

Abstract
Ductal carcinoma in situ (DCIS) is a non-malignant lesion of the breast with the potential to progress to invasive ductal carcinoma (IDC). The disappearance and breakdown of the myoepithelial cell layer and basement membrane in DCIS have been identified as major events in the development of breast cancer. The MCF10DCIS.com cell line is a well-established model which recapitulates the progression of breast cancer from DCIS to IDC. We have previously reported that a novel Gemini vitamin D analog, 1α,25-dihydroxy-20R-21(3-hydroxy-3-deuteromethyl-4,4,4-trideuterobutyl)-23-yne-26,27-hexafluoro-cholecalciferol (BXL0124) is a potent inhibitor of the growth of MCF10DCIS.com xenografted tumors without hypercalcemic toxicity. In the present study, we utilized the MCF10DCIS.com in vivo model to assess the effects of BXL0124 on breast cancer progression from weeks 1 to 4. Upon DCIS progression to IDC from weeks 3 to 4, tumors lost the myoepithelial cell layer and basement membrane as shown by immunofluorescence staining with smooth muscle actin (SMA) and laminin 5, respectively. Administration of BXL0124 maintained the critical myoepithelial cell layer as well as basement membrane, and animals treated with BXL0124 showed a 43% reduction in tumor volume by week 4. BXL0124 treatment decreased cell proliferation and maintained vitamin D receptor (VDR) levels in tumors. In addition, the BXL0124 treatment reduced the mRNA levels of matrix metalloproteinases (MMPs) starting at week 3, contributing to the inhibition of invasive transition. Our results suggest that the maintenance of DCIS plays a significant role in the cancer preventive action of the Gemini vitamin D BXL0124 during the progression of breast lesions.
Keywords: Vitamin D analog, Vitamin D receptor, Ductal Carcinoma in Situ, Invasive Ductal Carcinoma, Matrix Metalloproteinases
Introduction
Breast cancer is one of the most common cancers, and the second highest cancer-related cause of death among women (1). Regardless of its high prevalence in society, the etiology and pathogenesis of breast cancer have yet to be elucidated (2-5). Genetic, hormonal and environmental factors have been suggested as causes of breast cancer (6, 7). Early development of breast cancer consists of non-invasive lesions such as ductal carcinoma in situ (DCIS), and approximately 25% of breast abnormalities detected during screening are DCIS (8). DCIS is defined by enhanced proliferation of intraductal epithelial cells (9). DCIS is a pre-cancerous lesion and if left untreated, approximately 30 to 50% of cases will progress to invasive disease (10-12). Therefore, the transition from DCIS to invasive ductal carcinoma (IDC) is a crucial event in the advancement of breast tumors.
The human breast ductal structure is made up of an inner layer of luminal epithelial cells, surrounded by an outer layer of myoepithelial cells and is separated from the stroma by the basement membrane (13). In order for tumor cells to invade and metastasize, they must penetrate these two protective layers (14). The disappearance and breakdown of the myoepithelial cell layer and basement membrane in DCIS have been identified as major events in the progression from DCIS to IDC (15). Thus, it is important to find chemopreventive agents, such as vitamin D, that could inhibit the progression to later stages of breast cancer.
Exposure to sunlight has shown an inverse relationship to breast cancer risk (16). This is linked to the increased production of vitamin D3 upon exposure to ultraviolet light (17-20). Since this discovery, many studies have linked 1,25-dihydroxyvitamin D3 (1,25(OH)2D3), the active metabolite of vitamin D, and vitamin D analogs to the suppression of cancer cell invasion, proliferation, and metastasis (21-24). However, the pharmacological dose of 1,25(OH)2D3 required to elicit a response on breast tumors can induce hypercalcemic toxicity (24, 25). Therefore, non-calcemic vitamin D analogs have been of interest in the inhibition of breast cancer (21, 25-27). We have previously demonstrated that a Gemini vitamin D analog BXL0124 significantly decreased tumor growth of MCF10DCIS.com xenografts in vivo without hypercalcemic toxicity (25, 28). However, the preventive effects of BXL0124 on the early transition from DCIS to IDC have not been elucidated. We utilized the MCF10DCIS.com progression model to investigate whether BXL0124 blocks or delays the early transition of DCIS to IDC in vivo.
The MCF10DCIS.com cell line is one of the human MCF10A series of cell lines which is unique in the fact that it is representative of breast cancer progression in vitro and in vivo (29, 30). The MCF10DCIS.com cell line consistently produces comedo DCIS-like lesions in animal xenografts, which highly resemble the histopathology of human DCIS in xenograft models. The DCIS-like lesions have been shown to reproducibly progress to invasive tumors, providing a unique in vivo model to investigate the transition for DCIS to IDC (28, 29). The MCF10DCIS.com xenograft model forms histology that resembles human DCIS, however this is not classical DCIS, which are premalignant cells confined to the mammary ducts. Therefore, from this point forward when we refer to DCIS we are referring to the DCIS-like histology that is observed in the subcutaneous xenografts. In addition, Hu et al. showed similarities of cell type specific expression profiles between human DCIS samples and MCF10DCIS.com xenografts by comparing myoepithelial and epithelial cell gene expression profiles (15). The study showed a statistically significant enrichment of genes involved in the extracellular matrix, basement membrane structure, and development in the myoepithelial cell populations of both the human DCIS samples and MCF10DCIS.com xenografts. Since the enrichment patterns of genes were highly similar from human to the MCF10DCIS.com xenograft tumors, this is an excellent model to test the effects of chemopreventive agents on the structural, molecular, and phenotypic changes during the progression of DCIS to IDC. In this report, we investigated the effects of BXL0124 treatment on MCF10DCIS.com xenografts, and found that BXL0124 maintained the integrity of critical structures related to non-cancerous breast lesions which are typically lost during the progression to malignant disease.
Materials and Methods
Reagents and Cell Culture
Gemini vitamin D analog, 1α,25-dihydroxy-20R-21(3-hydroxy-3-deuteromethyl-4,4,4-trideuterobutyl)-23-yne-26,27-hexafluoro-cholecalciferol (BXL0124, >95% purity (31)) was provided by BioXell, Inc. (Nutley, NJ) and dissolved in DMSO. For in vivo animal experiments, BXL0124 was diluted in Cremophor EL: PBS (1:8, v/v). MCF10DCIS.com human breast cancer cell line was provided by Dr. Fred Miller at the Barbara Ann Karmanos Cancer Institute (Detroit, MI). The MCF10DCIS.com cell line was authenticated by short tandem repeat profiling at American Type Culture Collection (Manassas, VA). MCF10DCIS.com cells were maintained in monolayer cell culture in DMEM/F12 medium supplemented with 5% horse serum, 1% penicillin/streptomycin, and 1% HEPES solution at 37°C, 5% CO2.
Xenograft Animal Studies
All animal studies were approved by the Institutional Review Board for the Animal Care and Facilities Committee of Rutgers University. Female nude mice (5-6 weeks old) were purchased from Charles River Laboratories (Wilmington, MA). They were allowed to acclimate to the facilities for two weeks at which time they were injected (7-8 weeks old) with human MCF10DCIS.com cells into the dorsal area at 106 cells per site and treatment began the following day. Mice were treated with DMSO control or BXL0124 (0.1μg/kg Body Weight) intraperitoneally six times per week over the course of 4 weeks. Tumors were palpated twice per week and total body weights were measured weekly. Tumors were measured with a vernier caliper and tumor volume (V; cubed centimeters) was calculated using the equation V = D*d2/2 where D (centimeters) and d (centimeters) are the largest and smallest perpendicular diameters. Animals were sacrificed 1, 2, 3, and 4 weeks after injection at which time tumors were excised and blood drawn for further analysis. Tumors from 1, 2, 3, and 4 weeks were fixed in 10% formalin for 15 hours for immunohistochemical analysis. Tumors from 3 and 4 weeks were snap frozen in liquid nitrogen for qPCR analysis.
Determination of Serum Calcium Level
The determination of calcium concentration in serum was carried out by using the calcium reagent set from Pointe Scientific, Inc. (Canton, MI) following the manufacturers protocol. Briefly, serum (4 μL) was mixed with the diluted reagent (200 μL) in the wells of a 96-well plate and incubated at room temperature for 1 minute and absorbance read at 570 nm using a Tecan infinite M200 plate reader (Tecan, Durham, NC). The calcium concentrations were calculated from calcium standards provided by the manufacturer.
Immunohistochemical and Immunofluorescence Analysis
Subcutaneous tumors were fixed, embedded in paraffin and sectioned at 5 μm thickness. Individual tumors were analyzed histopathologically by hematoxylin and eosin (H&E) staining. For immunohistochemistry, sections were stained as previously described (32) with antibodies to proliferating cell nuclear antigen (PCNA) (1:8000, Dako, M0879, Carpinteria, CA) and vitamin D receptor (VDR) (1:200, Santa Cruz Biotechnology, sc-13133, Santa Cruz, CA). The sections were counterstained with Harris hematoxylin (Sigma Aldrich, St. Louis, MO). The PCNA nuclear intensity and VDR total pixel intensity were quantified by blinding the samples and having a third party carry out the analysis using a Scan Scope (Aperio, Vista, CA). For immunofluorescence staining, the slides were blocked in 10% goat serum, and then incubated sequentially overnight at 4°C with the combination of primary antibodies to smooth muscle actin (SMA) (1:200, Abcam, ab5694, Cambridge, MA), laminin 5 (1:50, Santa Cruz Biotechnology, sc13587, Santa Cruz, CA), pancytokeratin (panCK) (1:50, Dako, M3515, Carpinteria, CA) and TO-PRO-3 iodide nuclear antibody (Invitrogen, 1μM). Fluorophore-conjugated secondary antibodies (Alexa Fluor 488 or 546, 1:100, Invitrogen, Carlsbad, CA) were incubated at room temperature for 30 minutes. The images were taken using confocal microscopy with lasers at 488 nm, 546 nm, and 633nm (TO-PRO-3). Immunofluorescence was visualized using a Nikon Eclipse C1 Plus confocal microscope system.
Western Blot Analysis
The procedures have been described previously (32). Five tumor samples from each group were homogenized and pooled for analysis. Primary antibodies against VDR (1:200, Thermoscientific, MA1-710, Waltham, MA) and β-actin (1:2000, Sigma-Aldrich, A1978, St. Louis, MO) were used for analysis. Secondary antibodies were from Santa Cruz Biotechnology. Western Blots were quantified by using ImageJ software (US National Institutes of Health, Bethesda, MD) and calculating the relative density of the bands using the gel analyzer command.
Quantitative Real-Time Polymerase Chain Reaction Analysis
These procedures have been reported previously (33, 34). The Taqman® probe-based gene expression system from Applied Biosystems (Foster City, CA) was used to detect the genes of interest.
DCIS Quantification
Hematoxylin and eosin images of DCIS were quantified using ImageJ software (US National Institutes of Health, Bethesda, MD). The total tumor area was selected, and then the areas of DCIS within the tumor were calculated. The percentage of DCIS within each tumor was presented as the area of DCIS divided by the total tumor area.
Statistical analysis
Statistical significance was evaluated using the Student's t-test.
Results
Gemini Vitamin D Analog, BXL0124, Inhibits the Transition of Ductal Carcinoma In Situ to Invasive Ductal Carcinoma in MCF10DCIS.com Xenografts
Previous studies have shown that BXL0124 has an inhibitory effect on the growth of MCF10DCIS.com xenograft mammary tumors at five weeks (28). However, the effect of BXL0124 on DCIS progression to IDC has not been investigated. The early stages of breast cancer, specifically the DCIS to IDC transition, are of utmost importance from a prevention standpoint. Therefore, in the present study, we evaluated the effects of BXL0124 on the early transition of DCIS to IDC. In our preliminary studies, we first compared the DCIS progression of MCF10DCIS.com xenografts between the mammary fat pad and subcutaneous injections. The subcutaneous xenografts formed a higher number of comedo DCIS lesions and more consistently produced DCIS lesions compared to the mammary fat pad xenografts (Supplemental Fig. 1A and 1B), in agreement with a previous report by Hu et al (15). Comparison of four separate lesions from subcutaneous or mammary fat pad at week 3 xenografts shows that subcutaneous xenografts reproducibly form a higher number of DCIS lesions compared to the mammary fat pad xenografts (Supplemental Fig. 1B). The subcutaneous xenografts show a clear point DCIS to IDC transition so it is easy to determine whether pharmacological agents, such as BXL0124, inhibits this transition (Supplemental Fig. 2A), whereas the mammary fat pad xenografts it is not clear when or if the transition is blocked (Supplemental Fig. 2B). Therefore, we proceeded with the MCF10DCIS.com subcutaneous xenograft model for the clarity of studying the effects of pharmacological agents on DCIS progression to IDC. Using MCF10DCIS.com subcutaneous xenografts, we investigated the histopathological and molecular changes that occurred during the growth of tumors in nu/nu mice over the course of 4 weeks. Animals treated with BXL0124 showed a reduction in average tumor volume over the first 3 weeks and significant repression was observed by week 4 with a 43% reduction in tumor size (p < 0.05) (Fig. 1A). BXL0124 treatment did not cause any significant changes in bodyweight or serum calcium levels, indicating that there was no observed hypercalcemic toxicity associated with the given dose (Fig. 1B-C). H&E staining showed that MCF10DCIS.com cells subcutaneously xenografted into nu/nu mice formed lesions histologically resembling that of human DCIS. In the control group, DCIS lesions started to escape to an invasive-like stage at week 3 and the tumors advanced to IDC rapidly by week 4, at which time the tumors were 80% invasive (Fig. 1D). The tumors treated with BXL0124 formed DCIS lesions by week 3 and unlike control tumors, maintained these DCIS lesions through week 4, showing approximately 30% invasive histology (p < 0.05) (Fig. 1D).
Figure 1.

BXL0124 inhibits tumor growth and inhibits progression to IDC in MCF10DCIS.com subcutaneous xenografts in nu/nu mice. A. Average tumor volume at weekly time points is shown, * p < 0.05 (n=5 per group). Tumor volume (V; cubed centimeters) was calculated using the equation V = D*d2/2 where D (centimeters) and d (centimeters) are the largest and smallest perpendicular diameters. B. Average final bodyweight at autopsy is shown (n=5 per group) C. Serum calcium determination to assess hypercalcemic toxicity is shown (n=5 per group) D. A representative hematoxylin and eosin (H&E) staining showing the progression of MCF10DCIS.com subcutaneous xenografts in nu/nu mice from weeks 1, 2, 3, and 4 (40x). DCIS quantification, * p < 0.05 (n=4 per group).
BXL0124 Treatment Decreases the Proliferation of MCF10DCIS.com Tumors at Week 4
Cell proliferation was determined by measuring proliferating cell nuclear antigen (PCNA) by immunohistochemistry staining. The cell proliferation in MCF10DCIS.com tumors remained relatively low through week 3. As control tumors progressed to an invasive phenotype at week 4, there is a marked increase in the level of PCNA expression compared to previous weeks. Treatment with BXL0124 showed a significant decrease in PCNA levels compared to the control at week 4 (Fig. 2A). Four tumors from each group were blinded and were analyzed for PCNA staining intensity. The intensities were scored from 0+ (negative staining) to 3+ (strongest staining) for each individual cell. The sum of all positive staining including 1+, 2+, and 3+ was used to calculate the percentage of PCNA-positive cells. Quantification showed that 52% of the cells were PCNA-positive in the control group at week 4, whereas only 32% of cells were positive in the BXL0124-treated group at week 4 (p < 0.05) (Fig. 2B).
Figure 2.

BXL0124 treatment decreases the proliferation of MCF10DCIS.com tumors at week 4. A. A representative immunohistochemical analysis of PCNA in tumor samples from weeks 1, 2, 3, and 4 is shown (100x). PCNA-positive staining is found in the nucleus of the cells. B. Four tumors from each group were blinded and quantified, three representative areas from each tumor were quantified for the intensity of PCNA staining, mean +/- S.E.M. The staining intensities were scored from 0+ (negative staining) to 3+ (the strongest staining), * p < 0.05.
BXL0124 Treatment Maintains Vitamin D Receptor Levels in MCF10DCIS.com Tumors
MCF10DCIS.com xenograft tumors express vitamin D receptor (VDR). From weeks 1 to 3, VDR levels were similar between control and BXL0124 treatment groups. VDR was lost upon the rapid progression from DCIS to invasive tumors in the control group in week 4. However, treatment with BXL0124 not only maintained DCIS histology but also retained VDR levels at week 4 (Fig. 3A). It is interesting to note that VDR expression is lost where epithelial and stromal cells come in contact, suggesting that this cell-to-cell interaction could potentially reduce VDR levels. Four tumors from each group were blinded and analyzed for VDR staining intensity. The intensities were scored from 0+ (negative staining) to 3+ (strongest staining) based on pixel intensity of staining. The sum of all positive staining including 1+, 2+, and 3+ was used to calculate the percentage of VDR-positive cells. Quantification showed 66% VDR-positive staining in the control tumors, compared to 81% positive staining in the BXL0124 group at week 4 (p < 0.05) (Fig. 3B). VDR levels were also analyzed in xenograft tumors by Western blot, showing a 60% increase in the xenografts from BXL0124 treated mice compared to the control xenograft tumors (Fig. 3C).
Figure 3.

BXL0124 treatment maintains vitamin D receptor levels in MCF10DCIS.com tumors. A. A representative immunohistochemical analysis for VDR from weeks 1, 2, 3, and 4 is shown (40x). VDR-positive staining is found in the cytoplasm and nucleus of the cells. Whole tumor mounts of VDR expression from week 4 is shown as a contracted view. B. Four tumors from each group were blinded and quantified and three representative areas from each tumor were quantified for the intensity of VDR staining, mean +/- S.E.M. The staining intensities were scored from 0+ (negative staining) to 3+ (the strongest staining), * p < 0.05. C. The protein level of VDR was increased in the tumors of week 4 BXL0124 treated mice as shown by western blot analysis. Five xenograft tumors from each group were combined for pooled samples. β-actin was used as a loading control.
Treatment with BXL0124 Inhibits Progression to IDC by Maintaining the Myoepithelial Cell Layer
The main diagnostic feature which distinguishes DCIS from invasive lesions is the loss of the critical myoepithelial cell layer and basement membrane (15). To investigate the integrity of the cell layers throughout progression, we analyzed the myoepithelial marker smooth muscle actin (SMA) and the epithelial marker pancytokeratin (panCK) in xenograft tumors. Tumors at weeks 1 and 2 were composed primarily of epithelial cells as noted by panCK staining. There was a gradual establishment of the myoepithelial cell layer starting at week 2, fully forming around the epithelial cells in both the control and BXL0124 treated group by week 3 (Fig. 4). In the control group tumors, the myoepithelial cell layer spontaneously dissociated from its organized structure by week 4 as the tumors progressed to IDC. However, the course of BXL0124 treatment showed the formation of the myoepithelial cell layer in week 3 and maintained organization of the myoepithelial cell layer at week 4 (Fig. 4).
Figure 4.
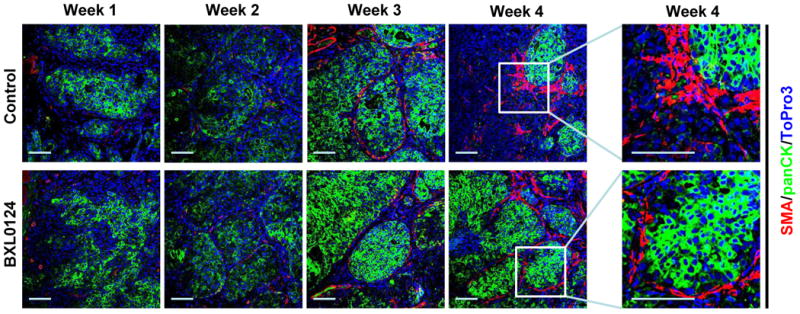
Treatment with BXL0124 inhibits progression to IDC by maintaining the myoepithelial cell layer. A representative immunofluorescence staining for tumor samples from weeks 1, 2, 3, and 4 with the myoepithelial cell marker smooth muscle actin (SMA, shown in red) and the epithelial cell marker pancytokeratin (panCK, shown in green) is shown (200x). Nuclei were stained with TO-PRO-3 (blue). Expanded magnification is shown for specific areas from week 4 tumors. Scale bars represent 100 μm.
Treatment with BXL0124 Maintains the Basement Membrane
To assess the effects of BXL0124 on the integrity of the basement membrane, co-immunofluoresence staining was carried out with a basement membrane marker, laminin 5, and a myoepithelial marker, SMA. The basement membrane formed by week 2, and was maintained in week 3 in both the control and BXL0124 treated groups. At week 4, the basement membrane is disrupted in the invasive-like tumors of the control group, as indicated by reduced and fragmented staining of laminin 5. This structure remained intact in the BXL0124 treated tumors at week 4 (Fig. 5).
Figure 5.
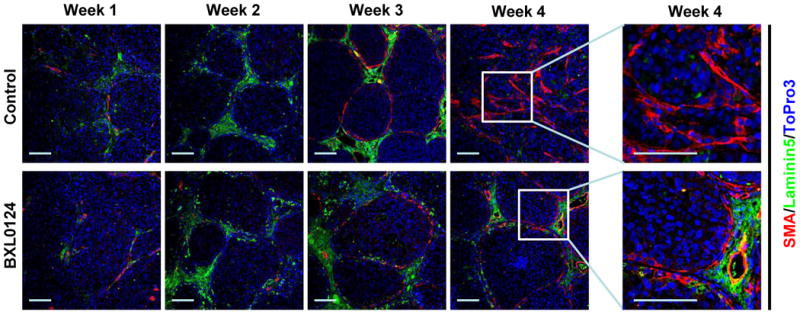
Treatment with BXL0124 maintains the basement membrane. A representative immunofluorescence staining with the myoepithelial cell marker SMA (shown in red) and the basal membrane marker laminin 5 (shown in green) on tumors from weeks 1, 2, 3, and 4 is shown (200x). Nuclei were stained with TO-PRO-3 (blue). Expanded magnification is shown for specific areas from week 4 tumors. Scale bars represent 100 μm.
BXL0124 Inhibits the mRNA Expression of the Matrix Metalloproteinases During DCIS to IDC Progression
Matrix metalloproteinases (MMPs) have been implicated in increased tumor growth, invasion, and metastasis (34, 35). Consequently, we assessed the mRNA levels of specific MMPs in MCF10DCIS.com tumor xenografts at weeks 3 and 4. Analysis of MCF10DCIS.com tumors showed a significant decrease in the mRNA levels of MMP2, 9, 14, and 15 upon BXL0124 treatment at week 3 while MMP16 did not change (Fig. 6A). The modulation of MMP2 and MMP14 expression by BXL0124 treatment persisted to week 4. These observations suggest that reduction of MMP expression by BXL0124 contributes to the inhibition of transition from DCIS to invasive carcinomas. Interestingly, despite the changes in protein levels of VDR we did not see a significant change in mRNA expression of the VDR gene at weeks 3 or 4 upon BXL0124 treatment (Fig. 6A). We did not detect significant biological changes in the vitamin D metabolizing genes CYP24A1 (catabolism) or CYP27B1 (synthesis) in week 3 and 4 xenograft samples (Supplemental Fig. 3). In order to determine if VDR is expressed in myoepithelial cells, we assessed the immunofluorescence staining of VDR together with the myoepithelial marker smooth muscle actin (SMA). VDR staining did not co-localize with SMA staining in any of the samples that were analyzed, indicating that VDR is not expressed in myoepithelial cells (Fig. 6B). In addition, the loss of VDR is evident in week 4 control xenografts, while VDR levels are maintained upon BXL0124 treatment at week 4 (Fig. 6B).
Figure 6.

BXL0124 inhibits the mRNA expression levels of the matrix metalloproteinases during DCIS to IDC progression. A. qPCR analysis of MMPs and VDR mRNA levels in MCF10DCIS.com tumor samples from weeks 3 and 4 is shown, mean +/- S.E.M., Cycle numbers are shown in parenthesis: MMP2 (25), MMP9 (28), MMP14 (23), MMP15 (28), MMP16 (25), and VDR (26). Statistical significance refers to the respective week control, * p < 0.05, ** p < 0.01, *** p < 0.001 (n=3-5 per group). B. A representative immunofluorescence staining with the myoepithelial cell marker, SMA (shown in red), and VDR (shown in green) on tumors from weeks 3 and 4 are shown (200x). Nuclei were stained with TO-PRO-3 (blue). Scale bars represent 100 μm.
Discussion
DCIS progression to IDC is defined by the escape of inner luminal epithelial cells through the outer layer of myoepithelial cells and the basement membrane, ultimately coming in contact with the stromal cell population (13). Previous reports have shown that myoepithelial cells can arise from the luminal cell population, but not vice versa (13, 36-38). In our study, the MCF10DCIS.com xenografts show growth of the epithelial cells in the early weeks followed by the formation of the myoepithelial cell layer which is consistent with those reports. BXL0124 does not seem to affect the rate of formation of the myoepithelial layer (Fig. 4. Weeks 1, 2, and 3), however it does significantly reduce the rate at which the myoepithelial cell layer is broken down (Fig. 4. Week 4), suggesting that BXL0124 inhibits the transition from DCIS to invasive carcinoma. When the staining of laminin 5 in the DCIS and IDC lesions are examined histologically, it is clear that BXL0124 treatment helps to maintain an intact, organized structure of the critical basement membrane (Fig. 5. Week 4). It is also interesting to note that the basement membrane forms first in week 2 followed by the myoepithelial cell layer at week 3. This suggests that the basement membrane might act as a scaffold for the formation of the myoepithelial cell layer, and the loss of this scaffold through enzymatic degradation could partially account for the disorganization of the myoepithelial cell layer observed in week 4 control tumors. Analyzing the effects of BXL0124 on the progression of DCIS to IDC (Fig. 1 D), we found that the treatment sustains DCIS lesions and prevents progression to IDC through maintenance of the critical myoepithelial cell layer and the basement membrane.
The progression from DCIS to IDC is believed to be provoked largely by the production of proteolytic enzymes (39). MMPs degrade proteins involved in extracellular matrix structure and molecules involved in cell-cell adhesion, which releases epithelial cells from their ordered layers and deregulates cell signaling, ultimately leading to extensive changes in gene transcription (40). MMP2 and MMP9, the gelatinases, are responsible for the degradation of type IV collagen as well as laminin 5, components of the basement membrane (39, 41). MMP2 is secreted in a latent form and requires activation by MMP14 to its pro-MMP2 form (42). Aside from its activating function, MMP14 as well as MMP15 and MMP16 have been shown to directly affect cell invasion by remodeling the basement membrane in vivo (43). Thus, reduction of MMP2, MMP9, MMP14 and MMP15 by BXL0124 in week 3 prior to the transition of DCIS to IDC likely contributes to the maintenance of the basement membrane in the tumor xenografts. This suggests that the down-regulation of MMPs by BXL0124 treatment could play a major role in the preservation of DCIS histology.
As tumors progress to IDC, there is a significant increase in cell proliferation which is consistent with previous findings that stromal and tumor epithelial cell interactions can enhance proliferation (44). The significant reduction of the proliferation rate in tumors from BXL0124 treated animals at week 4 demonstrates the potential of BXL0124 to slow the growth of DCIS epithelial cells. Since the effects of BXL0124 are known to be dependent on VDR (28, 34), we analyzed VDR levels over the course of tumor progression. It was previously shown that a decrease in protein levels of VDR have been correlated with the progression from benign to malignant breast lesions (45). In our study, the BXL0124 treatment not only maintained the integrity of the DCIS structure, but also retained VDR levels in the epithelial cells (Fig. 3A. Week 4). VDR protein levels were higher in BXL0124 treated group when compared to the control group at week 4. However, mRNA expression of VDR was unchanged in weeks 3 or 4, suggesting that loss of VDR in week 4 control tumors is likely due to protein degradation or possible post-translational regulation. Interestingly, loss of DCIS architecture seems to be a determining factor in the loss of VDR expression, suggesting that the interaction with the surrounding stromal cells could play a major role in the down-regulation of VDR. Whether adjacent stromal-to-epithelial cell contact or factors secreted from stromal cells contribute to the loss of VDR needs to be further investigated. Analysis of VDR and SMA co-localization studies show that VDR is not expressed in the myoepithelial cell layer or stroma but within the luminal cells. In invasive tumors, VDR negative cells include both tumor and stroma cells. Taken together, it appears that VDR is expressed in the luminal cell population and this expression is lost upon progression to IDC. This further suggests that BXL0124 does not act directly on the myoepithelial cell layer but may exert its anti-tumor effects through a paracrine mechanism from the luminal cells. The analysis of DCIS revealed increased VDR levels and decreased cell proliferation, providing a prevention strategy to inhibit the early progression of DCIS to IDC with the treatment of vitamin D or its analogs. These data suggest a novel mechanism in which down-regulation of VDR is synchronized with the loss of the critical myoepithelial cell layer and basement membrane.
Breast cancer progression is indicated by the loss of normal tissue architecture allowing invasion into surrounding tissue (46). Our study shows that BXL0124 has the potential to exert its effects indirectly on the myoepithelial cell population and directly on the luminal cell population to inhibit tumorigenesis. In the present study, BXL0124 inhibited the progression of DCIS to IDC by maintaining the integrity of the myoepithelial cell layer and basement membrane. The inhibitory activity of BXL0124 on MMP mRNA expression is likely a contributor to the maintenance of DCIS organization and inhibition of tumor progression. This study with a novel Gemini vitamin D analog BXL0124 in the MCF10DCIS.com model suggests a unique treatment modality preventing the progression of DCIS to an aggressive, invasive-like disease.
Supplementary Material
Acknowledgments
Financial Support: This work was supported in part by the National Institutes of Health National Cancer Institute R01 CA127645 (N. Suh), the National Institute of Environmental Health Sciences Grant ES005022 (H. Zarbl) and The Trustees Research Fellowship Program at Rutgers, The State University of New Jersey (N. Suh).
Footnotes
Conflicts of Interest: Authors have no potential conflicts of interest to disclose.
References
- 1.Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64:9–29. doi: 10.3322/caac.21208. [DOI] [PubMed] [Google Scholar]
- 2.Amir E, Freedman OC, Seruga B, Evans DG. Assessing women at high risk of breast cancer: a review of risk assessment models. J Natl Cancer Inst. 2010;102:680–91. doi: 10.1093/jnci/djq088. [DOI] [PubMed] [Google Scholar]
- 3.Troester MA, Swift-Scanlan T. Challenges in studying the etiology of breast cancer subtypes. Breast Cancer Res. 2009;11:104. doi: 10.1186/bcr2323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Almendro V, Fuster G. Heterogeneity of breast cancer: etiology and clinical relevance. Clin Transl Oncol. 2011;13:767–73. doi: 10.1007/s12094-011-0731-9. [DOI] [PubMed] [Google Scholar]
- 5.Pare R, Yang T, Shin JS, Tan PH, Lee CS. Breast cancer precursors: diagnostic issues and current understanding on their pathogenesis. Pathology. 2013;45:209–13. doi: 10.1097/PAT.0b013e32835f2249. [DOI] [PubMed] [Google Scholar]
- 6.Hu X, Stern HM, Ge L, O'Brien C, Haydu L, Honchell CD, et al. Genetic alterations and oncogenic pathways associated with breast cancer subtypes. Mol Cancer Res. 2009;7:511–22. doi: 10.1158/1541-7786.MCR-08-0107. [DOI] [PubMed] [Google Scholar]
- 7.Walsh T, Lee MK, Casadei S, Thornton AM, Stray SM, Pennil C, et al. Detection of inherited mutations for breast and ovarian cancer using genomic capture and massively parallel sequencing. Proc Natl Acad Sci U S A. 2010;107:12629–33. doi: 10.1073/pnas.1007983107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brinton LA, Sherman ME, Carreon JD, Anderson WF. Recent trends in breast cancer among younger women in the United States. J Natl Cancer Inst. 2008;100:1643–8. doi: 10.1093/jnci/djn344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Virnig BA, Tuttle TM, Shamliyan T, Kane RL. Ductal carcinoma in situ of the breast: a systematic review of incidence, treatment, and outcomes. J Natl Cancer Inst. 2010;102:170–8. doi: 10.1093/jnci/djp482. [DOI] [PubMed] [Google Scholar]
- 10.Page DL, Dupont WD, Rogers LW, Jensen RA, Schuyler PA. Continued local recurrence of carcinoma 15-25 years after a diagnosis of low grade ductal carcinoma in situ of the breast treated only by biopsy. Cancer. 1995;76:1197–200. doi: 10.1002/1097-0142(19951001)76:7<1197::aid-cncr2820760715>3.0.co;2-0. [DOI] [PubMed] [Google Scholar]
- 11.Collins LC, Tamimi RM, Baer HJ, Connolly JL, Colditz GA, Schnitt SJ. Outcome of patients with ductal carcinoma in situ untreated after diagnostic biopsy: results from the Nurses' Health Study. Cancer. 2005;103:1778–84. doi: 10.1002/cncr.20979. [DOI] [PubMed] [Google Scholar]
- 12.Sanders ME, Schuyler PA, Dupont WD, Page DL. The natural history of low-grade ductal carcinoma in situ of the breast in women treated by biopsy only revealed over 30 years of long-term follow-up. Cancer. 2005;103:2481–4. doi: 10.1002/cncr.21069. [DOI] [PubMed] [Google Scholar]
- 13.Gudjonsson T, Adriance MC, Sternlicht MD, Petersen OW, Bissell MJ. Myoepithelial cells: their origin and function in breast morphogenesis and neoplasia. J Mammary Gland Biol Neoplasia. 2005;10:261–72. doi: 10.1007/s10911-005-9586-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pandey PR, Saidou J, Watabe K. Role of myoepithelial cells in breast tumor progression. Front Biosci (Landmark Ed) 2010;15:226–36. doi: 10.2741/3617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hu M, Yao J, Carroll DK, Weremowicz S, Chen H, Carrasco D, et al. Regulation of in situ to invasive breast carcinoma transition. Cancer Cell. 2008;13:394–406. doi: 10.1016/j.ccr.2008.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Grant WB. An ecologic study of dietary and solar ultraviolet-B links to breast carcinoma mortality rates. Cancer. 2002;94:272–81. doi: 10.1002/cncr.10196. [DOI] [PubMed] [Google Scholar]
- 17.Garland CF, Garland FC, Gorham ED, Lipkin M, Newmark H, Mohr SB, et al. The role of vitamin D in cancer prevention. Am J Public Health. 2006;96:252–61. doi: 10.2105/AJPH.2004.045260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Garland CF, Garland FC. Do sunlight and vitamin D reduce the likelihood of colon cancer? Int J Epidemiol. 1980;9:227–31. doi: 10.1093/ije/9.3.227. [DOI] [PubMed] [Google Scholar]
- 19.Garland CF, Gorham ED, Mohr SB, Grant WB, Giovannucci EL, Lipkin M, et al. Vitamin D and prevention of breast cancer: pooled analysis. J Steroid Biochem Mol Biol. 2007;103:708–11. doi: 10.1016/j.jsbmb.2006.12.007. [DOI] [PubMed] [Google Scholar]
- 20.Garland FC, Garland CF, Gorham ED, Young JF. Geographic variation in breast cancer mortality in the United States: a hypothesis involving exposure to solar radiation. Prev Med. 1990;19:614–22. doi: 10.1016/0091-7435(90)90058-r. [DOI] [PubMed] [Google Scholar]
- 21.El Abdaimi K, Dion N, Papavasiliou V, Cardinal PE, Binderup L, Goltzman D, et al. The vitamin D analogue EB 1089 prevents skeletal metastasis and prolongs survival time in nude mice transplanted with human breast cancer cells. Cancer Res. 2000;60:4412–8. [PubMed] [Google Scholar]
- 22.Krishnan AV, Feldman D. Mechanisms of the anti-cancer and anti-inflammatory actions of vitamin D. Annu Rev Pharmacol Toxicol. 2011;51:311–36. doi: 10.1146/annurev-pharmtox-010510-100611. [DOI] [PubMed] [Google Scholar]
- 23.Matthews D, LaPorta E, Zinser GM, Narvaez CJ, Welsh J. Genomic vitamin D signaling in breast cancer: Insights from animal models and human cells. J Steroid Biochem Mol Biol. 2010;121:362–7. doi: 10.1016/j.jsbmb.2010.03.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Swami S, Krishnan AV, Wang JY, Jensen K, Horst R, Albertelli MA, et al. Dietary vitamin D(3) and 1,25-dihydroxyvitamin D(3) (calcitriol) exhibit equivalent anticancer activity in mouse xenograft models of breast and prostate cancer. Endocrinology. 2012;153:2576–87. doi: 10.1210/en.2011-1600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lee HJ, Paul S, Atalla N, Thomas PE, Lin X, Yang I, et al. Gemini vitamin D analogues inhibit estrogen receptor-positive and estrogen receptor-negative mammary tumorigenesis without hypercalcemic toxicity. Cancer Prev Res (Phila) 2008;1:476–84. doi: 10.1158/1940-6207.CAPR-08-0084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.So JY, Wahler JE, Yoon T, Smolarek AK, Lin Y, Shih WJ, et al. Oral Administration of a Gemini Vitamin D Analog, a Synthetic Triterpenoid and the Combination Prevents Mammary Tumorigenesis Driven by ErbB2 Overexpression. Cancer Prev Res (Phila) 2013;6:959–70. doi: 10.1158/1940-6207.CAPR-13-0087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Anzano MA, Smith JM, Uskokovic MR, Peer CW, Mullen LT, Letterio JJ, et al. 1 alpha,25-Dihydroxy-16-ene-23-yne-26,27-hexafluorocholecalciferol (Ro24-5531), a new deltanoid (vitamin D analogue) for prevention of breast cancer in the rat. Cancer Res. 1994;54:1653–6. [PubMed] [Google Scholar]
- 28.So JY, Lee HJ, Smolarek AK, Paul S, Wang CX, Maehr H, et al. A novel Gemini vitamin D analog represses the expression of a stem cell marker CD44 in breast cancer. Mol Pharmacol. 2011;79:360–7. doi: 10.1124/mol.110.068403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Miller FR, Santner SJ, Tait L, Dawson PJ. MCF10DCIS.com xenograft model of human comedo ductal carcinoma in situ. J Natl Cancer Inst. 2000;92:1185–6. doi: 10.1093/jnci/92.14.1185a. [DOI] [PubMed] [Google Scholar]
- 30.So JY, Lee HJ, Kramata P, Minden A, Suh N. Differential expression of key signaling proteins in MCF10 cell lines, a human breast cancer progression model. Mol Cell Pharmacol. 2012;4:31–40. [PMC free article] [PubMed] [Google Scholar]
- 31.Maehr H, Lee HJ, Perry B, Suh N, Uskokovic MR. Calcitriol derivatives with two different side chains at C-20. V. Potent inhibitors of mammary carcinogenesis and inducers of leukemia differentiation. J Med Chem. 2009;52:5505–19. doi: 10.1021/jm900780q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee HJ, Ju J, Paul S, So JY, DeCastro A, Smolarek A, et al. Mixed tocopherols prevent mammary tumorigenesis by inhibiting estrogen action and activating PPAR-gamma. Clin Cancer Res. 2009;15:4242–9. doi: 10.1158/1078-0432.CCR-08-3028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lee HJ, Liu H, Goodman C, Ji Y, Maehr H, Uskokovic M, et al. Gene expression profiling changes induced by a novel Gemini Vitamin D derivative during the progression of breast cancer. Biochem Pharmacol. 2006;72:332–43. doi: 10.1016/j.bcp.2006.04.030. [DOI] [PubMed] [Google Scholar]
- 34.So JY, Smolarek AK, Salerno DM, Maehr H, Uskokovic M, Liu F, et al. Targeting CD44-STAT3 signaling by Gemini vitamin D analog leads to inhibition of invasion in basal-like breast cancer. PLoS One. 2013;8:e54020. doi: 10.1371/journal.pone.0054020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kessenbrock K, Plaks V, Werb Z. Matrix metalloproteinases: regulators of the tumor microenvironment. Cell. 2010;141:52–67. doi: 10.1016/j.cell.2010.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Gudjonsson T, Villadsen R, Nielsen HL, Ronnov-Jessen L, Bissell MJ, Petersen OW. Isolation, immortalization, and characterization of a human breast epithelial cell line with stem cell properties. Genes Dev. 2002;16:693–706. doi: 10.1101/gad.952602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Pechoux C, Gudjonsson T, Ronnov-Jessen L, Bissell MJ, Petersen OW. Human mammary luminal epithelial cells contain progenitors to myoepithelial cells. Dev Biol. 1999;206:88–99. doi: 10.1006/dbio.1998.9133. [DOI] [PubMed] [Google Scholar]
- 38.Smalley MJ, Titley J, O'Hare MJ. Clonal characterization of mouse mammary luminal epithelial and myoepithelial cells separated by fluorescence-activated cell sorting. In Vitro Cell Dev Biol Anim. 1998;34:711–21. doi: 10.1007/s11626-998-0067-0. [DOI] [PubMed] [Google Scholar]
- 39.Duffy MJ, Maguire TM, Hill A, McDermott E, O'Higgins N. Metalloproteinases: role in breast carcinogenesis, invasion and metastasis. Breast Cancer Res. 2000;2:252–7. doi: 10.1186/bcr65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Radisky ES, Radisky DC. Matrix metalloproteinase-induced epithelial-mesenchymal transition in breast cancer. J Mammary Gland Biol Neoplasia. 2010;15:201–12. doi: 10.1007/s10911-010-9177-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Giannelli G, Falk-Marzillier J, Schiraldi O, Stetler-Stevenson WG, Quaranta V. Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5. Science. 1997;277:225–8. doi: 10.1126/science.277.5323.225. [DOI] [PubMed] [Google Scholar]
- 42.Sato H, Takino T, Okada Y, Cao J, Shinagawa A, Yamamoto E, et al. A matrix metalloproteinase expressed on the surface of invasive tumour cells. Nature. 1994;370:61–5. doi: 10.1038/370061a0. [DOI] [PubMed] [Google Scholar]
- 43.Hotary K, Li XY, Allen E, Stevens SL, Weiss SJ. A cancer cell metalloprotease triad regulates the basement membrane transmigration program. Genes Dev. 2006;20:2673–86. doi: 10.1101/gad.1451806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schnitt SJ. The transition from ductal carcinoma in situ to invasive breast cancer: the other side of the coin. Breast Cancer Res. 2009;11:101. doi: 10.1186/bcr2228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Lopes N, Sousa B, Martins D, Gomes M, Vieira D, Veronese LA, et al. Alterations in Vitamin D signalling and metabolic pathways in breast cancer progression: a study of VDR, CYP27B1 and CYP24A1 expression in benign and malignant breast lesions. BMC Cancer. 2010;10:483. doi: 10.1186/1471-2407-10-483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bombonati A, Sgroi DC. The molecular pathology of breast cancer progression. J Pathol. 2011;223:307–17. doi: 10.1002/path.2808. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.