

Abstract
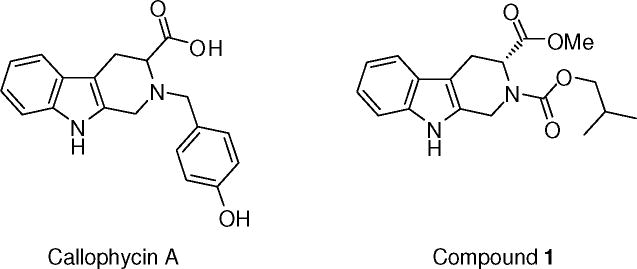
In previous studies, (R)-2-isobutyl 3-methyl 3,4-dihydro-1H-pyrido[3,4-b] indole-2,3(9H)-dicarboxylate (1), a callophycin A derivative, was found to strongly inhibit nitrite production in lipopolysaccharide (LPS)-stimulated RAW 264.7 cells, while (R)- or (S)-callophycin A showed only weak inhibition. We currently report additional studies to define the mechanisms underlying the inhibitory action of 1. Expression of inducible nitric oxide synthase (iNOS) was reduced at both protein and mRNA levels. Major upstream signaling molecules and transcription factors regulating iNOS expression were examined, but it was found that 1 did not affect the phosphorylated and total protein levels of p38 mitogen-activated protein kinase (p38 MAPK), Jun N-terminal kinase (JNK), extracellular signalregulated kinase 1/2 (ERK1/2), and signal transducer and activator of transcription 1 (STAT1), nor did it mediate the degradation of the inhibitor of nuclear factor-κBα-isoform (IκBα). However, starting at early time points, 1 consistently inhibited the phosphorylation of protein kinase B/Akt at serine 473. In addition, 1 suppressed the protein expression of octamer-binding transcription factor-2 (Oct-2) and the expression of microRNA 155 (miR-155). In sum, compound 1 inhibits LPS-induced nitrite production by a unique and complex mechanism. Reduction of iNOS expression is accompanied by inhibition of Akt activation, Oct-2 protein expression, and miR-155 expression.
Rhodophyta (red algae) have been used as food sources (e.g., Porphyra species and Palmaria palmata) as well as gelling and thickening agents due to a high content of sulfated polysaccharides.1 In addition, compounds or extracts from red algae have shown a variety of biological activities. In particular, of the Callophycus species (sp.), Callophycus serratus is frequently studied and has been reported to contain bromophycolides,2, 3 callophycoic acids,4 callophycols,4 and callophycolide A5 with antibacterial,4 antimalarial,4–6 or cancer cell cytotoxic activities.3, 4 Recently, callophycin A, bearing the tetrahydro-β-carboline scaffold, was isolated from Callophycus oppositifolius, and has been reported to exhibit cytotoxic potential against several human cancer cell lines7 and moderate cancer chemopreventive activities.8
Tetrahydro-β-carbolines also have been found in other marine organisms including tunicates, sponges, and marine-derived Streptomyces sp.: Eudistomins C, E, F, K and L from Eudistoma olivaceum,9 ma’eganedin A from Amphimedon sp.,10 shepherdine, 2,6-diacetyl-shepherdine and 1,6-dihydroxy-1,2,3,4-tetrahydro-β-carboline from Hyrtios reticulatus,11 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid12 and (−)(3S)-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid from the Chinese marine sponge Cinachyrella australiensis13 and Asteropus sp.,14 trans-1,2,3,4-tetrahydro-β-carboline-1,3-dicarboxylic acid, cis-1,2,3,4-tetrahydro-β-carboline-1,3-dicarboxylic acid and 1-methyltetrahydro-β-carboline from Haliplanella luciae,15 eudistomidins from Eudistoma glaucus,16, 17 and hyrtioreticulins A, B, and E and hyrtioerectine B from Hyrtios reticulatus.18 Among these naturally occurring tetrahydro-β-carboline compounds, 1-methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid,12 and eudistomidins G and B16 have shown cytotoxicity toward cancer cells, and hyrtioreticulins A and B inhibited ubiquitin-activating enzyme E1.18
Synthetic derivatives of tetrahydro-β-carbolines have shown inhibition of phosphodiesterase type-5,19 fatty acid amide hydrolase,20 mitogen-activated protein kinase (MAPK)-activated protein kinase 2 enzyme,21 aromatase,8 and NF-κB,8 interaction with transient receptor potential vanilloid type 120 and transient receptor potential channels,20 reduction of the Aβ1–42 neurotoxicity,22 induction of quinone reductase 1,8 anti-thrombotic activity,23 and anti-inflammatory activities.8, 24 However, in the literature, there are few comprehensive or detailed mechanistic studies on bioactive tetrahydro-β-carboline natural products and derivatives.
In previous work, we demonstrated that (R)-2-isobutyl 3-methyl 3,4-dihydro-1 H-pyrido[3,4-b] indole-2,3(9H)-dicarboxylate (1) exerted the most potent inhibitory effect on nitrite production in lipopolysaccharide (LPS)-induced RAW 264.7 cells with an IC50 value of 2.8 μM.8 We now report a more thorough account of the mechanism of action of 1, focusing on the expression or phosphorylation/activation levels of related proteins, mRNAs, and microRNA.
RESULTS AND DISCUSSION
Various factors can be involved in nitrite production, including catalytic activity or expression levels of iNOS, nitrite export machinery from cells to culture media, and so forth. In the current experimental setting, because the induction of iNOS enzyme is the major source of nitrite outburst following LPS exposure,25 the effects of (S)- and (R)-callophycin A, and compound 1 on protein expression levels were examined. The three compounds were evaluated under identical experimental conditions using Western blot analysis. As expected, based on the results of previous work,8 both (S)- and (R)-callophycin A barely inhibited LPS-induced iNOS expression, while compound 1 suppressed the expression essentially down to the basal level (Figure 1A). Therefore, further studies were conducted only with 1.
Figure 1.
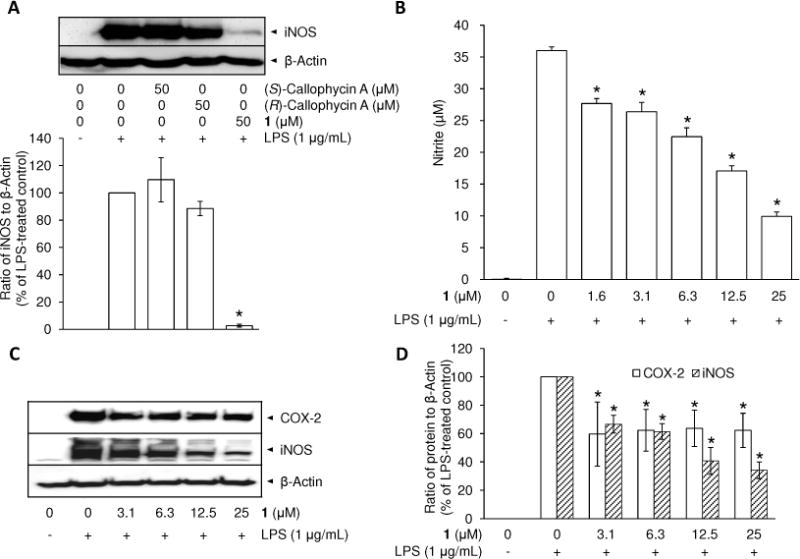
Comparison of the effect of (S)- and (R)-callophycin A, and 1 on iNOS protein expression (A), inhibitory effect of 1 on nitrite production (B), and protein expression of COX-2 and iNOS (C, D) in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells were pretreated with callophycin As or 1 for 15 min and stimulated with 1 μg/mL LPS for an additional 20 h. Protein expression levels of iNOS were examined by Western blot analyses (A). RAW 264.7 cells were pretreated with 1 (3.1–25 μM) for 15 min, and stimulated with 1 μg/mL LPS for 24 h. The level of nitrite was estimated using Griess reagent (B). The protein expression levels of COX-2 and iNOS were examined by Western blot analyses (C). After normalization to an internal standard β-actin, % expression relative to LPS-treated control (100%) is presented as means (n = 3) ± SD (D). An asterisk (*) indicates a significant difference between LPS-treated control and 1-pretreated/LPS-treated control with p values less than 0.05.
As shown in Figure 1B, the basal level of nitrite in the culture media of resting RAW 264.7 cells was undetectable. LPS-stimulation for 24 h led to a dramatic increase (36.0 ± 0.6 μM) which was attenuated by pretreatment with 1 in a concentration-dependent manner (IC50, 11.4 ± 1.4 μM). In order to examine iNOS protein expression, after collecting culture media for the nitrite assay, the treated cells were lysed. Pretreatment with 1 inhibited iNOS protein expression in a concentration-dependent manner (Figure 1C); at the highest concentration tested (25 μM), the expression level was reduced to 34.1 ± 5.8% in comparison with the LPS-treated control (Figure 1D). With the same experimental conditions, 1 moderately reduced another proinflammatory protein, COX-2, with an expression level of around 60% at each of the concentrations tested (3.1–25 μM).
Next, the effect of 1 on iNOS mRNA expression was examined. As indicated by RT-PCR analyses (Figure 2A), mRNA expression levels of iNOS were dramatically increased by 310 ± 50-fold following a 4 h treatment period with LPS in comparison with RAW 264.7 cells without LPS treatment (designated as 1-fold). Consistent with the reduction in protein expression, pretreatment with 1 prior to LPS treatment significantly suppressed iNOS mRNA expression in a concentration-dependent manner. At the highest concentration tested (25 μM), expression was reduced to 190 ± 50-fold after a 4 h incubation with LPS. As illustrated in time-dependent studies (Figure 2B), increases of 2530 ± 450- and 10150 ± 1850-fold were observed after 8 and 20 h incubation periods with LPS, respectively, while the pretreatment with 1 reduced the corresponding increases to 720 ± 80- and 530 ± 100-fold, respectively. Thus, the differences in iNOS mRNA expression levels between LPS-treated control groups, and 1-pretreated LPS-treated groups, were greater with longer incubation periods. COX-2 mRNA expression was also induced by treatment with LPS (490 ± 110-fold in comparison with basal levels), but this was not affected by pretreatment with 1 (Figure 2A).
Figure 2.
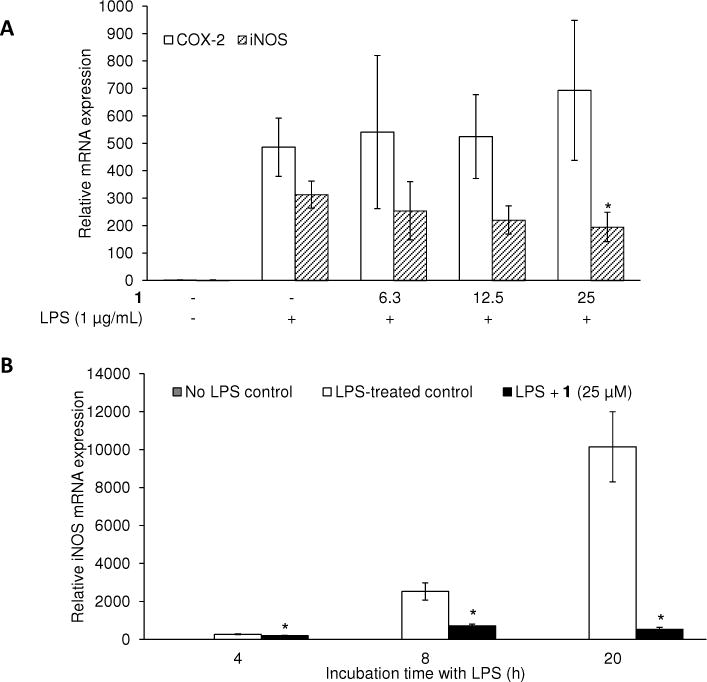
Effect of 1 on the mRNA expression levels of COX-2 and iNOS in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells were pretreated with the indicated concentrations of 1 for 15 min, and then stimulated with LPS. After a 4 h of incubation with LPS at various concentrations of 1 (A), or after the different incubation time points (4, 8 and 20 h) with the highest concentration at 25 μM (B), RNA was extracted and used in the RT-PCR reaction. After normalization to the internal standard β-actin, fold-changes relative to LPS-non-treated control is presented (n = 3). An asterisk (*) indicates a significant difference between LPS-treated control and 1-pretreated/LPS-treated control in each time point with p values less than 0.05.
The involvement of MAPKs and NF-κB in iNOS expression has been reported based on approaches using specific inhibitors. MAPK p38 inhibitor SB203580,26 ERK inhibitor PD98059,26 JNK inhibitor SP600125,27 and NF-κB inhibitor pyrrolidine dithiocarbamate,28 decreased LPS-induced expression of iNOS or production of NO in murine macrophage cell lines. Phosphorylation of specific residues on MAPKs is known to be essential for the kinase activation of MAPKs. The functional activation of MAPKs is mediated by dual phosphorylation on Thr180 and Tyr182 for p38 MAPK,29 Thr183 and Tyr185 for JNK,30 and Thr202 and Tyr204 for ERK.31 Therefore, the effect of 1 on the activation of MAPKs was next evaluated by examination of phosphorylation levels using Western blot analysis.
As previously reported, phosphorylation levels of MAPKs, including p38 MAPK, ERK1/2 and JNK, are rapidly increased following LPS stimulation and reach a maximal level after 10–30 min of incubation.32 Therefore, the phosphorylation levels of MAPKs were determined after 30 min of LPS treatment. As shown in Figure 3A, the LPS-treated control showed increased phosphorylation levels of each MAPK in comparison with untreated control cells, without changing total forms. However, pretreatment with 1 did not alter phosphorylation levels when tested at concentrations ranging from 3.1 to 25 μM. Therefore, as the levels of either phosphorylated or total MAPKs were not changed by treatment with 1 (Figure 3B, 3C), other pathways were explored to explain the reduction in NO production.
Figure 3.
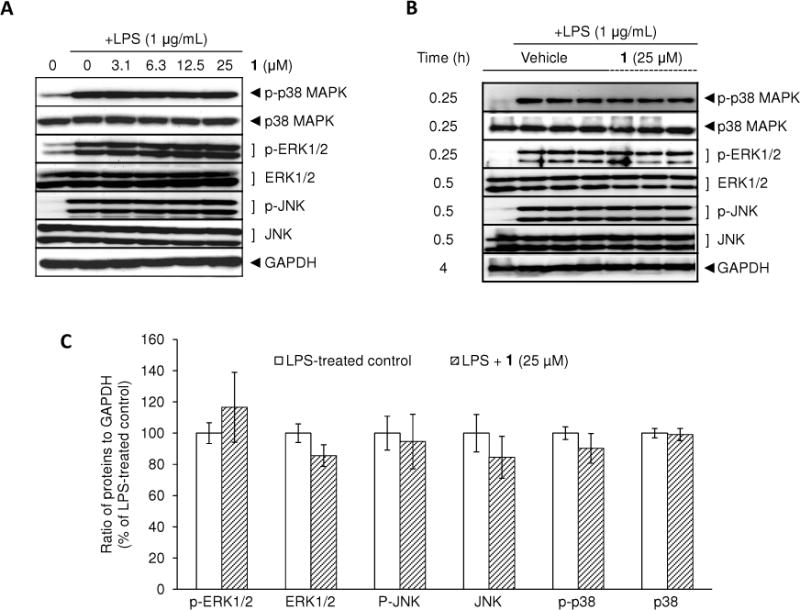
Effect of 1 on LPS-induced MAPKs phosphorylation in RAW 264.7 cells. RAW 264.7 cells were treated with various concentrations of 1 for 15 min prior to LPS (1 μg/mL) treatment for 30 min. The protein expression levels of MAPKs were determined using Western blot analyses (A). After normalization to the internal standard GAPDH, % expression of the 1 (25 μM) pretreatment group relative to LPS-treated control in each MAPK (100%) is presented (n = 3) (B, C).
IκB plays an essential role in the nuclear translocation of NF-κB because under resting conditions nearly all NF-κB resides in the cytosol as an inactivated state due to binding with IκBα. However, once IκBα is phosphorylated by IκB kinases, it is rapidly degraded, leading to the release and nuclear translocation of NF-κB.33 Therefore, protein levels of phosphorylated and total IκBα were examined following LPS-stimulation. Based on previous reports of the complete degradation and recovery of IκBα after LPS treatment,32 RAW 264.7 cells were pretreated with vehicle (0.5% DMSO) or 1 (25 μM) and incubated with LPS for 10, 15, 30, and 60 min. As shown in Figure 4A, the protein level of phosphorylated IκBα was increased while that of total IκBα was decreased after 10 min of LPS treatment. Pretreatment with 1 in the concentration range of 3.1–25 μM had no effect on the protein levels of either phosphorylated or total IκBα. To confirm these results, experiments were repeated only with the highest concentration of 1 with different incubation times. Although 1 did not attenuate the degradation of IκBα, it seemed to slightly facilitate the recovery of IκBα from degradation based on the result of a 30 min incubation with LPS (Figure 4B, 4C). In line with these results, pretreatment with 1 had no effect on the nuclear localization of the NF-κB major subunit p65 after 1 h of LPS treatment (Figure 4D).
Figure 4.
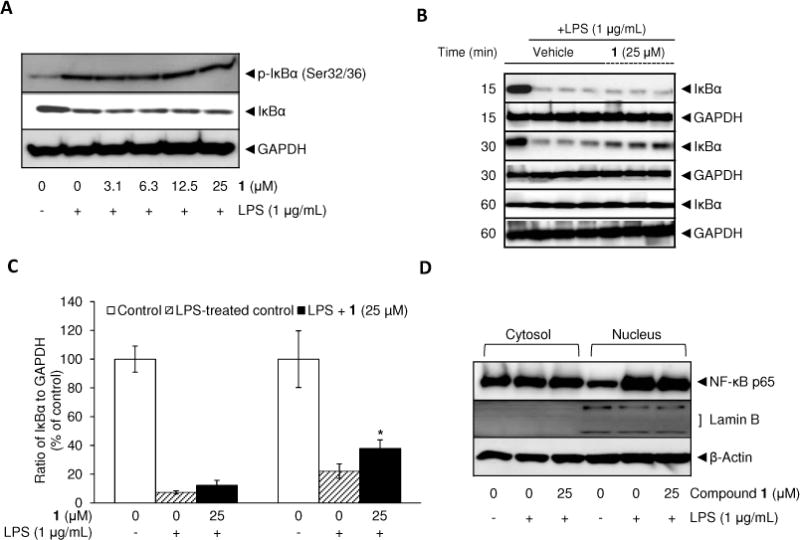
Effect of 1 on LPS-induced phosphorylation and degradation of IκBα (A–C) and nuclear localization of NF-κB p65 (D). RAW 264.7 cells were treated with various concentrations of 1 for 15 min prior to LPS (1 μg/mL) treatment for 10 min (A), or treated with the highest concentration (25 μM) for 15 min prior to LPS (1 μg/mL) treatment for different incubation times (15, 30, and 60 min) (B, C). Also, after 60 min incubation with LPS, nuclear and cytosolic proteins were extracted to determine nuclear translocation of the NF-κB subunit p65 (D). Lamin B was used as a nuclear marker. Protein expression levels were determined by Western blot analysis. After normalization to the internal standard GAPDH, % expression of the 1 (25 μM) pretreatment group or LPS-treated group relative to IκBα expression in LPS-non-treated control (100%) is presented (n = 3) (B, C). An asterisk (*) indicates a significant difference between LPS-treated control and 1-pretreated/LPS-treated control in each time point with p values less than 0.05.
After exposure to LPS, Toll-like receptor 4 (TLR4) propagates signals through two major adaptor proteins including myeloid differentiation factor 88 (MyD88) and Toll/IL-1 receptor (TIR)-containing adapter inducing IFN-β (TRIF). Through the TRIF pathway, phosphorylated/activated signal transducer and activator of transcription 1 (STAT1) functions as a transcription factor for iNOS gene expression.34 Therefore, the effect of 1 on the phosphorylation of STAT1 was examined to determine the potential of mediating an inhibitory effect on iNOS expression through the STAT1 pathway. As previously described,25 the protein levels of phosphorylated STAT1 peaked around 4 to 6 h of incubation with LPS. However, the protein expression levels of both phosphorylated and total STAT1 were not changed by the treatment of 1 (3.1–25 μM) prior to LPS exposure for 4 h (Figure 5A, 5B). In accordance with this result, nuclear localization of STAT1 by LPS was not changed by pretreatment with 1 (Figure 5C), so it does not appear this pathway is involved in the inhibitory effect mediated by 1.
Figure 5.
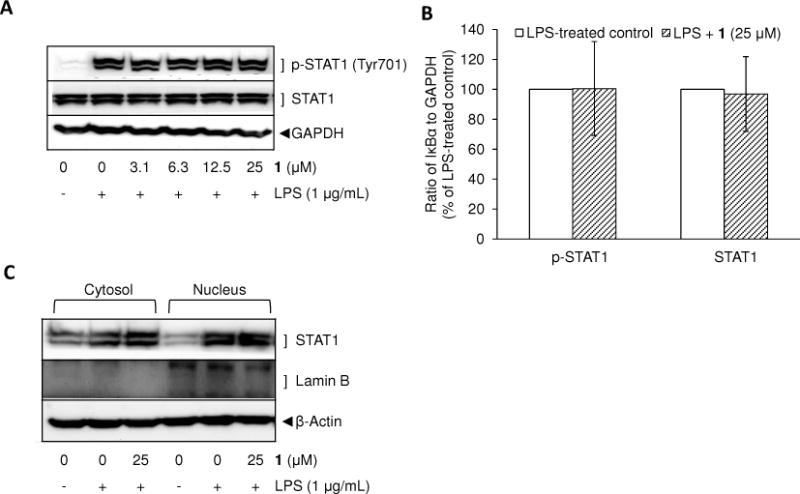
Effect of 1 on LPS-induced phosphorylation (A, B) and nuclear translocation of STAT1 (C). RAW 264.7 cells were treated with various concentrations of 1 for 15 min prior to LPS (1 μg/mL) treatment for 4 min (A). The experiments were repeated with the highest concentration of 1 (25 μM) (B). Also, after 6 h of incubation with LPS, the nuclear and cytosolic proteins were extracted to determine nuclear translocation of STAT1. The protein expression levels were determined by Western blot analysis. After normalization to the internal standard GAPDH, % expression of the 1 (25 μM) pretreatment group relative to p-STAT1 and STAT1 expression in LPS-treated control (100%) is presented (n = 3) (B).
Using a phosphatidylinositol 3-kinase (PI3K) inhibitor LY294002, it was shown that PI3K/Akt positively regulates iNOS expression in LPS-stimulated RAW 264.7 cells.35 Also, dominant-negative-Akt expression in cells attenuated LPS-induced iNOS expression.36 Based on this information, the effect of 1 on the activation of Akt was examined. As shown in Figure 6A, the phosphorylation level of Akt at Serine 473 started to increase from 10 min and attained the maximal level by around 6 h of incubation with LPS. Accordingly, the effect of 1 on the phosphorylation level of Akt was determined. Compound 1 did not affect the protein expression levels of total Akt at each incubation period with LPS (Figure 6B, 6C), but the treatment did lead to the attenuation of LPS-induced p-Akt at each incubation time point (15 min, 30 min, and 4 h) with significant differences relative to the expression levels of LPS-treated controls.
Figure 6.
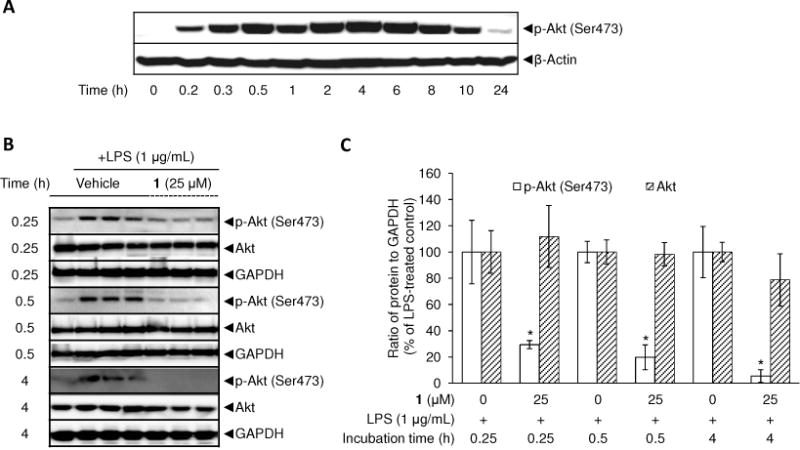
Effect of 1 on LPS-induced phosphorylation of Akt at Ser473 residue in RAW 264.7 cells. Time-course of p-Akt and Akt protein expression after LPS treatment was examined (A). RAW 264.7 cells were treated with 1 μg/mL LPS for 0.25, 0.5, and 4 h and lysed to determine protein expression levels of phosphorylated and total Akt using Western blot analyses (B). After normalization to the internal standard GAPDH, % expression of the 1 (25 μM) pretreatment group relative to p-Akt and Akt expression in LPS-treated control (100%) is presented (n = 3) (C). An asterisk (*) indicates a significant difference between LPS-treated control and 1-pretreated/LPS-treated control with p values less than 0.05.
Using inhibitors (e.g., LY294002 or an Akt inhibitor) or gene knock-out approaches (e.g., transfection of dominant-negative-Akt)36 with LPS-stimulated RAW 264.7 cells, the PI3K/Akt/mammalian target of rapamycin (m-TOR) signaling pathway has been shown to be involved in the induction of Oct-2 expression. Moreover, as determined by chromatin immunoprecipitation assays, the binding activity of octamer binding protein Oct-2 to the iNOS promoter region was increased in response to LPS, indicating that Oct-2 activation is a crucial step for transcriptional activation of the iNOS gene.37
As 1 inhibited the phosphorylation of Akt, the effect of this compound on Oct-2 protein expression levels was examined. Oct-2 protein expression levels showed maximal expression after 8 h of incubation with LPS (Figure 7A). Following pretreatment with 1, the protein levels of Oct-2 induced by LPS were reduced at 8 and 20 h but not at earlier time points (Figure 7A, 7B).
Figure 7.
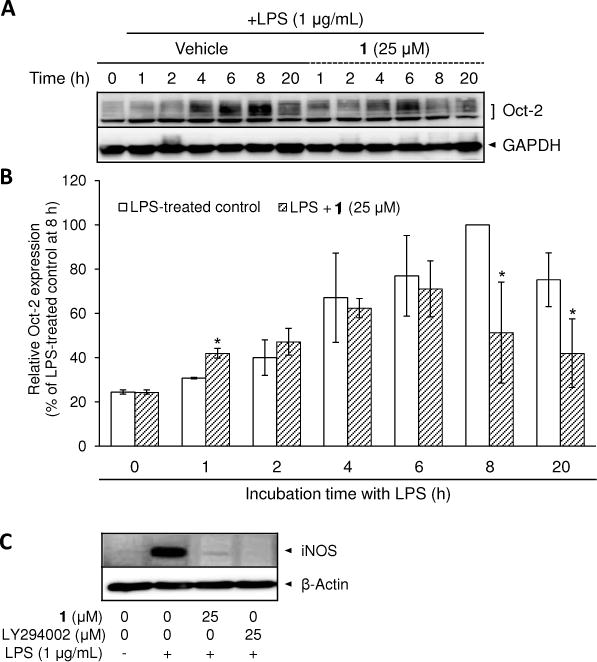
Effect of 1 and LY294002 on LPS-induced protein expression of Oct-2 and iNOS in RAW 264.7 cells. Time-course of Oct-2 protein expression after LPS treatment with or without 1-pretreatment was examined. RAW 264.7 cells incubated with 1 μg/mL LPS for 1, 2, 4, 6, 8, and 20 h were lysed to determine protein expression levels of Oct-2 using Western blot analyses (A). After normalization to the internal standard GAPDH, % expression of the 1 (25 μM) pretreatment group relative to Oct-2 expression in LPS-treated control (100%) is presented (n = 3) (B). The inhibition of LPS-induced iNOS protein expression by 1 and LY294002 has been examined after 20 h treatment with LPS (C). An asterisk (*) indicates a significant difference between LPS-treated control and 1-pretreated/LPS-treated control with p values less than 0.05.
LY294002 (10–50 μM) has been commonly used for inhibition of the PI3K/Akt signaling pathway in LPS-stimulated RAW 264.7 cells.38–40 In accordance with the result by Kim et al.,35 pretreatment with LY294002 (25 μM) inhibited iNOS protein expression induced by LPS down to the basal level in RAW 264.7 cells. The inhibitory effects of 1 and LY294002 on iNOS protein levels are basically equivalent (Figure 7C). The complete blockade of iNOS expression via LY294002 or 1 confirms the PI3K/Akt signaling pathway plays a crucial role in iNOS expression.
Following treatment with LPS (100 ng/mL), LY294002 (12.5–50 μM) is known to inhibit Oct-2 expression.36 To determine if a response similar to that of the positive control LY294002 was mediated by 1, we pretreated cells with 1 and LY294002 (25 μM) and examined expression levels. As shown in Figure S1, Supporting Information, the expression of Oct-2 reaches a maximum after about 8 h of incubation, and the inhibitory effect mediated by 1 was essentially equivalent to LY294002.
In addition, microRNAs (miRs) are small, non-coding RNAs that modulate gene expression post-transcriptionally. It was found that miR-155 was up-regulated by LPS induction.41 It has been reported that miR-155 repressed the mRNA and protein expression of Src homology-2 domain-containing inositol 5-phosphatase 1 (SHIP1), a negative regulator of Akt, via interaction with the 3′-untranslated region (3′UTR) of SHIP1 mRNA, leading to up-regulation of serine 473-phosphorylation of Akt in RAW 264.7 cells.42 miR-155 plays an important role in the immune response, but high expression levels are observed in cancers and leukemias.43 As shown in Figure 8, the expression level of miR-155 was increased by 50 ± 10- and 290 ± 90-fold following 4 and 20 h incubation with LPS, respectively, compared with the non-treated control. Compound 1 did not alter the expression level of miR-155 at 4 h, but after 20 h, the expression level was attenuated in a dose-dependent manner, down to 130 ± 20-fold, at a concentration of 25 μM.
Figure 8.
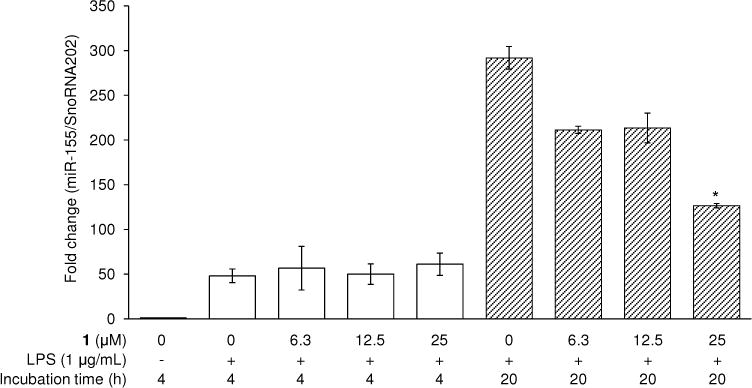
Effect of 1 on LPS-induced expression of miR-155 in RAW 264.7 cells. RAW 264.7 cells were treated with 1 (6.3–25 μM) prior to 1 μg/mL of LPS treatment for 4 and 20 h. RNA was extracted and used in the RT-PCR reaction. After normalization to the internal standard snoRNA202, fold-changes relative to vehicle-treated control are presented (n = 3). An asterisk (*) indicates a significant difference between LPS-treated control and 1-pretreated/LPS-treated control in each time point with p values less than 0.05.
Thus, some hints are beginning to emerge to explain the inhibitory activity of 1; the work is ongoing. Yet another explanation for inhibitory effects could be instability of iNOS mRNA. As reported by Lahti et al.,44 the rate of decrease of iNOS mRNA was similar in the presence or absence of the RNA synthesis inhibitor actinomycin D, so degradation is of relevance. In our preliminary work, the effect of 1 on mRNA degradation was examined. First, RAW 264.7 cells were exposed to LPS for 24 h, LPS-containing media was completely removed, cells were washed with PBS, and iNOS mRNA levels were determined as a function of time. In comparison with cells in which LPS remained in the incubation media, iNOS mRNA levels significantly diminished in a time-dependent manner (Figure S2; Supporting Information). Interestingly, when LPS was removed from the media but cells continued to be incubated in the presence of 1, the rate by which iNOS mRNA decreased was significantly accelerated. These results suggest 1 might be involved in the degradation/instability of iNOS mRNA.
In a related manner, JNK inhibitor SP60012545 and mTOR kinase inhibitor rapamycin46 have been reported to destabilize iNOS mRNA, thereby regulating the rate of iNOS mRNA degradation in LPS-activated J774 murine macrophage cells, and in a mixture of proinflammatory cytokine-stimulated astrocytes, respectively. mTOR is one of the downstream target molecules of Akt. Therefore, 1 might affect iNOS mRNA stability by a mode similar to rapamycin. Alternatively, regulators involved in mRNA stability, including tristetraprolin, KH-type splicing regulatory protein, and HuR,46 could be affected by treatment with 1.
In sum, 1 is a remarkable derivative of callophycin A that inhibits LPS-induced NO generation in RAW 264.7 cells by an elusive mechanism. After a good deal of work, we have observed the potential of 1 to inhibit Akt phosphorylation, Oct-2 expression and miR-155 expression, and possibly to accelerate iNOS mRNA degradation. When tested with LPS-stimulated RAW 264.7 cells at concentrations in the same range as our evaluations performed with 1, other natural products have been shown to reduce iNOS expression accompanied by inhibition of Akt phosphorylation: tanshinone IIA (1–20 μg/mL),38 a combination of luteolin (2–16 μM) and chicoric acid (4–32 μM),47 alantolactone (10 μM),39 ampelopsin (150 μg/mL),40 and miyabenol A (3–10 μM).48
On the other hand, some studies have shown a negative role of PI3K/Akt in inflammatory signaling. For example, increased Akt phosphorylation along with the inhibition of iNOS expression was observed with LPS-activated RAW 264.7 cells treated with resveratrol49 or icariin.50 Therefore, further studies are warranted to elucidate the link between the Akt signaling pathway and iNOS expression, and further studies are still required to unequivocally define the role of 1 in these processes.
EXPERIMENTAL SECTION
General Experimental Procedures
(S)- and (R)-Callophycin A and 1 were chemically synthesized as previously described.8 Structural characterization and purity of the synthesized compounds have been reported previously.8 LPS was purchased from Sigma-Aldrich Co. NE-PER Nuclear and Cytoplasmic Extraction Reagents were purchased from Thermo Fisher Scientific Inc. Cell Lysis Buffer (10 ×), antibodies against β-actin, GAPDH, COX-2, phospho (p)-ERK1/2 (Thr202/Tyr204), ERK1/2, p-p38 MAPK (Thr180/Tyr182), p38 MAPK, p-JNK (Thr183/Tyr185), JNK, NF-κB p65, IκBα, p-IκBα (Ser32/36), p-Akt (Ser473), pan-Akt, p-STAT1 (Tyr701), or STAT1 were purchased from Cell Signaling Technology, Inc. Antibodies against iNOS, lamin B, or Oct-2 were purchased from Santa Cruz Biotechnology, Inc. Enhanced chemiluminescence (ECL) detection kit was purchased from GE Healthcare Bio-Sciences. iScriptReverse Transcription Supermix for RT-qPCR was purchased from BioRad Laboratories, Inc. PerfeCTa SyBR Green FastMix, ROX was purchased from Quanta biosciences. FluoroTrans® PVDF transfer membranes (Pall Life Sciences, Ann Arbor, MI, USA). mmu-miR-155 (mmu-miR-155-5p), snoRNA202, TaqMan MicroRNA Reverse Transcription Kit, TaqMan® Universal PCR Master Mix, No AmpErase UNG, 1-Pack (1 × 5 mL) were purchased from Applied Biosystems by Life Technologies. Penicillin-streptomycin Solution (50×) was purchased from Mediatech, Inc. DMEM with 4.5 g/L glucose without L-glutamine and phenol red was purchased from Lonza Group Ltd.
Cell Culture
RAW 264.7 murine macrophage-like cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% (v/v) heat-inactivated fetal bovine serum, penicillin G potassium salt (100 IU/mL), and streptomycin sulfate (100 μg/mL) in a 37 °C humidified incubator with 5% CO2 in air.
Measurement of Nitrite Production
The assay was performed as previously described with a slight modification.51 RAW 264.7 cells were plated at a density of 5 × 105 per well in a 24-well culture plate and incubated in a 37 °C humidified incubator with 5% CO2 in air for 24 h. The incubated cells were pretreated with phenol red-free medium containing various concentrations of 1 for 15 min, followed by 1 μg/mL of LPS treatment for an additional 24 h. The nitrite concentration in the culture media was measured using Griess reagent.
Western Blot Analysis
The total cell lysate, or nuclear and cytosolic protein extracts were prepared according to the manufacturer’s instructions as described previously25, 32 and stored at −80 °C until use. Equal amounts of total protein in each cell lysate were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) (8–12%), and electrotransferred to PVDF membranes. After blocking with 5% skimmed milk in 0.1% Tween 20-containing TBS, membranes were incubated with corresponding primary antibodies, followed by incubation with secondary antibodies conjugated with horseradish peroxidase. Chemiluminescent signal from each immunoblot was detected using chargecoupled device-based digital image analysis system (Geliance 1000 imager, Perkin Elmer, Inc). Intensity of bands was quantified using ImageJ software from the National Institutes of Health (http://rsbweb.nih.gov/ij/).
Real Time-polymerase Chain Reaction (RT-PCR) Analysis
Total RNA extracted from cells using Trizol reagent52 was dissolved in RNase-free water. After the measurement of concentration and purity of isolated RNA using a BioSpec-nano spectrophotometer (Shimadzu Scientific Instruments, Inc.), 1 μg of total RNA was reverse transcribed using iScriptReverse Transcription Supermix for RT-qPCR on a ABI 7300 thermocycler (Applied Biosystems Inc.). The cDNA amplification on iNOS, COX-2, and β-actin was performed with SyBR green method as the fluorescence signal as previously described.25 Expression calculated using the 2−∆∆CT method was performed to analyze the results.51
miRNA Analysis
RNA was isolated by the method of Chomczynski and Sacchi52 using Trizol since it has been demonstrated as a gold standard for miRNA isolation.53 Isolated RNA was reverse transcribed with corresponding RT primers described in the previous report25 using TaqMan MicroRNA Reverse Transcription Kit according to the manufacturer’s protocol. Then, the product was amplified using TaqMan Universal PCR Master Mix, No AmpErase UNG, and corresponding TaqMan Small RNA Assay (20 ×) for mmu-miR-155 (mmu-miR-155-5p), and a control reference RNA snoRNA202.
Statistical Analysis
Data are presented as means ± standard deviation (SD) for the indicated number of three independent experiments. Statistical significance was determined by one-way analysis of variance (ANOVA) using Microsoft Excel 2007 software (Microsoft Corporation). P values less than 0.05 (p < 0.05) were considered statistically significant. The IC50 values were calculated using TableCurve software 2D (version 4) curve-fitting software (Jandel Scientific).
Supplementary Material
STRUCTURES.
Acknowledgments
This project was supported by a grant from the National Institute of General Medical Sciences, National Institutes of Health, award number: P20GM 103466. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Footnotes
SPECIAL ISSUE
This article is dedicated to Prof. Dr. Otto Sticher, of ETH-Zurich, Zurich, Switzerland, for his pioneering work in pharmacognosy and phytochemistry.
ASSOCIATED CONTENT
Supporting Information. Effect of 1 and LY294002 on Oct-2 protein expression in LPS-stimulated RAW 264.7 cells, and effect of 1 on the rate of mRNA decrease after LPS removal using RT-PCR. This material is available free of charge via the Internet at http://pubs.acs.org.
Notes
The authors declare no competing financial interest.
References
- 1.Park EJ, Pezzuto JM. Antioxid Redox Signal. 2013;19:115–138. doi: 10.1089/ars.2013.5235. [DOI] [PubMed] [Google Scholar]
- 2.Kubanek J, Prusak AC, Snell TW, Giese RA, Hardcastle KI, Fairchild CR, Aalbersberg W, Raventos-Suarez C, Hay ME. Org Lett. 2005;7:5261–5264. doi: 10.1021/ol052121f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kubanek J, Prusak AC, Snell TW, Giese RA, Fairchild CR, Aalbersberg W, Hay ME. J Nat Prod. 2006;69:731–735. doi: 10.1021/np050463o. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Lane AL, Stout EP, Hay ME, Prusak AC, Hardcastle K, Fairchild CR, Franzblau SG, Le Roch K, Prudhomme J, Aalbersberg W, Kubanek J. J Org Chem. 2007;72:7343–7351. doi: 10.1021/jo071210y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Stout EP, Prudhomme J, Roch KL, Fairchild CR, Franzblau SG, Aalbersberg W, Hay ME, Kubanek J. Bioorg Med Chem Lett. 2010;20:5662–5665. doi: 10.1016/j.bmcl.2010.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lane AL, Stout EP, Lin AS, Prudhomme J, Le Roch K, Fairchild CR, Franzblau SG, Hay ME, Aalbersberg W, Kubanek J. J Org Chem. 2009;74:2736–2742. doi: 10.1021/jo900008w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ovenden SPB, Nielson JL, Liptrot CH, Willis RH, Tapiolas DM, Wright AD, Motti CA. Phytochem Lett. 2011;4:69–71. [Google Scholar]
- 8.Shen L, Park EJ, Kondratyuk TP, Guendisch D, Marler L, Pezzuto JM, Wright AD, Sun D. Bioorg Med Chem. 2011;19:6182–6195. doi: 10.1016/j.bmc.2011.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Rinehart KL, Jr, Kobayashi J, Harbour GC, Gilmore J, Mascal M, Holt TG, Shield LS, Lafargue F. J Am Chem Soc. 1987;109:3378–3387. [Google Scholar]
- 10.Tsuda M, Watanabe D, Kobayashi Ji. Tetrahedron Lett. 1998;39:1207–1210. [Google Scholar]
- 11.Salmoun M, Devijver C, Daloze D, Braekman JC, van Soest RW. J Nat Prod. 2002;65:1173–1176. doi: 10.1021/np020009+. [DOI] [PubMed] [Google Scholar]
- 12.Ren H, Gu Q, Cui C, Zhu W. J Ocean Univ China. 2006;5:75–81. [Google Scholar]
- 13.Li L, Deng Z, Li J, Fu H, Lin W. Beijing Daxue Xuebao, Yixueban. 2004;36:12–17. [Google Scholar]
- 14.Li H, Dang HT, Li J, Sim CJ, Hong J, Kim D-K, Jung JH. Biochem Syst Ecol. 2010;38:1049–1051. [Google Scholar]
- 15.Lu Y, Fan C, Xi D, Zhao S, Miao Y. Zhongguo Haiyang Yaowu. 2010;29:45–49. [Google Scholar]
- 16.Takahashi Y, Ishiyama H, Kubota T, Kobayashi J. Bioorg Med Chem Lett. 2010;20:4100–4103. doi: 10.1016/j.bmcl.2010.05.071. [DOI] [PubMed] [Google Scholar]
- 17.Suzuki T, Kubota T, Kobayashi J. Bioorg Med Chem Lett. 2011;21:4220–4223. doi: 10.1016/j.bmcl.2011.05.072. [DOI] [PubMed] [Google Scholar]
- 18.Yamanokuchi R, Imada K, Miyazaki M, Kato H, Watanabe T, Fujimuro M, Saeki Y, Yoshinaga S, Terasawa H, Iwasaki N, Rotinsulu H, Losung F, Mangindaan RE, Namikoshi M, de Voogd NJ, Yokosawa H, Tsukamoto S. Bioorg Med Chem. 2012;20:4437–4442. doi: 10.1016/j.bmc.2012.05.044. [DOI] [PubMed] [Google Scholar]
- 19.El-Gamil DS, Ahmed NS, Gary BD, Piazza GA, Engel M, Hartmann RW, Abadi AH. Arch Pharm (Weinheim) 2013;346:23–33. doi: 10.1002/ardp.201200334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ortar G, De Petrocellis L, Moriello AS, Allarà M, Morera E, Nalli M, Di Marzo V. Bioorg Med Chem Lett. 2013;23:138–142. doi: 10.1016/j.bmcl.2012.10.137. [DOI] [PubMed] [Google Scholar]
- 21.Trujillo JI, Meyers MJ, Anderson DR, Hegde S, Mahoney MW, Vernier WF, Buchler IP, Wu KK, Yang S, Hartmann SJ, Reitz DB. Bioorg Med Chem Lett. 2007;17:4657–4663. doi: 10.1016/j.bmcl.2007.05.070. [DOI] [PubMed] [Google Scholar]
- 22.Jiaranaikulwanitch J, Govitrapong P, Fokin VV, Vajragupta O. Molecules. 2012;17:8312–8333. doi: 10.3390/molecules17078312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Yao K, Zhao M, Zhang X, Wang Y, Li L, Zheng M, Peng S. Eur J Med Chem. 2011;46:3237–3249. doi: 10.1016/j.ejmech.2011.04.037. [DOI] [PubMed] [Google Scholar]
- 24.Zeng L, Zhang J. Bioorg Med Chem Lett. 2012;22:3718–3722. doi: 10.1016/j.bmcl.2012.04.020. [DOI] [PubMed] [Google Scholar]
- 25.Park EJ, Pezzuto JM, Jang KH, Nam SJ, Bucarey SA, Fenical W. Nat Prod Commun. 2012;7:789–794. [PMC free article] [PubMed] [Google Scholar]
- 26.Kim SH, Kim J, Sharma RP. Pharmacol Res. 2004;49:433–439. doi: 10.1016/j.phrs.2003.11.004. [DOI] [PubMed] [Google Scholar]
- 27.Lin YC, Huang DY, Chu CL, Lin WW. Mol Immunol. 2010;47:1569–1578. doi: 10.1016/j.molimm.2010.01.008. [DOI] [PubMed] [Google Scholar]
- 28.Xie QW, Kashiwabara Y, Nathan C. J Biol Chem. 1994;269:4705–4708. [PubMed] [Google Scholar]
- 29.Raingeaud J, Gupta S, Rogers JS, Dickens M, Han J, Ulevitch RJ, Davis RJ. J Biol Chem. 1995;270:7420–7426. doi: 10.1074/jbc.270.13.7420. [DOI] [PubMed] [Google Scholar]
- 30.Dérijard B, Hibi M, Wu IH, Barrett T, Su B, Deng T, Karin M, Davis RJ. Cell. 1994;76:1025–1037. doi: 10.1016/0092-8674(94)90380-8. [DOI] [PubMed] [Google Scholar]
- 31.Roskoski R. Pharmacol Res. 2012;66:105–143. doi: 10.1016/j.phrs.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 32.Park EJ, Cheenpracha S, Chang LC, Kondratyuk TP, Pezzuto JM. Nutr Cancer. 2011;63:971–982. doi: 10.1080/01635581.2011.589960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ferreiro DU, Komives EA. Biochemistry. 2010;49:1560–1567. doi: 10.1021/bi901948j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gao JJ, Filla MB, Fultz MJ, Vogel SN, Russell SW, Murphy WJ. J Immunol. 1998;161:4803–4810. [PubMed] [Google Scholar]
- 35.Kim YH, Choi KH, Park JW, Kwon TK. Immunol Lett. 2005;99:45–50. doi: 10.1016/j.imlet.2004.12.007. [DOI] [PubMed] [Google Scholar]
- 36.Chou YY, Gao JI, Chang SF, Chang PY, Lu SC. FEBS J. 2011;278:85–96. doi: 10.1111/j.1742-4658.2010.07929.x. [DOI] [PubMed] [Google Scholar]
- 37.Lu SC, Wu HW, Lin YJ, Chang SF. Am J Physiol Cell Physiol. 2009;296:C1133–C1139. doi: 10.1152/ajpcell.00031.2009. [DOI] [PubMed] [Google Scholar]
- 38.Xu M, Cao F, Liu L, Zhang B, Wang Y, Dong H, Cui Y, Dong M, Xu D, Liu Y, Zhao P, Niu W, Li Z. Am J Respir Cell Mol Biol. 2011;45:1028–1035. doi: 10.1165/rcmb.2011-0113OC. [DOI] [PubMed] [Google Scholar]
- 39.Chun J, Choi RJ, Khan S, Lee DS, Kim YC, Nam YJ, Lee DU, Kim YS. Int Immunopharmacol. 2012;14:375–383. doi: 10.1016/j.intimp.2012.08.011. [DOI] [PubMed] [Google Scholar]
- 40.Qi S, Xin Y, Guo Y, Diao Y, Kou X, Luo L, Yin Z. Int Immunopharmacol. 2012;12:278–287. doi: 10.1016/j.intimp.2011.12.001. [DOI] [PubMed] [Google Scholar]
- 41.Tili E, Michaille JJ, Cimino A, Costinean S, Dumitru CD, Adair B, Fabbri M, Alder H, Liu CG, Calin GA, Croce CM. J Immunol. 2007;179:5082–5089. doi: 10.4049/jimmunol.179.8.5082. [DOI] [PubMed] [Google Scholar]
- 42.O’Connell RM, Chaudhuri AA, Rao DS, Baltimore D. Proc Natl Acad Sci USA. 2009;106:7113–7118. doi: 10.1073/pnas.0902636106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tili E, Michaille JJ, Croce CM. Immunol Rev. 2013;253:167–184. doi: 10.1111/imr.12050. [DOI] [PubMed] [Google Scholar]
- 44.Lahti A, Sareila O, Kankaanranta H, Moilanen E. BMC Pharmacol. 2006;6:5. doi: 10.1186/1471-2210-6-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lahti A, Jalonen U, Kankaanranta H, Moilanen E. Mol Pharmacol. 2003;64:308–315. doi: 10.1124/mol.64.2.308. [DOI] [PubMed] [Google Scholar]
- 46.Lisi L, Navarra P, Feinstein DL, Dello Russo C. J Neuroinflammation. 2011;8:1. doi: 10.1186/1742-2094-8-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Park CM, Jin KS, Lee YW, Song YS. Eur J Pharmacol. 2011;660:454–459. doi: 10.1016/j.ejphar.2011.04.007. [DOI] [PubMed] [Google Scholar]
- 48.Ku KT, Huang YL, Huang YJ, Chiou WF. J Agric Food Chem. 2008;56:8911–8918. doi: 10.1021/jf8019369. [DOI] [PubMed] [Google Scholar]
- 49.Zong Y, Sun L, Liu B, Deng YS, Zhan D, Chen YL, He Y, Liu J, Zhang ZJ, Sun J, Lu D. PLoS One. 2012;7:e44107. doi: 10.1371/journal.pone.0044107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Xu CQ, Liu BJ, Wu JF, Xu YC, Duan XH, Cao YX, Dong JC. Eur J Pharmacol. 2010;642:146–153. doi: 10.1016/j.ejphar.2010.05.012. [DOI] [PubMed] [Google Scholar]
- 51.Park EJ, Cheenpracha S, Chang LC, Pezzuto JM. Phytochem Lett. 2011;4:426–431. doi: 10.1016/j.phytol.2011.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chomczynski P, Sacchi N. Anal Biochem. 1987;162:156–159. doi: 10.1006/abio.1987.9999. [DOI] [PubMed] [Google Scholar]
- 53.Mraz M, Malinova K, Mayer J, Pospisilova S. Biochem Biophys Res Commun. 2009;390:1–4. doi: 10.1016/j.bbrc.2009.09.061. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.