

SUMMARY
Recent cell biological studies suggest that sphingolipids and cholesterol may cluster in biomembranes to form raft-like microdomains. Such lipid domains are postulated to function as platforms involved in the lateral sorting of certain proteins during their trafficking within cells as well as during signal transduction events. Here, the physical interactions that occur between cholesterol and sphingolipids in model membrane systems are discussed within the context of microdomain formation. A model is presented in which the role of cholesterol is refined compared to earlier models.
Keywords: Sphingolipid-cholesterol raft, Caveolar lipid, Lipid solubilization via detergent
INTRODUCTION
One of the most intriguing questions about cell membranes is why these supramolecular ensembles contain so many different types and species of lipids. After all, if the chief purpose of membranes is to compartmentalize cells while serving as selective permeability barriers in the export and import of various cellular materials, then only a handful of lipids should be necessary. With the realization that many membrane lipids themselves serve as reservoirs of metabolically derived messenger signals, the original question seems much less daunting. Yet, to effectively regulate such a complex ensemble necessitates an inherent organization much greater than originally envisioned when the bilayer lipids were thought to function mainly as a fluid solvent to facilitate protein diffusion (Singer and Nicolson, 1972). Hence, recent advances in the lipid cell signaling field not only have provided enlightenment regarding the need for diversity among membrane lipids but also have served to renew interest in the physical organization of lipids within biomembranes (e.g. Vaz and Almeida, 1993; Jacobson et al., 1995).
In particular, the postulated occurrence of sphingolipid-cholesterol rafts or microdomains in biomembranes has been at the forefront of new developments in the membrane trafficking and signaling field. In essence, strong and preferential lateral interactions that putatively occur between sphingolipids and cholesterol lead to formation of microdomains or ‘rafts’ within biomembranes. These sphingolipid-cholesterol rafts are postulated to be platforms that directly participate, i.e. function, in the lateral recruitment of certain types of proteins while trafficking through the Golgi as well as in the association of proteins with GPI-anchors to non-clatherin coated plasma membrane invaginations called caveolae (e.g. Bretscher and Munro, 1993; Parton and Simons, 1995; Simons and Ikonen, 1997). Hence, purely physical interactions between the lipids promote membrane domain formation. More importantly, the ability of the domains or rafts to laterally segregate proteins during the sorting process also is attributed to differences in the physical environment within the lipid domains themselves compared to other membrane regions (Schroeder et al., 1994; Simons and Ikonen, 1997). Because of the widespread interest in the involvement of GPI-anchored proteins in intracellular signal transduction events, the preceding ideas have received a good deal of attention and have been enthusiastically debated in the recent literature. Here, I will provide an overview of the recent advances relating to the physical interactions that occur between sphingolipids and other membrane lipids (e.g. cholesterol) in model membrane systems in the hopes that this will provide useful and fundamental insights into the situation(s) likely to exist in biomembranes. For comprehensive coverage of pre-1993 advances regarding sphingolipid physical properties, several reviews are available (e.g. Maggio, 1994; Thompson and Brown, 1988; Curatolo, 1987; Thompson et al., 1986; Thompson and Tillack, 1985; Barenholz and Thompson 1980). An excellent discussion of lipid domains can be found in a review by Welti and Glaser (1994). Discussions focused on cholesterol domains in membranes can be found in monographs (e.g. Finegold, 1993) as well as in reviews by Schroeder et al. (1991, 1996) and by Slotte (Slotte et al., 1994; Slotte, 1997). Other recent articles include commentary and pictorial schemes describing putative cell biological roles of sphingolipid-cholesterol rafts during the sorting and trafficking of GPI-anchored proteins and related functional activities in membrane signaling (Anderson, 1993; Lisanti et al., 1994; Futerman, 1995; Parton and Simons, 1995; Parton, 1996; Liu et al., 1997; Harder and Simons, 1997; Simons and Ikonen, 1997; Weimbs et al., 1997). While the focus here will be on mammalian lipids, readers are reminded that sphingolipids and sterols are ubiquitous among eukaryotes (Hori and Sugita, 1993; Lester and Dickson, 1993; van der Rest et al., 1995). Hence, specialized roles ascribed to sphingolipid-sterol rafts during the intracellular sorting of GPI-anchored proteins may be widespread among eukaryotes (e.g. Zhang and Thompson, 1997).
DIG COMPOSITION – REALITY OR ARTIFACT?
A key and controversial issue regarding sphingolipid-cholesterol rafts is whether the biochemical method used to isolate these structures actually harvests pre-existing microdomains or creates them artifactually. The original method of raft isolation was based on the resistance of certain membrane components (e.g. GPI-anchored proteins, sphingolipids, cholesterol) to solubilization by the detergent, Triton X-100, at 4°C (Brown and Rose, 1992). The idea that lateral phase separation of sphingolipids in biomembranes might explain their resistance to detergent extraction dates back to the work of Yu et al. (1973). However, despite the resistance to solubilization exhibited by select proteins and lipids, many different membrane proteins, the bulk of the phosphoglycerides and a large amount of cholesterol are solubilized (Brown and Rose, 1992). Separation of the soluble and insoluble components is achieved by density gradient centrifugation. Hence, sphingolipid-cholesterol rafts sometimes are referred to as detergent-insoluble glycolipid-enriched complexes (DIGs) (Parton and Simons, 1995). The presence of the cholesterol-binding protein, VIP-caveolin, allows discrimination between caveolae and other types of DIGs (Dupree et al., 1993). Using CHAPS instead of Triton X-100 reportedly results in sphingolipid-cholesterol rafts that are enriched in cholesterol, cerebrosides, sphingomyelin, and ganglioside GM3, but not lactosylceramide, Forssman antigen, or GPI-anchored proteins (Fiedler et al., 1993). Obviously, a key issue regarding such methodology is whether the postulated sphingolipid-cholesterol rafts exist in membranes prior to detergent treatment or whether the insoluble components in the low-density gradient fraction are remnants that once were independent and randomly dispersed in the membranes prior to the selective removal of their neighbors by the detergent. Similar concerns have been expressed regarding the putative association of GPI-anchored proteins and sphingolipids with caveolae because sequestration to caveolae becomes obvious only after cytoskeleton-independent crosslinking (Mayor et al., 1994; Kurzchalia et al., 1995; Gorodinsky and Harris, 1995; Mayor and Maxfield, 1995; Fujimoto, 1996). In this regard, newer means of isolating the DIGs are helping to resolve these issues. The newer detergent-free methods achieve varying degrees of enrichment (Schnitzer et al., 1995; Smart et al., 1995; Song et al., 1996; Stan et al., 1997). However, extensive characterization of the lipids associated with GPI-anchored proteins and/or VIP-caveolin in these newer preparations has not been reported. In this regard, the colloidal silica particle coating procedure of Schnitzer et al. (1995) and immunoisolation method of Stan et al. (1997) appear to be especially promising for defining caveolar lipids. Initial indications are that ganglioside GM1, which reportedly localizes to caveolae (Parton, 1994; Fra et al., 1995), is found in the preparations of Schnitzer et al. (1995) and Smart et al. (1995). Signaling lipids such as sphingomyelin and phosphoinositides also appear to be enriched in the caveolar fraction (Liu et al., 1997).
In the meantime, we must rely on other inferential approaches including the knowledge gathered from model membrane studies. For the past two decades, membrane biophysicists have studied the physical interactions that occur between sphingolipids and other membrane lipids while focusing primarily on model membranes. Although no single experimental approach categorically proves or disproves the existence of sphingolipid-enriched domains in membranes, the results provide fundamental insights into the ways that sphingolipids interact and mix with other lipid components in bilayers. When viewed collectively, the data also indicate that certain sphingolipids almost certainly form small microdomains in otherwise fluid-phase PC bilayers. What has remained less clear is how cholesterol impacts on the stability and size of sphingolipid-enriched microdomains. However, experiments involving phosphatidylcholines with di-saturated acyl chains, which mimic many features of naturally abundant sphingolipids, indicate that cholesterol can affect lateral phase separation when present at physiologically relevant concentrations (Almeida et al., 1993; Silvius et al., 1996). Interestingly, very recent experiments involving sphingolipids indicate that cholesterol can promote phase separation and, in doing so, change the physical properties of the resulting sphingolipid-cholesterol enriched phases (Ahmed et al., 1997).
EARLY REVELATIONS ABOUT SPHINGOLIPID LATERAL ORGANIZATION IN LIPID BILAYERS
The capacity of certain sphingolipids to form clusters or microdomains in phosphatidylcholine (PC) bilayers was postulated many years ago by T. E. Thompson and colleagues (Thompson and Tillack, 1985; Thompson et al., 1986). The ideas evolved from kinetic studies of the intervesicular transfer rates of different lipids (Correa-Freire, 1982; Frank et al., 1983; Brown et al., 1985; Brown and Thompson, 1987), from ultrastructural labeling studies in which lectins and toxins were used to probe the surface localization of glycosphingolipids in PC liposomes prior to visualization by freeze-etch electron microscopy (Tillack et al., 1982; Brown et al., 1983; Grant and Peters, 1984; Thompson et al., 1985), and from scattered spectroscopic and thermotropic studies of sphingolipid mixing behavior in PCs (e.g. Neuringer et al., 1979; Lee and Grant, 1980; Suzuki et al., 1981; Correa-Freire et al., 1979; Bach et al., 1982; Ruocco et al., 1983; Maggio et al., 1985). The key features of the ideas advanced by Thompson were that certain sphingolipids could laterally cluster into microdomains when present as minor constituents (<10 mol%) in liquid-crystalline phase PC vesicles. In fact, an early rendition of glycosphingolipid clustering suggested by Thompson et al. (1986) is shown in Fig. 1.
Fig. 1.

Schematic portraying the molecular arrangement of the glycosphingolipid, asialo-GM1, in a liquid-crystalline PC matrix. One half of the bilayer is depicted. Asialo-GM1 is denoted by the filled tetramerically linked hexagons which represent the four uncharged sugar residues of this glycolipid. The effect of cholesterol on the clustering of asialo-GM1 was not considered. (Adapted from Thompson et al. (1986), copyright 1986, Plenum Publishing Corp.)
Further studies revealed that not all sphingolipids laterally cluster in fluid-phase PC vesicles (Thompson et al., 1985; Brown and Thompson, 1987; Rock et al., 1990). Also, the phase state of the PC matrix, the thickness of the PC matrix, and the PC acyl chain composition all affect the lateral distribution and average cluster size of the glycosphingolipids (Tillack et al., 1982; Dwyer and Bloomfield, 1982; Peters and Grant, 1984; Melhorn et al., 1986). Especially interesting is the increased tendency of certain neutral sphingolipids to cluster in fluid-phase PCs which contain the physiologically relevant saturated sn-1, unsaturated sn-2 acyl motif compared to fluid phase PCs which contain disaturated acyl chains (Tillack et al., 1982). Yet, the average cluster size often is small and may not exceed twenty sphingolipid molecules (Rock et al., 1990). Ganglioside clustering in PC bilayers persists in the presence of cholesterol (Peters et al., 1984). In liposomes containing both gel and liquid-crystalline phase PCs, the phase state of the PC matrix and the ceramide composition influence the localization of the sphingolipid (see Rock et al., 1991; Palestini et al., 1995; and also Peters and Grant, 1984; Peters et al., 1984). Fluorescence studies of model membranes also suggest that cerebrosides and ceramides form microdomains in PC bilayers (Molotkovsky et al., 1991; Holopainen et al., 1997). Fluorescence imaging studies of liposomes and of lipid monolayers containing phosphoglycerides, sphingomyelin, and cholesterol have visualized large macrodomains that form in response to viral proteins (Luan et al., 1995) or in response to low monolayer surface pressures (Slotte, 1995; Mattjus et al., 1995).
Despite all the evidence gathered from the biophysical studies of model membranes in the early and mid 1980s indicating that certain sphingolipids are likely to microcluster in biomembranes (also see Tillack et al., 1983), interest in sphingolipid domain formation began to wane. As a result, many important questions regarding cholesterol’s role in sphingolipid domain formation have gone unanswered. Perhaps, this occurred because no cell biological function could be directly linked to sphingolipid microdomains despite interesting speculation that such structures might function as flexible and rapidly alterable recognition sites on cell surfaces (Thompson et al., 1986; Brown and Thompson, 1987) or might be involved in the trafficking of sphingolipids themselves to the apical plasma membrane surface of polarized epithelial cells (Simons and van Meer, 1988). However, interest in sphingolipid microdomain formation has been renewed with the postulated involvement of sphingolipid-cholesterol domains or rafts in the sorting of proteins in the Golgi as well as in the trafficking and localization of GPI-anchored proteins to caveolar plasma membrane regions. Before turning our attention to recent physical studies pertinent to understanding sphingolipidcholesterol interactions, we first examine the molecular structural features that are likely to play essential roles in regulating the lateral interactions among sphingolipids, phosphoglycerides and cholesterol.
SPHINGOLIPID STRUCTURE: WHAT’S DIFFERENT?
The key structural features that make sphingolipids different from glycerol-based membrane lipids are depicted in Fig. 2. First, consider the hydrocarbon regions of the molecules. Note that, with respect to molecular conformation, the sphingoid base is structurally equivalent to the combination provided by the glycerol base and the sn-1 acyl chain in a typical membrane phosphoglyceride. For this reason, a saturated sn-1 acyl chain in a phosphoglyceride needs only to be about 13.5 carbons long to be equivalent conformationally to an 18-carbon sphingoid base with a 4,5 trans double bond. Yet, saturated sn-1 chains 16 or 18 carbons in length generally predominate in membrane phosphoglycerides (Kuksis and Myher, 1976) while 18-carbon sphingoid bases predominate in the mammalian sphingolipids although the percentage of 20-carbon sphingoid base does increase in complex gangliosides (Yohe et al., 1976).
Fig. 2.
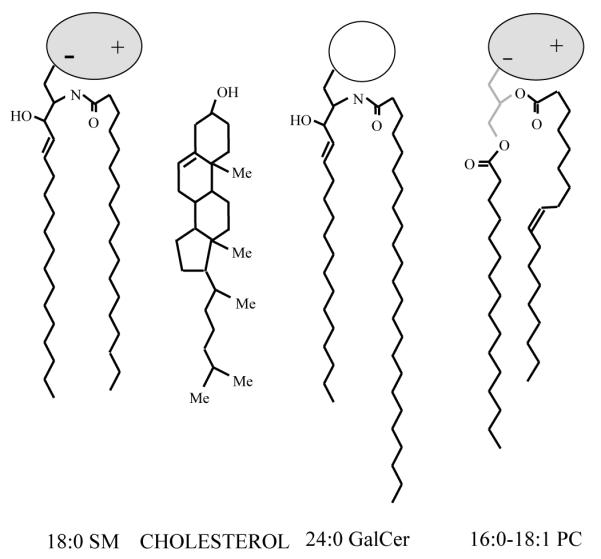
Structural features of typical sphingolipids, cholesterol, and phosphoglycerides. (Sphingomyelin = SM; galactosylceramide = GalCer; glycerophosphocholine = PC; methyl groups in cholesterol = Me). For simplicity, the hydrocarbon chains of the lipids are depicted as fully extended trans rotamers. In reality, free rotation about the carbon-carbon single bonds provides numerous trans-gauche isomers in the fluid but not the gel state. The cis double bond depicted in the sn-2 chain of PC leads to a t+gt− kink. Rotation about nearby carbon-carbon bonds increases the average molecular cross-sectional area. In PC, the glycerol base chain is depicted in gray and the highly hydratable, zwitterionic phosphorylcholine headgroup is depicted as a shaded ellipse oriented roughly parallel to the bilayer surface. In SM, the phosphorylcholine headgroup is depicted similarly and a stearoyl (18:0) acyl chain, typical of bovine brain SM, is shown amide-linked to the 18-carbon sphingosine base. In GalCer, the uncharged and lowly hydratable galactose headgroup is depicted as an unfilled circle, whereas, a lignoceroyl (24:0) acyl chain, typical of bovine brain GalCer, is shown amide-linked to the 18-carbon sphingosine base. In cholesterol, the 3-OH group as well as the methyl (Me) groups at positions 18,19, 21, and 26 all protrude from the β face of the molecule (see Bittman, 1997, for more detail).
When acylated, the sphingoid base is known as a ceramide. The amino group at the second carbon of the sphingoid base serves as the attachment site for the fatty acyl chain via an amide linkage. Typically, acyl chains are long and saturated (Curatolo, 1987). For instance, acyl chains of twenty four carbons are common in bovine brain cerebrosides and in sulfatides although shorter chains (e.g. 18:0) predominate in bovine brain sphingomyelin and gangliosides. The predominance of saturated acyl chains contributes to the high phase transition temperatures characteristic of sphingolipids (Koynova and Caffrey, 1995). Unsaturation in sphingolipid acyl chains is limited almost exclusively to single cis double bonds. Monounsaturated acyl chains are generally minor constituents. When monounsaturated acyl chains account for 30 mol% or more, the cis double bond is usually located at carbon positions (e.g. 24:1Δ15(c) = nervonate) that permit efficient interaction with the planar polycyclic ring system of membrane sterols. The acyl chains of sphingolipids rarely are polyunsaturated (Robinson et al., 1992). This situation markedly contrasts that of membrane phosphoglycerides which typically have polyunsaturated sn-2 chains.
Despite the differences in double bond frequency, the sphingolipid acyl chain and phosphoglyceride sn-2 acyl chain share certain conformational features in that the initial segments of their hydrocarbon chains extend roughly orthogonal to the sphingoid or sn-1 chain, respectively, but then bend sharply between carbons 2 and 3 and thereafter align roughly parallel to the sphingoid or sn-1 chain, respectively (Speyer et al., 1989; Hamilton et al., 1993; Ruocco et al., 1996). The net effect of the similarities and differences in hydrocarbon conformation and composition is to render many membrane sphingolipids rather asymmetric with respect to their hydrocarbon chain lengths (Fig. 2). Hence, naturally occurring sphingolipids have the potential to participate in transbilayer hydrocarbon interdigitation (e.g. Slater and Huang, 1992). This potential is not limited to sphingolipids with saturated acyl chains but persists when the acyl chains are long and monounsaturated acyl chains (Kulkarni and Brown, 1997).
Although sphingolipid transbilayer interdigitation in model membranes might appear to be irrelevant to plasma membranes, where the glycolipids and sphingomyelin are oriented mostly exofacially, one should keep in mind that pools of sphingomyelin are likely to be localized to the cytofacial membrane surface or compartmentalized into caveolae (Liu and Anderson, 1995; Zhang et al., 1997). Sphingomyelinase action during signaling events can produce ceramides, which regulate cell apoptosis. The availability of the signaling sphingomyelin pool to sphingomyelinase may be affected by transbilayer hydrocarbon interdigitation. In this capacity, cholesterol may play a role by controlling whether sphingolipid transbilayer interdigitation occurs. Adding cholesterol to sphingomyelins causes a thinning of gel-phase but not liquid-crystalline phase bilayers as well as more pronounced terminal methyl troughs at the bilayer center in electron density profiles (McIntosh et al., 1992; Maulik and Shipley, 1996a,b). Both these results are consistent with cholesterol interfering with transbilayer chain interdigitation. Also, the solid domains in 24:0 GalCer/di-16:0 PC mixtures in each half of the bilayer do not superimpose across the bilayer (Almeida et al., 1992). Boggs and Koshy (1994) observed no interdigitation of sulfatides with long saturated acyl chains in various PC liquid crystalline bilayers at physiological temperature in the presence of cholesterol. Hence, hydrocarbon chain interdigitation does not appear to be essential for stable domain formation among sphingolipids.
Next, consider the interfacial regions that bridge the nonpolar hydrocarbon regions to the polar headgroups. In sphingolipids, a hydroxyl group is located at the third carbon or at both the third and fourth carbons of the sphingoid base depending whether the base is sphingosine or phytosphingosine, respectively. Phytosphingosine is prevalent among nonneuronal sphingolipids such as those of the kidney and intestinal cells as well as in nonvertebrate eukaryotes (Karlsson, 1970). Moreover, the fatty acyl chains linked to the sphingoid bases are often 2-hydroxylated in simple sphingolipids (e.g. Johnson and Brown, 1992). As a result, an abundance of hydrogen bond donating functional groups are present. Yet, the fatty acyl carbonyl group resulting from the amide linkage with the sphingoid base amino group (at carbon two) provides hydrogen bond accepting capacity. Hence, both hydrogen bond donating and accepting functional groups occur in the interfacial region (e.g. Boggs, 1987; Pink et al., 1988). The situation markedly contrasts that of membrane phosphoglycerides which have two hydrogen bond accepting carbonyl groups in the same region.
Finally, the polar headgroup regions of both sphingolipids and phosphoglycerides provide additional diversity. In mammalian cells, the major nonglycosylated sphingolipid is sphingomyelin, in which phosphorylcholine headgroup is linked via a phosphate ester to the 1-hydroxyl group of ceramide. This headgroup is charged but zwitterionic at neutral pH due to the presence of both the phosphate and (trimethylammonio)ethyl groups and is identical to that of glycerophosphocholines. When one or more sugar residues is linked by a β-glycosidic bond to the 1-hydroxyl group of ceramide, a glycosphingolipid results. Tremendous diversity exists among the sugar headgroups (Stults et al., 1989). A glycosphingolipid becomes a ganglioside if one or more of its sugar residues is a sialic acid, which is negatively charged at physiological conditions. The hydroxyl groups of the sugar residues provide the possibility for additional intra- and/or intermolecular hydrogen bonding which can have dramatic effects on the sugar headgroup conformation (e.g. Nyholm and Pascher, 1993). In contrast, membrane phosphoglycerides achieve diversity by utilizing phosphorylcholine, phosphorylethanolamine, phosphorylserine, phosphorylinositol, or phosphorylglycerol as headgroups. Considering the diversity of structure that exists among phosphoglycerides and sphingolipids, it is readily apparent that nature has provided inherent flexibility into the potential ways that these lipids can organize with each other and with cholesterol in membranes.
CHOLESTEROL
While the basic bilayer structure of all eukaryotic membranes occurs due to the strong amphipathic nature of the phosphoglyceride and sphingolipid components, sterols also are essential components of eukaryotic membranes. In mammalian membranes, the sterol is always cholesterol. Mammalian cells do not grow without an endogenous or exogenous source of cholesterol. This sterol is not uniformly distributed among mammalian intracellular membranes but is concentrated largely to the plasma membrane (Lange et al., 1989). Lesser amounts occur in the Golgi, nuclear, and lysosomal membranes as well as in the endoplasmic reticulum and mitochondria.
The basic structural motif of cholesterol differs greatly from that of sphingolipids and phosphoglycerides. Cholesterol (cholest-5-en-3β-ol) contains a tetracyclic hydrocarbon ring system that is similar to perhydrogenated cyclopentanophenanthrene (Fig. 2), but is actually an unsaturated alcohol due to its 3-hydroxyl group and 5,6 double bond. Attached to the cyclopentanyl ring at carbon 17 is an isooctyl hydrocarbon side chain. The rings of cholesterol are fused in the trans-configuration, which results in a compact and relatively planar conformational arrangement. Also, the 3-OH group, the two methyl groups at the junction of the rings and the side chain are all located on the same side of the ring (β-configuration). Variations of these arrangements occur in related steroids.
The only polar functional group is the hydroxyl residue. Hence, cholesterol orients in bilayers with its 3-OH group positioned towards the aqueous interface in the vicinity of the ester bonds of phosphoglycerides. Hydrogen bonding of the 3-OH group directly to the oxygen atom at either the sn-1 or sn-2 position of phosphoglycerides rather than to interfacial water appears to be unlikely (Bittman, 1997). In contrast, hydrogen bonding of the 3-OH group to the amide group of sphingomyelin appears to be quite likely (Sankaram and Thompson, 1990; Bittman et al., 1994).
Cholesterol’s nonpolar tetracyclic planar ring system resides adjacent to hydrocarbon segments of sphingolipids and phosphoglycerides that are closer to the aqueous interface than the bilayer midplane (e.g. McIntosh et al., 1992; Morrow et al., 1995; Ruocco et al., 1996). This orientation and conformation accounts for cholesterol being able to order the adjacent hydrocarbon chains by diminishing trans-gauche isomerization about the carbon-carbon bonds. The hydrocarbon region affected by the rigid sterol ring generally lies from the second carbon to about the seventh to tenth carbon of the chain (Davies et al., 1990). The net effect is to reduce the transient ‘kinks’ in the chains and diminish ion leakage across fluid bilayer membranes. NMR data indicate the effective length of cholesterol (including its side-chain) within phospholipid bilayers is about 17.5 Å, a length that corresponds to a 17-carbon all-trans hydrocarbon chain (Wu and Chi, 1991). Yet, the penetration depth of cholesterol within PC bilayers varies in response to changes in sterol concentration depending upon the acyl chain length of the adjacent PC. As a result, the sterol side chain sometimes can partially penetrate into the opposing bilayer leaflet (e.g. Sankaram and Thompson, 1991; Harris et al., 1995). Also, sterol side-chain structure impacts significantly on interactions with phosphoglycerides (Bittman, 1997).
IMPACT OF CHOLESTEROL ON THE PHYSICAL PROPERTIES OF LIPID BILAYERS
Sphingolipids generally occur as minor lipid constituents of most cells making their isolation more difficult and labor intensive than that of phosphoglycerides. As a result, most physical studies have focused on the behavior of phosphoglycerides rather than sphingolipids. Moreover, because of the emphasis on phosphoglyceride behavior in membrane biophysics, production and use of sphingolipids with homogeneous acyl compositions has lagged behind that of phosphoglyceride molecular species. Hence, physical studies on sphingolipids and their mixing behavior with cholesterol have been relatively uncommon (e.g. Maggio, 1994). Instead, the bulk of the physical studies involving cholesterol have been carried out in model bilayer membranes composed of glycerophosphocholine (PC). Of the studies involving PC, the vast majority have focused on species in which both acyl chains are saturated (e.g. DMPC = di-14:0 PC, DPPC = di-16:0 PC) despite the fact that PCs having saturated sn-1 and unsaturated sn-2 acyl chains are more prevalent physiologically. This situation, which has begun to be remedied over the past decade, also can be traced to the historic difficulty in obtaining and working with pure PC species with the desired physiological acyl motif. As a result, a strange situation has developed in that the most-often studied PCs actually have chain structural motifs that mimic natural sphingolipids in certain aspects rather than naturally predominant PC species. This fact must be kept in mind when drawing conclusions from many earlier studies involving PC and sterol interactions.
The impact of cholesterol on phosphoglyceride bilayers has been reviewed recently in clear and succinct fashion (Welti and Glaser, 1994; Keough et al., 1996; Bittman, 1997). Rather than being redundant, the focus here will be primarily on the physical properties of sphingolipid and cholesterol mixtures likely to be important in understanding the recent cell biological data regarding the lipid composition of detergent-insoluble glycolipid rafts. For instance, the in-plane elasticity of simple sphingolipids is somewhat lower than that of phosphatidylcholines (Smaby et al., 1996a). Adding cholesterol increases the lipid packing densities of sphingolipids to levels that equal or exceed those of PCs (Smaby et al., 1996b). The increased lipid packing densities induced by cholesterol markedly affect their in-plane elasticities. For instance, adding 50 mol% cholesterol to di-saturated PC drastically reduces the in-plane elasticity (5- to 7-fold) (Needham et al., 1988; Smaby et al., 1997) to levels roughly comparable to those achieved when cholesterol is mixed with simple sphingolipids in bilayers or in monolayers (Needham and Nunn, 1990; McIntosh et al., 1992; Smaby, J. M. and Brown, R. E., unpublished data). In contrast, unsaturation in one or both of the PC acyl chains inhibits cholesterol’s ability to lower the in-plane elasticity in bilayers (Needham and Nunn, 1990) and in monolayers (Smaby et al., 1997). Interestingly, compared to cholesterol and di-18:1 PC mixtures, cholesterol and di-16:0 PC mixtures mimic more closely the low fluidity levels observed in detergent-insoluble glycolipid rafts (Schroeder et al., 1994). The preceding observations may explain the differential susceptibility of various membrane regions to solubilization by Triton X-100. Detergent partitioning at 4°C into putative sphingolipidcholesterol enriched microdomains is likely to be dramatically slowed by the tight and less elastic packing state of these membrane regions.
Regarding the impact of cholesterol on lipid domain organization and domain physical features, a few recent studies are particularly pertinent. 2H-NMR studies of galactosylceramide species mixed with cholesterol and with cholesterol/PC bilayers show changes in rotational mobilizations that could be due to phase separation (Hamilton et al., 1994). Subsequent NMR studies of globoside indicate that the surrounding lipid environment (including cholesterol) appear to affect the spacing and orientation of the glycolipids within the bilayer matrix (Jones et al., 1997). Other recent fluorescence and calorimetric investigations provide intriguing insights into how cholesterol might organize in sphingomyelin bilayers (Chong et al., 1996) as well as how cholesterol affects the domain structure in mixtures of sphingomyelin and ganglioside GM1 (Ferraretto et al., 1997). Also pertinent are the studies of how cholesterol concentration impacts on the mixing behavior and domain organization of surrounding matrix lipids. At 50 mol%, cholesterol promotes the intermixing of PC species (monounsaturated and disaturated) at temperatures where the two phosphoglycerides exhibit extensive (solid/fluid) lateral phase separation when the cholesterol is absent (Silvius, 1992). Interestingly, however, high cholesterol levels were not able to induce complete miscibility of unsaturated PCs with galactosylceramides. Subsequently, Silvius et al. (1996) showed that cholesterol abolishes the lateral segregation of PCs with different saturated acyl chain lengths at high concentrations, but promotes their lateral segregation at lower concentrations. Such effects extend to sphingolipids as well (Ahmed et al., 1997). Including 33 mol% cholesterol in model membranes of sphingomyelin or dipalmitoyl phosphatidylcholine greatly promotes phase separation. The nature of the separated phases indicates a coexistence of both liquid-disordered and liquid-ordered phases. The amount of liquid-ordered phase, as detected by fluorescence quenching, correlates with the detergent insolubility of the cholesterol-containing model membranes (Ahmed et al., 1997).
PUTATIVE ROLE OF CHOLESTEROL IN RAFT FUNCTION
Considering the structural features of naturally predominate sphingolipids and phosphoglycerides and the resulting consequences on their physical behavior, it is apparent that sphingolipids are likely candidates for participating in lipid microdomain formation in eukaryotic biomembranes. Cholesterol does not appear to be essential for sphingolipid microdomain formation. Yet, in the absence of cholesterol, the physical nature of the sphingolipid domains would be very different from when cholesterol is present. Without cholesterol, the sphingolipid hydrocarbon chains would be rigid due to their gel and/or lamellar crystalline character (Koynova and Caffrey, 1995). Individual molecules would undergo slower rotational diffusion and would be relatively confined within the surrounding domain lattice. In contrast, the liquid-ordered state created by high cholesterol concentrations (>25 mol%) would provide a domain environment that is tightly packed and of low in-plane elasticity (equaling or exceeding that of the sphingolipid gel state) but that maintains a relatively high degree of rotational diffusion within a less restrictive lattice environment. Such an environment could be regulated to facilitate the diffusion of GPI-anchored or other proteins into or out of such domains. The saturated acyl chains characteristic of GPI-anchors would find the cholesterol-sphingolipid liquid ordered state well-suited for dynamic association with such domains (Schroeder et al., 1994; Smaby et al., 1996b).
The formation of sphingolipid-cholesterol microdomains undoubtedly involves strong van der Waals stabilization contributed by the planar steroid ring interacting with the saturated hydrocarbon chains of the sphingolipids as well as the hydrogen bonding likely to occur between the 3-OH group of cholesterol and the amide bond of sphingolipids. The relatively ordered chain conformations of sphingolipids will be maintained in the presence of cholesterol and lead to chain-length mismatch with surrounding thinner regions that are enriched in saturated sn-1, unsaturated sn-2 phosphoglycerides. Despite interacting favorably with available sphingolipids, the shear mass of cholesterol in plasma membranes precludes this sterol from localizing exclusively to sphingolipid-cholesterol rafts. Cholesterol also will mix with the fluid-phase phosphoglycerides. Yet, the extent of mixing as well as the nature of the resulting packing state will depend upon various factors. For instance, the presence of cis unsaturation in the acyl chains will introduce kinks that will limit cholesterol’s ability to order, to mix homogeneously, and to reduce interfacial elasticity (e.g. Keough et al., 1996; Litman and Drake, 1996; Smaby et al., 1997). Also, headgroup composition will strongly affect the mixing behavior. Hence, at least two pools of cholesterol are likely to exist in cell membranes, one that is raft-associated and another that localizes to fluid phosphoglyceride regions (Harder and Simons, 1997; Simons and Ikonen, 1997).
Finally, small amounts of cholesterol may have a tendency to localize along the boundaries between sphingolipid-enriched domains and fluid-phase phosphoglyceride (e.g. Cruzeiro-Hansson et al., 1989) in an effort to minimize the unfavorable energetic effects created along the junctures. In doing so, cholesterol will alter the line tension at the boundaries along the mismatch region leading to changes in microdomain size, shape, and physical characteristics. Hence, the role of cholesterol is likely to be quite a bit more involved than functioning simply as a spacer lipid with the capacity to fill voids that form among nearby sphingolipids.
Acknowledgments
The support of USPHS grant GM45928 and the Hormel Foundation is gratefully acknowledged. I also thank the two anonymous reviewers for pointing out relevant articles that were in press while this Commentary was being assembled.
REFERENCES
- Ahmed SN, Brown DA, London E. On the origin of sphingolipid/cholesterol-rich detergent-insoluble cell membranes: physiologic concentrations of cholesterol and sphingolipid induce formation of a detergent-insoluble, liquid-ordered lipid phase in model membranes. Biochemistry. 1997;36:10944–10953. doi: 10.1021/bi971167g. [DOI] [PubMed] [Google Scholar]
- Almeida PF, Vaz WLC, Thompson TE. Lateral diffusion and percolation in two-phase, two-component lipid bilayers. Topology of the solid-phase domains in-plane and across the lipid bilayer. Biochemistry. 1992;31:7198–7210. doi: 10.1021/bi00146a024. [DOI] [PubMed] [Google Scholar]
- Almeida PF, Vaz WLC, Thompson TE. Percolation and diffusion in three-component lipid bilayers: effect of cholesterol on an equimolar mixture of two phosphatidylcholines. Biophys. J. 1993;64:399–412. doi: 10.1016/S0006-3495(93)81381-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson RGW. Caveolae, where incoming and outgoing messengers meet. Proc. Nat. Acad. Sci. USA. 1993;90:10909–10913. doi: 10.1073/pnas.90.23.10909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bach D, Miller IR, Sela BA. Calorimetric studies on various gangliosides and ganglioside-lipid interactions. Biochim. Biophys. Acta. 1982;686:233–239. doi: 10.1016/0005-2736(82)90117-1. [DOI] [PubMed] [Google Scholar]
- Barenholz Y, Thompson TE. Sphingomyelins in bilayers and biological membranes. Biochim. Biophys. Acta. 1980;604:129–158. doi: 10.1016/0005-2736(80)90572-6. [DOI] [PubMed] [Google Scholar]
- Bittman R. Has nature designed the cholesterol side chain for optimal interaction with phospholipids? In: Bittman R, editor. Subcellular Biochemistry, Volume 28: Cholesterol: Its Functions and Metabolism in Biology and Medicine. Plenum Press; New York: 1997. pp. 145–171. [DOI] [PubMed] [Google Scholar]
- Bittman R, Kasireddy CR, Mattjus P, Slotte JP. Interaction of cholesterol with sphingomyelin monolayers and vesicles. Biochemistry. 1994;33:11776–11781. doi: 10.1021/bi00205a013. [DOI] [PubMed] [Google Scholar]
- Boggs JM. Lipid intermolecular hydrogen bonding: influence on structural organization and membrane function. Biochim. Biophys. Acta. 1987;906:353–404. doi: 10.1016/0304-4157(87)90017-7. [DOI] [PubMed] [Google Scholar]
- Boggs JM, Koshy KM. Do the long fatty acid chains of sphingolipids interdigitate across the center of a bilayer of shorter chain symmetric phospholipids? Biochim. Biophys. Acta. 1994;1189:233–241. doi: 10.1016/0005-2736(94)90070-1. [DOI] [PubMed] [Google Scholar]
- Bretscher MS, Munro S. Cholesterol and the Golgi apparatus. Science. 1993;261:1280–1281. doi: 10.1126/science.8362242. [DOI] [PubMed] [Google Scholar]
- Brown DA, Rose JK. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. doi: 10.1016/0092-8674(92)90189-j. [DOI] [PubMed] [Google Scholar]
- Brown RE, Allietta M, Tillack TW, Thompson TE. Glycolipids are not extracted from phospholipids bilayers by binding to ferritin-lectin conjugates. Biochim. Biophys. Acta. 1983;731:495–499. [Google Scholar]
- Brown RE, Sugàr IP, Thompson TE. Spontaneous transfer of gangliotetraosylceramide between phospholipid vesicles. Biochemistry. 1985;24:4082–4091. doi: 10.1021/bi00336a042. [DOI] [PubMed] [Google Scholar]
- Brown RE, Thompson TE. Spontaneous transfer of ganglioside GM1 between phospholipid vesicles. Biochemistry. 1987;26:5454–5460. doi: 10.1021/bi00391a036. [DOI] [PubMed] [Google Scholar]
- Chong PL-G, Liu F, Wang MM, Truong K, Sugar IP, Brown RE. Fluorescence evidence for cholesterol regular distribution in phosphatidylcholine and in sphingomyelin lipid bilayers. J. Fluorescence. 1996;6:221–230. doi: 10.1007/BF00732825. [DOI] [PubMed] [Google Scholar]
- Correa-Freire MC, Freire E, Barenholz Y, Biltonen RL, Thompson TE. Thermotropic behavior of monoglucocerebrosidedipalmitoylphosphatidylcholine multilamellar liposomes. Biochemistry. 1979;18:442–445. doi: 10.1021/bi00570a008. [DOI] [PubMed] [Google Scholar]
- Correa-Freire MC, Barenholz Y, Thompson TE. Glucocerebroside transfer between phosphatidylcholine bilayers. Biochemistry. 1982;21:1244–1248. doi: 10.1021/bi00535a021. [DOI] [PubMed] [Google Scholar]
- Cruzeiro-Hansson L, Ipsen JH, Mouritsen OG. Intrinsic molecules in lipid membranes change the lipid-domain interfacial area: cholesterol at domain interfaces. Biochim. Biophys. Acta. 1989;979:166–176. doi: 10.1016/0005-2736(89)90432-x. [DOI] [PubMed] [Google Scholar]
- Curatolo W. The physical properties of glycolipids. Biochim. Biophys. Acta. 1987;906:111–136. doi: 10.1016/0304-4157(87)90008-6. [DOI] [PubMed] [Google Scholar]
- Davies MA, Schuster HF, Brauner JW, Mendelsohn R. Effects of cholesterol on conformational disorder in dipalmitoylphosphatidylcholine bilayers. A quantitative IR study of the depth dependence. Biochemistry. 1990;29:4368–4373. doi: 10.1021/bi00470a016. [DOI] [PubMed] [Google Scholar]
- Dupree P, Parton RG, Raposo G, Kurzchalia TV, Simons K. Caveolae and sorting in the trans-Golgi-network of epithelial cells. EMBO J. 1993;12:1597–1605. doi: 10.1002/j.1460-2075.1993.tb05804.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dwyer JD, Bloomfield VA. Cholera toxin mediated agglutination of ganglioside GM1 containing phospholipid vesicles and GM1-coated polystyrene spheres. Biochemistry. 1982;21:3231–3234. doi: 10.1021/bi00256a031. [DOI] [PubMed] [Google Scholar]
- Ferraretto A, Pitto M, Palestini P, Masserini M. Lipid domains in the membrane: Thermotropic properties of sphingomyelin vesicles containing GM1 ganglioside and cholesterol. Biochemistry. 1997;36:9232–9236. doi: 10.1021/bi970428j. [DOI] [PubMed] [Google Scholar]
- Fiedler K, Kobayashi T, Kurzchalia TV, Simons K. Glycosphingolipids-enriched, detergent-insoluble complexes in protein sorting in epithelial cells. Biochemistry. 1993;32:6365–6373. doi: 10.1021/bi00076a009. [DOI] [PubMed] [Google Scholar]
- Finegold L, editor. Cholesterol in Membrane Models. CRC Press; Boca Raton, FL: 1993. [Google Scholar]
- Fra AM, Masserini M, Palestini P, Sonnino S, Simons K. A photo-reactive derivative of ganglioside GM1 specifically cross-links VIP21-caveolin on the cell surface. FEBS Lett. 1995;375:11–14. doi: 10.1016/0014-5793(95)95228-o. [DOI] [PubMed] [Google Scholar]
- Frank A, Barenholz Y, Lichtenberg D, Thompson TE. Spontaneous transfer of sphingomyelin between phospholipid bilayers. Biochemistry. 1983;22:5647–5651. [Google Scholar]
- Fujimoto T. GPI-anchored proteins, glycosphingolipids, and sphingomyelin are sequestered to caveolae only after crosslinking. J. Histochem. Cytochem. 1996;44:929–941. doi: 10.1177/44.8.8756764. [DOI] [PubMed] [Google Scholar]
- Futerman AH. Inhibition of sphingolipid synthesis: effects on glycosphingolipid-GPI-anchored protein microdomains. Trends Cell Biol. 1995;5:377–379. doi: 10.1016/s0962-8924(00)89078-9. [DOI] [PubMed] [Google Scholar]
- Gorodinsky A, Harris DA. Glycolipid-anchored proteins in neuroblastoma cells from detergent-resistant complexes without caveolin. J. Cell Biol. 1995;129:619–627. doi: 10.1083/jcb.129.3.619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant CWM, Peters MW. Lectin-membrane interactions. Biochim. Biophys. Acta. 1984;799:403–422. doi: 10.1016/0304-4157(84)90018-2. [DOI] [PubMed] [Google Scholar]
- Hamilton KS, Jarrell HC, Briere KM, Grant CWM. Glycosphingolipid backbone conformation and behavior in cholesterol-containing phospholipids bilayers. Biochemistry. 1993;32:4022–4028. doi: 10.1021/bi00066a024. [DOI] [PubMed] [Google Scholar]
- Hamilton KS, Briere KM, Jarrell HC, Grant CWM. Acyl chain length effects related to glycosphingolipid crypticity in phospholipid membranes: probed by 2H-NMR. Biochim. Biophys. Acta. 1994;1190:367–375. doi: 10.1016/0005-2736(94)90096-5. [DOI] [PubMed] [Google Scholar]
- Harder T, Simons K. Calveolae, DIGs, and the dynamics of sphingolipid-cholesterol microdomains. Curr. Opin. Cell Biol. 1997;9:534–542. doi: 10.1016/s0955-0674(97)80030-0. [DOI] [PubMed] [Google Scholar]
- Harris JS, Epps DE, Davio SR, Kezdy FJ. Evidence for transbilayer tail-to-tail cholesterol dimers in dipalmitoylglycerophosphocholine liposomes. Biochemistry. 1995;34:3851–3857. doi: 10.1021/bi00011a043. [DOI] [PubMed] [Google Scholar]
- Holopainen JM, Lehtonen JYA, Kinnunen PKJ. Lipid microdomains in dimyristoylphosphatidylcholine-ceramide liposomes. Chem. Phys. Lipids. 1997;88:1–13. doi: 10.1016/s0009-3084(97)00040-6. [DOI] [PubMed] [Google Scholar]
- Hori T, Sugita M. Sphingolipids in lower animals. Prog. Lipid Res. 1993;32:25–45. doi: 10.1016/0163-7827(93)90003-f. [DOI] [PubMed] [Google Scholar]
- Jacobson K, Sheets ED, Simson R. Revisiting the fluid mosaic model of membranes. Science. 1995;268:1441–1442. doi: 10.1126/science.7770769. [DOI] [PubMed] [Google Scholar]
- Johnson SB, Brown RE. Simplified derivatization for determining sphingolipid fatty acyl composition by gas chromatography-mass spectrometry. J. Chromatogr. 1992;605:281–286. doi: 10.1016/0021-9673(92)85248-r. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones DH, Lingwood CA, Barber KR, Grant CWM. Globoside as a membrane receptor: A consideration of oligosaccharide communication with the hydrophobic domain. Biochemistry. 1997;36:8539–8547. doi: 10.1021/bi970183a. [DOI] [PubMed] [Google Scholar]
- Karlsson K-A. Sphingolipid long chain bases. Lipids. 1970;5:878–891. doi: 10.1007/BF02531119. [DOI] [PubMed] [Google Scholar]
- Keough KMW, Morrow MR, Bittman R. Phospholipidcholesterol bilayers. In: Grant DM, Harris RK, editors. Encyclopedia of Nuclear Magnetic Resonance. John Wiley; Chichester, New York: 1996. pp. 3601–3608. [Google Scholar]
- Koynova R, Caffrey M. Phases and phase transitions of the sphingolipids. Biochim. Biophys. Acta. 1995;1255:213–236. doi: 10.1016/0005-2760(94)00202-a. [DOI] [PubMed] [Google Scholar]
- Kuksis A, Myher JJ. Analyses of subsets of molecular species of glycerophospholipids. In: Paoletti R, et al., editors. Lipids. Vol. 1: Biochemistry. Raven Press; New York: 1976. pp. 23–38. [Google Scholar]
- Kulkarni VS, Brown RE. Thermotropic behavior of galactosylceramides with cis-monoenoic fatty acyl chains. Biophys. J. 1997 doi: 10.1016/s0005-2736(98)00076-5. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurzchalia TV, Hartmann E, Dupree P. Guilt by insolubility – does a protein’s detergent insolubility reflect a caveolar location? Trends Cell Biol. 1995;4:187–189. doi: 10.1016/s0962-8924(00)88990-4. [DOI] [PubMed] [Google Scholar]
- Lange Y, Swaisgood MH, Ramos BV, Steck TL. Plasma membranes contain half the phospholipid and 90% of the cholesterol and sphingomyelin in cultured human fibroblasts. J. Biol. Chem. 1989;264:3786–3793. [PubMed] [Google Scholar]
- Lee PM, Grant CWM. Ganglioside headgroup disorder as a sequel to lectin binding. Biochem. Biophys. Res. Commun. 1980;95:1299–1305. doi: 10.1016/0006-291x(80)91615-0. [DOI] [PubMed] [Google Scholar]
- Lester RL, Dickson RC. Sphingolipids with inositolphosphate-containing head groups. Advan. Lipid Res. 1993;26:253–274. [PubMed] [Google Scholar]
- Lisanti MP, Scherer PE, Tang Z-L, Sargiacomo M. Caveolae, caveolin and caveolin-rich membrane domains: a signaling hypothesis. Trends Cell Biol. 1994;4:231–235. doi: 10.1016/0962-8924(94)90114-7. [DOI] [PubMed] [Google Scholar]
- Litman BJ, Mitchell DC. A role for phospholipid polyunsaturation in modulating membrane protein function. Lipids. 1996;31:s-193–s-197. doi: 10.1007/BF02637075. [DOI] [PubMed] [Google Scholar]
- Liu P, Anderson RGW. Compartmentalized production of ceramide at the cell surface. J. Biol. Chem. 1995;270:271–27185. doi: 10.1074/jbc.270.45.27179. [DOI] [PubMed] [Google Scholar]
- Liu J, Oh P, Horner T, Rogers RA, Schnitzer JE. Organized endothelial cell surface signal transduction in caveolae distinct from glycosylphosphatidylinositol-anchored protein microdomains. J. Biol. Chem. 1997;272:7211–7222. doi: 10.1074/jbc.272.11.7211. [DOI] [PubMed] [Google Scholar]
- Luan P, Yang L, Glaser M. Formation of membrane domains created during the budding of vesicular stomatitis virus. A model for selective and protein sorting in biological membranes. Biochemistry. 1995;34:9874–9883. doi: 10.1021/bi00031a008. [DOI] [PubMed] [Google Scholar]
- Maggio B, Ariga T, Sturtevant JM, Yu RK. Thermotropic behavior of binary mixtures of dipalmitoylphosphatidylcholine and glycosphingolipids in aqueous dispersions. Biochim. Biophys. Acta. 1985;818:1–12. doi: 10.1016/0005-2736(85)90131-2. [DOI] [PubMed] [Google Scholar]
- Maggio B. The surface behavior of glycosphingolipids in biomembranes: a new frontier of molecular ecology. Prog. Biophys. Mol. Biol. 1994;62:55–117. doi: 10.1016/0079-6107(94)90006-x. [DOI] [PubMed] [Google Scholar]
- Mattjus P, Bittman R, Vilcheze C, Slotte JP. Lateral domain formation in cholesterol/-phospholipid monolayers as affected by the sterol side chain conformation. Biochim. Biophys. Acta. 1995;1240:237–247. doi: 10.1016/0005-2736(95)00179-4. [DOI] [PubMed] [Google Scholar]
- Maulik PR, Shipley GG. Interactions of N-stearoyl sphingomyelin with cholesterol and dipalmitoylphosphatidycholine in bilayer membranes. Biophys. J. 1996a;70:2256–2265. doi: 10.1016/S0006-3495(96)79791-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maulik PR, Shipley GG. N-palmitoyl sphingomyelin bilayers: structure and interactions with cholesterol and dipalmitoylphosphatidylcholine. Biochemistry. 1996b;35:8025–8034. doi: 10.1021/bi9528356. [DOI] [PubMed] [Google Scholar]
- Mayor S, Rothberg KG, Maxfield FR. Sequestration of GPI-anchored proteins in caveolae triggered by cross-linking. Science. 1994;264:1948–1951. doi: 10.1126/science.7516582. [DOI] [PubMed] [Google Scholar]
- Mayor S, Maxfield FR. Insolubility and redistribution of GPI-anchored proteins at the cell surface after detergent treatment. Mol. Biol. Cell. 1995;6:929–944. doi: 10.1091/mbc.6.7.929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McIntosh TJ, Simon SA, Needham D, Huang C. Structure and cohesive properties of sphingomyelin/cholesterol bilayers. Biochemistry. 1992;31:2012–2020. doi: 10.1021/bi00122a017. [DOI] [PubMed] [Google Scholar]
- Mehlhorn IE, Parraga G, Barber KR, Grant CWM. Visualization of domains in rigid ganglioside/phosphatidylcholine bilayers: Ca2+ effects. Biochim. Biophys. Acta. 1986;863:139–155. doi: 10.1016/0005-2736(86)90254-3. [DOI] [PubMed] [Google Scholar]
- Molotkovsky JG, Mikhalyov II, Imbs AB, Bergelson LD. Synthesis and characterization of new fluorescent glycolipid probes: Molecular organization of glycosphingolipids in mixed-composition lipid bilayers. Chem. Phys. Lipids. 1991;58:199–212. [Google Scholar]
- Morrow MR, Singh D, Lu D, Grant CWM. Glycosphingolipid fatty acid arrangement in phospholipid bilayers: cholesterol effects. Biophys. J. 1995;68:179–186. doi: 10.1016/S0006-3495(95)80173-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Needham D, McIntosh TJ, Evans E. Thermomechanical and transition properties of dimyristoylphosphatidylcholine/cholesterol bilayers. Biochemistry. 1988;27:4668–4673. doi: 10.1021/bi00413a013. [DOI] [PubMed] [Google Scholar]
- Needham D, Nunn RS. Elastic deformation and failure of lipid bilayer membranes containing cholesterol. Biophys. J. 1990;58:997–1009. doi: 10.1016/S0006-3495(90)82444-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neuringer LJ, Sears B, Jungalwala FB. Deuterium NMR studies of cerebroside-phospholipid bilayers. Biochim. Biophys. Acta. 1979;558:325–329. doi: 10.1016/0005-2736(79)90268-2. [DOI] [PubMed] [Google Scholar]
- Nyholm P-G, Pascher I. Orientation of the saccharide chains of glycolipids at the membrane surface: conformational analysis of the glucoseceramide and the glucose-glyceride linkages using molecular mechanics (MM3) Biochemistry. 1993;32:1225–1234. doi: 10.1021/bi00056a005. [DOI] [PubMed] [Google Scholar]
- Palestini P, Allietta M, Sonnino S, Tettamanti G, Thompson TE, Tillack TW. Gel phase preference of ganglioside GM1 at low concentrations in two-component, two-phase phosphatidylcholine bilayers depends upon the ceramide moiety. Biochim. Biophys. Acta. 1995;1235:221–230. doi: 10.1016/0005-2736(95)80008-4. [DOI] [PubMed] [Google Scholar]
- Parton RG. Ultrastructural localization of gangliosides: GM1 is concentrated in caveolae. J. Histochem. Cytochem. 1994;42:155–166. doi: 10.1177/42.2.8288861. [DOI] [PubMed] [Google Scholar]
- Parton RG, Simons K. Digging into caveolae. Science. 1995;269:1398–1399. doi: 10.1126/science.7660120. [DOI] [PubMed] [Google Scholar]
- Parton RG. Caveolae and caveolins. Curr. Opin. Cell Biol. 1996;8:542–548. doi: 10.1016/s0955-0674(96)80033-0. [DOI] [PubMed] [Google Scholar]
- Peters MW, Grant CWM. Freeze-etch study of an unmodified lectin interacting with its receptors in model membranes. Biochim. Biophys. Acta. 1984;775:273–282. doi: 10.1016/0005-2736(84)90181-0. [DOI] [PubMed] [Google Scholar]
- Peters MW, Barber KR, Grant CWM. Lateral distribution of gangliosides in bilayer membranes: Lipid and ionic effects. J. Neurosci. Res. 1984;12:343–353. doi: 10.1002/jnr.490120221. [DOI] [PubMed] [Google Scholar]
- Pink DA, MacDonald AL, Quinn B. Anisotropic interactions in hydrated cerebrosides. A theoretical model of stable and metastable states and hydrogen bond formation. Chem. Phys. Lipids. 1988;47:83–95. [Google Scholar]
- Robinson BS, Johnson DW, Poulos A. Novel molecular species of sphingomyelin containing 2-hydroxylated polyenoic very long chain fatty acids in mammalian testes and spermatozoa. J. Biol. Chem. 1992;267:1746–1751. [PubMed] [Google Scholar]
- Rock P, Allietta M, Young WW, Jr, Thompson TE, Tillack TW. Organization of glycosphingolipids in phosphatidylcholine bilayers: use of antibody molecules and Fab fragments as morphologic markers. Biochemistry. 1990;29:8484–8490. doi: 10.1021/bi00488a040. [DOI] [PubMed] [Google Scholar]
- Rock P, Allietta M, Young WW, Thompson TE, Tillack TW. Ganglioside GM1 and asialo-GM1 at low concentration are preferentially incorporated into the gel phase in two-component, two-phase phosphatidylcholine bilayers. Biochemistry. 1991;30:19–25. doi: 10.1021/bi00215a003. [DOI] [PubMed] [Google Scholar]
- Ruocco MJ, Shipley GG, Oldfield E. Galactocerebrosidephospholipid interactions in bilayer membranes. Biophys. J. 1983;43:91–101. doi: 10.1016/S0006-3495(83)84327-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruocco MJ, Siminovitch DJ, Long JR, Das Gupta SK, Griffin RG. 2H and 13C nuclear magnetic resonance study of N-palmitoylgalactosylsphingosine (cerebroside)/cholesterol bilayers. Biophys. J. 1996;71:1776–1788. doi: 10.1016/S0006-3495(96)79378-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sankaram MB, Thompson TE. Interactions of cholesterol with various glycerophospholipids and sphingomyelin. Biochemistry. 1990;29:10670–10675. doi: 10.1021/bi00499a014. [DOI] [PubMed] [Google Scholar]
- Sankaram MB, Thompson TE. Cholesterol-induced fluid-phase immiscibility in membranes. Proc. Nat. Acad. Sci. USA. 1991;88:8686–8690. doi: 10.1073/pnas.88.19.8686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder F, Jefferson JR, Kier AB, Knittel J, Scallen TJ, Wood WG, Hapala I. Membrane cholesterol dynamics: Cholesterol domains and kinetic pools. Proc. Soc. Exp. Biol. Med. 1991;196:235–252. doi: 10.3181/00379727-196-43185. [DOI] [PubMed] [Google Scholar]
- Schroeder R, London E, Brown D. Interactions between saturated acyl chains confer detergent resistance on lipids and glycosylphosphatidylinositol (GPI)-anchored proteins: GPI-anchored proteins in liposomes and cells show similar behavior. Proc. Nat. Acad. Sci. USA. 1994;91:12130–12134. doi: 10.1073/pnas.91.25.12130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schroeder F, Frolov AA, Murphy EJ, Atshaves BP, Jefferson JR, Pu L, Wood WG, Foxworth WB, Kier AB. Recent advances in membrane cholesterol domain dynamics and intracellular cholesterol trafficking. Proc. Soc. Exp. Biol. Med. 1996;213:150–177. doi: 10.3181/00379727-213-44047. [DOI] [PubMed] [Google Scholar]
- Schnitzer JE, McIntosh DP, Dvorak AM, Liu J, Oh P. Separation of caveolae from associated microdomains of GPI-anchored proteins. Science. 1995;269:1435–1439. doi: 10.1126/science.7660128. [DOI] [PubMed] [Google Scholar]
- Silvius JR. Cholesterol modulation of lipid intermixing in phospholipid and glycosphingolipid mixtures. Evaluation using fluorescent lipid probes and brominated lipid quenchers. Biochemistry. 1992;31:3398–3408. doi: 10.1021/bi00128a014. [DOI] [PubMed] [Google Scholar]
- Silvius JR, del Guidice D, Lafleur M. Cholesterol at different bilayer concentrations can promote or antagonize lateral segregation of phospholipids of differing acyl chain length. Biochemistry. 1996;35:15198–15208. doi: 10.1021/bi9615506. [DOI] [PubMed] [Google Scholar]
- Simons K, van Meer G. Lipid sorting in epithelial cells. Biochemistry. 1988;27:6197–6202. doi: 10.1021/bi00417a001. [DOI] [PubMed] [Google Scholar]
- Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387:569–572. doi: 10.1038/42408. [DOI] [PubMed] [Google Scholar]
- Singer SJ, Nicolson GL. The fluid mosaic model of the structure of cell membranes. Science. 1972;175:720–731. doi: 10.1126/science.175.4023.720. [DOI] [PubMed] [Google Scholar]
- Slater JL, Huang C. Lipid Bilayer Interdigitation. In: Yeagle P, editor. The Structure of Biological Membranes. CRC Press; Boca Raton, FL: 1992. pp. 175–210. [Google Scholar]
- Slotte JP, Pörn MI, Härmälä A-S. Flow and distribution of cholesterol – effects of phospholipids. Curr. Topics Membr. 1994;40:483–502. [Google Scholar]
- Slotte JP. Lateral domain formation in mixed monolayers containing cholesterol and dipalmitoylphosphatidylcholine or N-palmitoylsphingomyelin. Biochim. Biophys. Acta. 1995;1235:419–427. doi: 10.1016/0005-2736(95)80031-a. [DOI] [PubMed] [Google Scholar]
- Slotte JP. Cholesterol-sphingomyelin interactions in cells – effects on lipid metabolism. In: Bittman R, editor. Subcellular Biochemistry, Volume 28: Cholesterol: Its Functions and Metabolism in Biology and Medicine. Plenum Press; New York: 1997. pp. 277–293. [PubMed] [Google Scholar]
- Smaby JM, Kulkarni VS, Momsen MM, Brown RE. The interfacial elastic packing interactions of galactosylceramides, sphingomyelins, and phosphatidylcholines. Biophys. J. 1996a;70:868–877. doi: 10.1016/S0006-3495(96)79629-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smaby JM, Momsen MM, Kulkarni VS, Brown RE. Cholesterol-induced interfacial area condensations of galactosylceramides and sphingomyelins with identical acyl chains. Biochemistry. 1996b;35:5696–5704. doi: 10.1021/bi953057k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smaby JM, Momsen MM, Brockman HL, Brown RE. Phosphatidylcholine acyl unsaturation modulates the decrease in interfacial elasticity induced by cholesterol. Biophys. J. 1997;73:1492–1505. doi: 10.1016/S0006-3495(97)78181-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smart EJ, Ying YS, Mineo C, Anderson RGW. A detergent-free method for purifying caveolae membrane from tissue culture cells. Proc. Nat. Acad. Sci. USA. 1995;92:10104–10108. doi: 10.1073/pnas.92.22.10104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song KS, Li S, Okamoto T, Quilliam LA, Sargiacomo M, Lisanti MP. Co-purification and direct interaction of Ras with caveolin, an integral membrane protein of caveolae microdomains. Detergent-free purification of caveolae membranes. J. Biol. Chem. 1996;271:9690–9697. doi: 10.1074/jbc.271.16.9690. [DOI] [PubMed] [Google Scholar]
- Speyer JB, Weber RT, Das Gupta SK, Griffin RG. Anisotropic 2H NMR spin-lattice relaxation in Lα-phase cerebroside bilayers. Biochemistry. 1989;28:9569–9574. doi: 10.1021/bi00451a003. [DOI] [PubMed] [Google Scholar]
- Stan R-V, Roberts WG, Predescu D, Ihida K, Saucan L, Ghitescu L, Palade GE. Immunoisolation and partial characterization of endothelial plasmalemmal vesicles (caveolae) Mol. Biol. Cell. 1997;8:595–605. doi: 10.1091/mbc.8.4.595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stults CLM, Sweeley CC, Macher BA. Glycosphingolipids: structure, biological source, and properties. Meth. Enzymol. 1989;179:167–214. doi: 10.1016/0076-6879(89)79122-9. [DOI] [PubMed] [Google Scholar]
- Suzuki T, Utsumi H, Inoue K, Nojima S. Haptenic activity of galactosyl ceramide and its topographical distribution on liposomal membranes. I. Effect of cholesterol incorporation. Biochim. Biophys. Acta. 1981;644:183–191. doi: 10.1016/0005-2736(81)90374-6. [DOI] [PubMed] [Google Scholar]
- Thompson TE, Tillack TW. Organization of glycosphingolipids in bilayers and plasma membranes of mammalian cells. Annu. Rev. Biophys. Biophys. Chem. 1985;14:361–386. doi: 10.1146/annurev.bb.14.060185.002045. [DOI] [PubMed] [Google Scholar]
- Thompson TE, Allietta M, Brown RE, Johnson ML, Tillack TW. Organization of gangliosides GM1 in phosphatidylcholine bilayers. Biochim. Biophys. Acta. 1985;817:229–237. doi: 10.1016/0005-2736(85)90024-0. [DOI] [PubMed] [Google Scholar]
- Thompson TE, Barenholz Y, Brown RE, Correa-Freire M, Young WW, Tillack TW. Molecular organization of glycosphingolipids in phosphatidylcholine bilayers and biological membranes. In: Freysz L, Dreyfus H, Massarelli R, Gatt S, editors. Enzymes of Lipid Metabolism II. Plenum Pub. Corp.; New York: 1986. pp. 387–396. [Google Scholar]
- Thompson TE, Brown RE. Biophysical properties of gangliosides. In: Ledeen R, Hogan E, Tettamanti G, Yates A, Yu R, editors. New Trends in Ganglioside Research: Neurochemical and Neuroregenerative Aspects. Vol. 14. Liviana Press; Padova: 1988. pp. 65–78. (Fidia Res. Series). [Google Scholar]
- Tillack TW, Wong M, Allietta M, Thompson TE. Organization of the glycosphingolipid asialo-GM1 in phosphatidylcholine bilayers. Biochim. Biophys. Acta. 1982;691:261–273. doi: 10.1016/0005-2736(82)90415-1. [DOI] [PubMed] [Google Scholar]
- Tillack TW, Allietta M, Moran RE, Young WW. Localization of globoside and Forssman glycolipids on erythrocytes membranes. Biochim. Biophys. Acta. 1983;733:15–24. doi: 10.1016/0005-2736(83)90086-x. [DOI] [PubMed] [Google Scholar]
- van der Rest ME, Kamminga AH, Nakano A, Anraku Y, Poolman B, Konigs WN. The plasma membrane of Saccharomyces cerevisiae: structure, function, and biogenesis. Microbiol. Rev. 1995;59:304–322. doi: 10.1128/mr.59.2.304-322.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaz WL, Almeida PF. Phase topology and percolation in multiphase bilayers: Is the biological membrane a domain mosaic? Curr. Opin. Cell Biol. 1993;3:482–488. [Google Scholar]
- Weimbs T, Low SH, Chapin SJ, Mostov KE. Apical targeting in polarized epithelial cells: there’s more afloat than rafts. Trends Cell Biol. 1997;7:393–399. doi: 10.1016/S0962-8924(97)01130-6. [DOI] [PubMed] [Google Scholar]
- Welti R, Glaser M. Lipid domains in model and biological membranes. Chem. Phys. Lipids. 1994;73:121–137. doi: 10.1016/0009-3084(94)90178-3. [DOI] [PubMed] [Google Scholar]
- Wu W-G, Chi L-M. Conformational change of cholesterol side chain in lipid bilayers. J. Am. Chem. Soc. 1991;113:4683–4685. [Google Scholar]
- Yohe HC, Roark DE, Rosenberg A. C20-sphingosine as a determining factor in aggregation of gangliosides. J. Biol. Chem. 1976;251:7083–7087. [PubMed] [Google Scholar]
- Yu J, Fischmann DA, Steck TL. Selective solubilization of proteins and phospholipids from red blood cell membranes by nonionic detergents. J. Supramol. Struct. 1973;1:233–248. doi: 10.1002/jss.400010308. [DOI] [PubMed] [Google Scholar]
- Zhang P, Liu B, Jenkins GM, Hannun YA, Obeid LM. Expression of neutral sphingomyelinase identifies a distinct pool of sphingomyelin involved in apoptosis. J. Biol. Chem. 1997;272:9606–9612. doi: 10.1074/jbc.272.15.9609. [DOI] [PubMed] [Google Scholar]
- Zhang X, Thompson GA., Jr An apparent association between glycosylphosphatidylinositol-anchored proteins and a sphingolipid in Tetrahymena mimbres. Biochem. J. 1997;323:197–206. doi: 10.1042/bj3230197. [DOI] [PMC free article] [PubMed] [Google Scholar]