

Abstract
The requirement of center asymmetry for the creation of second harmonic generation (SHG) signals makes it an attractive technique for visualizing changes in interfacial layers such as the plasma membrane of biological cells. In this article, we explore the use of lipophilic SHG probes to detect minute perturbations in the plasma membrane. Three candidate probes, Di-4-ANEPPDHQ (Di-4), FM4-64, and all-trans-retinol, were evaluated for SHG effectiveness in Jurkat cells. Di-4 proved superior with both strong SHG signal and limited bleaching artifacts. To test whether rapid changes in membrane symmetry could be detected using SHG, we exposed cells to nanosecond-pulsed electric fields, which are believed to cause formation of nanopores in the plasma membrane. Upon nanosecond-pulsed electric fields exposure, we observed an instantaneous drop of ∼50% in SHG signal from the anodic pole of the cell. When compared to the simultaneously acquired fluorescence signals, it appears that the signal change was not due to the probe diffusing out of the membrane or changes in membrane potential or fluidity. We hypothesize that this loss in SHG signal is due to disruption in the interfacial nature of the membrane. The results show that SHG imaging has great potential as a tool for measuring rapid and subtle plasma membrane disturbance in living cells.
As the epicenter for many cellular functions, understanding the dynamics of the plasma membrane is important to monitoring biological phenomena. External forces acting upon the plasma membrane (e.g., electric, mechanical) have been shown to cause rapid disturbances, often resulting in dramatic changes in cell physiology (1–3). To understand this interaction, a minimally invasive, highly sensitive imaging technique that enables monitoring the structure of the plasma membrane is needed. Lipophilic dyes, which embed themselves into lipid membranes, are sensitive to the surrounding electric field and, therefore, report changes in membrane fluidity as well as voltage due to the capacitive nature of the membranes (4,5). This sensitivity is typically detected as a shift in the fluorescence emission spectrum. Localization of the fluorescence signal to only the plasma membrane is difficult because the probes also label internal membrane structures. Thus, to overcome this lack of spatial selectivity, second harmonic generation (SHG) has been used as an alternative to fluorescence for membrane imaging (6,7).
In SHG, a second-order nonlinear polarization is induced by electronic disruption of a probe molecule from the electromagnetic field of the incident laser beam. This polarization generates oscillating dipole moments that reradiate light at twice the energy of the excitation beam. The induction of this dipole is sensitive to the static electric field surrounding the probe and the steady-state molecular polarization of the probe molecule. These properties make SHG probes useful for monitoring changes in biological membranes.
First, as the voltage potential across the membrane changes, the static electric field around the probe also shifts, making the probe sensitive to these variations (7). Several SHG probes have, therefore, been employed to monitor plasma membrane potential (7,8).
Second, because the dipole is affected by the steady-state molecular polarization of the probe itself, a SHG signal is only produced in materials that lack a center of inversion symmetry. In the centrosymmetric case, any emitted radiation is cancelled out by destructive interference. The properties of an interfacial environment, such as a cellular plasma membrane, not only provide the necessary asymmetry, but cause the polarized lipophilic dyes to be aligned in respect to the interface, instead of being randomly distributed as they would in a bulk environment. This alignment allows the generation of a coherent SHG signal from the plasma membrane while the rest of the cell remains nearly signal-free (6,7).
We investigated whether the alignment sensitivity of the SHG response could be used to detect minute changes in the organization of the plasma membrane. Jurkat clone E6-1 human T-lymphocytes with a spherical morphology were selected for optimum signal clarity and cultured as directed by American Type Culture Collection (ATCC, Manassas, VA) with 1 I.U./mL penicillin and 0.1 μg/mL streptomycin. Cells were added to 35-mm poly-L-lysine-coated glass-bottomed dishes (MatTek, Ashland, MA) and incubated for 1 h in growth media to allow adherence. Before loading, the cells were rinsed with a buffer consisting of 135 mM NaCl, 5 mM KCl, 2 mM MgCl2, 10 mM HEPES, 10 mM glucose, 2 mM CaCl2, pH 7.4, 290–310 mOsm. SHG probes, Di-4-ANEPPDHQ (Di-4) (5 μM final concentration), FM4-64 (15 μM) or ATR (100 μM, 1 mg/mL BSA) were added to the buffer solution and incubated for 1 h. Cellular imaging was performed in the labeling buffer to limit diffusion of the probe molecules out of the cell membranes.
A Ti:sapphire oscillator at 980 nm (Coherent Chameleon, 130 fs, 80 MHz, ∼15 mW at the sample; Coherent Laser, Santa Clara, CA) was coupled through the scan head of a modified model No. TCS SP5 II (Leica Geosystems, Norcross, GA) for SHG and multiphoton-excited fluorescence imaging (40×, water, 1.1 NA) using resonant scanning. SHG signal was collected in transmission by a photomultiplier tube after 680-nm shortpass and 485/25-nm bandpass filters; simultaneous fluorescence signal was collected in the epi-direction by two non-descanned photomultiplier tubes with 540/60-nm and 650/60-nm bandpass filters.
Three SHG probes previously used to monitor voltage or membrane order in living cells were tested (8–10). Although ATR is reported to be effective in monitoring membrane voltage, we obtained nearly no SHG signal, despite successful loading as indicated by the fluorescence signal (Fig. 1). When FM4-64 and Di-4 were loaded to similar fluorescence intensities, nearly equivalent SHG signal was collected. Di-4 did appear to have a greater internalization of the dye. However, after the first frame, the FM4-64 signal dropped considerably (Fig. 1 b), an observation reported as a membrane voltage-independent bleaching effect (8). This drop in signal recovered after excitation was blocked for several seconds, but quantification of the response was difficult. Di-4 did not suffer as dramatic a drop in signal upon excitation, and still had sufficient SHG signal/noise after several seconds, so it was used in all further experiments.
Figure 1.
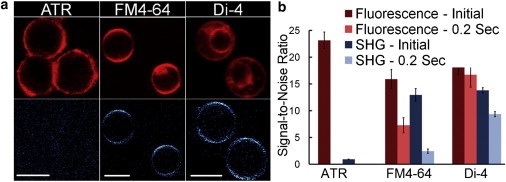
(a) Fluorescence (top) and SHG (bottom) images for the three probes. (b) Signal/noise for the fluorescence and SHG for the initial frame and shortly after beginning acquisition. Error bars represent the mean ± SE (n = 10). Scale bar is 10 μm.
To test whether Di-4 would report a rapid change in membrane organization, we applied a single nanosecond-duration pulsed electric field (nsPEF) to the labeled cell. These ultrabrief, high-intensity (MV/m) pulses differ from longer (μs-ms), lower-intensity (kV/m) pulses traditionally associated with electroporation in induced cellular response (3,11,12). Through selective uptake of small ions (Ca2+, Ti+) with limited uptake of propidium iodide, nsPEF have been previously postulated to cause nanopores (<2 nm diameter) in the plasma membrane. In contrast with a previous study observing poration resulting from traditional electroporation (13), the brevity of this apparently novel cellular insult allows for the decoupling of the mechanical effects of the pulse on the membrane from the electrical effects of the pulse itself. A single pulse, generated by a custom pulse generator, was delivered to the cells using a pair of 125-μm diameter tungsten electrodes, separated edge-to-edge by 150 μm, as previously described in Ibey et al. (14). For maximum visualization of changes in the SHG signal, a half-wave plate was placed before the scan head to align the polarization of the laser such that the brightest signals from the plasma membrane were at the poles facing the electrodes.
The Di-4 SHG signal in response to a single 16.7 kV/cm, 600-ns nsPEF is shown in Fig. 2. Before the pulse, the intensity of the SHG signal is high at each of these poles. Immediately after the pulse, the SHG intensity drops by ∼50% on the side of the cell facing the anodic electrode, whereas little intensity is lost at the other pole. This response is plotted in Fig. 2 (pulse applied at 2 s), where it is apparent that the response is near instantaneous with little recovery in signal in the 5 s postexposure. The SHG response matches the previously observed effect of this stimulus, where ion uptake displayed a polar dependence and persisted for a number of minutes (11,12). Images taken 5 min after an nsPEF exposure are also shown in Fig. 2. These images confirm the eventual recovery of the cell and the corresponding return of SHG signal to preexposure levels.
Figure 2.
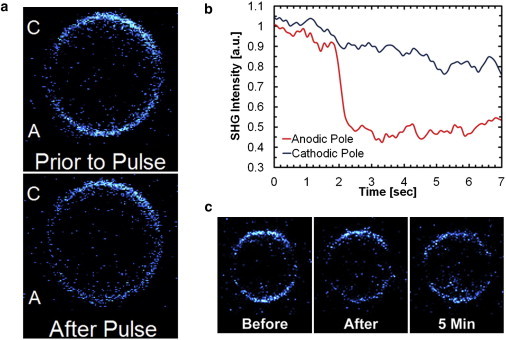
(a) SHG images showing drop in signal on the anodic (or A-pole) of the cell. (b) Time trace of SHG response with the electrical pulse applied at 2 s that shows a near-instantaneous drop in the SHG signal at the anodic pole of the cell. (c) SHG image preexposure, immediately postexposure, and then 5-min postexposure showing recovery of the SHG signal.
To decouple membrane disturbance from environmental changes around the membrane, we compared the SHG response to the simultaneously acquired fluorescence signal. Because fluorescence is not subject to the strict orientation requirement of SHG, the plasma membrane fluorescence signal provides an indication of the membrane fluidity and/or potential. Despite the dramatic shift in SHG intensity on the anodic pole upon the electrical pulse exposure (Fig. 3 a), the fluorescence channels display little response from the equivalent membrane sections with the exception of photobleaching and a slight increase in signal in both emission bands on the anodic side (Fig. 3, b and c). The shading in these graphs represents the mean ± SE for six cells. Although this slight increase may indicate that a small amount of dye is simply diffusing in or out of the membrane upon exposure, the fluorescence response is not as rapid or as lasting as the SHG response. Change in membrane fluidity or voltage can also be quantized using these fluorescence signals and a value known as the generalized polarization (GP) (4),
| (1) |
As with the raw intensity of the individual signals, the GP value for the membrane (Fig. 3 d) shows no significant shift, indicating that the membrane is likely not transitioning between a more raft- and fluidlike state. Thus, it seems likely that the dye was initially aligned in the tightly-packed ordered membrane so that the probes were able to generate a SHG photon. As shown in Fig. 3 e, we postulate that upon electrical pulse exposure, the membrane was disrupted by the formation with nanopores giving the probe molecules the flexibility to disorient within the membrane. The resulting alignment of the probes is more isotropic in nature, thereby limiting the probes probability of producing a SHG photon. The fluorescence signal remained, however, indicating that the probes remained active in the membrane.
Figure 3.
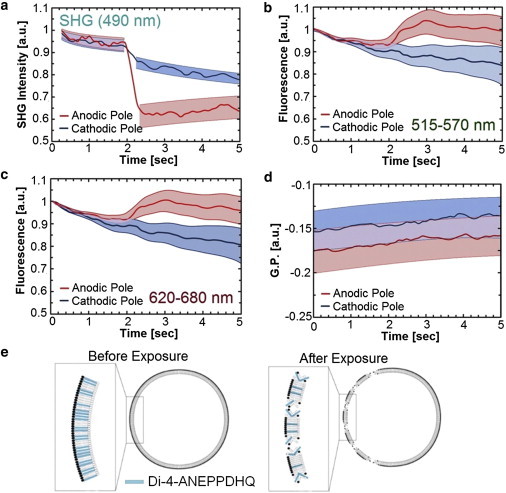
(a) Average SHG signal showing the dramatic drop in signal on the anodic pole at the pulse application (2 s). (b and c) Simultaneous TPF signals showing nearly no instantaneous change at the pulse application. (d) GP showing no observable changes in the membrane potential or fluidity after the pulse. (Shaded areas) Fit to the mean ± SE for each trace (n = 6). (e) Conceptualization of the hypothesized membrane disruption underlying the observed change in SHG response.
Thus, by taking advantage of the selection criteria of SHG, we were able to successfully use the SHG probe, Di-4, to monitor rapid disruption of the plasma membrane. Because SHG can only be generated when the probes are aligned in the plasma membrane, the SHG signal diminishes significantly upon disruption. The simultaneous collection of the multiphoton-excited fluorescence signal was advantageous in that it demonstrated that the probes did not simply diffuse out of the membrane, did not appear to be energetically disrupted by the electric pulse, and showed that the membrane changes were not simply a change in lipid order. We believe that this technique holds tremendous potential for use in the study of how external stimuli interact with and change the orientation of biological membranes. Such knowledge may allow for further understanding of how manipulation of cells and biological systems can be achieved using external stimuli.
Acknowledgments
This work was supported by the Air Force Office of Scientific Research’s Laboratory Research Independent Research (LRIR) grants No. 12RH01COR and No. 13RH08COR. E.K.M. performed this research under the 711 HPW Repperger Summer Research Internship program led by Dr. Morley Stone and Dr. David Luginbuhl. E.K.M. also thanks the Achievement Rewards for College Scientists (ARCS) Foundation for their support.
References and Footnotes
- 1.Neumann E., Schaefer-Ridder M., Hofschneider P.H. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982;1:841–845. doi: 10.1002/j.1460-2075.1982.tb01257.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Miller D.L., Pislaru S.V., Greenleaf J.E. Sonoporation: mechanical DNA delivery by ultrasonic cavitation. Somat. Cell Mol. Genet. 2002;27:115–134. doi: 10.1023/a:1022983907223. [DOI] [PubMed] [Google Scholar]
- 3.Vernier P.T., Sun Y., Gundersen M.A. Calcium bursts induced by nanosecond electric pulses. Biochem. Biophys. Res. Commun. 2003;310:286–295. doi: 10.1016/j.bbrc.2003.08.140. [DOI] [PubMed] [Google Scholar]
- 4.Parasassi T., Krasnowska E.K., Gratton E. Laurdan and prodan as polarity-sensitive fluorescent membrane probes. J. Fluoresc. 1998;8:365–373. [Google Scholar]
- 5.Jin L., Millard A.C., Loew L.M. Characterization and application of a new optical probe for membrane lipid domains. Biophys. J. 2006;90:2563–2575. doi: 10.1529/biophysj.105.072884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Moreaux L., Sandre O., Mertz J. Coherent scattering in multi-harmonic light microscopy. Biophys. J. 2001;80:1568–1574. doi: 10.1016/S0006-3495(01)76129-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Campagnola P.J., Wei M.D., Loew L.M. High-resolution nonlinear optical imaging of live cells by second harmonic generation. Biophys. J. 1999;77:3341–3349. doi: 10.1016/S0006-3495(99)77165-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sacconi L., Dombeck D.A., Webb W.W. Overcoming photodamage in second-harmonic generation microscopy: real-time optical recording of neuronal action potentials. Proc. Natl. Acad. Sci. USA. 2006;103:3124–3129. doi: 10.1073/pnas.0511338103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Theer P., Denk W., Detwiler P.B. Second-harmonic generation imaging of membrane potential with retinal analogues. Biophys. J. 2011;100:232–242. doi: 10.1016/j.bpj.2010.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Obaid A.L., Loew L.M., Salzberg B.M. Novel naphthylstyryl-pyridium potentiometric dyes offer advantages for neural network analysis. J. Neurosci. Methods. 2004;134:179–190. doi: 10.1016/j.jneumeth.2003.11.011. [DOI] [PubMed] [Google Scholar]
- 11.Pakhomov A.G., Bowman A.M., Schoenbach K.H. Lipid nanopores can form a stable, ion channel-like conduction pathway in cell membrane. Biochem. Biophys. Res. Commun. 2009;385:181–186. doi: 10.1016/j.bbrc.2009.05.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Beier H.T., Roth C.C., Ibey B.L. Resolving the spatial kinetics of electric pulse-induced ion release. Biochem. Biophys. Res. Commun. 2012;423:863–866. doi: 10.1016/j.bbrc.2012.06.055. [DOI] [PubMed] [Google Scholar]
- 13.Zalvidea D., Claverol-Tinturé E. Second harmonic generation for time-resolved monitoring of membrane pore dynamics subserving electroporation of neurons. Biomed. Opt. Express. 2011;2:305–314. doi: 10.1364/BOE.2.000305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ibey B.L., Mixon D.G., Pakhomov A.G. Plasma membrane permeabilization by trains of ultrashort electric pulses. Bioelectrochemistry. 2010;79:114–121. doi: 10.1016/j.bioelechem.2010.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]