

Abstract
Head direction (HD) cells respond when an animal faces a particular direction in the environment and form the basis for the animal's perceived directional heading. When an animal moves through its environment, accurate updating of the HD signal is required to reflect the current heading, but the cells still maintain a representation of HD even when the animal is motionless. This finding suggests that the HD system holds its current state in the absence of input, a view that we tested by rotating a head-restrained rat in the presence of a prominent visual landmark and then stopping it suddenly when facing the cell's preferred firing direction (PFD). Firing rates were unchanged for the first 100 ms, but then progressively decreased over the next 4 s and stabilized at ∼42% of their initial values. When the rat was stopped facing away from the PFD, there was no initial effect of braking, but the firing rate then increased steadily over 4 s and plateaued at ∼14% of its peak firing rate, substantially above initial background firing rates. In experiment 2, the rat was serially placed facing one of eight equidistant directions over 360° and held there for 30 s. Compared with the cell's peak firing rate during a passive rotation session, firing rates were reduced (51%) for in-PFD directions and increased (∼300%) from background levels for off-PFD directions, values comparable to those observed in the braking protocol. These differential HD cell responses demonstrate the importance of self-motion to the HD signal integrity.
Keywords: self-motion, vestibular, head direction cell, anterodorsal thalamus, passive, restraint
an individual's perceived directional orientation in the environment is represented by head direction (HD) cells in the brain, which are activated when the head faces a particular direction. Each HD cell selectively encodes a particular direction, and all possible directions are represented uniformly across the HD cell population (Taube 2007; Taube et al. 1990a). As an animal turns its head, the firing rates of HD cells that represent the direction the animal was previously facing decrease, while the firing rates increase for HD cells representing the new current direction. The continuous output of the HD cell network therefore represents the animal's perceived directional heading by the selection of which cells are activated at a given moment (Valerio and Taube 2012). The perception of directional heading encoded by the HD cell network is intertwined with the perception of changes in angular head movements, which simultaneously changes the active population of HD cells to represent the animal's current heading. The neural substrate for angular head movements is derived to a large extent from the vestibular sensory system. The vestibular semicircular canals send ascending sensory information about angular head velocity (AHV) to the HD cell network (see Shinder and Taube 2010 for review).
There are two basic properties that define HD cell responses: 1) the cell's firing rate, and 2) the cell's preferred firing direction (PFD). Consistent with the vestibular projection to the HD system, it was found that lesions of the vestibular sensory organs completely disrupt the HD signal (Stackman and Taube 1997) and result in spatial memory deficits (Baek et al. 2010; Russell et al. 2003; Wallace et al. 2002). The loss of direction-specific activity in HD cells could not be recovered with either proprioceptive feedback from head movements, or from orientation information from familiar visual landmarks. Conversely, the HD signal remained intact during passive head rotations that reduced or eliminated visual, motor, proprioceptive, and tactile information related to movement, as long as vestibular sensory functions were not impaired (Shinder and Taube 2011). Thus, while the motor command to execute a head movement cannot by itself generate the HD signal, vestibular input is both necessary and sufficient to generate the signal. In contrast to HD cell firing rates, other studies have demonstrated the importance that self-motion inputs (motor, proprioceptive feedback, motor efferent copy) serve in maintaining a stable PFD when the HD signal is updated as an animal locomotes into a novel environment (Stackman et al. 2003; Yoder et al. 2011).
The HD signal is thought to be generated within the connections between the dorsal tegmental and lateral mammillary nuclei (Bassett et al. 2007; Sharp et al. 2001a; Taube 2007). Key components for generating the HD signal are the AHV cells within these two nuclei. These cells fire as a function of the animal's AHV (Bassett and Taube 2001; Sharp et al. 2001b; Stackman and Taube 1998) and receive their primary inputs from the medial vestibular nucleus via the nucleus prepositus and supragenual nuclei (Biazoli et al. 2006). When an animal stops moving, vestibular inputs cease being modulated, as vestibular sensory input encodes changes in head movement, not head position. However, HD cells still maintain a consistent representation of the animal's current directional heading, even when the animal is not moving (Taube 1995; Taube et al. 1990a). These persistent HD cell responses indicated that the vestibular input may be signaling a correction or change in the animal's directional heading, but may not be continuously required for maintaining the signal when the animal is not turning its head (Taube and Bassett 2003). Thus how does the HD system maintain the current directional representation in the absence of changes in vestibular input? One possibility is that the tonic firing rates (5–20 spikes/s) of AHV cells within the dorsal tegmental nuclei when the animal is not moving continue to drive the HD cells (Bassett and Taube 2001). This view would be consistent with the observation that vestibular-only type I neurons within the vestibular nuclei also have non-zero background firing rates (about 40 spikes/s; Bush et al. 1993; Chen-Huang et al. 1997; Tomlinson and Robinson 1984). Alternatively, theories were developed that incorporated such properties of persistence into models explaining how the HD cell system functioned. In particular, these models contained an intrinsic attractor network (e.g., Knierim and Zhang 2012; Redish et al. 1996; Sharp et al. 2001a; Skaggs et al. 1995; Song and Wang 2005; Zhang 1996) or cellular mechanism within the HD cells (Yoshida and Hasselmo 2009), which could maintain the current state within the HD cells until AHV pushed the activity away from cells representing the previous directional heading and onto cells representing the current directional heading. This type of circuit could be dependent upon the vestibular head movement signal to update the HD response, but would not require input to maintain the signal when the animal is motionless. Furthermore, previous studies have examined the adaptation of a HD cell's firing rate over brief periods (<2 s) (Taube and Muller 1998), but no study has looked at adaptation properties over extended durations.
The purpose of this study was to assess the ability of the HD system to maintain a directional response over a period of time in the absence of motion by limiting the animal's movement through restraint. If HD cells decrease their firing rates in the absence of motion, then periodic sensory input is necessary to maintain a stable and robust HD signal. This property of HD cells is important to understand, as it may provide clues as to the types of inputs or intrinsic properties that sustain cell firing when an animal does not move, and maintains a stable directional heading. To test this hypothesis, we first rotated a head-fixed, body-restrained rat and then stopped and held it motionless for a period of time, while simultaneously recording HD cells from the anterodorsal thalamus (ADN). Passive head rotation during head-fixed restraint does not, by itself, impair HD cell responses, particularly in terms of peak firing rates (Shinder and Taube 2011). Here, we report that, when an animal is motionless, there is a marked and stable decrease in the cell's firing rate for directions within the cell's directional firing range and an increase for directions outside the cell's directional firing range. These findings demonstrate that the HD signal remains intact in the absence of motion, but with a reduced difference between peak and background firing rates.
METHODS
Animals.
Nine female Long-Evans rats were used in this study. Animals were individually housed, provided food and water ad libitum, and had a 12:12-h light-dark cycle. At the time of surgery, the animals were 161 ± 18 days old and weighed 285 ± 8 g. All procedures were approved by the institutional care and use committee at Dartmouth College, following the standards outlined in the National Institutes of Health Guide for the Care and Use of Laboratory Animals, as well as the guidelines provided by the Society for Neuroscience.
Surgery.
Each animal was implanted with a head bolt and an electrode array that was positioned above the ADN. The implant for electrophysiological recording has been described in detail elsewhere (Shinder and Taube 2011). Briefly, the head bolt was fixed to the head through polymer resin and skull screws. The implanted bolt coupled to an anchor that was mounted to the platform on which the animal was restrained, limiting head movement relative to the platform. Twenty nichrome wires (each wire was 25.4 μm in diameter, California Fine Wire) were threaded through a stainless steel cannula. This electrode array was then positioned relative to bregma using stereotaxic coordinates at 1.8 mm posterior, 1.3 mm lateral, and 3.7 mm ventral to the cortical surface.
All animals were acclimated to restraint prior to any electrophysiological recordings. Starting 1 wk following postsurgical recovery, the animals were restrained for 8 min by being held loosely in the experimenter's hands. This procedure was first done once a day for 3 days. Then, as electrophysiological screening began, the animal was restrained following the screening session for 8 min using a towel-wrapped restraint technique (Taube 1995; Golob et al. 1998). The towel-wrapped restraint acclimation was repeated once a day for 3 days. Following the subsequent screening session, the animal was placed in a restraint device with its head and body restrained for 8 min (Shinder and Taube 2011). The head-fixed restraint acclimation continued for at least 3 days before neurons were tested with the animal in the head-fixed restraint device. Following this procedure, animals tolerated restraint without significant periods of attempted movement. While restrained, bands were loosely placed around the body to hold the animal's fore- and hindlimbs. The rat could remove these bands by struggling against the restraint, but this behavior was rarely noted and resulted in the immediate cessation of experimentation.
Electrophysiological recordings occurred while the animal either foraged in an open cylinder (76 cm diameter, 50 cm tall) or was passively restrained in the same open cylinder. A cue card subtending ∼100° was mounted on the inside wall of the cylinder as a navigational landmark. This visual cue was visible to the animal in all experiments throughout the study. Symmetrical overhead lighting was provided by eight uniformly placed lights at the ceiling. The electrophysiological apparatus has been explained in detail elsewhere (Taube 1995). In brief, the restraint platform was not motorized and, when at rest, experienced no vibration. All movement of the head was detected with the use of two LEDs of different colors (1 red, 1 green) that were sampled at 60 Hz. Similarly, electrophysiological data were triggered, counted, and binned for each 1/60th s. During the braking studies, the electrophysiological data and the time stamps were recorded for each spike. Time stamps were later aligned in relation to the moment of braking.
All experimental sessions began with either an 8-min active foraging session or a 4- to 8-min passive rotation session in the cylinder, and most sessions contained both types. The passive rotation session was similar to that used and described in a previous study (Shinder and Taube 2011). Briefly, rats were wrapped in a cloth towel, restraint bands were placed over the cloth around both the forelimbs and hindlimbs, and the rat was placed in the cylindrical tube. Their heads were anchored to the restraint bar above their skull. The restraint device was mounted on a platform that could be rotated back and forth manually. A passive session consisted of rotating the rat back and forth at various speeds throughout a full 360°. Following this passive session, we performed one of three different protocols. [In a few cases (n = 6 out of 84 cases), we did not conduct a passive rotation session before commencing with one of the following protocols.]
Protocols: braking.
The braking protocol was designed to test the HD response while the animal was moving and motionless. Animals were head-fixed and had their bodies restrained within the circular tube that was mounted on a rotatable platform. By stopping the moving animal when it faced a particular direction, and maintaining it in a motionless state, the HD cell response was characterized for a given direction under passive motion and motionless conditions (Fig. 1A). A HD cell was initially identified and characterized while the animal actively foraged for food pellets. Then the HD cell's properties were monitored with continuous back-and-forth passive rotation in the horizontal plane, and a marker was placed on the outside of the enclosure that indicated the cell's PFD. The marker was not visible to the animal. Using this marker for the cell's PFD and other markers placed 90° around the enclosure from the PFD marker, the experimenter could position the rat to face into the cell's PFD or 90° away from the cell's PFD.
Fig. 1.
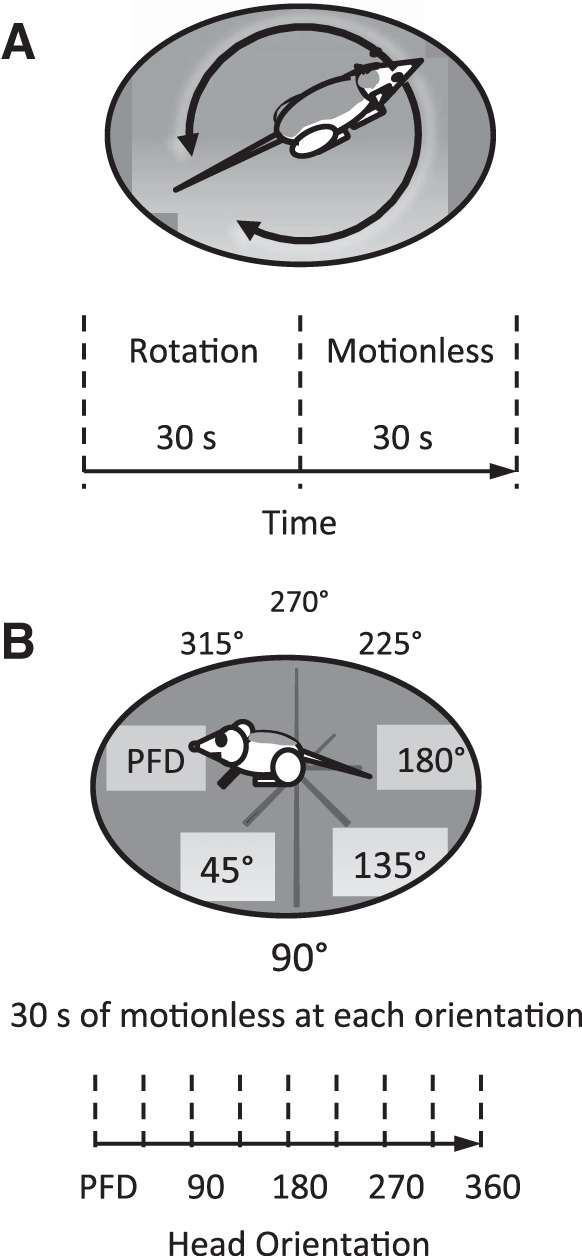
Two protocols were tested. A: braking protocol. The animal was rotated back and forth continuously in ∼300° sweeps and then stopped either at the cell's preferred firing direction (PFD) or 90° counterclockwise (CCW) to the cell's PFD. B: static placement protocol. The animal was placed in a stationary position for 30 s at eight equidistant directions around 360°, starting at the cell's PFD. At each position, the cell's response was recorded while the animal was in a motionless state. The major difference between the two protocols was in terms of the type of motion experienced by the rat prior to its static placement in a particular orientation. In the first protocol, the rat experienced 30 s of back-and-forth motion in the horizontal plane, while in the second protocol the animal experienced only a short 45° rotation over ∼1 s prior to placement at a static position.
We then characterized the difference between braking the animal while it faced the cell's PFD vs. braking the animal outside the cell's PFD. This comparison assessed whether motion was necessary for the maintenance of the HD cell response while the animal remained motionless, or if the HD cell's firing rate could be maintained at levels seen during passive rotation in the absence of movement. To characterize the HD cell's response to motionlessness, the animal was passively rotated back and forth continuously for 30 s inside the cylindrical enclosure, and then stopped for 30 s. The passive rotation used during the braking protocol was similar to the passive rotation session described above (Fig. 2A), with one exception: passive rotation during the relatively short 30-s braking protocol did not sample the full range of HDs in the environment. Rotation of the animal centered around the HD cell's PFD, but covered only ∼300° because there was a range of approximately ± 30° opposite the cell's PFD that was not sampled during the passive rotation. Following ∼30 s of back-and-forth passive rotation, the animal was stopped while facing either the cell's PFD (Figs. 1A and 2B) or 90° counterclockwise (CCW) to the cell's PFD. The selection of 90° away from the PFD stems from previous data indicating that the directional firing range for cells during passive rotation is ≤140° (±70° around the PFD; Shinder and Taube 2011; see also Table 1 for directional firing range of cells tested), and the mechanical inability of the apparatus to use 180° in the braking experiments. Thus, to avoid directions that may fall within the cell's directional firing range, we defined directions “not in the PFD” or “away from the PFD” as directions that were ≥90° away from the cell's PFD. Rotations were done by hand with the platform rotating just above the surface of the floor (1–5 mm). When the rotation was stopped, the platform rested motionless on the floor of the enclosure. The absence of any applied forces on the platform limited the possibility of imposed mechanical vibration during the periods of motionlessness. As shown in Fig. 2B, the last second just prior to motionlessness usually concluded with a final sweep from one side of the enclosure and ended with a small correction of the rotation aimed at positioning the animal to face the preplaced marker indicating the cell's PFD. Once stopped, recording continued while the animal sat motionless for 30 s. The sequence of 30 s of back-and-forth passive rotation followed by 30 s of static placement was repeated four times for each head orientation (either four times in the PFD, or four times 90° CCW to the PFD, with only one direction tested in any given HD cell). Finally, the animal was passively rotated back and forth in the same manner for an additional period to ensure that the directional response was still present. Rotational velocities ranged from 0 to 500°/s, with 95% of all rotation ≤245°/s (Fig. 2A). The velocity profile experienced during these rotations was similar to that produced previously, in which 95% of all passive rotation ≤275°/s, range: 0–495°/s (Shinder and Taube 2011). Fifty percent of the velocity experienced during the braking trials was >10°/s, since one-half of the time was spent motionless, and the other one-half was spent in passive rotation.
Fig. 2.
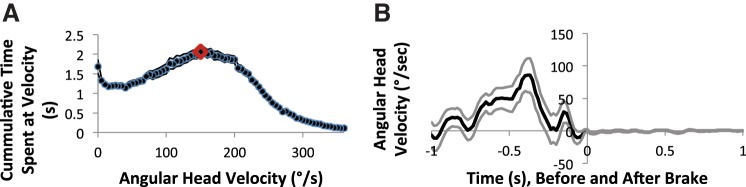
Angular head velocity profiles during rotation (A) and at the onset of motionlessness (B) experienced in the braking protocol. A: the amount of angular head velocity experienced during recordings of head direction (HD) cells during passive rotations in between periods of motionlessness (n = 41). The solid points represent the amount of time in seconds the rat spent rotating at a given angular head velocity, and the fine lines represent the standard error around each point. Head velocities were binned in 5°/s increments, and times were pooled by individual samples (1/60 s). Each recording period in between the four 30-s periods of motionlessness contained 90 s of rotation. The most time spent at any given rotational velocity was 2.1 s and occurred at 150°/s (red diamond). B: mean angular head velocity experienced immediately before and after the cessation (braking) of passive rotations. Each session was aligned to the time point of movement cessation for 163 trials across 41 cells. Angular head velocity was sampled at 60 Hz. The dark line represents the mean velocities, and the fine lines are the standard error around the mean.
Table 1.
HD cell firing rate responses across motion protocols
| Peak Firing Rate | Background* | Range | Rayleigh* | |
|---|---|---|---|---|
| Active | 56.0 ± 4.8 | 1.1 ± 0.2 | 117.7 ± 5.3 | 0.80 ± 0.01 |
| Passive | 55.3 ± 3.6 | 1.8 ± 0.3 | 132.4 ± 5.0 | 0.76 ± 0.02 |
| Braking passive | 67.0 ± 3.9 | 0.7 ± 0.1 | 123.7 ± 5.4 | 0.84 ± 0.01 |
| Mean Firing Rate Facing PFD | Mean Firing Rate Facing Away From PFD | |||
| Braking, % | 42.4 ± 3.2 (21.8 ± 2.3) | 14.0 ± 2.0 (9.5 ± 2.1) | ||
| Static with 1-s Interstimulus interval, % | 51.0 ± 3.5 (22.7 ± 2.0) | 11.8 ± 2.0 (4.6 ± 0.8) | ||
| Static with 30-s, Interstimulus interval, % | 69.0 ± 9.1 (16.7 ± 1.9) | 18.2 ± 3.4 (4.6 ± 0.8) |
Values are means ± SE. Firing rate responses across motion protocols are shown. Top: the firing rate responses of 40 head direction (HD) cells during rotation within the enclosure. Each cell was recorded during all three of the following conditions: active foraging, passive rotation, and passive rotation during the braking protocol. A multivariate ANOVA found significant differences in the background firing rate and Rayleigh values of those HD cells under the different recording conditions (
P < 0.001). Bottom: the mean response of HD cells when animals sat motionless for 30 s facing either the cell's preferred firing direction (PFD) or 90° away from the cell's PFD. The HD cell response is the mean firing rate during motionlessness as a percentage of the peak firing rate during passive rotation. The mean firing rates are shown in parentheses below the firing rate percentages. 90° away from the PFD was the direction sampled to define “away from PFD” for all three motionless conditions.
Static directional sampling.
The response during the braking protocol indicated that the HD cell's firing rate changed over the course of about 4 s as the rat remained motionless. This change occurred when the animal's head was stopped at the PFD or 90° CCW to the PFD. To further characterize the directional sensitivity during motionlessness following this unexpected response, we sampled HD cell responses using static placement of a restrained animal.
Following the initial recording of the HD cell in the active foraging and/or passive rotation session, the cell was recorded for 30 s while the animal was restrained in the restraint device and placed facing the cell's PFD (Fig. 1B). The animal remained motionless during the recording period and then was moved 45° CCW, and a second motionless session recorded for 30 s. This process was repeated until the animal was recorded in eight directions, 45° apart, spanning 360° of the enclosure. Each orientation of the rat contained unique visual views of the surrounding environment and theoretically enabled the rat to perceive its orientation. A final session repeated the first recording at the cell's PFD, to ensure the initial directional response was still present. Thus, we recorded a total of nine sessions at eight orientations, with the first and last sessions recorded facing the cell's PFD. Although the animal's orientation during the 45° rotational periods to the next position were not recorded, the rotational velocity was typically between 45 and 90°/s (which is above the vestibular threshold) for about 1 s. Directional rotation was consistently CCW. No overall asymmetry was noted in the HD responses reported below or in previous experiments (Blair and Sharp 1995; Shinder and Taube 2011; Taube 1995). Each time the animal was rotated in CCW steps, the direction at each static placement was normalized to the initial direction when the animal faced the cell's PFD. Thus the first direction in the series was always 0° when the animal faced the cell's PFD, and the second direction was 45°, and so on. The first and last directions tested were facing in the same direction after 360° of rotation and indicated that the peak in the firing rate response was maintained at the original 0° direction. Because all recordings were performed while the animal was motionless and because it was unlikely that stepping in the CW direction would have produced different results, the direction of rotation was not varied during the recording sessions.
Analysis.
For the braking experiments, peristimulus histograms of average firing rates binned at 10 Hz (100 ms) were computed from the spike time stamps. Determining the brake point by constructing histograms using 60-Hz bins would not be accurate because of jitter in the video signal due to video interlacing. Therefore, to determine the moment that movement ended, we computed the animal's AHV at 60 Hz. While the animal was rotated for 1 s prior to cessation of motion, the mean AHV (measured at 60 samples/s; 166 total recordings) was 25.3 ± 22.4°/s. However, while the animal sat motionless, the mean AHV for the first 1-s epoch of motionlessness on any given recording was −0.53 ± 1.67°/s. Therefore, the onset of the brake (the point at which head movement was stopped) was defined as the first 16.7-ms time point in which the AHV < 5°/s during the deceleration period. The measured AHV at this threshold was 0.77 ± 1.72°/s (Fig. 2B). Noise and other errors in the recorded signal that could have affected the measurement of the onset of the brake were reviewed and evaluated in each individual head velocity recording. Any epoch in which the time point of the brake could not be determined reliably was removed from analysis. Only 1 trial out of 166 trials across 41 cells was removed from analysis. Analyzed trials were 30 s in length and included 1 s prior to the brake point and 29 s following the brake point. All spike times were expressed relative to the time of the brake point. Therefore, a spike occurring at the brake point would have a 0-s time stamp, spikes occurring before the brake would have negative values, and positive values would reflect spikes occurring after the brake. The peristimulus firing rate was computed based upon histogram binning methods similar to those used for computing firing rate responses for longer recording sessions (Taube 1995). The spike times that occurred within a 16.6-ms bin were pooled to determine the firing rate for that histogram bin. Normalization of the cell's peristimulus firing rate histogram was performed by dividing each bin's firing rate by the peak firing rate determined from the 4-min session of continuous passive rotation (and not from the 30-s passive rotation periods within the braking protocol). Instantaneous firing rates were sometimes used to demonstrate the effects of motionlessness (Figs. 3–5) and were computed by taking the average of the inverse interspike interval before and after each spike time. Similar to the binned data, instantaneous firing rates were normalized by using the peak firing rate, which was determined from the passive rotation sessions. Peaks in firing rate were computed using filtered instantaneous firing rates. A sliding five-point average was repeated five times. A firing rate peak was detected if the firing rate at a given time point was higher than the firing rates of the three spike time points before and after the time point of interest. Interpeak intervals were the time periods between one peak and the next subsequent peak. The cell's background firing rate was defined as the mean of the firing rates for the remaining binned directions that were outside the cell's directional firing range (Taube et al. 1990a).
Fig. 3.

Top: the graph on the left displays the instantaneous firing rate of a representative HD cell during a single interruption (brake) of rotation when the animal was stopped facing the cell's PFD. Instantaneous firing rate is computed as the average of the inverse of the interspike interval before and after the time stamp of the current spike. Negative time points reflect the period of rotation prior to the brake, and positive times reflect the period after the brake when the animal was motionless. The instantaneous firing rate prior to the brake is depicted in black, from the brake to 1 s after the brake in red, from 1 to 4 s after the brake in green, and from 4 to 29 s after the brake in blue. The brown trace represents the instantaneous firing rates after being smoothed. The middle graph displays the four smoothed traces of instantaneous firing rate for each of the four braking trials performed on this cell. The brown trace is identical to that in the graph on the left. The other trials, in addition to the brown trace, are depicted in black, blue, and red. The graph on the right shows the directional tuning curves for this cell during active foraging (black) and passive rotation (green). Bottom: the firing rate of another cell for a single rotation and brake when the animal was stopped facing 90° away from the cell's PFD. Color traces are the same as in the top graphs.
Fig. 5.

A–D: individual trials and peak firing rates for a representative HD cell. Each graph displays the instantaneous firing rate with the time point of each detected peak in firing displayed above each graph (peak detection defined in methods). Each trial is colored differently and has different shaped markers. E: the intervals between firing rate peaks from the four trials are pooled from the data shown in A–D (black) and graphed as a histogram. The recorded interpeak intervals are displayed alongside interpeak interval averages taken from random shuffles of the recorded peak time points (red). The firing rate peaks, rather than the spike times, are shuffled to maintain an equivalent number of peaks. F: the histogram of interpeak interval values averaged across the HD cells following the onset of motionlessness while facing the HD cell's PFD in the braking protocol (n = 22). Again, the recorded data are shown in black, and the averaged interpeak interval computed from random shuffles of the recorded data is shown in red. G: the HD cell interpeak intervals for the animals that stopped while facing away from the recorded cell's PFD (n = 17, 2 HD cells had such low firing rates postbrake that firing rate peaks could not be detected).
To analyze the effect of firing rate adaptation and static placement of the animal on directional tuning, animals were serially placed in eight evenly distributed directions in the environment. The firing rate during 30 s of static placement for each orientation was compared with the cell's response during active exploration (8-min session) or passive rotation (4-min session) for the equivalent orientation. For each of the eight orientations analyzed, the firing rate was defined as the mean firing rate over five bins (±15°) centered on the sampled orientation. Because the peak firing rate occurs in only one bin, the average firing rate over the five bins that represent the cell's PFD will underestimate the true peak firing rate. Therefore, during active foraging sessions, the mean peak firing rate was similarly sampled over five bins (±15°) centered around the bin with the highest firing rate. The mean firing rate during motionlessness was also compared with the peak firing rate during rotation. The firing rate percentage was defined as the ratio of the mean firing rate during motionlessness divided by the peak firing rate during rotation. This percentage was averaged across cells in each condition and presented in Table 1.
Statistics.
Most statistical tests were done on mean firing rates sampled over 30 s between one condition and another condition. Therefore, we used noncircular statistics including t-tests and ANOVA tests comparing population means. The analysis of firing rate changes at the cessation of motion utilized linear regressions and ANOVAs of the firing rates at selected time intervals around the cessation of motion.
Histology.
Following experimentation, the animals were euthanized to process the brains for the localization of the recording electrode tracts. Prior to euthanization, electrolytic lesions (15 μA, 20 s) were made at the bottom of each tract. Following perfusion of the rat with 10% formalin, the brain was removed and allowed to sit in 10% formalin for at least 2 days. It was then placed in a 10% formalin + 2% potassium ferrocyanide solution, which enabled a Prussian blue reaction to occur and lead to visualization of the electrode tip in which current had been passed. The brains were placed in a 20% sucrose solution and later sectioned and mounted onto microscope slides. After staining with thionin, the sections were examined under a microscope to determine the lesion location and electrode tracks. All recording locations were confirmed to lie within the ADN.
RESULTS
The data are presented for two separate experiments. Some of the HD cells were recorded in both experiments. Forty-one HD cells were recorded in 4 animals in the “braking” experiments; 52 HD cells in 9 animals were recorded in the “static sampling of HD responses.” Of these, 20 HD cells were sampled in both experiments. A total of 55 HD cells were recorded in 9 animals for both experiments.
Braking.
To evaluate the influence of self-motion information on the HD cell signal, animals were passively rotated back and forth around the HD cell's PFD for ∼30 s and then stopped abruptly (Figs. 1A and 2B). We compared the cell's firing rate at time points just prior to when the animal stopped moving and following application of the brake. Twenty-two HD cells were recorded when the animal was stopped with its head pointing in the cell's PFD, and 19 HD cells were recorded when the animal was stopped while facing 90° away from the cell's PFD. The response of each cell was the average of four braking sessions (except for one cell in which there were only three repetitions). Each braking session was preceded by a back-and-forth passive rotation. Responses during the braking protocols were compared with the cell's characteristics determined from the 4-min passive rotation session (Table 1, top). Similar to previous findings, the responses during passive rotation were comparable to those seen during active foraging.
Representative examples for when the rat was stopped while facing into and away from the cell's PFD are shown in Fig. 3. For the cell at the top of the figure, a single trial is shown in the graph on the left in which the animal was stopped while facing the HD cell's PFD. The instantaneous firing rate prior to the brake was 49.8 spikes/s (mean response for the trace shown in black), and 40.2 spikes/s for the 100-ms period just prior to the brake. For the first 100 ms after the brake, the cell's firing rate was 36.9 spikes/s, and from the brake to 1 s after the brake the firing rate was 44.8 spikes/s (red). For the 1- to 4-s period after the brake (green trace), the mean firing rate decreased to 32.7 spikes/s (66% of the cell's peak firing rate). Thus the cell's firing rate was ∼40 spikes/s as the animal moved to face the PFD and decreased to ∼30 spikes/s following the cessation of motion. From 4 to 29 s postbrake (blue), the cell's firing rate averaged 26.9 spikes/s and was highly variable over time. Over this period, the cell's firing rate averaged 54% of its peak firing rate, despite the fact that the animal was continually facing the cell's PFD. This pattern of activity, with moments of increased or decreased firing, was observed across the plateau period for all cells. The middle graph at the top of Fig. 3 displays the smoothed instantaneous firing rate traces for all four braking trials and shows that the variations in firing rate over the plateau period were not consistent from trial to trial in the same cell. All other HD cells had similar variable firing rate patterns in the plateau phase from 4 to 29 s.
Figure 4A and Tables 1 and 2 depict the mean response across all 22 HD cells. Firing rate histograms plot the cell's response starting 1 s prior to cessation of motion to 29 s following termination of motion. For the mean response, each HD cell's firing rate histogram was averaged across four braking sessions and normalized based on the cell's peak firing rate during the 4-min session of passive rotation that was recorded prior to the braking protocol. The normalized histograms were then averaged across the population (22 HD cells; Fig. 4A). Cessation of movement had a marginal effect on HD cell firing rates during the first 100 ms after braking, with no significant difference between the HD cell's peak firing rate measured during passive rotation and the HD cell's firing rate 100 ms before or after the brake [Table 2, ANOVA: F(2,65) = 1.36, P = 0.27]. As with the representative cells in Fig. 3, following this initial 100-ms postbrake period, there was a gradual, but steady decrease in the cell's firing rate over the next 4 s, after which the firing rate leveled off for the remaining 25 s. The mean slopes for these two epochs were −10.73 ± 1.11 and −0.11 ± 0.11% of peak firing rate/s, respectively. The decreased firing rate plateaued at 42.4 ± 3.2% of the peak firing rate for the passive rotation session (Tables 1, bottom, and 2). The plateau in firing rate was on average about 40% of the cell's peak firing rate, but the reduction in firing rate for any given HD cell could range from about 10 to 70% (Fig. 4, bottom histogram). However, while there was substantial variability in the firing rate for any given HD cell from 5 to 29 s after cessation of movement (see Figs. 3 and 5, A–D), there was no apparent systematic drift in the mean firing rate during the plateau period. The firing rate peaks during each trial for an individual cell do not follow a particular pattern, but rather appear to be distributed randomly. Analyses of the time intervals between firing rate peaks supported this view [Fig. 5, E (example cell) and F (population of HD cells)].
Fig. 4.

The graphs display histograms of the mean firing rates of HD cells, normalized by the cell's peak firing rate measured during passive rotation, and binned in 100-ms increments. Time zero is the point at which the animal stopped moving. Negative times represent when the animal was passively rotating back and forth, and positive times represent the period when the animal was statically facing a fixed direction. The graphs in the top row represent the average binned firing rate response from 1 s prior to 29 s after the brake. The graphs in the middle row represent the average binned firing rate and the instantaneous firing rates from 100 ms before to 100 ms after the brake. The mean of the binned firing rates is represented by the black line in the top two graphs; the standard error is shown by gray shading around the line, and individual spikes are black circles (defined by the spike time and the average of the inverse interspike interval before and after that spike, divided by the HD cell's peak firing rate). A: sessions when the animal was stopped while facing the cell's PFD (n = 22 cells). B: sessions when the animal was stopped facing 90° CCW away from the cell's PFD (n = 19, different cells than in A). Bar graphs below are population histograms showing the mean HD cell responses during the steady state period (4–29 s) contributing to the graphs above. The population histograms display each HD cell's mean steady-state firing rate as a percentage of the cell's peak firing rate measured during passive rotation.
Table 2.
Firing rates before and after a brake in motion
| −0.1–0 s | 0–0.1 s | 0–1 s | 5–29 s | |
|---|---|---|---|---|
| Facing towards the PFD | 92.0 ± 6.0 | 87.1 ± 6.0 | 74.4 ± 4.8* | 42.4 ± 3.4* |
| Facing away from the PFD | 3.6 ± 1.3 | 1.4 ± 0.5 | 3.7 ± 0.7 | 14.0 ± 2.0* |
Values are means ± SE in %. Firing rates before and after a brake in motion are shown. HD cell normalized mean firing rates for specified epochs prior to or after a brake in motion. Firing rates are normalized and presented as a percentage of the peak firing rate of the previous passive session. The firing rates 100 ms before the brake and after the brake were not significantly different, but firing rates over a period of 1 s following the brake differed from the firing rates 100 ms prior to the brake when the animal was stopped while facing into the cell's PFD (ANOVA,
P = 0.03). In addition, the mean firing rate over the period from 5 to 29 s after the brake differed from the response 100 ms before the brake when facing both toward and away from the cell's PFD (ANOVA,
P < 0.001).
A different set of cells from those described above was tested under similar conditions, but the animal was stopped while facing 90° away from the cell's PFD (n = 19) (Figs. 3 and 4B; Table 2). This procedure was conducted to determine whether there was any change in the cell's background firing rate as a result of braking and the animal remaining motionless. In the representative response at the bottom of Fig. 3, a single braking trial is shown in the graph on the left in which the animal was stopped facing 90° away from the cell's PFD. This cell's mean firing rate prior to the brake was 87.4 ± 8.6 spikes/s (black), but was 0.0 ± 0.0 spikes/s over 100 ms before the brake. The firing rate over the first 100 ms after the brake remained at zero, and the mean firing rate over 1 s postbrake was 1.3 spikes/s (red); this rate was produced by a single spike occurring 0.99 s after the onset of the brake. However, from 1 to 4 s after the brake, the mean firing rate increased to 29.4 ± 1.9 spikes/s (green) and increased further to 38.3 ± 0.6 spikes/s (blue) from 4 to 29 s. Similar to the response above, there was no consistent pattern in the firing rate fluctuations during the plateau phase within a trial or across trials in the same cell (Fig. 3, bottom center). Across the population of cells tested, the mean background firing rate during passive rotation was 1.83 ± 0.32 spikes/s (Table 1, top). In comparison, the mean firing rate when the animal was 90° away from the cell's PFD at 100 ms prebrake was 1.97 ± 0.68 spikes/s, and was 1.05 ± 0.40 spikes/s at 100 ms postbrake. These pre- and postbrake firing rate values were not significantly different from the cell's background firing rate during the passive rotation session [Table 2, ANOVA of the background firing rate and the mean normalized firing rates 100 ms before and after braking: F(2,56) = 1.34, P = 0.27]. Again, however, there was an initial transition period that lasted about 4 s, in which the cells' firing rates increased steadily before leveling off (Fig. 4B). The slope from braking to 4 s was 3.22 ± 0.76% of peak firing rate/s or 2.33 ± 0.66 spikes/s2, and the slope for the last 24 s was −0.03 ± 0.03% of peak firing rate/s or −0.02 ± 0.02 spikes/s2. The mean firing rate from 5 to 29 s postbrake was 9.47 ± 2.05 spikes/s or 14.0 ± 2.0% of the cells' peak firing rates. Note, however, that this firing rate was 697 ± 162% of the cells' mean background firing rates during the passive rotation, which was on average 1.8 spikes/s (Table 1). Similar to the plateau in firing when the animal was stopped facing the cell's PFD, the plateau in the cell's firing rate varied over time (Fig. 3) and across cells when the animal was stopped facing away from the cell's PFD (Figs. 4B, bottom histogram, and 5G). The steady-state firing rate from 5 to 29 s after braking ranged from 1 to 33% of the cell's peak firing rate, or from 33 to 2,060% of the HD cell's background firing rate, with no apparent systematic changes in the firing rate after the initial 4-s increase in firing (Figs. 3, 4A, and 5, A–E). Interestingly, the number of firing rate peaks was much lower when facing away from the PFD than when facing into the PFD (Fig. 5, F and G). This result may be due to the lower firing rate when facing away from the PFD, but was not indicative of an active signal resonating throughout the HD circuit during motionlessness.
The firing rate levels during the plateau phase vary over a wide range. The firing rate response of HD cells does not change at the onset of the brake, and, to define this variability as an independent measure, the initial raw firing rates can be compared trial by trial to the later plateaued rate in that trial. When the animal was stopped facing into the HD cell's PFD, the relationship of the raw firing rates 100 ms before and after the brake was well-defined (Pearson's r = 0.772, slope = 0.784, R2 = 0.595). However, the relationship between peri-brake firing rates and the resulting average firing rate over the plateau period was not as clearly defined (100 ms before to plateau: r = 0.448, slope = 0.219, R2 = 0.201; 100 ms after brake to plateau: r = 0.531, slope = 0.264, R2 = 0.282). Thus the relative decrease in the firing rate from peak levels prior to the brake was not clearly predicted by the cell's peak firing rate. When the animal was stopped facing away from the HD cell's PFD, firing rates were very low (Fig. 4B, note only 24 spikes occurring over 200 ms across all 19 cells in the center graph). Thus trial-by-trial HD cell firing rates around the brake period were often zero, regardless of the cell's peak firing rate; therefore, a regression analysis could not be performed. Another possibility explaining the variability in plateau firing rates is that the range of firing rate levels during the plateau phase could be an effect of the amount of experience each animal had with the braking protocol. However, HD cells were recorded over 5 mo in the paradigm, and there was no indication for a relationship between experience and the amount the firing rate response plateaued [n = 4 rats; the HD cell's average normalized plateau firing rate across rats for the first HD cell recorded = 0.47 ± 0.06, for the second HD cell recorded = 0.44 ± 0.09, for the third HD cell recorded = 0.42 ± 0.03; ANOVA for effect of the number of cells found previously: F(2,10) = 0.112, P = 0.89].
In summary, during the braking protocol, cell firing rates behaved similarly during the passive rotation phase as in previous experiments [ANOVA of peak firing rate for active, passive, and passive braking protocols: F(2,119) = 2.51, P = 0.09; Table 1, top], but when the animal was stopped at the cells' PFDs, the firing rates decreased to about one-half of the normal peak firing rate. When the animals were stopped at orientations outside the cells' PFDs, the firing rates increased markedly above the normal background firing rate. The change in firing rates was not immediate, as the firing rates 100 ms following the brake did not differ from those 100 ms prior to the brake (Fig. 4). However, by the end of the first second following the brake, the firing rates were significantly shifted from those 100 ms prior to the brake (Fig. 4, Table 2). Thus the process initiated by the cessation of motion appeared to progressively affect the cell's firing rates until a plateau value was reached. It took about 4 s after the brake was applied for these firing rates to stabilize at their new values.
Static directional sampling.
Previous studies on HD cells have recorded them while the animal is usually moving around continuously, and there has been an absence of studying their properties when the animal remains motionless for relatively long time periods, such as longer than 2 s. To further address the properties of firing adaptation in more detail, we tested animals that were statically placed for 30 s at a time, facing eight different selected directions. The eight directions were in 45° increments ranging from 0 to 360° (Figs. 2B and 6). The visual landmark within the enclosure was present the entire time and was clearly viewable by the animal at each tested position. Prior to static testing, the cells were first recorded in an active, freely-moving session for 8 min and then in a 4- to 8-min passive rotation session that sampled the full range of HDs across 360°. Then each cell was recorded as the restrained animal was serially placed in each of the eight different directions (Fig. 6). For these restrained sessions, the cell was first recorded with the animal's head facing the cell's PFD. Then the animal was rotated (∼30–90°/s) to each of the other seven orientations and recorded again while the animal remained motionless for 30 s. The animal was then rotated a final 45°, which brought it back to the “starting orientation,” and on a final ninth session with the animal motionless for 30 s (green lines in Fig. 6).
Fig. 6.

Firing rates for each recording period for a representative HD cell during the static placement protocol. The cell's normalized firing rate responses over time are shown in the outer graphs for when the rat was facing motionless for 30 s in eight evenly spaced directions, starting and ending in the direction of the cell's PFD (0 and 360°). Each recording was started after the animal had been motionless for several seconds. The protocol used a short 1- to 2-s rotation in between each placement. The black circles represent the mean firing rate over time, and the gray horizontal line represents the mean firing rate over the entire 30-s period. The black markers and black line in the 0° firing rate graph denote when the animal faced into the cell's PFD at the first placement, and the red triangles and red line represent the second placement. The red and black lines overlap as the initial mean rate for this cell = 16.2 spikes/s, and repeating the orientation in the same direction lead to a mean rate = 16.4 spikes/s. The radial plot in the center represents this cell's average firing rate during active foraging, but realigned such that the PFD was shifted to 0°. The tick marks represent 20 spikes/s. The cell's peak firing rate was 27.4 spikes/s and the directional firing range was 118.3°. The graph plotted in the inset depicts the HD cell's tuning curve during an active foraging session (HD is plotted relative to the environment).
Thirty HD cells were tested in the above paradigm, and their responses were compared with the active foraging session and to responses in the 4- to 8-min passive rotation session (n = 29). In general, responses remained stable and similar between the first and last static sessions when the animal faced the recorded cell's PFD (correlation of peak firing rate measured at 0° and 360° = 0.82; slope = 0.89, P < 0.01, R2 = 0.68; t-test, P = 0.69). HD cell firing rates in the static sampling protocol were similar to the braking experiments reported above. There were similar responses in both protocols when the animals faced the cell's PFD (Table 1, bottom: braking and motionless directional placements with 1-s intervals between placements), and equivalent responses across protocols when they faced 90° away from the cell's PFD [ANOVA effect of condition: braking, the two directionally facing motionless cell groups with 1-s intervals between periods of motionlessness: F(2,158) = 0.65, P = 0.52; ANOVA effect for direction: facing toward, away from PFD: F(1,158) = 185.82, P < 0.0001; no significant interaction effect]. Static placement of the animal at the cell's PFD reduced firing rates to 49.6 ± 4.0% of peak values seen during active movement (n = 30). The firing rate at the PFD was reduced to 51.0 ± 3.5% compared with the peak firing rate response during the passive rotation session (Table 1, n = 29). In addition, elevated firing rates occurred in the static placement sessions when facing away from the cell's PFD, consistent with the braking experiments reported above (Fig. 7). The firing rate during the statically sampled response for the sampled directions that were not in the cell's PFD (defined as directions ≥90° away from the cell's PFD) was 10.7 ± 1.6% of the peak firing rate and 568 ± 110% of the background firing rate observed during active foraging sessions, and 9.5 ± 1.6% of the peak firing rate and 298 ± 49% of the background firing rate observed during passive rotation sessions (Fig. 7, red and green traces, respectively). The reduction in the firing rate difference between the PFD and “off-PFD” directions indicates that there is a decrease in the signal-to-noise ratio, such that any individual spike would be less predictive of the animal's HD.
Fig. 7.
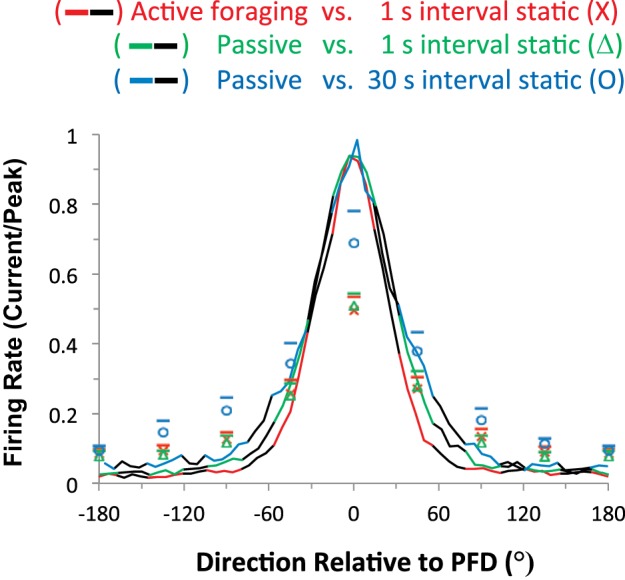
Mean responses of 30 HD cells normalized by peak firing rate recorded while the animal actively foraged in the environment (black and red line) and when the animal was passively restrained and placed statically facing selected directions in the environment (red x's represent the mean firing rate of the static session compared with the active foraging session). The ordinate plots the ratio of the firing rate for the given parameter condition compared with the cell's peak firing rate. The range of mean responses of the active foraging firing rate that were selected for comparison to the statically placed firing rate response are depicted by the red portions of the red and black line. All directions are relative to the HD cell's PFD (0 and 360° face into the cell's PFD, and 180° faces away from the cell's PFD in the opposite direction). The green and black line depicts the mean normalized firing rate responses of 29 HD cells that were recorded when the animal was passively restrained and rotated back and forth continuously through all directions. The green triangles represent the mean firing rate when the restrained animal was placed facing selected directions. The animal was moved directly from one orientation to the next without any intervening back-and-forth movements, as was done in the comparison to active foraging. As before, selected segments of the passive rotation response that were used for comparison to the static placements are shown by the green portions of the green and black line, and all directions are relative to the cell's PFD. The blue and black line represents the mean normalized firing rate responses of 18 HD cells under passive conditions in which the animals were rotated back and forth for 30 s in between static placements. The sampling periods of the rotation means used for statistical analyses are shown by the blue portions of the blue and black line, and the blue circles represent the mean firing rate while the animal was facing each direction. The bars above each marker represent the SEM.
It was possible that the single 1- to 2-s rotation of the animal to statically place it into position may not have allowed for sufficient accumulation of self-motion and visual information to enable accurate updating of the HD signal and represent the animal's current HD. Therefore, the passive rotation period was increased in between each static placement. We recorded 18 new HD cells while the animal was rotated back and forth passively, covering 360° over a 30-s period in between each static placement. The periods of motionlessness spanned the same eight HDs used in the static protocol above. Mean firing rates were normalized based on the cell's peak firing rate during the 4-min passive rotation session. Motionlessness still reduced the normalized mean peak firing rate when the rat faced the cell's PFD, as the mean firing rate in the static session was 69.0 ± 9.1% of the active peak firing rate (Fig. 7, blue trace). This value, however, is not reduced as much as the values found in the braking (42.4%) and static directional sampling (49.6%) sessions.
When the animal's head was facing away from the cell's PFD, the mean firing rate was 925 ± 283% of the background firing rate during passive rotation. Again, this increased rate compares to the rates in the braking protocol (697 ± 162%) and the directionally sampled motionless protocol with 1-s intervals between placements (298 ± 49% during passive rotation; 568 ± 110% during active session). The effect of the extended period of rotation in between static placements appears to be an increase in the firing rate at both the cell's PFD and away from the cell's PFD [Fig. 7; ANOVA comparing firing rates when facing the cell's PFD or 90° away in the static placement protocols: effect of direction, facing toward or away from PFD: F(1,153) = 158.56, P < 0.001; effect for the protocol used: F(2,153) = 5.00, P = 0.008; interaction effect of direction by protocol was not significant]. The increased firing rate appears as a positive bias in all directions without a significant effect on the separation between the peak and background firing rates. The origin of the bias in the motionless response produced by the 30 s of rotation in between motionless positioning is currently unknown. Braking data were not included in this analysis because the same cells were not used to test both HDs (PFD and 90° away from the PFD). In summary, there appears to be a small increase in the firing rates during the static placements due to the 30 s of passive rotation in between each placement.
DISCUSSION
To better understand the role that self-motion cues play in the HD cell signal, HD cells were monitored in the absence of self-motion. We have previously demonstrated that vestibular sensory information is necessary (Stackman and Taube 1997) and sufficient (Shinder and Taube 2011) for the generation of the HD signal. However, other self-motion cues, including visual, proprioceptive, and motor efference copy, are also important for controlling the alignment of the HD response with the environmental landmarks (Stackman et al. 2003; Yoder et al. 2011). Yet little is known about how self-motion information ultimately produces changes in HD cell firing rates, or the extent to which sensory cues interact in that process. Previous studies have noted that HD cells continue to fire while the animal is motionless, but they have also shown that HD cells contain a small, but significant, relationship with AHV, where higher angular head velocities are associated with higher firing rates (Blair and Sharp 1995; Taube 1995; Taube et al. 1990a). However, these studies did not assess the firing rate response to cessation of movement or to changes in directional tuning while the animal remained motionless for periods longer than 1–2 s. By developing a method in which HD cells could be recorded while the animal was restrained (Shinder and Taube 2011), passive restraint could be applied without impairing HD cell activity. The current findings indicate that the loss of self-motion input when head rotation is interrupted has no immediate or sizeable effect on the cell's response. However, from ∼100 ms after the head stopped moving, cell firing rates changed steadily for about 4 s and then stabilized. This change in firing rates occurred, despite the presence of a salient visible visual cue that could be used as a stable reference point. When the head was stopped facing the cell's PFD, firing rates decreased over 4 s, and, when the head was stopped facing away from the cell's PFD, cell firing rates increased over a similar time period. Importantly, the firing rate changed gradually toward a plateau value that was different for the in-PFD and out-of-PFD conditions. Thus, in both cases, although the HD cells remained directionally tuned (Fig. 7), the signal-to-noise ratio was reduced (Fig. 4).
One issue to consider that would result in this response to motionless is that the cell's PFD drifts or shifts away from its initial orientation following the onset of motionlessness. There are four possibilities as to how a cell's PFD could change over time during the motionless period: 1) the PFD drifts continuously at a slow rate, 2) the PFD drifts continuously at a fast rate, 3) the PFD drifts discontinuously (i.e., at different rates), and 4) the PFD shifts to a new orientation which is maintained across the motionless period. We now discuss each of these possibilities in order. First, if the PFD drifted at a slow rate away from the cell's initial PFD, then we would expect the cell's firing rate to slowly decrease throughout the motionless period to a point where the cell's firing rate reaches the background firing rate. However, the results showed that, when the animal was facing in the direction of the HD cell's PFD, the firing rate decreased progressively to a stable state, but never reached the background firing rate level. The steady-state (plateau) level was always well above the cell's background firing; indeed, although the mean firing rate during the plateau phase was decreased on average by 42%, it was 3,490% above the cell's background firing rate compared with the passive rotation session and 302% above the cell's firing rate during the plateau phase when the animal was positioned facing away from the cell's initial PFD. Thus a decreased firing rate during the motionless period is unlikely to be attributed to a slowly drifting PFD. Second, if the PFD drifted continuously at a fast rate, then we would expect periodic bursts of firing as the animal perceives its directional orientation passing through the cell's PFD. This possibility is unlikely because the analyses of high firing rate bursts during the motionless period (Fig. 5) showed that they were not periodic, but occurred randomly. Furthermore, there were far fewer bursts when the animal faced away from the PFD than when the animal faced into the cell's PFD. Thus, if the PFD was continuously drifting, the peak should drift through all directions equivalently, and a similar number of bursts would have been observed for directions outside the cell's PFD. Third, the PFD could be drifting or fluctuating over a confined range near the PFD. This possibility is what we might predict if the inhibitory control over the interconnection of HD cells is reduced. The number of bursts predicted when the animal faced away from the PFD would be one-half of the number seen when facing into the PFD. The prediction assumes the PFD drifts in each direction alternately. While the number of peaks seen in Fig. 5, F and G, is less than would be predicted, the prediction does not account for short drifts that do not extend far enough to influence firing rates away from the PFD. Furthermore, the amount of fluctuation in the PFD would be determined by the amount of reduction in inhibition on surrounding HD cells. This reduced inhibitory drive would affect the final plateau firing rate level without being influenced by the initial firing rates at the cessation of motion, in parallel with the observation above. Fourth, if the PFD shifted to a new stable orientation that was not at the cell's initial orientation, but still within its initial directional firing range, then we would expect the cell's firing rate to stabilize at a new reduced value, which is precisely what we observed. However, if this situation were to occur, then we would expect the cell's firing rate to remain at or near zero at directions outside the cell's directional firing range, unless the animal now perceived its orientation to fall within it. Background firing rates, although elevated a little during the motionless period, did not change for all directions outside the cell's directional firing range, suggesting that the cell's PFD remained relatively stable. Moreover, a stable new orientation cannot explain the variability in the plateau firing rates and cannot explain the arrhythmic bursting produced during the plateau phase. Furthermore, as mentioned above, the prominent and stable visual landmark was clearly visible throughout the motionless period. Given the salience of the cue card to control the cell's PFD (Goodridge and Taube 1995; Taube et al. 1990b), it is unlikely that the cell's PFD would have shifted to a new orientation. Finally, the results of experiment 2, where the same cell was tested at different orientations in 45° steps, did not indicate that there was a significant shift in the cell's PFD (orientations at 45 and 315° did not demonstrate firing rate activity indicative of a peak in firing rate; Figs. 6 and 7). In summary, it is unlikely that the cell's PFD was drifting slowly or shifting to a new orientation during the motionless period.
A previous study explored the extent to which HD cell firing adapts when the animal maintains the same directional heading over a period of time under unrestrained, freely-moving conditions (Taube and Muller 1998). The study analyzed episodes in which the animal's directional heading was maintained within a ±6° arc around the recorded cell's PFD for a minimum of 0.333 s. Episode times varied from 0.333 to 2 s, depending on the animal's behavior. The authors reported that HD cells in ADN, but not in the postsubiculum, displayed a small, but significant, adaptation over these short time periods (∼11% decrease from the cell's peak firing rate). In contrast, no adaptation was observed when a second set of recording sessions was analyzed for the same ADN and postsubicular cells, and the authors concluded that there was not strong evidence for the presence of significant adaptation. However, the relatively short time periods over which adaptation effects were searched for may have precluded the observation of any adaptation effects, since we found that the reduction in firing rate did not plateau until 4 s after the rat entered the cell's PFD. Although the animals in the previous study were freely moving, as opposed to head-fixed and restrained in the current experiments, we feel this explanation is unlikely to account for the differences, because passive rotation under the restrained conditions employed was not found to affect cell firing rates substantially over short time periods (Shinder and Taube 2011). However, as noted in that previous study, other restraint methods have led to differences in HD cell firing rates following restraint (Knierim et al. 1995; Taube 1995; Taube et al. 1990b; Zugaro et al. 2001, 2002), and this effect may also impact the HD response to motionlessness. In summary, adaptation effects do occur in HD cells, but they are only observed when looking at time periods of motionlessness that are longer than 1 s.
The change in firing rate that is initiated by cessation of motion takes ∼4 s to stabilize, whether the animal is facing into the HD cell's PFD or away from it. Therefore, in the absence of movement, background firing rates increase for cells whose PFDs are not in the animal's current direction, and peak firing rates decrease for cells representing the animal's current heading. At the cessation of movement, the HD cell response stabilizes progressively and slowly. It is possible that this change in firing could be due to a progressive decrease in the AHV response following the brake in motion. There is a postrotary vestibular response that extends neuronal firing beyond the cessation of head movement, which is found in AHV signals within the vestibular nuclei (Yokota et al. 1992). While the postrotary, or velocity storage effect seen in the vestibular signal indicates that the AHV signal could have lingering postmotion effects, the responses that extend seconds past the cessation of motion require continuous rotational stimuli, which were not used in these experiments (Raphan et al. 1979). Alternatively, the progressive change in HD cell firing following motionlessness could be due to intrinsic feedback mechanisms, similar to those associated with attractor networks (Knierim and Zhang 2012; Redish et al. 1996; Sharp et al. 2001; Skaggs et al. 1995; Song and Wang 2005; Zhang 1996). Normally, the firing rate of a HD cell would represent the contributions of both the intrinsic feedback circuit and the AHV input. When the AHV signal is removed during motionless periods, the firing rate of the HD cell would be reduced to a level that is sustainable by the intrinsic feedback circuit alone. Intrinsic mechanisms could also explain the variability in the plateau phase if individual cells differed by the strength of inhibitory connectivity controlling each cell, and relaxing that inhibition occurred on similar time scales with differing magnitudes.
As all animals, including humans, pause in between movements, the striking finding of this study is the large difference in the performance of the HD system when self-motion information is not available to maintain the HD signal. The difference in the response may not be behaviorally relevant because the HD system output still represents the animal's current directional heading until the animal moves again. Normally, the environment does not change around the animal. Thus any deficiencies in maintaining the alignment of the HD response with the environment would not be perceived by the animal. However, these conditions are now commonly found in virtual environments, and these virtual environments provide a means to evaluate the spatial knowledge that is acquired and utilized when the HD system is functioning without movement. Remaining motionless while exploring a virtual environment results in errors of perceived spatial orientation compared with exploring the same environment with active movement (Chance et al. 1998; Klatzky et al. 1998) and reduces the efficiency of the search (Ruddle and Lessels 2006). However, virtual spatial learning produces similar, although not identical, spatial knowledge about the environment (Williams et al. 2007). For example, subjects were capable of performing a series of spatial tasks (execution of turns, reproduction of distances traveled, triangle completion task) when only visual cues (optic flow) were present in a virtual environment, but their performance did not attain the level that occurred when self-motion cues were available (Riecke et al. 2002). The difficulty that is created by learning spatial information about an environment while remaining motionless is that it subsequently makes it more difficult to navigate accurately in that environment from different orientations. The absence of self-motion thus impairs our normal orientation invariance: the ability to be aware of the spatial relationships of an environment from any orientation (Montello et al. 2004; Peruch et al. 1995; Richardson et al. 1999; Sun et al. 2004). The HD system may normally contribute to the orientation invariance of spatial memories, defining the spatial relationships of objects and landmarks independently of the orientation in which they were experienced. It is also possible that the HD cell network influences these spatial relationships by connecting related spatial views to create a contiguous spatial representation of the environment. Not all spatial representations in the brain may be affected by the impairments of the HD system imposed by motionlessness, since hippocampal place cells that represent our current place in the environment are capable of representing location information in a changing virtual environment without actual self-motion (Harvey et al. 2009). However, the present findings suggest that the HD system helps the brain to define spatial relationships independently of the orientation in which they were experienced, such that spatial learning becomes impaired when self-motion input to the HD pathway is disrupted (Baek et al. 2010; Russell et al. 2003; Wallace et al. 2002).
In summary, the HD system displays evidence of reduced information, or signal-to-noise content, when an animal is motionlessness. The firing rates decrease for cells representing the animal's current heading and increase for cells that do not represent the animal's current heading. Under normal conditions seen in everyday life, there would be little pressure for the brain to do more than maintain a representation of the last directional heading until a self-movement signal was detected to reestablish normal HD responding. However, this mechanism of HD cell updating fails to equivalently represent directional heading during movement and motionlessness. Therefore, without continuous input from vestibular self-motion cues, the HD system may not maintain the current orientation in the environment as reliably as when these cues are present. This finding opens the possibility to differences in the information conveyed by the HD system when environmental (landmark) and self-motion cues are in conflict, particularly when self-motion cues indicate no change in directional heading, and landmark cues indicate a change in orientation. Such conditions are seen with increasing frequency and include virtual reality, vestibular lesion or dysfunction, and microgravity. Maintaining some level of sensory self-motion stimulation related to the HD system, while experiencing changes in external landmark cues, may improve HD signal integrity, as well as the spatial and navigational perceptual functions associated with HD cell functioning (Valerio and Taube 2012).
GRANTS
This work was supported by National Institutes of Health Grants NS-053907 and DC-009318.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: M.E.S. and J.S.T. conception and design of research; M.E.S. performed experiments; M.E.S. analyzed data; M.E.S. and J.S.T. interpreted results of experiments; M.E.S. and J.S.T. prepared figures; M.E.S. drafted manuscript; M.E.S. and J.S.T. edited and revised manuscript; M.E.S. and J.S.T. approved final version of manuscript.
ACKNOWLEDGMENTS
Present address of M. E. Shinder: Department of Neuroscience, Baylor College of Medicine, One Baylor Plaza, BCM 295, Houston, TX 77030.
REFERENCES
- Baek JH, Zheng Y, Darlington CL, Smith PF. Evidence that spatial memory deficits following bilateral vestibular deafferentation in rats are probably permanent. Neurobiol Learn Mem 94: 402–413, 2010 [DOI] [PubMed] [Google Scholar]
- Bassett JP, Taube JS. Neural correlates for angular head velocity in the rat dorsal tegmental nucleus. J Neurosci 21: 5740–5751, 2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassett JP, Tullman ML, Taube JS. Lesions of the tegmento-mammillary circuit in the head direction system disrupts the head direction signal in the anterior thalamus. J Neurosci 27: 7564–7577, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biazoli CE, Jr, Goto M, Campos AM, Canteras NS. The supragenual nucleus: a putative relay station for ascending vestibular signs to head direction cells. Brain Res 1094: 138–148, 2006 [DOI] [PubMed] [Google Scholar]
- Blair HT, Sharp PE. Anticipatory head direction signals in anterior thalamus: evidence for a thalamocortical circuit that integrates angular head motion to compute head direction. J Neurosci 15: 6260–6270, 1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bush GA, Perachio AA, Angelaki DE. Encoding of head acceleration in vestibular neurons. I. Spatiotemporal response properties to linear acceleration. J Neurophysiol 69: 2039–2055, 1993 [DOI] [PubMed] [Google Scholar]
- Chance SS, Gaunet F, Beall AC, Loomis JM. Locomotion mode affects the updating of objects encountered during travel: the contribution of vestibular and proprioceptive inputs to path integration. Presence (Camb) 7: 1678–178, 1998 [Google Scholar]
- Chen-Huang C, McCrea RA, Goldberg JM. Contributions of regularly and irregularly discharging vestibular-nerve inputs to the discharge of central vestibular neurons in the alert squirrel monkey. Exp Brain Res 114: 405–422, 1997 [DOI] [PubMed] [Google Scholar]
- Golob EJ, Wolk DA, Taube JS. Recordings of postsubiculum head direction cells following lesions of the laterodorsal thalamic nucleus. Brain Res 780: 9–19, 1998 [DOI] [PubMed] [Google Scholar]
- Goodridge JP, Taube JS. Preferential use of the landmark navigational system by head direction cells in rats. Behav Neurosci 109: 49–61, 1995 [DOI] [PubMed] [Google Scholar]
- Harvey CD, Collman F, Dombeck DA, Tank DW. Intracellular dynamics of hippocampal place cells during virtual navigation. Nature 461: 941–946, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klatzky Loomis JM, Beall ACRL, Chance SS, Golledge RG. Spatial updating of self-position and orientation during real, imagined, and virtual locomotion. Psychol Sci 9: 293–298, 1998 [Google Scholar]
- Knierim JJ, Kudrimoti HS, McNaughton BL. Place cells, head direction cells, and the learning of landmark stability. J Neurosci 15: 1648–1659, 1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knierim JJ, Zhang K. Attractor dynamics of spatially correlated neural activity in the limbic system. Annu Rev Neurosci 35: 267–285, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Montello DR, Waller D, Hegarty M, Richardson AE. Spatial memory of real environments, virtual environments, and maps. In: Human Spatial Memory, Remembering Where, edited by Allen GL. Mahwah, NJ: Lawrence Erlbaum, 2004, p. 251–286 [Google Scholar]
- Peruch P, Vercher JL, Gauthier GM. Acquisition of spatial knowledge through visual exploration of simulated environments. Ecol Psychol 7: 1–20, 1995 [Google Scholar]
- Raphan T, Matsuo V, Cohen B. Velocity storage in the vestibulo-ocular reflex arc (VOR). Exp Brain Res 35: 229–248, 1979 [DOI] [PubMed] [Google Scholar]
- Redish AD, Elga AN, Touretzky DS. A coupled attractor model of the rodent head direction system. Netw Comput Neural Syst 7: 671–685, 1996 [Google Scholar]
- Richardson AE, Montello DR, Hegarty M. Spatial knowledge acquisition from maps and from navigation in real and virtual environments. Mem Cognit 27: 741–750, 1999 [DOI] [PubMed] [Google Scholar]
- Riecke BE, van Veen HAHC, Bülthoff HH. Visual homing is possible without landmarks: a path integration study in virtual reality. Presence (Camb) 11: 443–473, 2002 [Google Scholar]
- Ruddle RA, Lessels S. For efficient navigation search, humans require full physical movement, but not a rich visual scene. Psychol Sci 17: 460–465, 2006 [DOI] [PubMed] [Google Scholar]
- Russell NA, Horii A, Smith PF, Darlington CL, Bilkey DK. Bilateral peripheral vestibular lesions produce long-term changes in spatial learning in the rat. J Vestib Res 13: 9–16, 2003 [PubMed] [Google Scholar]
- Sharp PE, Blair HT, Cho J. The anatomical and computational basis of the rat head-direction cell signal. Trends Neurosci 24: 289–294, 2001a [DOI] [PubMed] [Google Scholar]
- Sharp PE, Tinkelman A, Cho J. Angular velocity and head direction signals recorded from the dorsal tegmental nucleus of gudden in the rat: implications for path integration in the head direction cell circuit. Behav Neurosci 115: 571–588, 2001b [PubMed] [Google Scholar]
- Shinder ME, Taube JS. Differentiating ascending vestibular pathways to the cortex involved in spatial cognition. J Vestib Res 20: 3–23, 2010 [DOI] [PubMed] [Google Scholar]
- Shinder ME, Taube JS. Active and passive movement are encoded equally by head direction cells in the anterodorsal thalamus. J Neurophysiol 106: 788–800, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skaggs WE, Knierim JJ, Kudrimoti HS, McNaughton BL. A model of the neural basis of the rat's sense of direction. Adv Neural Inf Process Syst 7: 173–180, 1995 [PubMed] [Google Scholar]
- Song P, Wang XJ. Angular path integration by moving “hill of activity”: a spiking neuron model without recurrent excitation of the head-direction system. J Neurosci 25: 1002–1014, 2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stackman RW, Golob EJ, Basset JP, Taube JS. Passive transport disrupts directional path integration by rat head direction cells. J Neurophysiol 90: 2862–2874, 2003 [DOI] [PubMed] [Google Scholar]
- Stackman RW, Taube JS. Firing properties of head direction cells in rat anterior thalamic neurons: dependence upon vestibular input. J Neurosci 17: 4349–4358, 1997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stackman RW, Taube JS. Firing properties of rat lateral mammillary single units: head direction, head pitch, and angular head velocity. J Neurosci 18: 9020–9037, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun JJ, Chan GSW, Campos JL. Active navigation and orientation-free spatial representations. Mem Cognit 32: 51–71, 2004 [DOI] [PubMed] [Google Scholar]
- Taube JS. Head direction cells recorded in the anterior thalamic nuclei of freely moving rats. J Neurosci 15: 70–86, 1995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taube JS. The head direction signal: origins and sensory-motor integration. Annu Rev Neurosci 30: 181–207, 2007 [DOI] [PubMed] [Google Scholar]
- Taube JS, Bassett JP. Persistent neural activity in head direction cells. Cereb Cortex 13: 1162–1172, 2003 [DOI] [PubMed] [Google Scholar]
- Taube JS, Muller RU. Comparison of head direction cell activity in the postsubiculum and anterior thalamus of freely moving rats. Hippocampus 8: 87–108, 1998 [DOI] [PubMed] [Google Scholar]
- Taube JS, Muller RU, Ranck JB., Jr Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis. J Neurosci 10: 420–435, 1990a [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taube JS, Muller RU, Ranck JB., Jr Head-direction cells recorded from the postsubiculum in freely moving rats. II. Effects of environmental manipulations. J Neurosci 10: 436–447, 1990b [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tomlinson RD, Robinson DA. Signals in vestibular nucleus mediating vertical eye movements in the monkey. J Neurophysiol 51: 1121–1136, 1984 [DOI] [PubMed] [Google Scholar]
- Valerio S, Taube JS. Path integration: how the head direction signal maintains and corrects spatial orientation. Nat Neurosci 15: 1445–1453, 2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallace DG, Hines DJ, Pellis SM, Whishaw IQ. Vestibular information is required for dead reckoning in the rat. J Neurosci 22: 10009–10017, 2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams B, Narasimham G, Rump B, McNamara TP, Carr TH, Rieser J, Bodenheimer B. Exploring large virtual environments with an HMD when physical space is limited. In: APGV '07, Proceedings of the 4th Symposium on Applied Perception in Graphics and Visualization. New York: ACM, 2007, p. 41–48 [Google Scholar]
- Yoder RM, Clark BJ, Brown JE, Lamia MV, Valerio S, Shinder ME, Taube JS. Both visual and idiothetic cues contribute to head direction cell stability during navigation along complex routes. J Neurophysiol 105: 2989–3001, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yokota J, Reisine H, Cohen B. Nystagmus induced by electrical stimulation of the vestibular and prepositus hypoglossi nuclei in the monkey: evidence for site of induction of velocity storage. Exp Brain Res 92: 123–138, 1992 [DOI] [PubMed] [Google Scholar]
- Yoshida M, Hasselmo ME. Persistent firing supported by an intrinsic cellular mechanism in a component of the head direction system. J Neurosci 29: 4945–4952, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang K. Representation of spatial orientation by the intrinsic dynamics of the head-direction cell ensemble: a theory. J Neurosci 16: 2112–2126, 1996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zugaro MB, Berthoz A, Wiener SI. Peak firing rates of rat anterodorsal thalamic head direction cells are higher during faster passive rotations. Hippocampus 12: 481–486, 2002 [DOI] [PubMed] [Google Scholar]
- Zugaro MB, Tabuchi E, Fouquier C, Berthoz A, Wiener SI. Active locomotion increases peak firing rates of anterodorsal thalamic head direction cells. J Neurophysiol 86: 692–702, 2001 [DOI] [PubMed] [Google Scholar]