

Abstract
Background
We have previously demonstrated that heparin-binding EGF-like growth factor (HB-EGF) and mesenchymal stem cell (MSC) administration protect the intestines from ischemia/reperfusion (I/R) injury in vivo, with amniotic fluid-derived MSC (AF-MSC) being more efficacious than bone marrow-derived MSC (BM-MSC). The goal of the current study was to determine whether the protective effects of HB-EGF were from direct effects on MSC or via alternative mechanisms.
Methods
Murine MSC were transfected with an HB-EGF plasmid or control plasmid by electroporation. Mice were subjected to segmental intestinal I/R injury and received either BM-MSC or AF-MSC either with or without exogenous HB-EGF, or BM-MSC or AF-MSC that endogenously over-expressed HB-EGF. MSC engraftment, intestinal histologic injury, and intestinal permeability were quantified.
Results
There was increased MSC engraftment into injured compared to uninjured intestine. HB-EGF increased AF-MSC engraftment into injured intestine. Administration of HB-EGF and MSC improved intestinal histology and intestinal permeability after I/R injury, with AF-MSC being most efficacious. The effect of HB-EGF on MSC was similar when the growth factor was administered exogenously, or when it was overexpressed endogenously.
Conclusions
The effect of HB-EGF on AF-MSC was similar with both exogenous administration or endogenous overexpression of the growth factor, implying that HB-EGF has a direct effect on AF-MSC. This information may assist in guiding potential future AF-MSC-based therapies for patients at risk of intestinal ischemic injuries.
Keywords: heparin-binding EGF-like growth factor, mesenchymal stem cells, bone marrow, amniotic fluid, ischemia/reperfusion, intestinal injury
INTRODUCTION
Mesenchymal stem cells (MSC) are non-hematopoietic, pluripotent, self-renewing progenitor cells with a characteristic spindle-shaped morphology that contribute to the maintenance and regeneration of various connective tissues [1]. They are mobilized from bone marrow in response to tissue injury and aid in repair after a variety of end-organ injury-models including models of myocardial infarction [2], spinal cord injury [3], renal ischemia/reperfusion (I/R) injury [4], intestinal radiation injury [5] and ischemic cerebral injury[6]. MSC can engraft into injured tissues where they differentiate to replace injured cells [7,8]. MSC also exert paracrine functions by secreting protective factors that act on injured cells [9,10]. The robust, self-renewing, multilineage differentiation potential of MSC makes these cells very desirable candidates for possible clinical cellular therapy [11].
Populations of MSC derived from bone marrow (BM-MSC) [12] and amniotic fluid (AF-MSC) [13] have been isolated. Both MSC subtypes have dynamic differentiation potential and can be induced into cells of all three embryonic lineages [14,15]. AF-MSC may actually have more potent proliferation [16] and differentiation potential compared to BM-MSC [17]. Furthermore, unlike embryonic stem cells, AF-MSC are not tumorigenic [15]. These characteristics make AF-MSC a very attractive option for potential cellular therapy.
Heparin-binding EGF-like growth factor (HB-EGF) was initially identified as a secreted product of cultured human macrophages [18] that is a member of the epidermal growth factor (EGF) family [19]. We have previously demonstrated that HB-EGF promotes angiogenesis [20], and acts as a potent intestinal cytoprotective agent in animal models of intestinal I/R injury [21], hemorrhagic shock and resuscitation (HS/R) [22] and experimental necrotizing enterocolitis (NEC) [23]. We have also demonstrated that HB-EGF can protect native intestinal stem cells (ISC) in a rat model of NEC [24]. HB-EGF promotes MSC proliferation and migration, and protects MSC from apoptosis, with a more profound effect on AF-MSC compared to BM-MSC [25]. Furthermore, HB-EGF and BM-MSC act synergistically to reduce intestinal injury and improve survival in experimental NEC [26] and in murine I/R injury [27]. The goal of the current study was to further explore the synergistic effects of HB-EGF and MSC in a model of intestinal I/R injury in order to determine whether HB-EGF improves the efficacy of MSC via direct interactions or by separate mechanism(s) working in parallel.
METHODS
Isolation of BM-MSC
MSC isolation was performed with approval from the Institutional Animal Care and Use Committee of the Research Institute of Nationwide Children’s Hospital (protocol #AR06-0002). BM-MSC were harvested from adult pan-EGFP C57/BL6 mice following previously described protocols [12]. Briefly, mice were euthanized by cervical dislocation, and the femurs and tibias were removed and dissected free of surrounding tissue using sterile technique. The marrow was flushed out with 2 ml of phosphate-buffered saline (PBS) using a sterile syringe and 20 gauge needle. The marrow pellet was dispersed by gentle pipetting and transferred to uncoated cell culture flasks.
Isolation of AF-MSC
Amniotic fluid was obtained via amniocentesis of pan-EGFP C57/BL6 mice using an adaptation of previously described techniques [13]. Female mice at 12.5 days gestation were anesthetized with 2.5% tribromoethanol via intraperitoneal (IP) injection. The abdominal skin was shaved and scrubbed with 70% ethanol. A midline laparotomy was performed and the gravid uterus identified. The uterus was opened and amniocentesis was performed under direct vision of the individual placentas using a 23 gauge needle. Amniotic fluid samples were transferred to uncoated cell culture flasks. Pan-EGFP mice were used for isolation of BM-MSC and AF-MSC since all of the cells of their body, including their MSC, are fluorescently labeled for ease of tracking of the cells after transplantation.
Cell Culture and Differentiation
After harvesting, BM-MSC and AF-MSC were cultured in Dulbecco’s modified Eagle’s medium: Nutrient Mixture F-12 with GlutaMax (DMEM/F12; Invitrogen, Carlsbad, CA) supplemented with 10% MSC Qualified Fetal Bovine Serum (FBS; Invitrogen, Carlsbad, CA) and gentamycin (5 μg/ml) (Invitrogen, Carlsbad, CA) in uncoated cell culture flasks at 37°C in a humidified atmosphere of 5%CO2/95%Nitrogen. After 24 h, non-adherent cells were washed away with PBS and discarded. Adherent MSC were purified and expanded during successive passages. MSC were passaged once they achieved 80% confluence to expand the primary cultures. MSC from passages 4–9 were used for all experiments. Differentiation assays were performed with cells at passage 4–5, and the remaining assays were performed using cells at passage 5–9.
MSC pluripotency was confirmed using the STEMPRO Adipogenesis Differentiation Kit (Invitrogen, Carlsbad, CA) and the STEMPRO Osteogenesis Differentiation Kit (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions, as we have described previously [25].
MSC Transfection
pCMV6-Entry vector carrying a full length human HB-EGF plasmid was obtained from ORIGENE (Rockville, MD). Passage 2–4 EGFP-positive BM-MSC or AF-MSC harvested from pan-EGFP C57/BL6 mice as described above were transfected with 4 μg plasmid DNA or scrambled control plasmid DNA by electroporation using a Nucleofector Kit (Lonza, Walkersville, MD). Transfected cells were seeded in 35 mm dishes and incubated in GlutaMax DMEM/F12 medium containing 10% FBS for 24 h prior to transplantation. To determine transfection efficiency, BM-MSC or AF-MSC were co-transfected with 0.8 μg RFP and 3.2 μg of human HB-EGF plasmid DNA, fixed for fluorescence microscopy, and transfection efficiency determined by quantifying RFP-positive cells. A portion of the transfected cells were used for RNA isolation using an mirVanaTM Isolation Kit (Ambion, Grand Island, NY) and subsequently subjected to real-time RT-PCR analysis of human HB-EGF mRNA expression. Human HB-EGF primers: Forward primer sequence (5′ sense) 5′-CTCTCCCTGCCAAGTCTCAG-3′; Reverse primer sequence (3′ antisense): 5′-CTGCATGGAGTAGCACCAGA-3′. Data were normalized to GAPDH and shown relative to HB-EGF gene expression.
Murine Ischemia/Reperfusion Model
All operative procedures were performed with approval from the Institutional Animal Care and Use Committee of the Research Institute of Nationwide Children’s Hospital (protocol #AR000903). 8 to 10 week old male wild-type FVB mice weighing ~20 grams were used for all experiments. Animals were anesthetized with 3% isoflurane and the abdomen was shaved and cleansed with 70% isopropanol. Anesthesia was maintained using 1% isoflurane and body temperature was maintained with a heating pad.
A midline laparotomy was made and branches of the superior mesenteric artery were identified. Branches supplying the distal six centimeters of the terminal ileum were occluded with non-traumatic vascular clamps. For sham operated animals, branches were identified but no vascular clamps were applied. The abdomen was temporarily closed and segmental mesenteric artery occlusion (sMAO) to the terminal ileum was maintained for 60 min. Vascular clamps were then removed and 0.2 mL of either HB-EGF (1000 μg/kg) or PBS was administered via an intraluminal injection into the more proximal small intestine using a 0.3 mL low-dose U-100 insulin syringe and a 29-guage needle (Becton Dickinson, Franklin Lakes, NJ). The abdominal wall was sutured closed in layers. Animals were kept warm and observed until fully awake and ambulatory. Animals were euthanized 24 h after reperfusion via cervical dislocation under anesthesia.
MSC Preparation and Administration
Adherent BM-MSC or AF-MSC were trypsinized (0.25% trypsin; Cellgro, Manassas, VA) for 5 min and then neutralized with DMEM/F12 supplemented with 10% FBS. Cells were quantified using a hemocytometer. MSC were centrifuged, washed with sterile PBS, re-suspended in PBS to a concentration of 5x106 MSC/mL, and then gently mixed to prevent aggregation. Animals received 0.2 mL of either MSC suspension (1x106 total MSC) or PBS 2 h after reperfusion via intraperitoneal (IP) injection with 29-guage needles.
MSC Intestinal Engraftment
Mice were randomized into eight groups: (1) sham operated mice (sham) with control-transfected BM-MSC; (2) sham + HB-EGF-transfected BM-MSC; (3) sMAO + control-transfected BM-MSC; (4) sMAO + HB-EGF-transfected BM-MSC; (5) sham + control-transfected AF-MSC; (6) sham + HB-EGF-transfected AF-MSC; (7) sMAO + control-transfected AF-MSC or (8) sMAO + HB-EGF-transfected AF-MSC. There were ≥ 5 mice in each experimental group. Terminal ileum from sham operated mice as well as uninjured jejunum from mice exposed to sMAO were both used as controls. After euthanasia, the terminal ileum and jejunum were harvested and fixed in solution containing 1% paraformaldehyde, 15% picric acid and 0.1 M sodium phosphate buffer (pH 7.0) and shaken gently at 4°C overnight. Thirty μm frozen sections were mounted in DAPI containing mounting media (Vector Laboratories, Burlingame, CA). Fluorescence was observed using a Zeiss Axioskop fluorescent microscope (Carl Zeiss, New York, NY). MSC quantification was performed by counting EGFP-positive cells per crypt-villus axis in three separate intestinal sections per mouse at 200X magnification.
Histologic Injury Score
Mice were randomized into eleven groups: (1) sham; (2) sMAO; (3) sMAO + intraluminal HB-EGF; (4) sMAO + BM-MSC; (5) sMAO + intraluminal HB-EGF + BM-MSC; (6) sMAO + control-transfected BM-MSC; (7) sMAO + HB-EGF-transfected BM-MSC; (8) sMAO + AF-MSC; (9) sMAO + intraluminal HB-EGF + AF-MSC; (10) sMAO + control-transfected AF-MSC or (11) sMAO + HB-EGF-transfected AF-MSC. There were ≥ 5 mice in each experimental group. After euthanasia, the terminal ileum was harvested and fixed in 10% formalin overnight. Paraffin-embedded sections were prepared and stained with hematoxylin and eosin. Histologic scoring of the depth of tissue injury was performed as described by Chiu et al [28] with modifications as follows: 0, no damage; 1, subepithelial space at villous tip; 2, loss of mucosal lining of the villous tip; 3, loss of less than half of the villous structure; 4, loss of more than half of villous structure; and 5, transmural necrosis. Sections were evaluated blindly by two observers, with all scores averaged.
Intestinal Permeability Assay
Mice were randomized into the same groups listed above for histologic injury scoring. There were ≥ 5 mice in each experimental group. After euthanasia, terminal ileal mucosal barrier function was assessed using the ex vivo isolated everted gut sac method as we have previously described [29]. In brief, six-centimeter segments of terminal ileum were harvested, everted, and incubated in ice-cold Krebs-Henseleit bicarbonate buffer (KHBB) at pH 7.4. Fluorescein-isothiocynate dextran (FD4, molecular weight; 4000 Da) was used as a permeability probe. Everted gut sacs were gently distended by injecting 0.4 mL of KHBB and suspending the sacs in KHBB with added FD4 (60 μg/mL), representing the FD4muc, for 30 min. The incubation medium was maintained at 37°C and was continuously bubbled with a gas mixture containing 95% O2 and 5% CO2. Gut length (L) and diameter (D) were measured, and the intraluminal KHBB (FD4ser) was collected and measured (intraluminal volume). Both FD4muc and FD4ser were measured with a fluorescence spectrophotometer (SpectraMax Plus, Molecular Devices, CA). Gut permeability was expressed as the mucosal-to-serosal clearance of FD4 as follows:
Statistical Analyses
Experimental data were expressed as mean ± SD. Intragroup statistical differences were determined with ANOVA analysis and statistical significances between individual experimental groups were determined using a post-hoc Student’s t-test. Differences were considered to be statistically significant if p<0.05.
RESULTS
HB-EGF plasmid transfection leads to increased HB-EGF mRNA expression in MSC
EGFP-positive BM-MSC or AF-MSC were co-transfected with human HB-EGF and with RFP, and transfection efficiency was determined by the ratio of RFP-positive cells (red) to EGFP-labeled MSC (green) (Figure 1A). Approximately 48% transfection efficiency was achieved. Real time RT-PCR analysis showed that the expression of human HB-EGF in the HB-EGF-transfected BM-MSC and AF-MSC was significantly higher than that in either non-transfected MSC or in scrambled control plasmid transfected MSC (Figure 1B and 1C).
Figure 1. Transfection efficiency of human HB-EGF in MSC.

(A) EGFP-positive BM-MSC were co-transfected with RFP and either human HB-EGF or scrambled control plasmids. RFP positive cells (red) were visualized under fluorescence microscopy. Similar results were found for AF-MSC (not shown). Total RNA was extracted from non-transfected, scrambled control plasmid transfected, or HB-EGF transfected BM-MSC (B) or AF-MSC (C) and real-time RT-PCR performed. Data were normalized to GAPDH and are shown relative to the expression of the HB-EGF gene (*p<0.05 vs. non-transfected or scramble transfected controls).
MSC Preferentially Engraft into Injured Intestine and endogenous HB-EGF Promotes AF-MSC Engraftment
Representative intestinal sections displaying MSC engraftment are shown in Figure 2A. There was increased intestinal engraftment into injured terminal ileum compared to either uninjured jejunum from the same animal, or to uninjured terminal ileum from sham operated animals for both BM-MSC (Figure 2B) and for AF-MSC (Figure 2C). Administration of HB-EGF overexpressing BM-MSC led to similar engraftment compared to scrambled control transfected BM-MSC (Figure 2B). On the other hand, administration of HB-EGF overexpressing AF-MSC led to significantly increased engraftment compared to scrambled control transfected AF-MSC (Figure 2C). HB-EGF overexpression had no effect on MSC engraftment into uninjured intestine (Figure 2B,C).
Figure 2. MSC intestinal engraftment.

(A) Shown are representative fluorescent microscopy images of: a) uninjured jejunum from an animal subjected to sMAO and treated with scrambled control-transfected BM-MSC, b) terminal ileum from an animal treated subjected to sMAO and treated with scrambled control-transfected BM-MSC, c) terminal ileum from an animal subjected to sMAO and treated with HB-EGF-transfected BM-MSC, d) uninjured jejunum from an animal subjected to sMAO and treated with scrambled control-transfected AF-MSC, e) terminal ileum from an animal treated subjected to sMAO and treated with scrambled control-transfected AF-MSC, and f) terminal ileum from an animal subjected to sMAO and treated with HB-EGF-transfected AF-MSC. (B) Quantification of BM-MSC intestinal engraftment. Mice were subjected to sMAO or to sham surgery. There was increased BM-MSC engraftment into sMAO injured ileum compared to sMAO uninjured jejunum or sham uninjured ileum. Values represent mean ± SD with ≥ 5 animals per group. *p< 0.01 compared to sMAO uninjured jejunum or to sham operated ileum. (C) Quantification of AF-MSC intestinal engraftment. Mice were subjected to sMAO or to sham surgery. There was increased AF-MSC engraftment into sMAO injured ileum compared to sMAO uninjured jejunum or sham uninjured ileum. In addition, there was increased engraftment of HB-EGF-transfected AF-MSC compared to scrambled control-transfected AF-MSC into sMAO inured ileum. Values represent mean ± SD with ≥ 5 animals per group. *p< 0.01 compared to sMAO uninjured jejunum or to sham operated ileum; **p< 0.01 compared to sMAO uninjured jejunum or to sham operated ileum, and p<0.05 compared to sMAO injured ileum treated with scrambled control-transfected AF-MSC.
MSC Transplantation Improves Intestinal Histologic Injury
As expected, there was significantly increased intestinal histologic injury in the terminal ileum in animals subjected to sMAO compared to sham operated animals (3.38 ± 0.64 vs. 0.06 ± 0.17; p<0.01) (Figure 3A,B). Administration of intraluminal HB-EGF led to significantly decreased terminal ileal histologic injury in animals subjected to sMAO (0.93 ± 0.35 vs. 3.38 ± 0.64; p<0.01). Administration of BM-MSC significantly decreased terminal ileal histologic injury in both the absence (0.9±0.42; p<0.01) or presence (0.8±0.45; p<0.01) of intraluminal HB-EGF (Figure 3A). Treatment with scrambled control transfected BM-MSC (0.75 ± 0.5; p<0.01) or with HB-EGF transfected BM-MSC (0.75 ± 0.65; p<0.01) also significantly decreased terminal ileal histologic injury. There were no significant differences between the individual treatment groups. Administration of AF-MSC also decreased terminal ileal histologic injury both in the absence (1 ± 0.61; p<0.01) or presence (0.85 ± 0.41; p<0.01) of intraluminal HB-EGF. Treatment with scrambled control transfected AF-MSC (1 ± 0.41; p<0.01) or with HB-EGF-transfected AF-MSC (0.88 ± 0.25; p<0.01) also significantly decreased terminal ileal histologic injury (Figure 3B). Again, there were no significant differences between individual treatment groups.
Figure 3. Intestinal histologic injury.

(A) BM-MSC, (B) AF-MSC. Mice were subjected to sham surgery or to sMAO and received the treatments indicated. All treatment groups had improved intestinal histologic injury compared to sMAO injury alone. Values represent mean ± SD with ≥ 5 animals per group. *p < 0.01 compared to sham surgery, **p < 0.01 compared to sMAO alone.
MSC Transplantation and HB-EGF Improve Gut Barrier Function with Synergistic Efficacy
There was significantly increased intestinal permeability, indicative of impaired gut barrier function, in terminal ileum from animals subjected to sMAO compared to sham operated animals (96.5 ± 21.9 vs. 1.7 ± 2.3; p<0.01) (Figure 4A,B). Animals subjected to sMAO but treated with intraluminal HB-EGF had significantly improved gut barrier function compared to untreated animals (40 ± 5.3 vs. 96.5 ± 21.9; p<0.01). Administration of BM-MSC improved gut barrier function in both the absence (31.8 ± 6.9; p<0.01) or presence (32.2 ± 8.4; p<0.01) of intraluminal HB-EGF (Figure 4A). Gut barrier function was also improved with administration of either scrambled control transfected BM-MSC (34.8 ± 6.1; p<0.01) or HB-EGF transfected BM-MSC (33.6 ± 12.1; p<0.01). There were no significant differences between the individual treatment groups. On the other hand, administration of AF-MSC improved gut barrier function in both the absence (41.6 ± 10.5; p<0.01) or presence (17.4 ± 8.2; p<0.01) of intraluminal HB-EGF, with significantly improved gut barrier function for combined therapy with intraluminal HB-EGF and AF-MSC compared to either intraluminal HB-EGF alone (p<0.05) or AF-MSC alone (p<0.05) (Figure 4B). Administration of scrambled control transfected AF-MSC (39.9 ± 15.6; p<0.01) or HB-EGF transfected AF-MSC (12.2 ± 6.2; p<0.01) also both significantly improved gut barrier function. There was additional improved gut barrier function with administration of HB-EGF transfected AF-MSC compared to scrambled control transfected AF-MSC (p<0.05).
Figure 4. Intestinal permeability.
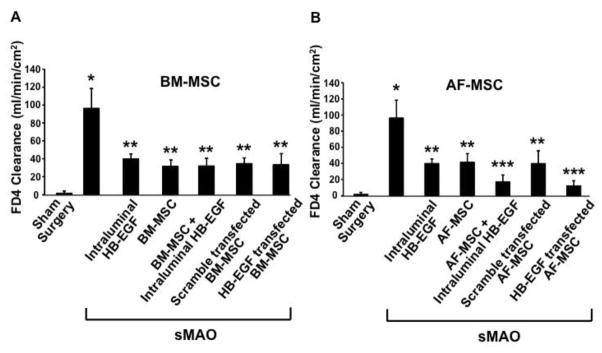
(A) BM-MSC, (B) AF-MSC. Mice were subjected to sham surgery or to sMAO and received the treatments indicated. All treatment groups had improved intestinal permeability injury compared to sMAO injury alone. There was an additional improvement in intestinal permeability for the AF-MSC+HB-EGF and the HB-EGF transfected AF-MSC treatment groups. Values represent mean ± SD with ≥ 5 animals per group. *p < 0.01 compared to sham surgery, **p < 0.01 compared to sMAO; ***p < 0.01 compared to sMAO and p<0.05 compared to other (non-sham) treatment groups.
DISCUSSION
MSC have emerged as an attractive target for regenerative medicine. MSC administration has been shown to be beneficial in multiple end-organ ischemic injury models [2–6]. Improving and augmenting MSC transplantation methodology to modulate therapeutic efficacy is an area of great interest. Others have focused on the transplantation of different MSC sub-types from a variety of primary tissues and optimizing these MSC populations via pre-transplantation conditioning. In the current study we have focused on the administration of HB-EGF in an effort to augment MSC function.
HB-EGF has previously been demonstrated to have potent mitogenic activity for a variety of cell types, including smooth muscle cells, epithelial cells, fibroblasts, keratinocytes and renal tubular cells [30], and is a known chemotactic agent for various cell types [31,32]. It exerts its effects by binding to cell-surface EGF receptors (EGFR) ErbB1 and ErbB4 [33,34] and to the HB-EGF-specific receptor Nardilysin (Nrdc) [35,36]. We have previously characterized the in vitro interactions between HB-EGF and MSC, demonstrating that HB-EGF promotes MSC proliferation and migration, and protects MSC from anoxia-induced apoptosis [25]. HB-EGF has a more potent effect on AF-MSC compared to BM-MSC in vitro [25]. We have also shown that HB-EGF protects resident intestinal stem cells during NEC [24] and that BM-MSC transplantation improves survival in experimental NEC [26]. We also showed that HB-EGF administration combined with MSC protects murine intestines from I/R injury in vivo [27]. We found synergistic efficacy when a combination of AF-MSC and HB-EGF was administered, while this synergism was not observed with administration of BM-MSC and HB-EGF. This parallels our in vitro differential effects of HB-EGF on BM-MSC and AF-MS. What was not clear, however, was whether this apparent in vivo synergism was via a direct effect of HB-EGF on MSC, or if it was predominantly via parallel additive effects of HB-EGF on the intestines themselves. The latter is possible since multiple studies from our lab have shown that HB-EGF is directly able to protect the intestinal mucosa from various forms of intestinal injury. We attempted to address this question by over-expressing endogenous HB-EGF in MSC and comparing the efficacy of these genetically altered MSC to that of non-HB-EGF-transfected MSC administered in combination with intraluminal exogenous HB-EGF.
Our results using a murine intestinal I/R injury model demonstrate that in BM-MSC and AF-MSC, endogenous overexpression of HB-EGF produces similar effects to co-administration of MSC with exogenous HB-EGF. When administered as monotherapy, BM-MSC and AF-MSC protect the intestines from injury to a similar extent. No additional improvement in protection from injury occurred when exogenous HB-EGF was administered in combination with BM-MSC transplantation or when endogenous HB-EGF expression was upregulated in BM-MSC. However, when exogenous HB-EGF administration was combined with AF-MSC transplantation, or when endogenous HB-EGF was over-expressed in AF-MSC, there was significantly increased MSC engraftment into injured intestine, and significantly improved gut barrier function. This supports our in vitro data demonstrating that HB-EGF has a more potent protective effect on AF-MSC compared to BM-MSC [25].
One possible confounding factor in our comparison of HB-EGF over-expression in the different MSC populations studied was the fact that there was increased HB-EGF mRNA expression in AF-MSC compared to BM-MSC. This could partially explain the differential behavior pattern observed between AF-MSC and BM-MSC. However the synergistic effect of exogenous HB-EGF on AF-MSC but not on BM-MSC was noted when equal amounts of exogenous HB-EGF were co-administered. Thus, there appears to be a fundamental difference in the interactions between HB-EGF and AF-MSC compared to BM-MSC.
The mechanisms explaining the differential effects of HB-EGF on AF-MSC compared to BM-MSC have yet to be fully elucidated. Specific interactions between HB-EGF and MSC are currently incompletely understood. Others have shown that HB-EGF promotes MSC proliferation and prevents MSC differentiation via the HER-1 cell surface receptor [37]. It is well known that EGFR plays a pivotal role in proliferation as well as paracrine and autocrine signaling [38]. This has been further demonstrated specifically in BM-MSC [39]. A possible explanation we are currently exploring to explain our observed behavior patterns is a differential expression profile of the known HB-EGF cell surface receptor proteins between the two MSC populations. Previous observations have demonstrated that hypoxia activates many survival and stress pathways, including down regulation of p38MAPK [40], down regulation of PI3K and ERK 1–2 [41], and up regulation of the Akt inhibitor PTEN [42]. We have shown that HB-EGF activates these pathways in various cell types [43], and future experiments will clarify the exact mechanisms utilized by HB-EGF in its effects on MSC.
We have demonstrated that HB-EGF administration with MSC transplantation leads to preferential engraftment of MSC into injured intestine, and improves intestinal histology and gut barrier function, after murine intestinal I/R injury. Both exogenously administered and endogenously overexpressed HB-EGF promotes increased AF-MSC engraftment and have synergistic efficacy with AF-MSC transplantation, leading to further improvement in gut barrier function. These data support the utilization of HB-EGF as a potential method to improve the efficacy of AF-MSC transplantation therapy, and indicates that HB-EGF exerts its effects, at least partially, via direct effects on MSC. Although our results imply a direct effect of HB-EGF on AF-MSC, ongoing studies will clarify the specific mechanistic role(s) of HB-EGF on MSC more directly.
Acknowledgments
This work was supported by NIH R01 GM61193 (GEB). We thank Palak Painter for her help with maintaining animal breeding colonies.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Pittenger MF, Mackay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 2.Kawada H, Fujita J, Kinjo K, et al. Nonhematopoietic mesenchymal stem cells can be mobilized and differentiate into cardiomyocytes after myocardial infarction. Blood. 2004;104:3581–3587. doi: 10.1182/blood-2004-04-1488. [DOI] [PubMed] [Google Scholar]
- 3.Koda M, Okada S, Nakayama T, et al. Hematopoietic stem cell and marrow stromal cell for spinal cord injury in mice. Neuroreport. 2005;16:1763–1767. doi: 10.1097/01.wnr.0000183329.05994.d7. [DOI] [PubMed] [Google Scholar]
- 4.Togel F, Hu Z, Weiss K, et al. Administered mesenchymal stem cells protect against ischemic acute renal failure through differentiation-independent mechanisms. Am J Physiol Renal Physiol. 2005;289:F31–42. doi: 10.1152/ajprenal.00007.2005. [DOI] [PubMed] [Google Scholar]
- 5.Zhang J, Gong JF, Zhang W, et al. Effects of transplanted bone marrow mesenchymal stem cells on the irradiated intestine of mice. J Biomed Sci. 2008;15:585–594. doi: 10.1007/s11373-008-9256-9. [DOI] [PubMed] [Google Scholar]
- 6.Wei L, Fraser JL, Lu ZY, et al. Transplantation of hypoxia preconditioned bone marrow mesenchymal stem cells enhance angiogenesis and neurogenesis after cerebral ischemia in rats. Neurobiol Dis. 2012;46:635–645. doi: 10.1016/j.nbd.2012.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yan Q, Ruan JW, Ding Y, et al. Electro-acupuncture promotes differentiation of mesenchymal stem cells, regeneration of nerve fibers and partial functional recovery after spinal cord injury. ExpToxicol Pathol. 2011;63:151–156. doi: 10.1016/j.etp.2009.11.002. [DOI] [PubMed] [Google Scholar]
- 8.Wong CY, Cheong SK, Mok PL, et al. Differentiation of human mesenchymal stem cells into mesangial cells in post-glomerular injury murine model. Pathology. 2008;40:52–57. doi: 10.1080/00313020701716367. [DOI] [PubMed] [Google Scholar]
- 9.Zarjou A, Kim J, Traylor AM, et al. Paracrine effects of mesenchymal stem cells in cisplatin-induced renal injury requiresheme oxygenase-1. Am J Physiol Renal Physiol. 2011;300:F254–62. doi: 10.1152/ajprenal.00594.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Weil BR, Markel TA, Herrmann JL, et al. Mesenchymal stem cells enhance the viability and proliferation of human fetal intestinal epithelial cells following hypoxic injury via paracrine mechanisms. Surgery. 2009;146:190–197. doi: 10.1016/j.surg.2009.03.031. [DOI] [PubMed] [Google Scholar]
- 11.Baksh D, Song L, Tuan RS. Adult mesenchymal stem cells: characterization, differentiation, and application in cell and gene therapy. J Cell Mol Med. 2004;8:301–316. doi: 10.1111/j.1582-4934.2004.tb00320.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Phinney DG, Kopen G, Isaacson RL, et al. Plastic adherent stromal cells from the bone marrow of commonly used strains of inbred mice: variations in yield, growth, differentiation. J Cell Biochem. 1999;72:570–585. [PubMed] [Google Scholar]
- 13.Baghaban EM, Jahangir S, Aghdami N. Mesenchymal stem cells from murine amniotic fluid as a model for preclinical investigation. Arch Iran Med. 2011;14:96–103. [PubMed] [Google Scholar]
- 14.Delo DM, De Coppi P, Bartsch G, Jr, et al. Amniotic fluid and placental stem cells. Methods Enzymol. 2006;419:426–438. doi: 10.1016/S0076-6879(06)19017-5. [DOI] [PubMed] [Google Scholar]
- 15.DeCoppi P, Bartsch G, Jr, Siddiqui MM, et al. Isolation of amniotic stem cell lines with potential for therapy. Nat Biotechnol. 2007;25:100–106. doi: 10.1038/nbt1274. [DOI] [PubMed] [Google Scholar]
- 16.Yan ZJ, Hu YQ, Zhang HT, et al. Comparison of the neural differentiation potential of human mesenchymal stem cells from amniotic fluid and adult bone marrow. Cell Mol Neurobiol. 2013;33:465–475. doi: 10.1007/s10571-013-9922-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Peister A, Woodruff MA, Prince JJ, et al. Cell sourcing for bone tissue engineering: amniotic fluid stem cells have a delayed, robust differentiation compared to mesenchymal stem cells. Stem Cell Res. 2011;7:17–27. doi: 10.1016/j.scr.2011.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Besner G, Higashiyama S, Klasbrun MP. Isolation and characterization of a macrophage- derived heparin-binding growth factor. Cell Regul. 1990;1:811–819. doi: 10.1091/mbc.1.11.811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Higashiyama S, Abraham JA, Miller J, et al. A heparin-binding growth factor secreted by macrophage-like cells that is related to EGF. Science. 1991;251:936–939. doi: 10.1126/science.1840698. [DOI] [PubMed] [Google Scholar]
- 20.Mehta VB, Besner GE. HB-EGF promotes angiogenesis in endothelial cells via PI3-kinase and MAPK signaling pathways. Growth Factors. 2007;25:253–263. doi: 10.1080/08977190701773070. [DOI] [PubMed] [Google Scholar]
- 21.El-Assal ON, Paddock H, Marquez A, et al. Heparin-binding epidermal growth factor-like growth factor gene disruption is associated with delayed intestinal restitution, impaired angiogenesis, and poor survival after intestinal ischemia in mice. J Pediatr Surg. 2008;43:1182–1190. doi: 10.1016/j.jpedsurg.2008.02.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.El-Assal ON, Radulescu A, Besner GE. Heparin-binding EGF-like growth factor preserves mesenteric microcirculatory blood flow and protects against intestinal injury in rats subjected to hemorrhagic shock and resuscitation. Surgery. 2007;142:234–242. doi: 10.1016/j.surg.2007.04.003. [DOI] [PubMed] [Google Scholar]
- 23.Yu X, Radulescu A, Zorko N, et al. Heparin-binding EGF-like growth factor increases intestinal microvascular blood flow in necrotizing enterocolitis. Gastroenterology. 2009;137:221–230. doi: 10.1053/j.gastro.2009.03.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Chen CL, Yu X, James IO, et al. Heparin-binding EGF-like growth factor protects intestinal stem cells from injury in a rat model of necrotizing enterocolitis. Lab Invest. 2012;92:331–334. doi: 10.1038/labinvest.2011.167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Watkins DJ, Zhou Y, Chen CL, et al. Heparin-binding EGF-like growth factor protects mesenchymal stem cells. J Surg Res. 2012;177:359–364. doi: 10.1016/j.jss.2012.05.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yang J, Watkins DJ, Chen CL, et al. Heparin-binding epidermal growth factor-like growth factor and mesenchymal stem cells act synergistically to prevent experimental necrotizing enterocolitis. J Am Coll Surg. 2012;215:534–545. doi: 10.1016/j.jamcollsurg.2012.05.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Watkins DJ, Yang J, Matthews MB, et al. Synergistic effects of HB-EGF and mesenchymal stem cells in a murine model of intestinal ischemia/reperfusion injury. J Pediatr Surg. 2013;48:1323–1329. doi: 10.1016/j.jpedsurg.2013.03.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chiu CJ, McArdle AH, Brown R, et al. Intestinal mucosal lesion in low-flow states. I. A morphological, hemodynamic, and metabolic reappraisal. Arch Surg. 1970;101:478–483. doi: 10.1001/archsurg.1970.01340280030009. [DOI] [PubMed] [Google Scholar]
- 29.Zhang H, Radulescu A, Besner GE. Heparin-binding epidermal growth factor-like growth factor is essential for preservation of gut barrier function after hemorrhagic shock and resuscitation in mice. Surgery. 2009;146:334–339. doi: 10.1016/j.surg.2009.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Davis-Fleischer KM, Besner GE. Structure and function of heparin-binding EGF-like growth factor (HB-EGF) Front Biosci. 1998;3:d288–299. doi: 10.2741/a241. [DOI] [PubMed] [Google Scholar]
- 31.Tokumara S, Higashiyama S, Endo T, et al. Ectodomain shedding of epidermal growth factor receptor ligands is required for keratinocyte migration in cutaneous wound healing. J Cell Biol. 2000;151:209–220. doi: 10.1083/jcb.151.2.209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Faull RJ, Stanley JM, Fraser S, et al. HB-EGF is produced in the peritoneal cavity and enhances mesothelial cell adhesion and migration. Kidney Int. 2001;59:614–624. doi: 10.1046/j.1523-1755.2001.059002614.x. [DOI] [PubMed] [Google Scholar]
- 33.Beerli RR, Hynes NE. Epidermal growth factor-related peptides activate distinct subsets of ErbB receptors and differ in their biological activities. J Biol Chem. 1996;271:6071–6076. doi: 10.1074/jbc.271.11.6071. [DOI] [PubMed] [Google Scholar]
- 34.Elenius K, Paul S, Allison G, et al. Activation of HER4 by heparin-binding EGF-like growth factor stimulates chemotaxis but not proliferation. EMBO J. 1997;16:1268–1278. doi: 10.1093/emboj/16.6.1268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nishi E, Prat A, Hospital V, et al. N-arginine dibasic convertase is a specific receptor for heparin-binding EGF-like growth factor that mediates cell migration. EMBO J. 2001;20:3342–3350. doi: 10.1093/emboj/20.13.3342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hospital V, Prat A. Nardilysin, a basic residues specific metallopeptidase that mediates cell migration and proliferation. Protein PeptLett. 2004;11:501–508. doi: 10.2174/0929866043406508. [DOI] [PubMed] [Google Scholar]
- 37.Krempera M, Pasini A, Rigo A, et al. HB-EGF/HER-1 signaling in bone marrow mesenchymal stem cells: inducing cell expansion and reversibly preventing multilineage differentiation. Blood. 2005;106:59–66. doi: 10.1182/blood-2004-09-3645. [DOI] [PubMed] [Google Scholar]
- 38.Wilson KJ, Mill C, Lambert S, et al. EGFR ligands exhibit functional differences in models of paracrine and autocrine signaling. Growth Factors. 2012;30:107–116. doi: 10.3109/08977194.2011.649918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kerpedjieva SS, Kim DS, Barbeau DJ, et al. EGFR ligands drive multipotent stromal cells to produce multiple growth factors and cytokines via early growth response-1. Stem Cells Dev. 2012;21:2541–2551. doi: 10.1089/scd.2011.0711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ren H, Accili D, Duan C. Hypoxia converts the myogenic action of insulin-like growth factors into mitogenic action by differentially regulating multiple signaling pathways. Proc Natl Acad Sci U S A. 2010;107:5857–5862. doi: 10.1073/pnas.0909570107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xu R, Chen J, Cong X, et al. Lovastatin protects mesenchymal stem cells against hypoxi- and serum deprivation-induced apoptosis by activation of pi3k/akt and erk1/2. J Cell Biochem. 2008;103:256–289. doi: 10.1002/jcb.21402. [DOI] [PubMed] [Google Scholar]
- 42.Peterson KM, Aly A, Lerman A, et al. Improved survival of mesenchymal stromal cell after hypoxia precontioning: role of oxidative stress. Life Sci. 2011;88:65–73. doi: 10.1016/j.lfs.2010.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.El-Assal ON, Besner GE. HB-EGF enhances restitution after intestinal ischemia/reperfusion via PI3K/Akt and MEK/ERK1/2 activation. Gastroenterology. 2005;129:609–625. doi: 10.1016/j.gastro.2005.05.054. [DOI] [PubMed] [Google Scholar]