natural History 28: 257-352. doi: 10.1080/00222939400770131 [Google Scholar]
- Meyer HA. (2012) A new water bear, Minibiotus julianae, from the Caribbean Island of Dominica (Tardigrada: Eutardigrada: Parachela: Macrobiotidae). Proceedings of the Biological Society of Washington 125(1): 54-60. doi: 10.2988/11-15.1 [Google Scholar]
- Meyer HA, Domingue MD. (2011) Minibiotus acadianus (Eutardigrada: Macrobiotidae), a new species of Tardigrada from southern Louisiana, USA. Western North American Naturalist 71(1): 38-43. doi: 10.3398/064.071.0106 [Google Scholar]
- Meyer HA, Hinton JG. (2009) The Tardigrada of southern Africa, with the description of Minibiotus harrylewisi, a new species from KwaZulu-Natal, South Africa (Eutardigrada: Macrobiotidae). African Invertebrates 50(2): 255-268. doi: 10.5733/afin.050.0203 [Google Scholar]
- Meyer HA, Lyons AM, Nelson DR, Hinton JG. (2011) Tardigrada of Michigan, Northern USA, with the description of Minibiotus jonesorum sp. n. (Eutardigrada: Macrobiotidae). Journal of Zoological Systematics and Evolutionary Research 49(S1): 40–47. doi: 10.1111/j.1439-0469.2010.00596.x [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł. (2003a) Minibiotus constellatus, a new species of Tardigrada (Eutardigrada, Macrobiotidae) from Peru. Genus 14(2): 295-305 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł. (2003b) A description of the new tardigrade Macrobiotus reinhardti (Eutardigrada, Macrobiotidae, harmsworthi group) with some remarks on the oral cavity armature within the genus Macrobiotus Schultze. Zootaxa 331: 1-24 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł. (2004) Minibiotus eichhorni sp. n., a new species of eutardigrade (Eutardigrada: Macrobiotidae) from Peru. Annales Zoologici 54: 673-676 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł. (2006a) Macrobiotus huziori, a new species of Tardigrada (Eutardigrada: Macrobiotidae) from Costa Rica (Central America). Zootaxa 1169: 47-59 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł. (2006b) A new species Macrobiotus magdalenae (Tardigrada: Eutardigrada: Macrobiotidae, richtersi group) from Costa Rican rain forest (Central America). New Zealand Journal of Zoology 33: 189-196. doi: 10.1080/03014223.2006.9518444 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł. (2013) The Tardigrada Register: a comprehensive online data repository for tardigrade taxonomy. Journal of Limnology 72(S1): 175–181. doi: 10.4081/jlimnol.2013.s1.e22 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł, Claxton SK. (2005) Minibiotus weglarskae, a new species of Tardigrada (Eutardigrada: Macrobiotidae) from Mongolia. Zootaxa 1008: 47-56 [Google Scholar]
- Michalczyk Ł, Kaczmarek Ł, Weglarskae B. (2006) Macrobiotus sklodowskae sp. n. (Tardigrada: Eutardigrada: Macrobiotidae, richtersi group) from Cyprus. Zootaxa 1371: 45-46 [Google Scholar]
- Michalczyk Ł, Wełnicz W, Frohme M, Kaczmarek Ł. (2012a) Redescriptions of three Milnesium Doyère, 1840 taxa (Tardigrada: Eutardigrada: Milnesiidae), including the nominal species for the genus. Zootaxa 3154: 1-20 [Google Scholar]
- Michalczyk Ł, Wełnicz W, Frohme M, Kaczmarek Ł. (2012b) Corrigenda of Zootaxa 3154: 1–20 Redescriptions of three Milnesium Doyère, 1840 taxa (Tardigrada: Eutardigrada: Milnesiidae), including the nominal species for the genus. Zootaxa 3393: 66-68 [Google Scholar]
- Miller WR, McInnes SJ, Bergstrøm, DM. (2005) Tardigrades of the Australian Antarctic: Hypsibius heardensis (Eutardigrada: Hypsibiidae: dujardini group) a new species from sub-Antarctic Heard Island. Zootaxa 1022: 57-64 [Google Scholar]
- Murray J. (1905) The Tardigrada of the Forth Valley. The Annals of Scottish Natural History 55: 160-164 [Google Scholar]
- Pilato G. (1981) Analisi di nuovi caratteri nello studio degli Eutardigradi. Animalia 8: 51-57 [Google Scholar]
- Pilato G. (1987) Revision of the genus Diphascon Plate, 1889, with remarks on the subfamily Itaquasconinae (Eutardigrada, Hypsibiidae). In: Bertolani (Ed) Biology of Tardigrades Selected Symposia and Monographs U.Z.I., 1: 337–357
- Pilato G. (2013) The taxonomic value of the structures for the insertion of the stylet muscles in the Eutardigrada, and description of a new genus. Zootaxa 3721(4): 365-378. doi: 10.11646/zootaxa.3721.4.4 [DOI] [PubMed] [Google Scholar]
- Pilato G, Binda MG. (1997/1998) A comparison of Diphascon (D.) alpinum Murray, 1906, D. (D.) chilenense Plate, 1889 and D. (D.) pingue Marcus, 1936 (Tardigrada), and description of a new species. Zoologischer Anzeiger 236: 181-185 [Google Scholar]
- Pilato G, Binda MG. (1999) Three new species of Diphascon of the pingue group (Eutardigrada, Hypsibiidae) from Antarctica. Polar Biology 21: 335-342. doi: 10.1007/s003000050370 [Google Scholar]
- Pilato G, Claxton SK. (1988) Tardigrades from Australia. 1. Macrobiotus hieronimi and Minibiotus maculartus, two new species of eutardigrades. Animalia 15: 83-89 [Google Scholar]
- Pilato G, Lisi O. (2006a) Macrobiotus rigidus sp. n., a new species of eutardigrade from New Zealand. Zootaxa 1109: 49-55 [Google Scholar]
- Pilato G, Lisi O. (2006b) Notes on some tardigrades from southern Mexico with description of three new species. Zootaxa 1236: 53-68 [Google Scholar]
- Pilato G, Lisi O. (2009a) Tardigrades of the Seychelles Islands, with the description of three new species. Zootaxa 2124: 1-20 [Google Scholar]
- Pilato G, Lisi O. (2009b) Description of three new species of Tardigrada from Seychelles. Zootaxa 2005: 24-34 [Google Scholar]
- Pilato G, Binda MG, Lisi O. (2003) Remarks on some species of tardigrades from South America with the description of Minibiotus sidereus n. sp. Zootaxa 195: 1-8 [Google Scholar]
- Pilato G, Binda MG, Lisi O. (2004) Notes on tardigrades of the Seychelles with the description of three new species. Italian Journal of Zoology 71: 171-178. doi: 10.1080/11250000409356569 [Google Scholar]
- Pilato G, Binda MG, Lisi O. (2006a) Three new species of eutardigrades from the Seychelles. New Zealand Journal of Zoology 33: 39-48. doi: 10.1080/03014223.2006.9518429 [Google Scholar]
- Pilato G, Binda MG, Lisi O. (2006b) Eutardigrada from New Zealand, with descriptions of two new species. New Zealand Journal of Zoology 33: 49-63. doi: 10.1080/03014223.2006.9518430 [Google Scholar]
- Pilato G, Binda MG, Napolitano A, Moncada E. (2000) The specific value of Macrobiotus coronatus DeBarros 1942, and description of two new species of the harmsworthi group (Eutardigrada). Bollettino delle sedute della Accademia Gioenia di Scienze Naturali in Catania 33: 103-120 [Google Scholar]
- Pilato G, Kaczmarek Ł, Michalczyk Ł, Lisi O. (2003) Macrobiotus polonicus, a new species of Tardigrada from Poland (Eutardigrada, Macrobiotidae, ‘hufelandi group’). Zootaxa 258: 1-8 [Google Scholar]
- Pilato G, Kiosya Y, Lisi O, Sabella G. (2012) New records of Eutardigrada from Belarus with the description of three new species. Zootaxa 3179: 39-60 [Google Scholar]
- Ramazzotti G. (1959) Il gruppo dell’ Echiniscus viridis con la nuova species E. perviridis e Macrobiotus pustulatus altra nuova specie (Tardigrada). Atti della Societa ltaliana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 98: 303-309 [Google Scholar]
- Ramazzotti G. (1962) Tardigradi del Cile, con descrizione di quattro nuove specie e di una varietá. Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano 101: 275-287 [Google Scholar]
- Ramazzotti G, Maucci W. (1983) Il Phylum Tardigrada. Memorie dell’Istituto Italiano di Idrobiologia 41: 1-1012 [Google Scholar]
- Rossi G, Claps M, Ardohain D. (2009) Tardigrades from northwestern Patagonia (Neuquén Province, Argentina) with the description of three new species. Zootaxa 2095: 21-36 [Google Scholar]
- Šatkauskienė I, Vosyliūtė R. (2010) Microfauna of Moss (Bryophyta: Bryopsida) from four regions of Lithuania. Acta Zoologica Lituanica 20: 225-231. doi: 10.2478/v10043-010-0024-5 [Google Scholar]
- Schill RO, Forster F, Dandekar T, Wolf N. (2010) Using compensatory base change analysis of internal transcribed spacer 2 secondary structures to identify three new species in Paramacrobiotus (Tardigrada). Organisms Diversity & Evolution 10(4): 287-296. doi: 10.1007/s13127-010-0025-z [Google Scholar]
- Tumanov DV. (2005a) Two new species of Macrobiotus (Eutardigrada, Macrobiotidae) from Tien Shan (Kirghizia), with notes on Macrobiotus tenuis group. Zootaxa 1043: 33-46 [Google Scholar]
- Tumanov DV. (2005b) Notes on the Tardigrada of Thailand, with description of Macrobiotus alekseevi sp. n. (Eutardigrada, Macrobiotidae). Zootaxa 999: 1-16 [Google Scholar]
- Tumanov DV. (2006) Five new species of the genus Milnesium (Tardigrada, Eutardigrada, Milnesiidae). Zootaxa 1122: 1-23 [Google Scholar]
- Zawierucha K, Kaźmierski A. (2012) The first records of tardigrades (Tardigrada, Eutardigrada) from Estonia. Zoology and Ecology 22: 111-113. doi: 10.1080/21658005.2012.696885 [Google Scholar]
- Ziemelis A, Purina L, Ozolinš A. (2012) A Short-term study of population Dynamics of tardigrades in the moss Leucodon sciuroides (Hedw.) SCHWÄGR. Latvijas Universitātes 70 zinātniskā konference Bioloăijas sekcija, Zooloăijas un dzīvnieku ekoloăijas apakšsekcija (conference abstract). [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.