

Abstract
Background and objectives
Recent data indicate that extinguished fear often returns when the testing conditions differ from those of treatment. Several manipulations including extensive extinction training, extinction in multiple contexts, and spacing the extinction trials and sessions reduce the return of fear. Moreover, extensive extinction and extinction in multiple context summated, and the spacing of the extinction trials and sessions summated to further reduce the return of fear. Here we evaluated whether these techniques also attenuate the context specificity of latent inhibition, and whether they summate to further decrease fear responding at test.
Methods
In two experiments, with rats as subjects in a lick suppression preparation, we assessed the effects of massive CS preexposure, CS preexposure in multiple contexts, and of spacing the CS-preexposure trials and sessions, in reducing the context specificity of latent inhibition.
Results
Fear responding was attenuated by all four manipulations. Moreover, extensive CS preexposure in multiple contexts, and conjoint spacing of the CS-preexposure trials and sessions, were more effective in reducing the context specificity of latent inhibition than each manipulation alone.
Limitations
Our experimental designs evaluated degrees of context specificity of latent inhibition but omitted groups in which latent inhibition was assessed without a context shift away from the context of latent inhibition treatment. This precluded us from drawing conclusions concerning absolute (as opposed to relative) levels of recovery from latent inhibition.
Conclusions
Techniques effective in decreasing the return of conditioned fear following extinction are also effective in decreasing the context specificity of latent inhibition in an animal model of anxiety. Fear and anxiety disorders might be prevented in anxious human participants with the same techniques used here, but that is still an empirical question.
Keywords: Extinction, Exposure therapy, Latent inhibition, Context specificity of latent inhibition, Return of fear
Introduction
In a fear-conditioning preparation a neutral stimulus (conditioned stimulus; CS; e.g., a light) is presented just prior to an aversive event (unconditioned stimulus; US; e.g., a mild footshock) during acquisition training, which gives the CS behavioral control over fear responses elicited previously solely by the US. Presumably a CS–US fear association underlies such learning. Given that fear associations of this kind are believed to be involved in the onset of pathological fear and anxiety disorders in humans (e.g., Field, 2006; Laborda, Miguez, Polack, & Miller, 2012; Mineka & Oehlberg, 2008), studying manipulations that attenuate fear responding in animal models may be informative to clinicians interested in developing better approaches to prevent and/or eliminate fear and anxiety disorders.
Fear responding to a CS is susceptible to reduction through extinction treatment which consists of presentations of the CS alone (e.g., Pavlov, 1927); however, the effect of extinction treatment is labile and extinguished CSs are likely to elicit fear responding once again under a number of circumstances. For example, an extinguished CS elicits more fear responding when, after acquisition training and extinction treatment in two different contexts, it is tested back in the acquisition context or in a third context (ABA and ABC renewal; e.g., Bouton & Bolles, 1979; Bouton & King, 1983; Rosas, García-Gutiérrez, & Callejas-Aguilera, 2007), than when it is tested in the extinction context. Similarly, an extinguished CS elicits more fear responding when it is tested a long time following extinction treatment than when it is tested soon following this manipulation (spontaneous recovery; Pavlov, 1927). Thus, fear conditioning models the acquisition of anxiety disorders, fear extinction models exposure therapy, and the return of fear following context changes and/or the passage of time models relapse after exposure therapy (e.g., Bouton, 1988; Bouton & Nelson, 1998; Laborda, McConnell, & Miller, 2011).
Research on extinction has identified some behavioral manipulations that attenuate the return of fear (for reviews, see Bouton, Woods, Moody, Sunsay, & García-Gutiérrez, 2006; Laborda et al., 2011). For example, the return of fear has been attenuated in our fear-conditioning preparation with rats as experimental subjects by: a) using a massive number of extinction trials (Denniston, Chang, & Miller, 2003), b) delivering extinction treatment in multiple contexts (Gunther, Denniston, & Miller, 1998), c) spacing the extinction trials (Urcelay, Wheeler, & Miller, 2009), and d) spacing the extinction sessions (Laborda, Miguez, & Miller, 2014). Moreover, Laborda et al. (2014) provided evidence suggesting that the mechanisms underlying the effects of session spacing are different from those of trial spacing.
Recently, we found that these manipulations not only decrease the return of fear but also that their effects summate to better reduce the reappearance of fear responses. Laborda and Miller (2013) replicated and extended Denniston et al.’s (2003) protective effect of massive numbers of extinction trials and Gunther et al.’s (1998) protective effect of extinction being administered in multiple contexts, in a preparation in which spontaneous recovery and renewal otherwise summated to produce strong return of fear of the CS (see Rosas & Bouton, 1998; Rosas, Vila, Lugo, & López, 2001). They found that extended extinction treatment in multiple contexts was more effective in attenuating the return of fear than each of these techniques by itself. Similarly, Laborda et al. (2014) replicated and extended Urcelay et al.’s (2009) trial spacing effect, also in a preparation in which renewal and spontaneous recovery summated eliciting strong return of fear. More importantly, they found that conjoint spacing of extinction trials and spacing of extinction sessions was more effective in attenuating the return of fear than either treatment alone.
Just as fear responding is reduced by presentations of the CS alone after acquisition training (i.e., extinction; e.g., Pavlov, 1927), presentations of the CS alone before acquisition training also reduce fear responding (i.e., latent inhibition, also known as the CS-preexposure effect; e.g., Lubow, 1973a; Lubow & Moore, 1959). Moreover, just as extinction seems to be specific to the context in which it occurs, responding to a latently inhibited CS seems to be minimized when testing occurs in the context of preexposure. The context specificity of latent inhibition is also evidenced when subjects receive preexposure treatment and acquisition training in two different contexts and are tested back in the acquisition context or in a novel context, relative to subjects tested in the context of latent inhibition treatment (e.g., Bailey & Westbrook, 2008; Maes, 2002). The observed context specificity of latent inhibition is consistent with the view that CS preexposure creates a memory of the nonreinforced CS that later competes with the memory of the CS being reinforced (i.e., the CS–US association) and contradicts the initial view of latent inhibition that CS preexposure simply retards subsequent acquisition of the CS–US association (for a theoretical review, see Escobar & Miller, 2012).
Just as extinction of fear conditioning models exposure therapy, latent inhibition of fear conditioning can be viewed as a technique that prevents the development and/or the expression of conditioned fear, and it potentially models a means of preventing anxiety disorders (e.g., Lubow, 1973b, 1998). For instance, cues apt to be presented in the presence of expected traumatic events could be preexposed to attenuate the formation of aversive associations, which might reduce the likelihood of developing pathological fear reactions to these events or at least reduce the strength of the expression of such reactions (e.g., prevention of posttraumatic stress disorder in soldiers). At least in certain circumstances, one might want latent inhibition to generalize to contexts and times other that those of latent inhibition treatment; hence, as with the return of fear, the context specificity of latent inhibition is an effect we might want to thwart in applied settings.
Can the context specificity of latent inhibition be prevented or at least attenuated through the use of techniques similar to those that have proven successful in attenuating the return of fear after extinction treatment? The small existing literature concerning this suggests these manipulations might prove effective in making latent inhibition more enduring. For example, Wheeler, Chang, and Miller (2003) found that both massive CS-preexposure treatment and CS preexposure in multiple contexts attenuated the contextual specificity of latent inhibition. In the present research, we evaluated whether techniques that summate to reduce the return of fear after extinction, massive nonreinforcement in multiple contexts, and conjoint spacing of nonreinforcement trials and sessions, also summate to prevent or at least attenuate the context specificity of latent inhibition.
Experiment 1
The purpose of the present series of experiments was to evaluate whether some behavioral manipulations that attenuate the return of conditioned fear produced by a change in context from that of extinction following extinction treatment can also reduce the context specificity of latent inhibition, potentially identifying a number of new parallels between extinction phenomena and latent inhibition phenomena. More specifically, Experiment 1 (see Table 1 for the experimental design) was intended to evaluate whether massive CS-preexposure trials in multiple contexts attenuate the context specificity of CS preexposure, just as massive extinction trials in multiple contexts has shown to reduce the return of fear (Laborda & Miller, 2013). All subjects were exposed to a procedure with CS–US pairings occurring in a context different from the context(s) in which CS preexposure had occurred, and testing occurring in a third distinct context. No control for latent inhibition (i.e., a group in which latent inhibition is assessed without a context shift away from the context of CS preexposure) was included because our goal was not to demonstrate an absolute latent inhibition effect, but to assess differences in degrees of latent inhibition. A 2 × 2 factorial design was used with the two variables being CS preexposure in one or multiple different contexts and the number of preexposure trials being moderate or massive (i.e., many).
Table 1.
Design summary of Experiment 1.
| Groups | Preexposure | Acquisition | Test | ||
|---|---|---|---|---|---|
| Single/Moderate | 30 X− (A) | 30 X− (A) | 30 X− (A) | 3 X+ (D) | X (E) |
| (B) | (B) | (B) | |||
| (C) | (C) | (C) | |||
| Single/Massive | 150 X− (A) | 150 X− (A) | 150 X− (A) | 3 X+ (D) | X (E) |
| (B) | (B) | (B) | |||
| (C) | (C) | (C) | |||
| Multiple/Moderate | 30 X− (A) | (A) | (A) | 3 X+ (D) | X (E) |
| (B) | 30 X− (B) | (B) | |||
| (C) | (C) | 30 X− (C) | |||
| Multiple/Massive | 150 X− (A) | (A) | (A) | 3 X+ (D) | X (E) |
| (B) | 150 X− (B) | (B) | |||
| (C) | (C) | 150 X− (C) | |||
Note: X was a 10-s click train. “+” denotes reinforcement with a mild footshock. “−” denotes no reinforcement. Letters in parenthesis indicate contexts. Context A, B, and C were counterbalanced. Numbers preceding letters indicate number of trials in that phase.
Methods
Subjects
The subjects were 24 male (249–404 g) and 24 female (186–268 g), experimentally naive, Sprague–Dawley descended rats obtained from our own breeding colony. Subjects were randomly assigned to one of four groups (ns = 12; Single/Moderate, Single/Massive, Multiple/Moderate, and Multiple/Massive), counterbalanced within groups for sex. The animals were individually housed in standard hanging stainless steel wire mesh cages in a vivarium maintained on a 16/8 h light/dark cycle. Experimental manipulations occurred near the middle portion of the light phase. The animals received free access to Purina Lab Chow, whereas water availability was limited to 30 min per day following a progressive deprivation schedule initiated one week prior to the start of the study. From the time of weaning until the start of the study, all animals were handled for 30 s, three times per week.
Apparatus
Thirty experimental chambers of three different basic types were used to construct five distinct contexts. Chamber O was a 30-cm long, 30-cm wide, and 27-cm high operant chest. The sidewalls of the chamber were made of stainless steel sheet metal, and the front wall, back wall, and ceiling of the chamber were made of clear Plexiglas. The floor was constructed of 0.3 cm diameter rods, spaced 1.3 cm center-to-center, connected by NE-2 neon bulbs that allowed a constant-current footshock to be delivered by means of a high voltage AC circuit in series with a 1.0-MΩ resistor. Each of twelve copies of Chamber O was housed in an environmental isolation chest that was dimly illuminated by a house light (1.12-Watt, #1820 incandescent bulb) mounted on one wall of the experimental chamber. Each Chamber O could be equipped with a water-filled lick tube that extended 1-cm into a cylindrical niche, which was 4.5 cm in diameter, left right centered, with its bottom 1.75-cm above the floor of the apparatus and 5.0 cm deep. There was a photobeam detector 1-cm in front of the lick tube that was broken whenever the subject licked the tube.
Chamber V was a 27-cm long box in a truncated-V shape (29.5-cm height, 21.5-cm wide at top, and 5.5-cm wide at bottom). The floor was comprised of two 27-cm long, 2-cm wide stainless plates, with a 1.5-cm gap between the two plates. A constant-current footshock could be delivered through the metal walls and floor of the chamber. The ceiling was clear Plexiglas, the front and back walls were black Plexiglas, and the sidewalls were stainless steel. Each of six copies of Chamber V was housed in a separate sound- and light-attenuating environmental isolation chest. The chamber was illuminated by a 7-W (nominal at 120 VAC, but driven at 50 VAC) light bulb, which was mounted on the inside wall of the environmental enclosure, approximately 30-cm from the center of the experimental chamber. The light entered the chamber primarily by reflection from the ceiling of the environmental chest.
Chamber R was rectangular, measuring 24.0 × 9.0 × 12.5 cm (l × w × h). The walls and ceiling of Chamber R were clear Plexiglas, and the floor was comprise of stainless steel rods measuring 0.5-cm diameter, spaced 1.3-cm apart (center to center). The rods were connected by NE-2 bulbs, which allowed the delivery of a constant-current footshock. Each of twelve copies of Chamber R was housed in separate light- and sound-attenuating environmental isolation chambers. Each chamber was dimly illuminated by a 2-W (nominal at 120 VAC, but driven at 50 VAC) incandescent house light mounted on an inside wall of the environmental chest located approximately 30-cm from the animal enclosure.
A 45-Ω speaker on one inside wall of the isolation chests could deliver a click train (6 Hz, 6 dB above background). Ventilation fans in each enclosure provided a constant 76-dB background noise. All auditory cues were measured on the C-scale. The light intensities inside the three chambers were approximately equal due to the differences in opaqueness of the walls of Chambers V, R, and O.
A 10-s click train served as CS X and a 0.7-mA, 0.5-s footshock served as the US. Five physical contexts were used, three for CS-preexposure sessions (Contexts A, B, and C), one for fear acquisition training (Context D), and one for testing (Context E). The physical contexts used as Contexts A, B, and C were counterbalanced within groups. The three CS-preexposure contexts were a) an instance of Chamber R with house light (HL) off, with a block of wood with two drops of 98% methyl salicylate inside the isolation chest, and with Plexiglas covering the grid floor, b) an instance of Chamber R different from the one used in (a) with HL on, no odor cue added, and no Plexiglas floor, and c) an instance of Chamber V with HL on. Context D consisted of an instance of Chamber O with HL off and a block of wood with two drops of lemon essence located inside the isolation chest. Finally, Context E was an instance of Chamber O (different from the one used as Context D) with HL on, and Plexiglas covering the grid floor. No odor cue was used in this context.
Procedures
See Table 1 for the experimental design of Experiment 1.
Acclimation
On Day 1, all subjects were acclimated to Context E (the test context) in a 30-min session. In this session, subjects had free access to water-filled lick tubes and no nominal stimuli were programmed to occur.
Preexposure
On Days 2–4, all subjects received a daily CS-preexposure session in the appropriated CS-preexposure context for that day and one daily session of mere exposure to each of the two CS-preexposure contexts not used for CS preexposure on that day, with the order of sessions counterbalanced within groups. Subjects in Condition Single received a daily CS-preexposure session in Context A, and a daily exposure session to each of Contexts B and C. Subjects in Condition Multiple received a daily CS-preexposure session in three different contexts (Context A on Day 2, Context B on Day 3, and Context C on Day 4) and a daily exposure session to the two CS-preexposure contexts not used for CS-preexposure training on that day (Contexts B and C on Day 2, Contexts A and C on Day 3, and Context A and B on Day 4). Condition Moderate received a total of 90 CS-preexposure trials divided equally among the three days (i.e., 30 trials in one of the three sessions on each of the three days), while Condition Massive received 450 CS-preexposure trials divided equally among the three days (i.e., 150 trials in one of the three sessions on each of the three days). CS-preexposure sessions included nonreinforced presentations of CS X, while context exposure sessions (which were administered to equally expose subjects to all three of the pre-exposure contexts) did not include presentations of CS X. All pre-exposure sessions were 40 min in duration and there was a 10-min intersession interval. For all groups, CS-preexposure trials had an intertrial interval (ITI) of 6 s from CS termination to CS onset. To accomplish this, while avoiding confounding session duration and trial spacing, Condition Moderate received its CS-preexposure trials in a block at the end of the CS-preexposure sessions, whereas Condition Massive received its CS-preexposure trials throughout its CS-preexposure sessions.
Acquisition
On Day 5, all subjects received 3 presentations of X coterminating with the US in a 45-min conditioning session in Context D. The reinforced trials occurred at 4, 18, and 30 min into the session.
Reacclimation
On Day 6, all subjects were reacclimated in two 30-min sessions in Context E. In these sessions, subjects had free access to water-filled lick tubes and no nominal stimuli were programmed to occur. The purpose of these sessions was to reestablish stable drinking behavior (which might have been differentially disrupted by the footshock US), thereby providing similar baseline behavior across all groups upon which conditioned lick suppression could be assessed.
Tests
On Days 7 and 8, all subjects were tested for conditioned lick suppression to the target cue X in Context E. On each test day, subjects were placed in the test chamber and time spent drinking was recorded. Immediately after completion of an initial 5 cumulative seconds of licking in the absence of any nominal stimulus, subjects were presented with CS X for 10 min. Thus, all subjects were drinking at the time of CS onset. Time to complete an additional 5 cumulative seconds of licking in the presence of the target cue was recorded and interpreted as reflecting fear induced in the subject by the CS. The test sessions were 11 min in duration with a ceiling score of 10 min to complete 5 cumulative seconds of drinking in the presence of the target cue. All test scores (i.e., time to complete 5 s of licking prior to CS onset and time to complete 5 s of drinking in the presence of the CS) were converted to log10 to better approximate the within-group normal distributions assumed by parametric statistical tests. Two test days were used because suppression across groups on the first test day tended to be quite high, potentially allowing a ceiling effect to mask differences between the groups. Following the convention of our laboratory, all animals that took more than 60 s to complete their first 5 cumulative seconds of licking (i.e., prior to CS onset) during the test sessions were scheduled to be eliminated from the study because such long latencies were considered indicative of unusually great fear of the test context.
Results and discussion
The results of Experiment 1 are depicted in Fig. 1. In this experiment relatively strong fear expression was expected in all subjects because they were trained in a design that encouraged the context specificity of the CS-preexposure effect; however, in three of the four groups we observed some attenuation of fear expression relative to equivalently treated subjects that received no CS-alone presentations (e.g., McConnell, Wheeler, Urcelay, & Miller, 2009; Experiment 1; such a group that yielded a mean of 2.0 log s). Subjects in Group Single/Moderate exhibited numerically the strongest fear across tests, followed by subjects in Group Single/Massive, Group Multiple/Moderate, and Group Multiple/Massive, in that order. Importantly, when both behavioral manipulations were administered together in the Multiple/Massive group, the expression of fear was weaker than when either manipulation was administered alone. The following statistical analyses support these conclusions.
Fig. 1.
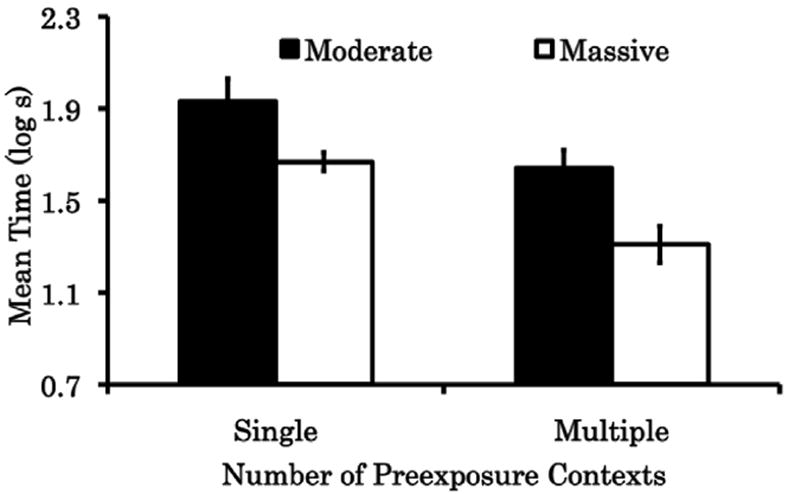
Results of Experiment 1. Mean log10 time to complete 5 cumulative seconds of licking in the presence of the preexposed CS X pooled across two tests (Days 7 and 8). Single = condition that received CS-preexposure trials in a single context (A); Multiple = condition that received CS-preexposure trials in three different contexts (A, B, and C); Moderate = condition that received a moderate number of CS-preexposure trials (90); Massive = condition that received a massive number of CS-preexposure trials (450). 0.7 log s represents the minimum possible time to complete 5 cumulative second of licking. Brackets represent the standard error of the means. Higher scores indicate more conditioned fear. See text and Table 1 for further details.
One animal from Group Multiple/Moderate and two from Group Single/Massive were eliminated from the study because they met the elimination criterion of displaying an unusually great fear of the test context (i.e., they took more than 60 s to complete 5 cumulative seconds of licking prior to CS onset in the first test session). A 2 (Contexts of preexposure [between subjects]: Single vs. Multiple) × 2 (Number of preexposure trials [between subjects]: Moderate vs. Massive) × 2 (Tests [within subjects]: 1 vs. 2) analysis of variance (ANOVA) applied to the pre-CS test data (in log s) showed no significant main effect nor interaction (all ps > .05), indicating that the experimental groups did not significantly differ in their baseline fear response. However, the same analysis proved significant when the CS data (in log s) were examined. This analysis showed a main effect of Contexts, F(1, 41) = 16.14, p < .01, MSE = 0.15, Cohen’s f = 0.41, indicating that CS preexposure in multiple context reduced the expression of fear more than to CS preexposure in a single context. The main effect of Number of preexposure trials was also significant, F(1, 41) = 12.88, p < .01, Cohen’s f = 0.36, indicating that a massive number of CS-preexposure trials also reduced the expression of fear relative to a moderate number of CS-preexposure trials. The interaction between these factors did not prove significant, F(1, 41) = 0.18, p = .67, suggesting that these effects summated (instead of interacted) to attenuate fear acquisition. Consistent with the expectation that less conditioned fear would be observed on Test 2 than Test 1 because of extinction of the CS that occurred on Test 1, the main effect of Tests was significant, F(1, 41) = 12.24, p < .01, MSE = 0.15. However, this factor did not significantly interact with any other factor (smallest p = .22).
Two planned comparisons were then performed. A significant difference was found when Group Single/Massive was compared with Group Multiple/Massive, F(1, 41) = 9.64, p < .01, indicating that fear acquisition was attenuated significantly more following massive CS preexposure in multiple contexts than following massive CS preexposure in a single context. A significant difference was also found when Group Multiple/Moderate was compared with Group Multiple/Massive, F(1, 41) = 8.28, p < .01, indicating that fear expression was attenuated more following massive CS preexposure in multiple contexts than following moderate CS preexposure in multiple contexts. These tests corroborate the additive effect of our manipulations in attenuating the context specificity of latent inhibition.
Experiment 2
Experiment 2 was intended to evaluate whether conjoint spacing of the CS-preexposure trials and CS-preexposure sessions summate in attenuating the context specificity of latent inhibition, just as conjoint spacing of extinction trials and extinction sessions have shown to summate in reducing the return of fear (Laborda et al., 2014). A 2 × 2 factorial design was used with one factor being whether CS-preexposure trials were massed or spaced within sessions and the other factor being whether CS-preexposure sessions were massed or spaced (see Table 2 for the experimental design).
Table 2.
Design summary of Experiment 2.
| Groups | Preexposure | Acquisition | Test |
|---|---|---|---|
| MassTrials/MassSessions | 90 X− (A) | 3 X+ (B) | X (C) |
| 6-s intertrial intervals | |||
| 10-min intersession intervals | |||
| MassTrials/SpacSessions | 90 X− (A) | 3 X+ (B) | X (C) |
| 6-s intertrial intervals | |||
| 7-d intersession intervals | |||
| SpacTrials/MassSessions | 90 X− (A) | 3 X+ (B) | X (C) |
| 600-s intertrial intervals | |||
| 10-min intersession intervals | |||
| SpacTrials/SpacSessions | 90 X− (A) | 3 X+ (B) | X (C) |
| 600-s intertrial intervals | |||
| 7-d intersession intervals |
Note: X was a 10-s click train. “+” denotes reinforcement with a mild footshock. “−” denotes no reinforcement. Letters in parenthesis indicate contexts. Numbers preceding letters indicate number of trials in that phase.
Methods
Subjects
The subjects were 48 male (228–381 g) and 48 female (160–240 g), experimentally naive, Sprague–Dawley descended rats obtained from our own breeding colony. Subjects were randomly assigned to one of four groups (ns = 12 in each of two replications; MassTrials/MassSessions, MassTrials/SpacSessions, SpacTrials/MassSessions, and SpacTrials/SpacSessions), counterbalanced within groups for sex. Housing and maintenance of subjects was as in Experiment 1.
Apparatus
As in Experiment 1, a 10-s click train served as CS X and a 0.7-mA 0.5-s footshock served as the US. Twelve each of Chambers R, V, and O served as three distinct contexts, one for CS-preexposure sessions (Context A), one for acquisition training (Context B), and one for testing (Context C). Context A was an instance of Chamber R or V (counterbalanced within groups) with HL on and with no odor cue nor Plexiglas floor. Context B consisted on an instance of Chamber O with HL off, and a block of wood with two drops of lemon essence located inside the isolation chest. No Plexiglas floor was used in Context B. Context C was an instance of Chamber O, different from the one used as Context B, with HL on and Plexiglas floor in place. No odor cue was used in Context C.
Procedures
The present experiment was conducted in two exact replications.
Acclimation
On Day 1, all subjects were acclimated to Context C in a 30-min session as in Experiment 1.
Preexposure
Across Days 2–16, all subjects received a total of 90 CS-preexposure trials in Context A divided equally across three sessions. Subjects in Condition MassTrials received 30 CS-preexposure trials with an ITI (from CS onset to CS onset) of 16 s in three 8-min training sessions. Subjects in Condition SpacTrials received 30 CS-preexposure trials with an ITI of 600 s in three 300-min training sessions. For practical reasons, session duration and trial spacing needed to be confounded in the present experiment; we address this confound on the Result and Discussion subsection (3.2). Subjects in Condition MassSessions received their three CS-preexposure sessions with an intersession interval of 10 min (all on Day 16), while subjects in Condition SpacSessions received their three CS-preexposure sessions with an intersession interval of 7 days (Days 2, 9, and 16). No lick tubes were present during these sessions.
Acquisition, reacclimation, and tests
All subjects received acquisition training in Context B (Day 17) with no lick tube present, reacclimation in Context C (Day 18), and were tested in Context C (Days 19 and 20), respectively. All procedures were the same as in Experiment 1.
Results and discussion
The results of Experiment 2 are depicted in Fig. 2. As in Experiment 1, in this experiment relatively strong fear expression was expected in all subjects because they were trained in a design that encouraged the context specificity of the CS-preexposure effect; however, in three of the four groups we observed some attenuation of fear responding relative to equivalently treated subjects that received no CS-alone presentations (e.g., McConnell et al., 2009; Experiment 1; a group that yielded a mean of 2.0 log s). Subjects in Group MassTrials/MassSessions exhibited numerically the strongest fear across tests, followed by subjects in Group SpacTrials/MassSessions, Group MassTrials/SpacSessions, and Group Spac-Trials/SpacSessions, in that order. Moreover, when both behavioral manipulations were administered together in the SpacTrials/SpacSessions group, the expression of fear was weaker than when either manipulation was administered alone. The following statistical analyses support these conclusions.
Fig. 2.
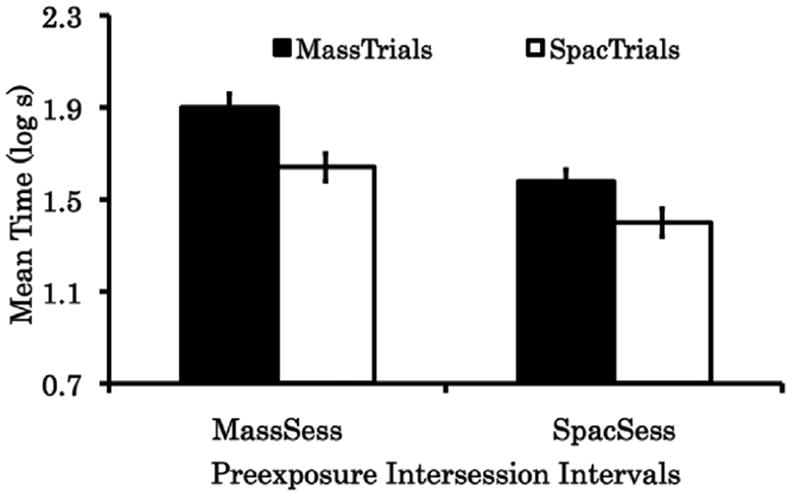
Results of Experiment 2. Mean log10 time to complete 5 cumulative seconds of licking in the presence of the preexposed CS X pooled across two tests (Days 7 and 8). MassTrials = condition that received CS-preexposure trials with short intertrial intervals (6 s); SpacTrials = condition that received CS-preexposure trials with long intertrial intervals (600 s); MassSess = condition that received CS-preexposure trials with short intersession intervals (10 min); SpacSess = condition that received CS-preexposure trials with long intersession intervals (7 days). 0.7 log s represents the minimum possible time to complete 5 cumulative second of licking. Brackets represent the standard error of the means. Higher scores indicate more conditioned fear. See text and Table 2 for further details.
Four animals from Group MassTrials/SpacSessions, one from Group MassTrials/MassSessions, and one from Group SpacTrials/MassSessions were eliminated from the study because of their displaying an unusually great fear of the test context (i.e., took more than 60 s to complete 5 cumulative seconds of licking prior to CS onset in the first test session). As this experiment was conducted in two exact replications, an initial analysis evaluated whether these replications differed with respect to responding to the target cue. To evaluate this, a 2 (Trial spacing [between subjects]: MassTrials vs. SpacTrials) × 2 (Session spacing [between subjects]: MassSessions vs. SpacSessions) × 2 (Replication [between subjects]: 1 vs. 2) × 2 (Tests [within subjects]: 1 vs. 2) ANOVA on the CS suppression scores (log s) was performed and found no significant main effect of replication nor interaction with replication (all ps > .05). Thus, we pooled subjects across replications and this factor was excluded from further analyses.
A 2 (Trial spacing [between subjects]: MassTrials v. SpacTrials) × 2 (Session spacing [between subjects]: MassSessions v. SpacSessions) × 2 (Tests [within subjects]: 1 v. 2) ANOVA applied to the pre-CS test data (log s) showed no significant main effect nor interaction (all ps > .05), indicating that the experimental groups did not significantly differ in their baseline fear response. However, the same analysis proved to be significant when the CS data (log s) was examined. This analysis showed a main effect of Trial spacing, F(1, 86) = 13.09, p < .01, MSE = 0.09, Cohen’s f = 0.26, indicating that spaced CS-preexposure trials successfully reduced behavioral control by CS X relative to massed CS-preexposure trials. The main effect of Session spacing was also significant, F(1, 86) = 21.58, p < .01, Cohen’s f = 0.34, indicating that spaced CS-preexposure sessions also reduced behavioral control by CS X relative to massed CS-preexposure sessions. Additionally, the interaction between these factors did not prove significant, F(1, 86) = 0.48, p = .49, suggesting that these effects summated (instead of interacted) to attenuate behavioral control by CS X. The main effect of Tests was also significant, F(1, 86) = 120.51, p < .01, MSE = 0.09. However, this factor did not significantly interact with any other factor (smallest p = .82), which is consistent with the general level of responding being significantly lower in Test 2 than Test 1 due to extinction during Test 1, but maintaining the same pattern of results.
Two planned comparisons were then performed. A significant difference was found when Group MassTrials/SpacSessions was compared with Group SpacTrials/SpacSessions, F(1, 86) = 4.17, p < .05, indicating that fear responding was attenuated more following spaced CS-preexposure trials than following massed CS-preexposure trials in spaced CS-preexposure sessions. A significant difference was also found when Group SpacTrials/MassSessions was compared with Group SpacTrials/SpacSessions, F(1, 86) = 8.20, p < .01, indicating that fear responding was attenuated more following spaced CS-preexposure sessions than following massed CS-preexposure sessions given spaced extinction trials. Both of these tests corroborate the additive effect of our manipulations in attenuating the expression of fear associations.
As mentioned earlier, for practical reasons session duration and trial spacing were confounded in the present experiment, which might seem to complicate interpretation of the trial spacing effect. However, latent inhibition has been found to hinge on associations between the CS-preexposure context and the CS, and exposure to the CS-preexposure context alone during long ITIs constitutes context extinction, which has been found to weaken latent inhibition (Escobar, Arcediano, & Miller, 2002; Graham, Barnet, Gunther, & Miller, 1994). Hence, if the longer CS-preexposure session duration in the SpacTrials condition were the critical variable, it should have reduced latent inhibition, whereas we observed enhanced latent inhibition, consistent with better learning during the latent inhibition treatment as a result of trial spacing. This conforms with the general observation that the spacing of training trials enhances the consequences of training (e.g., Barela, 1999; Urcelay et al., 2009).
General discussion
Fear responding can be thwarted by pre-acquisition exposure to a to-be-conditioned stimulus. However, just as reduced fear responding after extinction treatment is labile and context specific, reduced fear responding after CS preexposure is labile and context shifts enhance responding to such preexposed cues (e.g., Bailey & Westbrook, 2008; Maes, 2002). Moreover, latent inhibition is susceptible to enhancement by some experimental manipulations, which might translate into less context specificity of latent inhibition when subjects are tested outside the context of CS preexposure. For example, Leung, Killcross, and Westbrook (2011) have recently reported that additional compound preexposure (i.e., A and B presented simultaneously) to elementally preexposed cues (i.e., A and B present separately) enhances the effect of elemental preexposure, decreasing conditioned fear responding to A more than does additional preexposure to the target cue alone (i.e., A). Future research should evaluate whether this manipulation is also capable of reducing the context specificity of latent inhibition.
In the present series of experiments, four manipulations were evaluated in their effectiveness in reducing the context specificity of latent inhibition. In Experiment 1, all subjects were given CS-preexposure trials followed by Pavlovian fear conditioning in a situation in which strong fear responding was expected due to testing occurring outside the context of preexposure (i.e., in a neutral but familiar context). However, three of the groups were expected to show reduced fear responding relative to Group Single/Moderate because they received massive preexposure trials (Group Single/Massive), preexposure in multiple contexts (Group Multiple/Moderate), or both of these manipulations concurrently (Group Multiple/Massive). Just as these manipulations were effective in attenuating the return of fear following extinction (e.g., Laborda & Miller, 2013), these manipulations were effective in attenuating the context specificity of latent inhibition, thereby replicating Wheeler et al.’s (2003) results and furthermore demonstrating that extensive CS-preexposure and CS-preexposure in multiple context summate to reduce the context specificity of latent inhibition. Importantly, our results are novel in demonstrating that extensive preexposure treatment in multiple contexts is more effective in thwarting fear responding than either manipulation alone (i.e., giving additional preexposure trials or giving preexposure in multiple contexts).
In Experiment 2, all subjects were also given CS-preexposure trials followed by Pavlovian fear conditioning in a situation in which strong fear responding was expected again due to testing occurring outside of the context of latent inhibition treatment. However, three of the groups were expected to show reduced fear responding relative to Group MassTrials/MassSessions because they received spaced CS-preexposure sessions (Group MassTrials/SpacSessions), spaced CS-preexposure trials (Group SpacTrials/MassSessions), or both manipulations concurrently (Group Spac-Trials/SpacSessions). Just as these manipulations were effective in attenuating the return of fear following extinction (e.g., Laborda et al., 2014), these manipulations were effective in attenuating the context specificity of latent inhibition. Critically, our results suggest that conjoint spacing of CS-preexposure trials and spacing of CS-preexposure sessions is more effective in thwarting fear responding than either manipulation alone. Summation of these two effects was expected but not certain, as spacing trials or spacing sessions could have had asymptotic effects, not leaving room for evidencing the effects of a second manipulation.
Extensive (i.e., massive) CS-preexposure treatment and the spacing of CS-preexposure trials might be considered manipulations that fortify what is learned during nonreinforcement, whereas CS-preexposure treatment in multiple physical contexts and spacing the CS-preexposure sessions might arguably be considered manipulations that increase the presence of facilitatory cues from nonreinforcement at testing (Laborda & Miller, 2012; Miller & Laborda, 2011). Using multiple physical contexts during CS preexposure likely increases the number of potential features of the contexts of nonreinforcement that may be present if testing occurs in a new context. Moreover, CS preexposure delivered in spaced sessions may work in a similar way. Long intervals between preexposure sessions may create different temporal contexts (e.g., Bouton, 2010), making this technique effectively preexposure treatment in multiple temporal contexts, which should encourage generalization of the latent inhibition learning to new temporal contexts. It is noteworthy that although trial spacing and session spacing superficially sound similar, the fact that added time between trials is time spent in the treatment context but added time between sessions is time spent in the home cage suggests the existence of different mechanisms underlying the effect of these two manipulations (Laborda & Miller, 2012; Miller & Laborda, 2011).
Evaluating manipulations that potentially enhance what is learned during CS preexposure can be of interest for researchers interested in developing effective interventions to prevent or at least attenuate the acquisition of anxiety disorders. Craske and Zucker (2002) recently presented such a model for intervention. For them, a successful approach to prevention would include a careful analysis of risk factors for anxiety disorders, in order to target appropriate populations and potential cues that might come to elicit anxiety (e.g., battle stimuli for soldiers). Craske and Zucker acknowledge the importance of cue preexposure in preventing fear associations with examples including vicarious (e.g., parent modeling nonfearful reactions to a target event) and direct cue preexposure (see also Mineka & Zinbarg, 1996). Along the same lines, Weinstein (1990) suggested that, among other interventions for preventing the development of childhood anxiety to dental procedures, children should receive non-traumatic exposure to the treatment situation prior to invasive interventions. Considering the results presented here, this manipulation should be more effective if many spaced exposure events occur in spaced exposure sessions in multiple contexts; however, this is a recommendation that still needs empirical testing.
Future research should evaluate how to best translate the results reported here to applied situations. Extensive CS preexposure in multiple contexts and conjoint spacing of CS-preexposure trials and spacing of CS-preexposure sessions have the potential to decrease fear responding in humans, but the implementation of such interventions in applied settings has yet to be assessed.
Acknowledgments
role of funding organizations
National Institute of Mental Health Grant 33881 supported this research. Mario A. Laborda was partially funded by Program U-Apoya, University of Chile. The authors thank Cara Burney, Henry Cham, Peter Gerhardstein, Lisa Mash, Gonzalo Miguez, Juan M. Rosas, and Deanne Westerman for their comments on an earlier version of this manuscript.
Footnotes
declaration of interest
The authors declare having no conflicts of interest.
References
- Bailey GK, Westbrook RF. Extinction and latent inhibition of within-event learning are context specific. Journal of Experimental Psychology: Animal Behavior Processes. 2008;34:106–118. doi: 10.1037/0097-7403.34.1.106. [DOI] [PubMed] [Google Scholar]
- Barela PB. Theoretical mechanisms underlying the trial-spacing effect in Pavlovian fear conditioning. Journal of Experimental Psychology: Animal Behavior Processes. 1999;25:177–193. doi: 10.1037//0097-7403.25.2.177. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Context and ambiguity in the extinction of emotional learning: implications for exposure therapy. Behaviour Research and Therapy. 1988;26:137–149. doi: 10.1016/0005-7967(88)90113-1. [DOI] [PubMed] [Google Scholar]
- Bouton ME. The multiple forms of “context” in associative learning theory. In: Mesquita B, Barrett LF, Smith ER, editors. The mind in context. New York, NY: Guilford Press; 2010. pp. 233–258. [Google Scholar]
- Bouton ME, Bolles RC. Contextual control of the extinction of conditioned fear. Learning and Motivation. 1979;10:445–466. [Google Scholar]
- Bouton ME, King DA. Contextual control of the extinction of conditioned fear: tests for the associative value of the context. Journal of Experimental Psychology: Animal Behavior Processes. 1983;9:248–265. [PubMed] [Google Scholar]
- Bouton ME, Nelson JB. The role of context in classical conditioning: some implications for cognitive behavior therapy. In: O’Donohue W, editor. Learning and behavior therapy. Boston, MA: Allyn and Bacon; 1998. pp. 59–84. [Google Scholar]
- Bouton ME, Woods AM, Moody EW, Sunsay C, García-Gutiérrez A. Counteracting the context-dependence of extinction: relapse and tests of some relapse prevention methods. In: Craske MG, Hermans D, Vansteenwegen D, editors. Fear and learning: From basic processes to clinical implications. Washington, DC: American Psychological Association; 2006. pp. 175–196. [Google Scholar]
- Craske MG, Zucker BG. Prevention of anxiety disorders: a model for intervention. Applied & Preventive Psychology. 2002;10:155–175. [Google Scholar]
- Denniston JC, Chang RC, Miller RR. Massive extinction attenuates the renewal effect. Learning and Motivation. 2003;34:68–86. [Google Scholar]
- Escobar M, Arcediano F, Miller RR. Latent inhibition and contextual associations. Journal of Experimental Psychology: Animal Behavior Processes. 2002;28:123–136. [PubMed] [Google Scholar]
- Escobar M, Miller RR. Associative accounts of causal judgments. In: Zentall TR, Wasserman EA, editors. The Oxford handbook of comparative cognition. New York, NY: Oxford University Press; 2012. pp. 157–174. [Google Scholar]
- Field AP. Is conditioning a useful framework for understanding the development and treatment of phobias? Clinical Psychology Review. 2006;26:857–875. doi: 10.1016/j.cpr.2005.05.010. [DOI] [PubMed] [Google Scholar]
- Grahame NJ, Barnet RC, Gunther LM, Miller RR. Latent inhibition as a performance deficit resulting from CS–context associations. Animal Learning & Behavior. 1994;55:1–14. [Google Scholar]
- Gunther LM, Denniston JC, Miller RR. Conducting exposure treatment in multiple contexts can prevent relapse. Behaviour Research and Therapy. 1998;36:75–91. doi: 10.1016/s0005-7967(97)10019-5. [DOI] [PubMed] [Google Scholar]
- Laborda MA, McConnell BL, Miller RR. Behavioral techniques to reduce relapse after exposure therapy: applications of studies of experimental extinction. In: Schachtman TR, Reilly S, editors. Associative learning and conditioning theory: Human and non-human applications. New York, NY: Oxford University Press; 2011. pp. 79–103. [Google Scholar]
- Laborda MA, Miguez G, Miller RR. Preventing return of fear in an animal model of anxiety: Additive effects of spacing extinction trials and sessions. 2014 doi: 10.1016/j.beth.2012.11.001. submitted for publication. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laborda MA, Miguez G, Polack CW, Miller RR. Animal models of psychopathology: historical models and the Pavlovian contribution. Terapia Psicológica. 2012;40:45–59. [Google Scholar]
- Laborda MA, Miller RR. Reactivated memories compete for expression after Pavlovian extinction. Behavioural Processes. 2012;90:20–27. doi: 10.1016/j.beproc.2012.01.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laborda MA, Miller RR. Preventing return of fear in an animal model of anxiety: additive effects of massive extinction and extinction in multiple contexts. Behavior Therapy. 2013;44:249–261. doi: 10.1016/j.beth.2012.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leung HT, Killcross AS, Westbrook RF. Additional exposures to a compound of two preexposed stimuli deepen latent inhibition. Journal of Experimental Psychology: Animal Behavior Processes. 2011;37:394–406. doi: 10.1037/a0023714. [DOI] [PubMed] [Google Scholar]
- Lubow RE. Latent inhibition. Psychological Bulletin. 1973a;79:398–407. doi: 10.1037/h0034425. [DOI] [PubMed] [Google Scholar]
- Lubow RE. Latent inhibition as a means of behavior prophylaxis. Psychological Reports. 1973b;32:1247–1252. doi: 10.2466/pr0.1973.32.3c.1247. [DOI] [PubMed] [Google Scholar]
- Lubow RE. Latent inhibition and behavioral pathology: prophylactic and other possible effects of stimulus preexposure. In: O’Donohue W, editor. Learning and behavior therapy. Boston, MA: Allyn and Bacon; 1998. pp. 107–121. [Google Scholar]
- Lubow RE, Moore AU. Latent inhibition: the effect of nonreinforced preexposure to the conditioned stimulus. Journal of Comparative and Physiological Psychology. 1959;52:415–419. doi: 10.1037/h0046700. [DOI] [PubMed] [Google Scholar]
- Maes JHR. No sex difference in contextual control over the expression of latent inhibition and extinction in Pavlovian fear conditioning in rats. Neurobiology of Learning and Memory. 2002;78:258–278. doi: 10.1006/nlme.2002.4058. [DOI] [PubMed] [Google Scholar]
- McConnell BL, Wheeler DS, Urcelay GP, Miller RR. Protection from latent inhibition provided by a conditioned inhibitor. Journal of Experimental Psychology: Animal Behavior Processes. 2009;35:498–508. doi: 10.1037/a0014627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller RR, Laborda MA. Preventing recovery from extinction and relapse: a product of current retrieval cues and memory strengths. Current Directions in Psychological Science. 2011;20:325–329. [Google Scholar]
- Mineka S, Oehlberg K. The relevance of recent developments in classical conditioning to understanding the etiology and maintenance of anxiety disorders. Acta Psychologica. 2008;127:567–580. doi: 10.1016/j.actpsy.2007.11.007. [DOI] [PubMed] [Google Scholar]
- Mineka S, Zinbarg R. Conditioning and ethological models of anxiety disorders: stress-in-dynamic-context anxiety models. In: Hope DA, editor. Nebraska symposium on motivation: Perspectives on anxiety, panic, and fear. Vol. 43. Lincoln, NE: University of Nebraska Press; 1996. pp. 135–210. [PubMed] [Google Scholar]
- Pavlov IP. In: Conditioned reflexes. Anrep GV, editor. London: Oxford University Press; 1927. [Google Scholar]
- Rosas JM, Bouton ME. Context change and retention interval can have additive, rather than interactive, effects after taste aversion extinction. Psychonomic Bulletin and Reviews. 1998;5:79–83. [Google Scholar]
- Rosas JM, García-Gutiérrez A, Callejas-Aguilera JE. AAB and ABA renewal as a function of the number of extinction trials in conditioned taste aversion. Psicológica. 2007;28:129–150. [Google Scholar]
- Rosas JM, Vila NJ, Lugo M, López L. Combined effect of context change and retention interval on interference in causal judgment. Journal of Experimental Psychology: Animal Behavior Processes. 2001;27:153–164. [PubMed] [Google Scholar]
- Urcelay GP, Wheeler DS, Miller RR. Spacing extinction trials alleviates renewal and spontaneous recovery. Learning & Behavior. 2009;37:60–73. doi: 10.3758/LB.37.1.60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weinstein P. Breaking the worldwide cycle of pain, fear and avoidance: uncovering risk factors and promoting prevention for children. Annals of Behavioral Medicine. 1990;12:141–147. [Google Scholar]
- Wheeler DS, Chang RC, Miller RR. Massive preexposure and preexposure in multiple contexts attenuate the context specific of latent inhibition. Learning & Behavior. 2003;31:378–386. doi: 10.3758/bf03195998. [DOI] [PubMed] [Google Scholar]