

Abstract
Binary toxin (CDT) is frequently observed in Clostridium difficile strains associated with increased severity of C. difficile infection (CDI). CDT belongs to the family of binary ADP-ribosylating toxins consisting of two separate toxin components: CDTa, the enzymatic ADP-ribosyltransferase which modifies actin, and CDTb which binds to host cells and translocates CDTa into the cytosol. CDTb is activated by serine proteases and binds to lipolysis stimulated lipoprotein receptor. ADP-ribosylation induces depolymerization of the actin cytoskeleton. Toxin-induced actin depolymerization also produces microtubule-based membrane protrusions which form a network on epithelial cells and increase bacterial adherence. Multiple clinical studies indicate an association between binary toxin genes in C. difficile and increased 30-d CDI mortality independent of PCR ribotype. Further studies including measures of binary toxin in stool, analyses of CDI mortality caused by CDT-producing strains, and examination of the relationship of CDT expression to TcdA and TcdB toxin variants and PCR ribotypes are needed.
Keywords: Clostridium difficile infection, binary toxin, CDT, mechanism, mortality, disease severity, toxinotyping, PCR ribotyping
Introduction
Clostridium difficile is a human and animal pathogen causing intestinal infections following disturbance of the gut microbiota, usually as a result of prior antibiotic treatment. Since the discovery of C. difficile as the major cause of pseudomembranous colitis (PMC), pathogenesis has been linked to production of toxins and two large, single unit, glucosylating toxins, toxin A (TcdA) and toxin B (TcdB), are considered the main virulence factors. Since 1987 a third toxin, binary toxin, unrelated to the glucosylating toxins has been known to be produced by some C. difficile strains. Binary toxin positive strains were previously infrequently found as a cause of C. difficile infection (CDI) in human populations but have become increasingly prevalent in the past 10 y.1-4 For epidemiological studies C. difficile strains can be typed by PCR ribotyping, pulse field gel electrophoresis (PFGE) or restriction endonuclease analysis (REA).1-3 Strains can be also distributed into toxinotypes (designated by Roman numerals) based on the changes in the toxin A and B coding pathogenicity locus (PaLoc).4 Binary toxin is present only in a minority of PCR ribotypes, PFGE types, and REA types but is found in most variant (non-toxinotype 0) toxinotypes.
The discovery that the human epidemic strain types of C. difficile identified by PCR ribotyping as type 027, by REA as group BI, and PFGE as type NAP1 (referred to collectively as 027/BI/NAP1) produce binary toxin in addition to toxins A and B has stimulated investigation of the possible role of binary toxin in the pathogenesis of CDI.1,2 However, these strain types also possess other changes including high-level fluoroquinolone resistance and presence of an 18 bp deletion and a stop codon in the tcdC gene encoding an anti-sigma factor involved in down regulation of toxin A and B production.1,2 In addition, another epidemic strain type in humans and animals reported in the Netherlands and other countries, PCR ribotype 078, REA group BK, and PFGE type NAP7 or NAP7,8 (078/BK/NAP7,8) also possesses binary toxin and has a deletion and stop codon in the tcdC gene.3-5 While these two clonal strain groupings have been associated with increased disease severity and poor outcome in some settings, it is recognized that these strain type designations have not always correlated with disease severity, particularly in non-epidemic settings. In addition, many other strains carry these genetic alterations, including binary toxin.6 The role of bacterial factors other than toxins A and B that may lead to enhanced virulence is still debated, but recent data on binary toxin, including its mechanism of action and evolving epidemiology suggest re-consideration of the importance of this toxin in the pathogenesis of CDI.
Background and History of Discovery of Binary Toxin CDT
During the initial studies on C. difficile cytotoxicity two large protein toxins were purified and named toxin A (TcdA) and toxin B (TcdB). Clinical studies had confirmed that symptomatic patients were infected with C. difficile strains producing both TcdA and TcdB. Initial focus was therefore on the purification of the toxins and production of antibodies that were subsequently used for development of rapid tests for C. difficile diagnostics. Other studies addressed the effects of both toxins in animal models (hamsters) and the molecular mechanism of action within cells. Several groups have shown that the cell cytoskeleton was primarily affected in toxin treated cells. At that time other clostridial toxins which directly or indirectly modified actin in the cytoskeleton were already known.7,8 They belonged to two groups: clostridial iota toxin-like binary toxins and C2-like toxins, and both were ADP-ribosyltransferases.
To test whether the actin modifying activity of C. difficile TcdA and TcdB is also a result of ADP-ribosyltransferase activity, M. Popoff and coworkers tested several C. difficile strains. ADP-ribosyltransferase activity was discovered in a single strain in addition to cytotoxicity (TcdB) and enterotoxicity (TcdA) and this strain and toxin properties were described in the first reports of C. difficile binary toxin.9,10 Interestingly, these reports described strain CD 196, which was a historical PCR ribotype 027 strain.
The next report on C. difficile binary toxin was not published until 199711 and described the sequence of binary toxin encoding genes, the similarity with iota toxin clostridial binary toxins, and the small proportion of tested clinical strains that had binary toxin genes and/or ADP-ribosylating activity (5 of 24 strains). Three years later binary toxin positive strains were reported in C. difficile strains isolated from horses, but not from cats and dogs.12 In the same year it was reported that binary toxin genes were present in some PCR ribotypes recovered from clinical (human) specimens. According to the number of strains included in different PCR ribotypes in the large Cardiff PCR ribotype library it was estimated that 6.4% of toxigenic strains had the genes for binary toxin, however, not all gene positive strains had detectable ADP-ribosylating activity.13
CdtLoc: Binary Toxin Encoding Locus
The genes encoding binding (CDTb) and enzymatic activity (CDTa) were sequenced in 1997.11 Later the 6.2 kb region was additionally characterized and named the Cdt locus or CdtLoc.14 This region included both toxin genes (cdtA and cdtB) as well as the gene encoding regulatory protein (cdtR). In contrast to the PaLoc (coding for TcdA and TcdB) for which several different truncated versions were known4 it appeared that the CdtLoc was present either as a whole or as a single truncated version.15 Strains that have neither the whole or truncated CdtLoc have a unique 68 bp sequence in this chromosomal location (Fig. 1).14

Figure 1. Schematic representation of the CDT region and flanking genes. The regions from the nontoxigenic isolate CD37 (A), the binary toxin-negative isolate strain 630 (B), and the binary toxin-positive isolate QCD-32 g58 (C) are shown. The positions of the 5′ flanking genes CD2601 and CD2602, the 3′ flanking gene trpS, the response regulator gene cdtR, and the CDT binary toxin-encoding genes cdtAB, or their pseudogenes, are shown. For each variant of the CDT region the positions of the 5′ and 3′ conserved boundaries are shown, and the size of the entire CdtLoc is indicated. The unique 68-bp sequence that is present in CD37 and other nontoxigenic isolates in place of the CdtLoc is shown in bold. Adapted from J. Bacteriol. 2007;89:7290–7301, with permission from American Society for Microbiology.14
Genes for CDTa and CDTb could be variable due to some point mutations but this variability has not been studied as extensively as for TcdA and TcdB toxinotypes. Initially some partial sequences were reported.13,16 Popoff et al. have studied variability in the CDT regulatory gene cdtR and could distinguished five lineages which correlated well with types of two genes involved in regulation of toxins A and B, tcdC and tcdR.17 So far the most detailed study has included ten strains from 6 ribotypes (027, 078 and four local designations) and four toxinotypes (III, IV, V, IX).18 They could differentiate the CdtLoc into three groups which largely overlap with ribotypes and toxinotypes. However, some interesting exceptions were also detected, e.g., one strain determined as toxinotype III/NAP1/ribotype AD (different from 027) grouped with strains from toxinotype IX and not with strains from the toxinotype III/NAP1/027 group.
Basic Description of the Toxin
CDT belongs to the family of binary ADP-ribosylating toxins. Other members of this toxin family are C. botulinum C2 toxin, C. perfringens iota toxin, C. spiroforme toxin and the B. cereus/thuringiensis vegetative insecticidal proteins (VIP).19 All these toxins consist of two separate toxin components. One component is the biologically active component and possesses ADP-ribosyltransferase activity to modify actin and the second component is involved in binding of the toxin to host cells and is responsible for the translocation of the enzyme component into the cytosol.
Structure of CDTa
The enzyme component of CDT (strain CD196, O32738 or Q9KH42;UniProtKB) consists of 463 amino acids with a mass of ~53 kDa11,14,20 (Fig. 2). The first 43 amino acids are probably a signal sequence and are cleaved by proteolysis.11 Therefore, the mature toxin component CDTa has a mass of ~48 kDa. This part exhibits about 84 and 82% sequence identity with the enzyme components of C. perfringens iota toxin (Ib) and C. spiroforme toxin (CSTb), respectively. Other members of the family of binary actin ADP-ribosylating toxins are less related to the enzyme component of CDT with 34 and 29% sequence identity of B. cereus/thuringiensis Vip2 and C. botulinum toxin C2 toxin (C2I), respectively. Nevertheless the overall structures of the enzyme components of these toxins are very similar. CDTa has been crystalized recently.21 As shown for other toxins, CDTa has a 2 domain structure. Both domains exhibit similar folding most likely resulting from duplication of an ancient ADP-ribosyltransferase gene.22 The N-terminal part is covered by residues 1–215 (note, numbering refers to mature toxin) and consist of 5 α-helices and 8 β-strands, which are probably involved in the interaction with the binding component CDTb, whereas the C-terminal part with residues 224–420 carries ADP-ribosyltransferase activity. As in the N-terminus, the C-terminal part consists of 5 α-helices and 8 β-strands. This part contains the catalytic site. Many bacterial ADP-ribosyltransferases, which in toto exhibit only minimal sequence similarity, belong to the so-called R-S-E class of ADP-ribosyltransferases, which share a typical arginine (R) residue, an S-T-S motif and an EXE motif. In CDT, the crucial R is Arg-302. The STS motif starts with Ser-345 and the EXE motif with Glu385. The second Glu387 of the EXE motif is the so called catalytic glutamate.21 Exchange of this residue blocks the ADP-ribosyltransferase activity as well as the NAD-hydrolase activity, which is observed in the presence of H2O as a substrate.23 Recently, the structure of iota toxin, which is highly related to CDT, has been solved in complex with its substrate actin, leading to important insights into the molecular mechanisms of toxin-induced ADP-ribosylation of actin.24
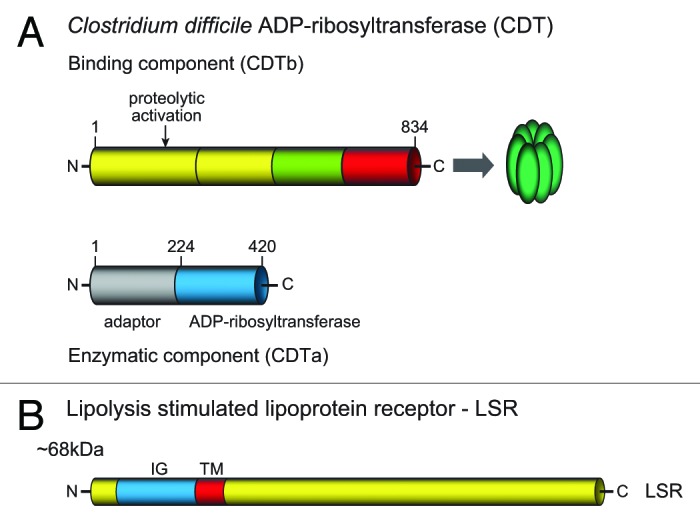
Figure 2. Scheme of the structure of CDT. (A) The binary toxin consists of a binding component (CDTb) and an enzymatic component (CDTa). Both are expressed with leader sequences, which are not shown here. CDTb is activated by proteolytic cleavage to release ~20 and 75 kDa fragments. The 75 kDa fragment is the active binding component, which oligomerizes to form heptamers. (B) The lipolysis stimulated lipoprotein receptor (LRS) is the target cell receptor of CDT. The protein possesses an extracellular part with an immunoglobulin-like structure, a transmembrane region and a large intracellular part.
Structure of CDTb
The binding component of CDTb consists of 876 amino acids with a molecular mass of 98.8 kDa (strain CD196, O32739 or A8DS70, UniProtKB) (Fig. 2). CDTb is about 80, 79, 44 and 36% identical with the binding components of iota toxin (Ib), C. spiroforme toxin (CSTb), C. botulinum C2 toxin (C2II) and B. cereus/thuringiensis VIP1, respectively. CDTb shares significant sequence similarity (36% identity) with the protective antigen (PA) of B. anthracis toxin.25 This finding is of major importance, because our knowledge about the structure-function relationship of PA was more advanced25 and much was learned from this toxin about activation, membrane interaction, pore formation and translocation of the binding components of binary actin ADP-ribosylating toxins (see below).
CDTb is expressed with a signal sequence of 42 amino acids. The binding component is divided into 4 domains. So far the binding domain has been crystalized from C2II26and PA27 but not from CDTb, Ib or CSTb. Mainly deduced from the similar structure of PA,25 the functions of the domains of CDTb are as follows: The N-terminal 257 residues form the activation domain I, which is followed by domain II until residue 480. Domain II is involved in membrane insertion and pore formation. Domain III is responsible for oligomerization and the C-terminal domain IV (from amino acid 592 to 876) is involved in receptor binding. The latter domain is very similar between CDTb, Ib and CSTb but does not share significant sequence similarity with C2II, VIP2 or PA. Moreover, the binding components of CDT, iota toxin and C. spiroforme toxin can be exchanged among each other, whereas C2II is not able to deliver the enzyme component of these toxins into target cells. This finding classifies CDT, iota toxin and CST into the subfamily of iota-like binary toxins.19
As known for all the binding components, CDTb has to be activated by serine-type proteases.11 Activating cleavage of the full length binding component occurs most likely between Lys209 and Leu210 to release a 20 kDa peptide and a 75 kDa peptide. The large peptide is the activated binding component and forms most likely heptamers. Heptamer formation has been shown for C2II,28 PA (although octamer formation was also suggested29) and for iota toxin.30,31 Whether the activation occurs before or after receptor binding is not clear. Both monomeric and heptameric CDTb bind to its cell surface receptor LSR (lipolysis stimulated lipoprotein receptor).
LSR: lipolysis-stimulated lipoprotein receptor
Using a gene trapping approach and haploid cells (HAP1 cells), the host cell receptor for CDT was identified as the lipolysis-stimulated lipoprotein receptor (LSR).32 Knockout of the LSR gene in HAP1 cells resulted in specific resistance toward CDT, whereas the related C2 toxin still killed the cells. Expression of recombinant LSR in LSR−/− HAP1 cells reconstituted the toxicity. Moreover, it was observed that HeLa cells do not express LSR and are insensitive to CDT. Accordingly, expression of LSR in HeLa caused CDT-induced cytotoxicity. Moreover, a direct interaction of the binding component of CDT to LSR or its extracellular part could be shown.32 LSR was found to be a type I single-pass transmembrane protein of 581 amino acids and a mass of ~65 kDa (Fig. 2). It has an extracellular immunoglobulin–like domain and a long intracellular part. Three splice variants have been described33 but their specific functions were not clear. LSR was found highly expressed in liver and also in many other tissues including gut (both small intestine and colon), lung, kidney, adrenal glands, testes and ovaries.34 Importantly, LSR was also discovered to be the receptor for C. perfringens iota toxin32 and C. spiroforme toxin CST.35
What is the physiological function of LSR? Earlier studies have shown that LSR was involved in lipoprotein clearance.36,37 It was reported that fatty acids, which are released by lipolysis activate the binding of lipoproteins to LSR.38 The receptor was suggested to be involved in uptake and removal of apoB, E-containing triglyceride–rich lipoproteins and in clearance of low density lipoproteins (LDL) by an LDL-receptor independent pathway.
Recently, it was shown that LSR is involved in formation of tricellular tight junctions.39 This type of tight junction occurs where three epithelial cells meet each other. LSR is apparently essential for establishment of high transepithelial electrical resistance of epithelial monolayers. It recruits with its cytoplasmic tail the tricellular tight junction protein tricellulin, which is structurally similar to tight junction protein occludin.40 Accordingly, knock-down of LSR causes redistribution of tricellulin. LSR belongs to a family of immunoglobulin-like domain-containing receptor (ILDR), which has two other members ILDR2 and 3. Because ILDR2 and 3 are only ~30% identical with LSR, it is questionable that they are receptors for CDT.41
Recently, it has been suggested that in addition to LSR, CD44 may be involved in binding and/or endocytosis of iota-like toxins.42 CD44 is a multifunctional glycoprotein on the surface of mammalian cells, which is associated with lipid rafts and forms cell-surface clusters. Although its role in toxin binding is not clear, CD44 knockout mice are at least partially resistant to iota toxin toxicity, suggesting a function in up-take of binary iota-like toxins.
CDT uptake
Studies on the uptake of CDT into target cells are scarce (see model Fig. 3). However, high sequence similarity of CDT with iota toxin and usage of the same cell surface receptor (LSR) by both toxins allow drawing conclusions from uptake studies with iota toxins. Studies with iota toxin suggest that the monomeric activated binding component interacts with the cell surface receptor (LSR) followed by accumulation in lipid rafts, oligomerization and binding of the enzyme component.43-45 Thereafter, the toxin-receptor complex is internalized to reach endosomal compartments. The low pH of endosomes is probably a trigger for membrane insertion of the binding component and pore formation to allow translocation of the enzyme component into the cytosol. This is deduced from the finding that bafilomycin A, which blocks vesicle proton ATPase and thereby inhibits acidification of endosomes, prevents translocation of the enzyme component.30,46 However, pore formation by oligomerized binding component, resulting in potassium release,31 also occurs without acidification. In some cells [e.g., human epithelial carcinoma cells (A431)], the binding component of iota toxin causes necrosis without the enzyme component.47 From early endosomes, the binding component traffics to late endosomes and is probably then degraded in lysosomes.48

Figure 3. Up-take and mode of action of CDT. The proteolytically activated binding component of CDTb forms heptamers and binds to its cell surface receptor LSR. Alternatively, monomeric CDT binds to the receptor and, thereafter, polymerizes to form heptamers. Then, the enzymatic component CDTa interacts with CDTb. The LSR-CDT complex is endocytosed. At low pH of endosomes, the binding component inserts into the endosomal membrane and forms a pore. Through the pore, the enzymatic component is translocated into the cytosol. This process depends on cytosolic chaperon system, including heat shock proteins (HSP, cyclophillin A and FK506-binding protein 51). In the cytosol, CDTa ADP-ribosylates actin. ADP-ribosylated actin is not able to polymerize and is trapped in its monomeric form. Moreover, ADP-ribosylated actin acts like a capping protein to block polymerization at the barbed (plus) ends of F-actin. This causes enhanced depolymerization of the actin cytoskeleton. The depolymerization of cortical actin, which is located beneath the cell membrane, results in formation of long protrusions, which are microtubule based. The protrusions form a network on the surface of epithelial cells, which increases the interaction interface, and enhances adherence and colonization of clostridia.
The mechanism of pore formation by CDTb-like proteins is best understood from PA and C2II. PA forms heptamers (or octamers) as a prepore, which at low pH, change their structure and insert into membranes by forming a 14 (or 16)-stranded β-barrel structure with a pore lumen of about 1.5 nm.49,50 Similar pore formation is suggested for all binding components of the binary actin ADP-ribosylating toxins and have been frequently studied in artificial black lipid membranes with iota toxin and C2 toxin.51-55 How the enzyme component of CDT is translocated through the pore, is deduced from studies with iota toxin and C2 toxin. The enzyme component is probably unfolded into a molten globule structure for translocation as suggested for transport through the PA50,56 and C2 toxin pore.57
Interestingly translocation of the enzyme component of all binary actin ADP-ribosylating toxins, including CDT, depends on intracellular helper proteins. Using heat shock protein 90 (Hsp90)-inhibitors geldanamycin and radicicol, it was first shown that the up-take of C2I and Ia depends on the chaperon Hsp90.58,59 Later the involvement of the petidyl-prolyl cis/trans-isomerase cyclophilin A60and the FK506-binding protein 51 was reported.61 These data indicate that the translocation process of ADP-ribosylating toxins through pores formed by oligomeric toxin binding components is supported by a complex intracellular machinery. Understanding of these processes will certainly offer new targets for anti-toxin strategies.
Mechanism of Action of Binary Toxin
Actin, the substrate of CDT
After translocation into the cytosol, CDTa ADP-ribosylates actin. Actin is a 45 kDa cytoskeletal protein, which is abundant in all cells and tissues and highly conserved (e.g., ~90% sequence identity between human and yeast actin).62 Six mammalian actin isoforms are known, which differ maximally in 25 amino acid residues. Actin is involved in a large array of cellular functions. It participates in establishment of cell morphology, is crucial for cell adhesion, motility and cytokinesis. It plays crucial roles in phagocytosis, endocytosis, intracellular traffic and secretory processes. Moreover, actin plays important roles in cellular signaling and regulation of transcription. All these functions depend on tightly regulated polymerization and depolymerization of monomeric actin (G-actin) to form long double stranded helical filaments.63-65
CDT ADP-ribosylates actin
CDT ADP-ribosylates actin at arginine-177.23 The same residue is also modified by the other binary actin ADP-ribosylating toxins, including iota toxin,66 CST, VIP and C2 toxin.19,67,68 Photorhabdus luminescens toxins Photox,69 Aetx70 and Salmonella effector SpvB71 also ADP-ribosylate actin at the same site. Thus, all these toxins may cause the same pathophysiological consequences in target cells. Modification of arginine-177 by CDT and by other toxins inhibited polymerization of actin70 (Fig. 3). Moreover, toxin-ADP-ribosylated actin acted like a capping protein, which binds to the barbed (fast polymerizing) ends of actin filaments thereby inhibiting polymerization of non-modified actin72 and interfering with gelsolin (an 82-kD lipid-binding actin regulatory protein) functions.73 By contrast the pointed end of actin filaments is free. Here actin usually depolymerizes. Thus, ADP-ribosylation of actin in arginine-177 induces depolymerization of actin filaments, eventually resulting in complete destruction of the actin cytoskeleton.68,74
CDT and microtubule formation
ADP-ribosylation of actin by CDT affects microtubule structure and bacterial adherence. Treatment of colonic epithelial cells with CDT causes major changes in the organization of microtubules.75 This cytoskeleton system consists of α-/β-tubulin heterodimers to form long intracellular filaments, which exhibit a polarized structure with a fast growing plus end and a slowly growing minus end. Whereas the minus end is often stabilized at the microtubule organization center, the plus ends show dynamic instability with rapid polymerization and depolymerization cycles. When microtubules polymerize into the direction of the cell membrane, their growth is usually blocked when they reach the cortical actin network, which is located beneath the cell membrane. Here, capture proteins stabilize microtubules and may stop further growth.76,77
However, when epithelial cells are treated with actin-depolymerizing toxins, including CDT, growth of microtubules is not blocked at the cell membrane. At sites where the toxin-induced depolymerization of cortical actin occurs, long membrane protrusions are observed, which are filled with microtubules (Fig. 4).75 These protrusions are formed by growth of microtubule plus ends .The protrusions are 0.05–0.5 µm in diameter and have tentacle-like extensions of up to several hundred micrometers. Electron microscopic studies show that these tentacles and protrusions form a network of filaments on the surface of epithelial cells, which increase the adherence of bacteria. Importantly, while CDT-induced protrusions are best studied in cell culture, they also occur in intact tissue and in vivo studies have shown that CDT increases adherence of C. difficile in a mouse model of infection.75 All these data suggest that destruction of the actin cytoskeleton induces increased adherence and colonization of the toxin-producing bacteria (Fig. 4).
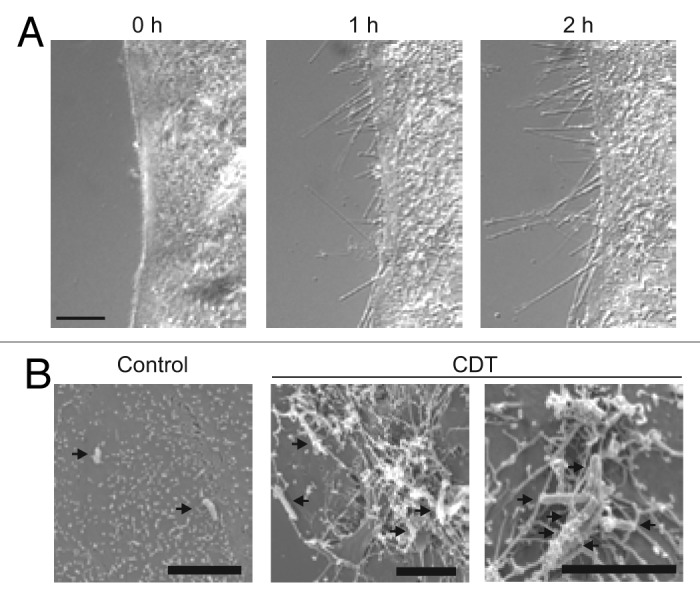
Figure 4. Microscopic pictures of CDT-induced protrusions. (A) The microscopic pictures show protrusion formation after treatment of human carcinoma Caco-2 cells after CDT treatment. (B) Electron microscopic studies show clostridia embedded in the meshwork of protrusions (pictures are modified from Schwan et al.75).
Evolving Epidemiology of Binary Toxin Positive C. difficile Strains
Toxinotyping and PCR ribotyping have been widely used to characterize C. difficile strains and the presence of binary toxin genes has been correlated with different toxinotypes and ribotypes. Toxinotyping categorized strains on the basis of genetic variations within the pathogenicity locus (PaLoc) encoding genes for the glucosylating toxins A and B and 3 accessory genes and was not obviously linked to the binary toxin genes which reside at a chromosomal site distant from the PaLoc.14 However, virtually all strains that contained complete cdtA and cdtB genes belonged to one of several variant toxinotypes (non-toxinotype 0)4,13,15 or “non-toxigenic” strains that did not have a PaLoc.78 The reason for this correlation is unknown, however a large proportion of toxinotype 0 strains contained truncated forms or pseudogenes (Fig. 1) of the binary toxin genes (79% of all tested toxinotype 0 strains).14,15 Not all variant toxinotypes have been shown to contain binary toxin genes, including the wide-spread toxinotype VIII which does not produce toxin A yet caused clinical syndromes similar to toxin A and B-producing strains (Table 1).13,15
Table 1. Correlation of Toxinotypes and PCR ribotypes among binary toxin-positive C. difficile strains (CDT+)*.
| Toxin production type | Toxinotypes | Ribotypes | Molecular background of PaLoc | Epidemiologic associations |
|---|---|---|---|---|
| A+B+CDT+ | minor types I, XXIV |
ND | A+B+: no major PaLoc deletions | |
| major types III IV V VI VII IX XIV XV XXII XXIII |
027, 034, 075, 080 023, 063 (058, 058) 078, 126, 045, 066 045, 063, 066 063 019 111 122 ND ND4,13,80,110 |
Toxinotype III/Ribotype 027 has been the major epidemic strain in North America since 20011,2 and has been epidemic in Europe.88 Toxinotype V/Ribotype 078 is the most common strain recovered from pigs/ calves93 and is an increasing cause of clinical CDI in the Netherlands3; and 3rd most prevalent in EU.91 Ribotype 023 is among 10 most prevalent ribotypes in EU.91 |
||
| A-B+CDT+ | X | 03613 | TcdA: rearrangement in PaLoc and large deletion probably causing changes in regulation and low or no transcription of truncated TcdA | |
| XVI, XVII, some V-like strains |
ND ND ND |
TcdA: mechanism unknown | ||
| XXX XXXI |
280, 281 237118 |
TcdA not present | ||
| A-B-CDT+ | XIa, XIb | 033 Yes13 |
A-B-: only small nonfunctional part of PaLoc present | Uncommon strains found mainly in asymptomatic patients78 |
| some strains without PaLoc | ND | A-B-: no PaLoc | Uncommon strains found mainly in asymptomatic patients78 |
CDT+: Presence of full length CDT locus, implying the potential for expression of binary toxin. Some A+B+ strains contain portions of the CDT locus, but are predicted as non-binary toxin producing strains (CDT-); ND, not done.
PCR ribotyping, a more generalizable typing method is based on amplification of the 16S-23S rRNA gene intergenic spacer region and therefore, is not obviously linked to binary toxin genes similar to toxinotyping. Certain ribotypes are consistently associated with the presence of binary toxin, including the epidemic PCR ribotype 027/toxinotype III strains (also known as BI/NAP1)13 (Table 1).
Prior to the emergence of the 027/BI/NAP1 strain in the early 2000s most surveys of C. difficile strains documented a prevalence of binary toxin genes of < 10%.79-84 Only 5.5% of strains in isolates from the Anaerobe Reference Unit at Cardiff were positive for binary toxin genes when reported in 2000.13 Likewise, a 5-y survey of consecutive clinical isolates in one hospital in Chicago between 1996 and 2001 found a binary toxin gene prevalence of 5.8%.85 The epidemiology of binary toxin positive C. difficile strains in the US changed dramatically over the next decade, largely as the result of the emergence of the epidemic 027/BI/NAP1 strain. In the US, the 027/BI/NAP1 strain was reported as the predominant strain in 8 hospital CDI outbreaks in 7 states between 2000 and 2003.1 In one affected hospital in Pittsburgh, 17 colectomies were performed for fulminant CDI in 2000 compared with an average of 2.7 per year over the previous decade.86 By 2009, this strain was endemic throughout the US, accounting for 61% of all clinical isolates from 25 acute healthcare facilities in Chicago.87 Shortly after the reports in the US a multi-hospital regional CDI outbreak occurred in Canada involving over 12 hospitals in the Montreal area between 2003 and 2004.2 Increased rates and severity of CDI were reported with an estimated 2,000 deaths directly attributable to CDI. The 027/BI/NAP1 strain accounted for 82% of the clinical isolates during this outbreak.2 This strain was subsequently documented in several European countries where outbreaks of severe CDI were also noted.88
The proportion of binary toxin positive strains has also increased in human CDI cases independent from the spread of type 027/BI/NAP1. In Italy strains isolated in different time intervals were compared and binary toxin positive strains represented 0% (before 1990), 24% (1991 to 1999), and 45% (2000 to 2001).89 The same trend was also observed in two large European studies in which the majority of European Union countries have participated. In the first study strains were collected during a 2 mo period in 2005 from 14 countries and 17.2% of all toxinogenic isolates were binary toxin positive (determined as presence of nontruncated binary toxin genes).90 In the second study performed in a 1 mo period in year 2008, strains were obtained from 34 countries and the proportion of binary positive strains among all toxinogenic strains was 23%.91 Only 5% of the isolates were ribotype 027.
Binary toxin-positive (CDT+) strains were also frequently recovered from animals. Interestingly, while CDT+ strains have represented less than 10% of all human isolates prior to the outbreak of 027/BI/NAP1, they were typically associated with animals and have represented from 20 to 100% of all animal isolates.92 One particular CDT+ group predominated among pigs and calves and has been characterized as PCR ribotype 078 and toxinotype V.92,93 Toxinotype V strains recovered from these animals raised for food production appeared to be clonally related to type V strains recovered from humans with CDI.3,94
Although some investigators have reported the recovery of C. difficile, including CDT+ strains, in meat destined for human consumption,95-97 foodborne transmission of C. difficile has not been confirmed.98 Goorhuis et al. reported an increase in the incidence of ribotype 078 strains among patients in the Netherlands from 3% to 13% between 2005 and 2008.3 Compared with CDI cases associated with 027 strains, 078 cases were younger and more frequently had community-associated disease but had similar rates of severe diarrhea and attributable mortality.3 Additionally, binary toxin positive strains other than ribotype 078 have been reported to often be associated with community onset CDI.82,99 Further characterization of CDT+ strains would be helpful to clarify the epidemiology of binary toxin in C. difficile.
Importance of Binary Toxin as a Possible Pathogenesis Factor in C. difficile
The pathogenic potential of binary toxin was somewhat masked by studies of CDI severity that focused on specific PCR ribotypes, PFGE types or REA group strains of C. difficile that possess binary toxin, but also have other genetic characteristics that might account for enhanced virulence. Among these potential virulence factors were possible increased sporulation compared with other strains, better adhesion properties, and deletions and mutations in the tcdC gene that could result in increased production of TcdA or TcdB toxins. For example, a recent study comparing the association of CDI severity with PCR ribotype 027 failed to find an association between severe infection and ribotype 027 as the cause of CDI using multivariate analysis, even though ribotype 027 had a higher proportion of severe disease than any other ribotype.100 Previous similar studies without as large CDI patient populations have also failed to show an association of the PCR ribotype 027 strain with severe CDI.101-103 In contrast, Petrella et al.104 reported a significantly lower treatment response rate and higher recurrence rate for 027/NAP1/BI strains than for other strains in two large clinical trials of vancomycin and fidaxomicin for CDI treatment. Miller et al. reported a Canada-wide survey in 2005 by the Canadian Nosocomial Infection Surveillance Program (CNISP) of 1008 patients with clinical data and infecting strain analysis in which 31% were infected with the 027/BI/NAP1 strain.105 This study confirmed a strong age association with infection due to this epidemic strain: 12.5% of NAP1-associated infections compared with 5.9% of non-NAP1-associated infections resulted in a severe outcome (p < 0.001). Patients 60–90 y of age infected with the NAP1 strain were approximately twice as likely to die or have a severe outcome compared with same aged patients with other strains. An important deficiency of strain-specific studies as a measure of the effect of binary toxin was that they may unknowingly harbor other binary toxin positive strains in the control groups, since the number of ribotypes known to carry binary toxin is quite extensive (Table 1). If this were to occur the severity of disease in the control group could be increased if large numbers of binary toxin-positive strains were included within the comparator group.
One study has attempted to differentiate the possible clinical effects of binary toxin from other genetic properties such as tcdC mutations in clinical isolates obtained over a 2-y period at one NHS Foundation Trust in London.106 The prevalence of tcdC mutations and binary toxin genes was analyzed in 207 C. difficile isolates and compared with measured risk factors of patient age and laboratory findings as well as patient outcomes (disease severity, ICU admission, mortality, recurrence and length of stay). The prevalence of tcdC truncating mutations was 15% whereas binary toxin genes were present in nearly twice as many isolates, 28%. Patients infected with isolates containing tcdC truncating mutations had significantly elevated C-reactive protein and peripheral white blood cell (WBC) counts compared with patients not infected with these strains, but there was no difference in patient outcome. In contrast, patients infected with C. difficile strains containing binary toxin not only had significantly higher peripheral WBC, but also had a significantly higher 30-d all-cause mortality (31% vs 14%, P = 0.02). In this study only 8% of isolates were ribotype 027 whereas 28% of isolates contained binary toxin genes.
A similar analysis was done on 212 C. difficile isolates containing genes for toxins A and B compared with 265 isolates containing genes for toxins A, B, and binary toxin, including 193 isolates of PCR ribotype 027 and 72 isolates that were non-027.107 Isolates were forwarded to the National Reference Laboratory at Statens Serum Institut, Copenhagen by departments of clinical microbiology if they were resistant to moxifloxacin, a severe clinical course was observed, or if a CDI outbreak was suspected. All isolates were genotyped to detect genes for toxins A, B, and binary toxin and PCR ribotyped. Of the isolates possessing toxin A, B, and binary toxin genes that were not ribotype 027, 24 (33%) were PCR ribotype 078, 26 (36%) were PCR ribotype 066, and 22 (31%) were PCR ribotype 023 and 9 other PCR ribotypes. The case fatality rate at 30 d after diagnosis was 54/193 (28%) for PCR ribotype 027 strains, 20/72 (27.8%) for binary toxin-positive non-027 isolates, and 36/212 (17%) for patients infected with toxin A and B positive isolates. Among binary toxin-positive non-027 isolates, the 30-d mortality was 29.2% for ribotype 078, 30.8% for ribotype 066, and 22.7% for ribotype 023 and others. Based on the similar case-fatality rates for strains possessing toxins A, B, and binary toxin, these strains were compared with the case-fatality rate for strains possessing only toxins A and B. In univariate analysis, the relative risk of death within 30 d was 1.8 (95% CI 1.2–2.7) for binary toxin-positive strains and by multivariate analysis adjusting for age, gender, and geographic region the relative risk was 1.6 (95% CI 1.0–2.4).
In an earlier study with fewer patients Barbut et al.99 identified 26 patients in 1999 and 2000 who had CDI caused by binary toxin-positive isolates and did a case control study of 42 patients with CDI caused by isolates that possessed only toxin A and B genes (not binary toxin genes) who were hospitalized at the same time on the same ward. Patients infected with binary toxin-positive strains were more likely to be community-associated (P = 0.017) and were more likely to have CDI as the cause of hospitalization (P = 0.003) by univariate analysis. In a later study conducted at one large teaching hospital in Paris from 2000 to 2004 Barbut et al.108 analyzed 131 C. difficile strains causing CDI. Binary toxin-positive isolates were found in 11% and were responsible for more severe diarrhea (P = 0.01) and higher case-fatality rate (P = 0.03) than isolates that did not contain binary toxin. Binary toxin-positive isolates were found in toxinotypes I, III, IV, V, VI, IX, XII, XIV, and XXIV, and only one of the toxinotype III isolates was related to the epidemic ribotype 027 strain. These observations suggest a possible link of the presence of binary toxin in a wide variety of C. difficile toxin variant strains to increased CDI mortality.
An additional study analyzed potential virulence factors, including tcdC deletions and binary toxin genes, in C. difficile isolates obtained from 69 consecutive patients who were followed for CDI recurrences.109 In this study, 41% of patients had at least one CDI recurrence and 38% of the isolates from the initial episode were PCR ribotype 027. The presence of binary toxin gene was significantly associated with recurrence (P = 0.02) and need for hospital admission with recurrence (P = 0.02). tcdC deletion and ribotype 027 were not associated with recurrence.
Attributable mortality and severe diarrhea were similar in PCR ribotype 078 (n = 54) and 027 (n = 124) in one study from the Netherlands and both were greater than in 501 non-027/078 cases.3 More recently, PCR ribotype 078 strains of C. difficile that produce binary toxin were found to be associated with the highest 14-d mortality (16/63, 25%) in a very large patient study from the United Kingdom, exceeding the next highest 14-d mortality of 20% (111/560) for ribotype 027 strains that also produce binary toxin.110
The effect of binary toxin in animal studies of CDI has been difficult to assess. Geric et al.78 were able to identify a prevalence of at least 2% binary toxin-positive genes in isolates that did not possess toxin A and B genes. These isolates provided an opportunity to test the virulence in animal models of strains possessing only binary toxin genes. Supernatants from four A-/B-/CDT+ strains were found to cause marked fluid accumulation in the rabbit ileal loop model.111 However, when hamsters were infected with these strains following clindamycin, they colonized the gastrointestinal tract but did not cause diarrhea or death. It was speculated that binary toxin by itself may not be sufficient to cause disease. More recently, Kuenhe et al.112 published data on the use of ClosTron technology to inactivate tcdA, tcdB and binary toxin (cdtA) genes. Isolates lacking toxin A or toxin B remained fully virulent in the hamster model, but when both toxin A and toxin B were inactivated and only the binary toxin genes remained active, 3 of 8 animals died. The symptoms in these animals were not typical of hamster CDI. The animals had signs of wet tail but had no cecal lesions, however, they did demonstrate hemorrhage and inflammation in their small intestines which was not seen in any other animals. In addition, when toxin B was inactivated, the presence of binary toxin with toxin A caused significantly more rapid hamster death (p < 0.05) than when only toxin A was present. These observations suggest that there may be an important role for binary toxin in C. difficile pathogenesis.
Future Implications for Binary Toxin Research
It is clear that the pathogenic potential of C. difficile organisms containing binary toxin has not been fully explored. It is also apparent that there is not a direct correlation between presence of the cdtA and cdtB genes and production of CDTa and CDTb polypeptides and binary toxin CDT.113 Detection of the binary toxin genes may not be sufficient to indicate the presence of the toxin in stool as measured by CDTb in stool. Only 19 of 36 (53%) of stools containing CDT+ isolates had CDTb detected in the stool specimen.113 Levels of CDTb detected in broth culture also correlated poorly with levels in stool specimens with generally higher levels in stool than in broth supernatants.113 Given these limitations of genetic detection for binary toxin CDT, there is a clinical need for detection of binary toxin in the stool of patients with CDI. At present there are no binary toxin clinical assays available, however, a prototype novel enzyme immunoassay for CDTb has been developed for research purposes by TechLab, Inc.113
In an oral presentation at the 4th International Clostridium difficile Symposium Heinrichs et al. (Heinrichs JH, Wang S., Miezeiewski M, Secore S, Xie A, Zorman J, et al. Design, production and pre-clinical evaluation of a novel toxin-based vaccine for the prevention of Clostridium difficile disease. 4th International Clostridium difficile Symposium September 2012; Bled, Slovenia, Abstract 01, www.icds.si/abstracts.php) showed preliminary pre-clinical data on the benefit of including binary toxin antigens in a C. difficile candidate vaccine. Their group first developed a recombinant vaccine targeting toxin A and toxin B that was highly effective in preventing disease in the hamster model when challenged with C. difficile strain VPI10463 that produces very high levels of toxin A and toxin B but lacks binary toxin. However, when hamsters were challenged with C. difficile strain BI17, a NAP1/BI/027 strain that produces toxins A, B, and binary toxin, the hamsters were not protected by this vaccine. They then developed CDTa and CDTb antigens recombinantly and tested them independently and together in combination with the TcdA and TcdB antigens in trivalent and tetravalent antigen combinations. The combination of all 4 antigens provided significantly superior protection of hamsters compared with TcdA and TcdB divalent vaccine (p < 0.0001) and was superior to TcdA, TcdB and either CDTa or CDTb antigens in trivalent combinations. This protective effect was shown for an additional NAP1/BI/027 strain, strain BI6, and for challenge with strain 630 (binary toxin negative strain) as well. The authors also showed that neutralizing antibody to binary toxin was associated with protection in the hamsters challenged with strain BI17. In contrast to the results of Kuehne et al.,112 they found a lack of fatal illness in hamsters challenged with a C. difficile strain that produced only binary toxin and not toxin A or B, although some of the animals developed soft stools and wet-tail consistent with the hamster observations of Kuehne et al. using their model.112
Although the results of the above study are still preliminary, if confirmed, they suggest significant pathogenic capability of binary toxin in combination with toxins A and B in the hamster model as also shown by Kuehne et al.,112 which could account for the increased mortality and disease severity associated with binary toxin-containing C. difficile strains infecting humans. Whether binary toxin targeting for vaccine development is required has not been determined, and existing clinical data for monoclonal antibodies directed against only toxin A and toxin B indicated that prevention of CDI recurrence following CDI treatment was achieved for strains that make binary toxin.114 Lowy et al. showed that the recurrence rate among patients with the epidemic BI/NAP1/027 strain was 8% for the antibody group and 32% for the placebo group (P = 0.06) suggesting that at least for recurrent CDI, antibody directed at binary toxin was apparently not necessary.114 However, protection against primary challenge with C. difficile by monoclonal antibodies or vaccines is more difficult than prevention of CDI recurrence in the hamster model115 and prevention of primary CDI may be an important distinction requiring the addition of binary toxin antigen to the vaccine mix.
One additional area of experimental interest is the use of binary toxin as an intracellular toxin delivery system to dissect aspects of cell function.116 The AB binary toxins are particularly well suited for this purpose as 1) their toxins are particularly potent due to the enzymatic A-domain, 2) they possess highly sophisticated mechanisms to translocate their A-domain across cell membranes, and 3) the toxins are highly specific with regard to their host cell substrates.117 Most work has been done with the C. botulinum C2 binary toxin. The receptor for C2 toxin seems to be universally present on vertebrate cells making it a useful system for study of multiple cell lines. The ability of the enzymatic (ADP-ribosylating) portion of C2 toxin to depolymerize F-actin has been used to study the function of the actin skeleton in cellular processes such as exocytosis, cell migration, leukocyte activation and endothelium permeability.117 Construction of chimeric fusion proteins by mixing the A and B domains of various binary toxins to achieve more selective binding of toxin to specific cell types such as tumor cells can be achieved. In addition, the A domain may be substituted with proteins or nucleic acids that can be introduced into the cytosol of specific cell types to correct deficiencies. Specific antibody domains can also be fused to the A-domain to create targeted immunotoxins.116 There are significant challenges as the pore size of the B-domain requires unfolding of proteins in order to translocate followed by refolding of the protein which requires a host cell chaperone. Increased knowledge of these mechanisms and their structure-function relationships should allow binary toxins to be useful carriers of nucleic acids and foreign proteins into mammalian cells.
Conclusions
Binary toxin CDT may be an important virulence factor of C. difficile for which the significance remains to be determined. Several clinical studies suggest an association between the presence of binary toxin in infecting C. difficile strains and increased mortality of the patients. Further studies of the effects of binary toxin-producing strains are needed including measures of binary toxin in stool, analyses of strains carrying binary toxin across multiple PCR ribotypes, the relationship of TcdA and TcdB variants to the presence of binary toxin in C. difficile strains, and further study of the epidemiology, mortality and risk factors for CDI caused by binary toxin-positive C. difficile strains.
Acknowledgments
Supported by research grants from the US Department of Veterans Affairs Research Service (Johnson S and Gerding DN). The authors thank Debbie C Walton, Jill Williams, and Karyn J Davis for editorial assistance.
Disclosure of Potential Conflicts of Interest
Gerding DN holds patents for prevention and treatment of CDI licensed to ViroPharma and is a consultant for ViroPharma, Merck, Roche, Novartis, Optimer, Cubist, Cangene, Sanofi Pasteur, Summit PLC and Actelion. Johnson S has served as a consultant for Optimer, Pfizer, and Bio-K+. Rupnik M has served as consultant for Astellas, Cubist and Summit PLC. Aktories K reports no potential conflicts.
References
- 1.McDonald LC, Killgore GE, Thompson A, Owens RC, Jr., Kazakova SV, Sambol SP, Johnson S, Gerding DN. An epidemic, toxin gene-variant strain of Clostridium difficile. N Engl J Med. 2005;353:2433–41. doi: 10.1056/NEJMoa051590. [DOI] [PubMed] [Google Scholar]
- 2.Loo VG, Poirier L, Miller MA, Oughton M, Libman MD, Michaud S, Bourgault AM, Nguyen T, Frenette C, Kelly M, et al. A predominantly clonal multi-institutional outbreak of Clostridium difficile-associated diarrhea with high morbidity and mortality. N Engl J Med. 2005;353:2442–9. doi: 10.1056/NEJMoa051639. [DOI] [PubMed] [Google Scholar]
- 3.Goorhuis A, Bakker D, Corver J, Debast SB, Harmanus C, Notermans DW, Bergwerff AA, Dekker FW, Kuijper EJ. Emergence of Clostridium difficile infection due to a new hypervirulent strain, polymerase chain reaction ribotype 078. Clin Infect Dis. 2008;47:1162–70. doi: 10.1086/592257. [DOI] [PubMed] [Google Scholar]
- 4.Rupnik M. Heterogeneity of large clostridial toxins: importance of Clostridium difficile toxinotypes. FEMS Microbiol Rev. 2008;32:541–55. doi: 10.1111/j.1574-6976.2008.00110.x. [DOI] [PubMed] [Google Scholar]
- 5.Debast SB, van Leengoed LA, Goorhuis A, Harmanus C, Kuijper EJ, Bergwerff AA. Clostridium difficile PCR ribotype 078 toxinotype V found in diarrhoeal pigs identical to isolates from affected humans. Environ Microbiol. 2009;11:505–11. doi: 10.1111/j.1462-2920.2008.01790.x. [DOI] [PubMed] [Google Scholar]
- 6.Barbut F, Rupnik M. Editorial commentary: 027, 078, and others: going beyond the numbers (and away from the hypervirulence) Clin Infect Dis. 2012;55:1669–72. doi: 10.1093/cid/cis790. [DOI] [PubMed] [Google Scholar]
- 7.Aktories K, Bärmann M, Ohishi I, Tsuyama S, Jakobs KH, Habermann E. Botulinum C2 toxin ADP-ribosylates actin. Nature. 1986;322:390–2. doi: 10.1038/322390a0. [DOI] [PubMed] [Google Scholar]
- 8.Aktories K, Wille M, Just I. Clostridial actin-ADP-ribosylating toxins. Curr Top Microbiol Immunol. 1992;175:97–113. doi: 10.1007/978-3-642-76966-5_5. [DOI] [PubMed] [Google Scholar]
- 9.Popoff MR, Boquet P. Clostridium spiroforme toxin is a binary toxin which ADP-ribosylates cellular actin. Biochem Biophys Res Commun. 1988;152:1361–8. doi: 10.1016/S0006-291X(88)80435-2. [DOI] [PubMed] [Google Scholar]
- 10.Popoff MR, Rubin EJ, Gill DM, Boquet P. Actin-specific ADP-ribosyltransferase produced by a Clostridium difficile strain. Infect Immun. 1988;56:2299–306. doi: 10.1128/iai.56.9.2299-2306.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Perelle S, Gibert M, Bourlioux P, Corthier G, Popoff MR. Production of a complete binary toxin (actin-specific ADP-ribosyltransferase) by Clostridium difficile CD196. Infect Immun. 1997;65:1402–7. doi: 10.1128/iai.65.4.1402-1407.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Braun M, Herholz C, Straub R, Choisat B, Frey J, Nicolet J, Kuhnert P. Detection of the ADP-ribosyltransferase toxin gene (cdtA) and its activity in Clostridium difficile isolates from Equidae. FEMS Microbiol Lett. 2000;184:29–33. doi: 10.1111/j.1574-6968.2000.tb08985.x. [DOI] [PubMed] [Google Scholar]
- 13.Stubbs S, Rupnik M, Gibert M, Brazier J, Duerden B, Popoff M. Production of actin-specific ADP-ribosyltransferase (binary toxin) by strains of Clostridium difficile. FEMS Microbiol Lett. 2000;186:307–12. doi: 10.1111/j.1574-6968.2000.tb09122.x. [DOI] [PubMed] [Google Scholar]
- 14.Carter GP, Lyras D, Allen DL, Mackin KE, Howarth PM, O’Connor JR, Rood JI. Binary toxin production in Clostridium difficile is regulated by CdtR, a LytTR family response regulator. J Bacteriol. 2007;189:7290–301. doi: 10.1128/JB.00731-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Stare BG, Delmée M, Rupnik M. Variant forms of the binary toxin CDT locus and tcdC gene in Clostridium difficile strains. J Med Microbiol. 2007;56:329–35. doi: 10.1099/jmm.0.46931-0. [DOI] [PubMed] [Google Scholar]
- 16.Pituch H, Kreft D, Obuch-Woszczatynski P, Wultańska D, Meisel-Mikołajczyk F, Łuczak M, van Belkum A. Clonal spread of a Clostridium difficile strain with a complete set of toxin A, toxin B, and binary toxin genes among Polish patients with Clostridium difficile-associated diarrhea. J Clin Microbiol. 2005;43:472–5. doi: 10.1128/JCM.43.1.472-475.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bouvet PJ, Popoff MR. Genetic relatedness of Clostridium difficile isolates from various origins determined by triple-locus sequence analysis based on toxin regulatory genes tcdC, tcdR, and cdtR. J Clin Microbiol. 2008;46:3703–13. doi: 10.1128/JCM.00866-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Metcalf DS, Weese JS. Binary toxin locus analysis in Clostridium difficile. J Med Microbiol. 2011;60:1137–45. doi: 10.1099/jmm.0.028498-0. [DOI] [PubMed] [Google Scholar]
- 19.Barth H, Aktories K, Popoff MR, Stiles BG. Binary bacterial toxins: biochemistry, biology, and applications of common Clostridium and Bacillus proteins. Microbiol Mol Biol Rev. 2004;68:373–402. doi: 10.1128/MMBR.68.3.373-402.2004. [table.] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Chang SY, Song KP. ADP-ribosylating binary toxin genes of Clostridium difficile strain CCUG 20309. DNA Seq. 2001;12:115–20. doi: 10.3109/10425170109047564. [DOI] [PubMed] [Google Scholar]
- 21.Sundriyal A, Roberts AK, Shone CC, Acharya KR. Structural basis for substrate recognition in the enzymatic component of ADP-ribosyltransferase toxin CDTa from Clostridium difficile. J Biol Chem. 2009;284:28713–9. doi: 10.1074/jbc.M109.043018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Han S, Craig JA, Putnam CD, Carozzi NB, Tainer JA. Evolution and mechanism from structures of an ADP-ribosylating toxin and NAD complex. Nat Struct Biol. 1999;6:932–6. doi: 10.1038/13300. [DOI] [PubMed] [Google Scholar]
- 23.Gülke I, Pfeifer G, Liese J, Fritz M, Hofmann F, Aktories K, Barth H. Characterization of the enzymatic component of the ADP-ribosyltransferase toxin CDTa from Clostridium difficile. Infect Immun. 2001;69:6004–11. doi: 10.1128/IAI.69.10.6004-6011.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Tsuge H, Nagahama M, Oda M, Iwamoto S, Utsunomiya H, Marquez VE, Katunuma N, Nishizawa M, Sakurai J. Structural basis of actin recognition and arginine ADP-ribosylation by Clostridium perfringens iota-toxin. Proc Natl Acad Sci U S A. 2008;105:7399–404. doi: 10.1073/pnas.0801215105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Young JA, Collier RJ. Anthrax toxin: receptor binding, internalization, pore formation, and translocation. Annu Rev Biochem. 2007;76:243–65. doi: 10.1146/annurev.biochem.75.103004.142728. [DOI] [PubMed] [Google Scholar]
- 26.Schleberger C, Hochmann H, Barth H, Aktories K, Schulz GE. Structure and action of the binary C2 toxin from Clostridium botulinum. J Mol Biol. 2006;364:705–15. doi: 10.1016/j.jmb.2006.09.002. [DOI] [PubMed] [Google Scholar]
- 27.Petosa C, Collier RJ, Klimpel KR, Leppla SH, Liddington RC. Crystal structure of the anthrax toxin protective antigen. Nature. 1997;385:833–8. doi: 10.1038/385833a0. [DOI] [PubMed] [Google Scholar]
- 28.Barth H, Blocker D, Behlke J, Bergsma-Schutter W, Brisson A, Benz R, Aktories K. Cellular uptake of Clostridium botulinum C2 toxin requires oligomerization and acidification. J Biol Chem. 2000;275:18704–11. doi: 10.1074/jbc.M000596200. [DOI] [PubMed] [Google Scholar]
- 29.Kintzer AF, Thoren KL, Sterling HJ, Dong KC, Feld GK, Tang II, Zhang TT, Williams ER, Berger JM, Krantz BA. The protective antigen component of anthrax toxin forms functional octameric complexes. J Mol Biol. 2009;392:614–29. doi: 10.1016/j.jmb.2009.07.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Blöcker D, Behlke J, Aktories K, Barth H. Cellular uptake of the Clostridium perfringens binary iota-toxin. Infect Immun. 2001;69:2980–7. doi: 10.1128/IAI.69.5.2980-2987.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Nagahama M, Nagayasu K, Kobayashi K, Sakurai J. Binding component of Clostridium perfringens iota-toxin induces endocytosis in Vero cells. Infect Immun. 2002;70:1909–14. doi: 10.1128/IAI.70.4.1909-1914.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Papatheodorou P, Carette JE, Bell GW, Schwan C, Guttenberg G, Brummelkamp TR, Aktories K. Lipolysis-stimulated lipoprotein receptor (LSR) is the host receptor for the binary toxin Clostridium difficile transferase (CDT) Proc Natl Acad Sci U S A. 2011;108:16422–7. doi: 10.1073/pnas.1109772108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yen FT, Masson M, Clossais-Besnard N, André P, Grosset JM, Bougueleret L, Dumas JB, Guerassimenko O, Bihain BE. Molecular cloning of a lipolysis-stimulated remnant receptor expressed in the liver. J Biol Chem. 1999;274:13390–8. doi: 10.1074/jbc.274.19.13390. [DOI] [PubMed] [Google Scholar]
- 34.Mesli S, Javorschi S, Bérard AM, Landry M, Priddle H, Kivlichan D, Smith AJ, Yen FT, Bihain BE, Darmon M. Distribution of the lipolysis stimulated receptor in adult and embryonic murine tissues and lethality of LSR-/- embryos at 12.5 to 14.5 days of gestation. Eur J Biochem. 2004;271:3103–14. doi: 10.1111/j.1432-1033.2004.04223.x. [DOI] [PubMed] [Google Scholar]
- 35.Papatheodorou P, Wilczek C, Nölke T, Guttenberg G, Hornuss D, Schwan C, Aktories K. Identification of the cellular receptor of Clostridium spiroforme toxin. Infect Immun. 2012;80:1418–23. doi: 10.1128/IAI.06378-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yen FT, Mann CJ, Guermani LM, Hannouche NF, Hubert N, Hornick CA, Bordeau VN, Agnani G, Bihain BE. Identification of a lipolysis-stimulated receptor that is distinct from the LDL receptor and the LDL receptor-related protein. Biochemistry. 1994;33:1172–80. doi: 10.1021/bi00171a017. [DOI] [PubMed] [Google Scholar]
- 37.Yen FT, Roitel O, Bonnard L, Notet V, Pratte D, Stenger C, Magueur E, Bihain BE. Lipolysis stimulated lipoprotein receptor: a novel molecular link between hyperlipidemia, weight gain, and atherosclerosis in mice. J Biol Chem. 2008;283:25650–9. doi: 10.1074/jbc.M801027200. [DOI] [PubMed] [Google Scholar]
- 38.Mann CJ, Khallou J, Chevreuil O, Troussard AA, Guermani LM, Launay K, Delplanque B, Yen FT, Bihain BE. Mechanism of activation and functional significance of the lipolysis-stimulated receptor. Evidence for a role as chylomicron remnant receptor. Biochemistry. 1995;34:10421–31. doi: 10.1021/bi00033a014. [DOI] [PubMed] [Google Scholar]
- 39.Masuda S, Oda Y, Sasaki H, Ikenouchi J, Higashi T, Akashi M, Nishi E, Furuse M. LSR defines cell corners for tricellular tight junction formation in epithelial cells. J Cell Sci. 2011;124:548–55. doi: 10.1242/jcs.072058. [DOI] [PubMed] [Google Scholar]
- 40.Ikenouchi J, Furuse M, Furuse K, Sasaki H, Tsukita S, Tsukita S. Tricellulin constitutes a novel barrier at tricellular contacts of epithelial cells. J Cell Biol. 2005;171:939–45. doi: 10.1083/jcb.200510043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Higashi T, Tokuda S, Kitajiri S, Masuda S, Nakamura H, Oda Y, Furuse M. Analysis of the ‘angulin’ proteins LSR, ILDR1 and ILDR2--tricellulin recruitment, epithelial barrier function and implication in deafness pathogenesis. J Cell Sci. 2013;126:966–77. doi: 10.1242/jcs.116442. [DOI] [PubMed] [Google Scholar]
- 42.Wigelsworth DJ, Ruthel G, Schnell L, Herrlich P, Blonder J, Veenstra TD, Carman RJ, Wilkins TD, Van Nhieu GT, Pauillac S, et al. CD44 Promotes intoxication by the clostridial iota-family toxins. PLoS One. 2012;7:e51356. doi: 10.1371/journal.pone.0051356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nagahama M, Sakaguchi Y, Kobayashi K, Ochi S, Sakurai J. Characterization of the enzymatic component of Clostridium perfringens iota-toxin. J Bacteriol. 2000;182:2096–103. doi: 10.1128/JB.182.8.2096-2103.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nagahama M, Yamaguchi A, Hagiyama T, Ohkubo N, Kobayashi K, Sakurai J. Binding and internalization of Clostridium perfringens iota-toxin in lipid rafts. Infect Immun. 2004;72:3267–75. doi: 10.1128/IAI.72.6.3267-3275.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hale ML, Marvaud JC, Popoff MR, Stiles BG. Detergent-resistant membrane microdomains facilitate Ib oligomer formation and biological activity of Clostridium perfringens iota-toxin. Infect Immun. 2004;72:2186–93. doi: 10.1128/IAI.72.4.2186-2193.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Marvaud JC, Smith T, Hale ML, Popoff MR, Smith LA, Stiles BG. Clostridium perfringens iota-toxin: mapping of receptor binding and Ia docking domains on Ib. Infect Immun. 2001;69:2435–41. doi: 10.1128/IAI.69.4.2435-2441.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nagahama M, Umezaki M, Oda M, Kobayashi K, Tone S, Suda T, Ishidoh K, Sakurai J. Clostridium perfringens iota-toxin b induces rapid cell necrosis. Infect Immun. 2011;79:4353–60. doi: 10.1128/IAI.05677-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nagahama M, Umezaki M, Tashiro R, Oda M, Kobayashi K, Shibutani M, Takagishi T, Ishidoh K, Fukuda M, Sakurai J. Intracellular trafficking of Clostridium perfringens iota-toxin b. Infect Immun. 2012;80:3410–6. doi: 10.1128/IAI.00483-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Krantz BA, Trivedi AD, Cunningham K, Christensen KA, Collier RJ. Acid-induced unfolding of the amino-terminal domains of the lethal and edema factors of anthrax toxin. J Mol Biol. 2004;344:739–56. doi: 10.1016/j.jmb.2004.09.067. [DOI] [PubMed] [Google Scholar]
- 50.Krantz BA, Finkelstein A, Collier RJ. Protein translocation through the anthrax toxin transmembrane pore is driven by a proton gradient. J Mol Biol. 2006;355:968–79. doi: 10.1016/j.jmb.2005.11.030. [DOI] [PubMed] [Google Scholar]
- 51.Schmid A, Benz R, Just I, Aktories K. Interaction of Clostridium botulinum C2 toxin with lipid bilayer membranes. Formation of cation-selective channels and inhibition of channel function by chloroquine. J Biol Chem. 1994;269:16706–11. [PubMed] [Google Scholar]
- 52.Bachmeyer C, Benz R, Barth H, Aktories K, Gilbert M, Popoff MR. Interaction of Clostridium botulinum C2 toxin with lipid bilayer membranes and Vero cells: inhibition of channel function by chloroquine and related compounds in vitro and intoxification in vivo. FASEB J. 2001;15:1658–60. doi: 10.1096/fj.00-0671fje. [DOI] [PubMed] [Google Scholar]
- 53.Blöcker D, Bachmeyer C, Benz R, Aktories K, Barth H. Channel formation by the binding component of Clostridium botulinum C2 toxin: glutamate 307 of C2II affects channel properties in vitro and pH-dependent C2I translocation in vivo. Biochemistry. 2003;42:5368–77. doi: 10.1021/bi034199e. [DOI] [PubMed] [Google Scholar]
- 54.Lang AE, Neumeyer T, Sun J, Collier RJ, Benz R, Aktories K. Amino acid residues involved in membrane insertion and pore formation of Clostridium botulinum C2 toxin. Biochemistry. 2008;47:8406–13. doi: 10.1021/bi800615g. [DOI] [PubMed] [Google Scholar]
- 55.Knapp O, Benz R, Gibert M, Marvaud JC, Popoff MR. Interaction of Clostridium perfringens iota-toxin with lipid bilayer membranes. Demonstration of channel formation by the activated binding component Ib and channel block by the enzyme component Ia. J Biol Chem. 2002;277:6143–52. doi: 10.1074/jbc.M103939200. [DOI] [PubMed] [Google Scholar]
- 56.Krantz BA, Melnyk RA, Zhang S, Juris SJ, Lacy DB, Wu Z, Finkelstein A, Collier RJ. A phenylalanine clamp catalyzes protein translocation through the anthrax toxin pore. Science. 2005;309:777–81. doi: 10.1126/science.1113380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Haug G, Wilde C, Leemhuis J, Meyer DK, Aktories K, Barth H. Cellular uptake of Clostridium botulinum C2 toxin: membrane translocation of a fusion toxin requires unfolding of its dihydrofolate reductase domain. Biochemistry. 2003;42:15284–91. doi: 10.1021/bi0354278. [DOI] [PubMed] [Google Scholar]
- 58.Haug G, Aktories K, Barth H. The host cell chaperone Hsp90 is necessary for cytotoxic action of the binary iota-like toxins. Infect Immun. 2004;72:3066–8. doi: 10.1128/IAI.72.5.3066-3068.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Haug G, Leemhuis J, Tiemann D, Meyer DK, Aktories K, Barth H. The host cell chaperone Hsp90 is essential for translocation of the binary Clostridium botulinum C2 toxin into the cytosol. J Biol Chem. 2003;278:32266–74. doi: 10.1074/jbc.M303980200. [DOI] [PubMed] [Google Scholar]
- 60.Kaiser E, Kroll C, Ernst K, Schwan C, Popoff M, Fischer G, Buchner J, Aktories K, Barth H. Membrane translocation of binary actin-ADP-ribosylating toxins from Clostridium difficile and Clostridium perfringens is facilitated by cyclophilin A and Hsp90. Infect Immun. 2011;79:3913–21. doi: 10.1128/IAI.05372-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Kaiser E, Böhm N, Ernst K, Langer S, Schwan C, Aktories K, Popoff M, Fischer G, Barth H. FK506-binding protein 51 interacts with Clostridium botulinum C2 toxin and FK506 inhibits membrane translocation of the toxin in mammalian cells. Cell Microbiol. 2012;14:1193–205. doi: 10.1111/j.1462-5822.2012.01788.x. [DOI] [PubMed] [Google Scholar]
- 62.Pollard TD, Cooper JA. Actin, a central player in cell shape and movement. Science. 2009;326:1208–12. doi: 10.1126/science.1175862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Dominguez R, Holmes KC. Actin structure and function. Annu Rev Biophys. 2011;40:169–86. doi: 10.1146/annurev-biophys-042910-155359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Lanzetti L. Actin in membrane trafficking. Curr Opin Cell Biol. 2007;19:453–8. doi: 10.1016/j.ceb.2007.04.017. [DOI] [PubMed] [Google Scholar]
- 65.Disanza A, Steffen A, Hertzog M, Frittoli E, Rottner K, Scita G. Actin polymerization machinery: the finish line of signaling networks, the starting point of cellular movement. Cell Mol Life Sci. 2005;62:955–70. doi: 10.1007/s00018-004-4472-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Vandekerckhove J, Schering B, Bärmann M, Aktories K. Clostridium perfringens iota toxin ADP-ribosylates skeletal muscle actin in Arg-177. FEBS Lett. 1987;225:48–52. doi: 10.1016/0014-5793(87)81129-8. [DOI] [PubMed] [Google Scholar]
- 67.Vandekerckhove J, Schering B, Bärmann M, Aktories K. Botulinum C2 toxin ADP-ribosylates cytoplasmic beta/gamma-actin in arginine 177. J Biol Chem. 1988;263:696–700. [PubMed] [Google Scholar]
- 68.Aktories K, Lang AE, Schwan C, Mannherz HG. Actin as target for modification by bacterial protein toxins. FEBS J. 2011;278:4526–43. doi: 10.1111/j.1742-4658.2011.08113.x. [DOI] [PubMed] [Google Scholar]
- 69.Visschedyk DD, Perieteanu AA, Turgeon ZJ, Fieldhouse RJ, Dawson JF, Merrill AR. Photox, a novel actin-targeting mono-ADP-ribosyltransferase from Photorhabdus luminescens. J Biol Chem. 2010;285:13525–34. doi: 10.1074/jbc.M109.077339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Braun M, Stuber K, Schlatter Y, Wahli T, Kuhnert P, Frey J. Characterization of an ADP-ribosyltransferase toxin (AexT) from Aeromonas salmonicida subsp. salmonicida. J Bacteriol. 2002;184:1851–8. doi: 10.1128/JB.184.7.1851-1858.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hochmann H, Pust S, von Figura G, Aktories K, Barth H. Salmonella enterica SpvB ADP-ribosylates actin at position arginine-177-characterization of the catalytic domain within the SpvB protein and a comparison to binary clostridial actin-ADP-ribosylating toxins. Biochemistry. 2006;45:1271–7. doi: 10.1021/bi051810w. [DOI] [PubMed] [Google Scholar]
- 72.Wegner A, Aktories K. ADP-ribosylated actin caps the barbed ends of actin filaments. J Biol Chem. 1988;263:13739–42. [PubMed] [Google Scholar]
- 73.Wille M, Just I, Wegner A, Aktories K. ADP-ribosylation of gelsolin-actin complexes by clostridial toxins. J Biol Chem. 1992;267:50–5. [PubMed] [Google Scholar]
- 74.Aktories K, Wegner A. ADP-ribosylation of actin by clostridial toxins. J Cell Biol. 1989;109:1385–7. doi: 10.1083/jcb.109.4.1385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Schwan C, Stecher B, Tzivelekidis T, van Ham M, Rohde M, Hardt WD, Wehland J, Aktories K. Clostridium difficile toxin CDT induces formation of microtubule-based protrusions and increases adherence of bacteria. PLoS Pathog. 2009;5:e1000626. doi: 10.1371/journal.ppat.1000626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Lansbergen G, Akhmanova A. Microtubule plus end: a hub of cellular activities. Traffic. 2006;7:499–507. doi: 10.1111/j.1600-0854.2006.00400.x. [DOI] [PubMed] [Google Scholar]
- 77.Siegrist SE, Doe CQ. Microtubule-induced cortical cell polarity. Genes Dev. 2007;21:483–96. doi: 10.1101/gad.1511207. [DOI] [PubMed] [Google Scholar]
- 78.Geric B, Johnson S, Gerding DN, Grabnar M, Rupnik M. Frequency of binary toxin genes among Clostridium difficile strains that do not produce large clostridial toxins. J Clin Microbiol. 2003;41:5227–32. doi: 10.1128/JCM.41.11.5227-5232.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Popoff MR, Rubin EJ, Gill DM, Boquet P. Actin-specific ADP-ribosyltransferase produced by a Clostridium difficile strain. Infect Immun. 1988;56:2299–306. doi: 10.1128/iai.56.9.2299-2306.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Rupnik M, Kato N, Grabnar M, Kato H. New types of toxin A-negative, toxin B-positive strains among Clostridium difficile isolates from Asia. J Clin Microbiol. 2003;41:1118–25. doi: 10.1128/JCM.41.3.1118-1125.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gonçalves C, Decré D, Barbut F, Burghoffer B, Petit JC. Prevalence and characterization of a binary toxin (actin-specific ADP-ribosyltransferase) from Clostridium difficile. J Clin Microbiol. 2004;42:1933–9. doi: 10.1128/JCM.42.5.1933-1939.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Terhes G, Urbán E, Sóki J, Hamid KA, Nagy E. Community-acquired Clostridium difficile diarrhea caused by binary toxin, toxin A, and toxin B gene-positive isolates in Hungary. J Clin Microbiol. 2004;42:4316–8. doi: 10.1128/JCM.42.9.4316-4318.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pituch H, Rupnik M, Obuch-Woszczatyński P, Grubesic A, Meisel-Mikołajczyk F, Luczak M. Detection of binary-toxin genes (cdtA and cdtB) among Clostridium difficile strains isolated from patients with C. difficile-associated diarrhoea (CDAD) in Poland. J Med Microbiol. 2005;54:143–7. doi: 10.1099/jmm.0.45799-0. [DOI] [PubMed] [Google Scholar]
- 84.Alonso R, Martín A, Peláez T, Marín M, Rodríguez-Creixéms M, Bouza E. Toxigenic status of Clostridium difficile in a large Spanish teaching hospital. J Med Microbiol. 2005;54:159–62. doi: 10.1099/jmm.0.45809-0. [DOI] [PubMed] [Google Scholar]
- 85.Geric B, Rupnik M, Gerding DN, Grabnar M, Johnson S. Distribution of Clostridium difficile variant toxinotypes and strains with binary toxin genes among clinical isolates in an American hospital. J Med Microbiol. 2004;53:887–94. doi: 10.1099/jmm.0.45610-0. [DOI] [PubMed] [Google Scholar]
- 86.Dallal RM, Harbrecht BG, Boujoukas AJ, Sirio CA, Farkas LM, Lee KK, Simmons RL. Fulminant Clostridium difficile: an underappreciated and increasing cause of death and complications. Ann Surg. 2002;235:363–72. doi: 10.1097/00000658-200203000-00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Black SR, Weaver KN, Jones RC, Ritger KA, Petrella LA, Sambol SP, Vernon M, Burton S, Garcia-Houchins S, Weber SG, et al. Clostridium difficile outbreak strain BI is highly endemic in Chicago area hospitals. Infect Control Hosp Epidemiol. 2011;32:897–902. doi: 10.1086/661283. [DOI] [PubMed] [Google Scholar]
- 88.Kuijper EJ, Coignard B, Tüll P, ESCMID Study Group for Clostridium difficile. EU Member States. European Centre for Disease Prevention and Control Emergence of Clostridium difficile-associated disease in North America and Europe. Clin Microbiol Infect. 2006;12(Suppl 6):2–18. doi: 10.1111/j.1469-0691.2006.01580.x. [DOI] [PubMed] [Google Scholar]
- 89.Spigaglia P, Mastrantonio P. Comparative analysis of Clostridium difficile clinical isolates belonging to different genetic lineages and time periods. J Med Microbiol. 2004;53:1129–36. doi: 10.1099/jmm.0.45682-0. [DOI] [PubMed] [Google Scholar]
- 90.Barbut F, Mastrantonio P, Delmée M, Brazier J, Kuijper E, Poxton I, European Study Group on Clostridium difficile (ESGCD) Prospective study of Clostridium difficile infections in Europe with phenotypic and genotypic characterisation of the isolates. Clin Microbiol Infect. 2007;13:1048–57. doi: 10.1111/j.1469-0691.2007.01824.x. [DOI] [PubMed] [Google Scholar]
- 91.Bauer MP, Notermans DW, van Benthem BH, Brazier JS, Wilcox MH, Rupnik M, Monnet DL, van Dissel JT, Kuijper EJ, ECDIS Study Group Clostridium difficile infection in Europe: a hospital-based survey. Lancet. 2011;377:63–73. doi: 10.1016/S0140-6736(10)61266-4. [DOI] [PubMed] [Google Scholar]
- 92.Rupnik M. Is Clostridium difficile-associated infection a potentially zoonotic and foodborne disease? Clin Microbiol Infect. 2007;13:457–9. doi: 10.1111/j.1469-0691.2007.01687.x. [DOI] [PubMed] [Google Scholar]
- 93.Keel K, Brazier JS, Post KW, Weese S, Songer JG. Prevalence of PCR ribotypes among Clostridium difficile isolates from pigs, calves, and other species. J Clin Microbiol. 2007;45:1963–4. doi: 10.1128/JCM.00224-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Jhung MA, Thompson AD, Killgore GE, Zukowski WE, Songer G, Warny M, Johnson S, Gerding DN, McDonald LC, Limbago BM. Toxinotype V Clostridium difficile in humans and food animals. Emerg Infect Dis. 2008;14:1039–45. doi: 10.3201/eid1407.071641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Rodriguez-Palacios A, Staempfli HR, Duffield T, Weese JS. Clostridium difficile in retail ground meat, Canada. Emerg Infect Dis. 2007;13:485–7. doi: 10.3201/eid1303.060988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Songer JG, Trinh HT, Dial SM, Brazier JS, Glock RD. Equine colitis X associated with infection by Clostridium difficile NAP1/027. J Vet Diagn Invest. 2009;21:377–80. doi: 10.1177/104063870902100314. [DOI] [PubMed] [Google Scholar]
- 97.Hensgens MP, Keessen EC, Squire MM, Riley TV, Koene MG, de Boer E, Lipman LJ, Kuijper EJ, European Society of Clinical Microbiology and Infectious Diseases Study Group for Clostridium difficile (ESGCD) Clostridium difficile infection in the community: a zoonotic disease? Clin Microbiol Infect. 2012;18:635–45. doi: 10.1111/j.1469-0691.2012.03853.x. [DOI] [PubMed] [Google Scholar]
- 98.Gould LH, Limbago B. Clostridium difficile in food and domestic animals: a new foodborne pathogen? Clin Infect Dis. 2010;51:577–82. doi: 10.1086/655692. [DOI] [PubMed] [Google Scholar]
- 99.Barbut F, Decré D, Lalande V, Burghoffer B, Noussair L, Gigandon A, Espinasse F, Raskine L, Robert J, Mangeol A, et al. Clinical features of Clostridium difficile-associated diarrhoea due to binary toxin (actin-specific ADP-ribosyltransferase)-producing strains. J Med Microbiol. 2005;54:181–5. doi: 10.1099/jmm.0.45804-0. [DOI] [PubMed] [Google Scholar]
- 100.Walk ST, Micic D, Jain R, Lo ES, Trivedi I, Liu EW, Almassalha LM, Ewing SA, Ring C, Galecki AT, et al. Clostridium difficile ribotype does not predict severe infection. Clin Infect Dis. 2012;55:1661–8. doi: 10.1093/cid/cis786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Cloud J, Noddin L, Pressman A, Hu M, Kelly C. Clostridium difficile strain NAP-1 is not associated with severe disease in a nonepidemic setting. Clin Gastroenterol Hepatol. 2009;7:868–73, e2. doi: 10.1016/j.cgh.2009.05.018. [DOI] [PubMed] [Google Scholar]
- 102.Wilson V, Cheek L, Satta G, Walker-Bone K, Cubbon M, Citron D, Gerding DN, Llewelyn MJ. Predictors of death after Clostridium difficile infection: a report on 128 strain-typed cases from a teaching hospital in the United Kingdom. Clin Infect Dis. 2010;50:e77–81. doi: 10.1086/653012. [DOI] [PubMed] [Google Scholar]
- 103.Morgan OW, Rodrigues B, Elston T, Verlander NQ, Brown DF, Brazier J, Reacher M. Clinical severity of Clostridium difficile PCR ribotype 027: a case-case study. PLoS One. 2008;3:e1812. doi: 10.1371/journal.pone.0001812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Petrella LA, Sambol SP, Cheknis A, Nagaro K, Kean Y, Sears PS, Babakhani F, Johnson S, Gerding DN. Decreased cure and increased recurrence rates for Clostridium difficile infection caused by the epidemic C. difficile BI strain. Clin Infect Dis. 2012;55:351–7. doi: 10.1093/cid/cis430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Miller M, Gravel D, Mulvey M, Taylor G, Boyd D, Simor A, Gardam M, McGeer A, Hutchinson J, Moore D, et al. Health care-associated Clostridium difficile infection in Canada: patient age and infecting strain type are highly predictive of severe outcome and mortality. Clin Infect Dis. 2010;50:194–201. doi: 10.1086/649213. [DOI] [PubMed] [Google Scholar]
- 106.Goldenberg SD, French GL. Lack of association of tcdC type and binary toxin status with disease severity and outcome in toxigenic Clostridium difficile. J Infect. 2011;62:355–62. doi: 10.1016/j.jinf.2011.03.001. [DOI] [PubMed] [Google Scholar]
- 107.Bacci S, Mølbak K, Kjeldsen MK, Olsen KE. Binary toxin and death after Clostridium difficile infection. Emerg Infect Dis. 2011;17:976–82. doi: 10.3201/eid1706.101483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Barbut F, Gariazzo B, Bonné L, Lalande V, Burghoffer B, Luiuz R, Petit JC. Clinical features of Clostridium difficile-associated infections and molecular characterization of strains: results of a retrospective study, 2000-2004. Infect Control Hosp Epidemiol. 2007;28:131–9. doi: 10.1086/511794. [DOI] [PubMed] [Google Scholar]
- 109.Stewart DB, Berg A, Hegarty J. Predicting recurrence of C. difficile colitis using bacterial virulence factors: binary toxin is the key. J Gastrointest Surg. 2013;17:118–24, discussion 124-5. doi: 10.1007/s11605-012-2056-6. [DOI] [PubMed] [Google Scholar]
- 110.Walker AS, Eyre DW, Wyllie DH, Dingle KE, Griffiths D, Shine B, Oakley S, O’Connor L, Finney J, Vaughan A, et al. Infections in Oxfordshire Research Database Relationship between bacterial strain type, host biomarkers, and mortality in Clostridium difficile infection. Clin Infect Dis. 2013;56:1589–600. doi: 10.1093/cid/cit127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Geric B, Carman RJ, Rupnik M, Genheimer CW, Sambol SP, Lyerly DM, Gerding DN, Johnson S. Binary toxin-producing, large clostridial toxin-negative Clostridium difficile strains are enterotoxic but do not cause disease in hamsters. J Infect Dis. 2006;193:1143–50. doi: 10.1086/501368. [DOI] [PubMed] [Google Scholar]
- 112.Kuehne SA, Collery MM, Kelly ML, Cartman ST, Cockayne A, Minton NP. The importance of toxin A, toxin B and CDT in virulence of an epidemic Clostridium difficile strain. J Infect Dis. 2013 doi: 10.1093/infdis/jit426. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Carman RJ, Stevens AL, Lyerly MW, Hiltonsmith MF, Stiles BG, Wilkins TD. Clostridium difficile binary toxin (CDT) and diarrhea. Anaerobe. 2011;17:161–5. doi: 10.1016/j.anaerobe.2011.02.005. [DOI] [PubMed] [Google Scholar]
- 114.Lowy I, Molrine DC, Leav BA, Blair BM, Baxter R, Gerding DN, Nichol G, Thomas WD, Jr., Leney M, Sloan S, et al. Treatment with monoclonal antibodies against Clostridium difficile toxins. N Engl J Med. 2010;362:197–205. doi: 10.1056/NEJMoa0907635. [DOI] [PubMed] [Google Scholar]
- 115.Babcock GJ, Broering TJ, Hernandez HJ, Mandell RB, Donahue K, Boatright N, Stack AM, Lowy I, Graziano R, Molrine D, et al. Human monoclonal antibodies directed against toxins A and B prevent Clostridium difficile-induced mortality in hamsters. Infect Immun. 2006;74:6339–47. doi: 10.1128/IAI.00982-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Barth H, Blöcker D, Aktories K. The uptake machinery of clostridial actin ADP-ribosylating toxins--a cell delivery system for fusion proteins and polypeptide drugs. Naunyn Schmiedebergs Arch Pharmacol. 2002;366:501–12. doi: 10.1007/s00210-002-0626-y. [DOI] [PubMed] [Google Scholar]
- 117.Barth H, Stiles BG. Binary actin-ADP-ribosylating toxins and their use as molecular Trojan horses for drug delivery into eukaryotic cells. Curr Med Chem. 2008;15:459–69. doi: 10.2174/092986708783503195. [DOI] [PubMed] [Google Scholar]
- 118.Elliott B, Squire MM, Thean S, Chang BJ, Brazier JS, Rupnik M, Riley TV. New types of toxin A-negative, toxin B-positive strains among clinical isolates of Clostridium difficile in Australia. J Med Microbiol. 2011;60:1108–11. doi: 10.1099/jmm.0.031062-0. [DOI] [PubMed] [Google Scholar]