

Abstract
BACKGROUND/OBJECTIVE
Licorice has been shown to possess cancer chemopreventive effects. However, glycyrrhizin, a major component in licorice, was found to interfere with steroid metabolism and cause edema and hypertension. The roasting process of licorice modifies the chemical composition and converts glycyrrhizin to glycyrrhetinic acid. The purpose of this study was to examine the anti-carcinogenic effects of the ethanol extract of roasted licorice (EERL) and to identify the active compound in EERL.
MATERIALS/METHODS
Ethanol and aqueous extracts of roasted and un-roasted licorice were prepared. The active fraction was separated from the methylene chloride (MC)-soluble fraction of EERL and the structure of the purified compound was determined by nuclear magnetic resonance spectroscopy. The anti-carcinogenic effects of licorice extracts and licochalcone A was evaluated using a MTT assay, Western blot, flow cytometry, and two-stage skin carcinogenesis model.
RESULTS
EERL was determined to be more potent and efficacious than the ethanol extract of un-roasted licorice in inhibiting the growth of DU145 and MLL prostate cancer cells, as well as HT-29 colon cancer cells. The aqueous extracts of un-roasted and roasted licorice showed minimal effects on cell growth. EERL potently inhibited growth of MCF-7 and MDA-MB-231 breast, B16-F10 melanoma, and A375 and A2058 skin cancer cells, whereas EERL slightly stimulated the growth of normal IEC-6 intestinal epithelial cells and CCD118SK fibroblasts. The MC-soluble fraction was more efficacious than EERL in inhibiting DU145 cell growth. Licochalcone A was isolated from the MC fraction and identified as the active compound of EERL. Both EERL and licochalcone A induced apoptosis of DU145 cells. EERL potently inhibited chemically-induced skin papilloma formation in mice.
CONCLUSIONS
Non-polar compounds in EERL exert potent anti-carcinogenic effects, and that roasted rather than un-roasted licorice should be favored as a cancer preventive agent, whether being used as an additive to food or medicine preparations.
Keywords: Roasted licorice roots, apoptosis, licochalcone A, cancer
INTRODUCTION
Over two-thirds of cancer-related deaths could likely be prevented through lifestyle modification, particularly through dietary means. [1] Cancer chemoprevention was defined as the use of synthetic and naturally occurring compounds to prevent, reverse, or delay the process of carcinogenesis, an overall process in which distinct changes of cellular and molecular events occur in a multistep manner. [2] Recently, interest in dietary plant bioactive components that show no/low toxicity for potential cancer chemoprevention has increased substantially. [3] Apoptosis is defined as a controlled type of cell death, and apoptotic processes become deregulated in cancer cells (Reviewed in [4]). Because apoptosis can be induced by a variety of physiological and pharmacological agents, plant bioactive compounds that effectively induce apoptosis of cancer cells can be used as cancer chemopreventive agents.
Licorice is a well-known plant, which is used to add flavor to foods, beverages, and tobacco, and it is used as a medicinal plant (reviewed in [5]). There is much literature on the biological effects of the constituents of licorice, particularly in terms of their anti-inflammatory, anti-microbial, anti-oxidant, anti-ulcer, cytoprotective, and cytotoxic activities (reviewed in [5,6]). However, some studies found that licorice interfered with steroid metabolism and caused edema and hypertension. [7,8,9] Glycyrrhizin, a major component of licorice, inhibits the metabolism of cortisol (reviewed in [10]) and can lead to acute and chronic cases of hypertension and hypokalemia. [11] In traditional oriental medicine roasted licorice has been used rather than raw licorice. The roasting process of licorice results the conversion of glycyrrhizin to glycyrrhetinic acid, [12,13] and thus, reduces the levels of glycyrrhizin. [14] Additionally, it has been shown that roasted licorice is more potent than un-roasted licorice in inhibiting allergic [14] and inflammatory [15] responses, and in protecting from neuronal injury by transient forebrain ischemia. [13]
Despite the considerable number of physiological studies which have been conducted on licorice, there have been no studies to directly compare the anti-carcinogenic effect of un-roasted licorice with that of roasted licorice. In the present study, we observed that the ethanol extract of roasted licorice (EERL), containing an increased amount of non-polar components, exhibits remarkably more potent inhibitory effect on the growth of cancer cells than the ethanol extract of un-roasted licorice (EEUL). We also demonstrated that the topical application of EERL inhibits 7,12-dimethylbenz(a)anthracene (DMBA)-initiated, 12-O-tetradecanoylphorbol-13-acetate (TPA)-promoted skin papilloma formation in mice. Additionally, we identified licochalcone A as the active compound of EERL, and demonstrated that both EERL and licochalcone A potently induced apoptosis of DU145 human prostate cancer cells.
MATERIALS AND METHODS
Materials
The following reagents were purchased from the indicated suppliers: 3-[4,5-dimethylthiazol-2-ly]-2,5-diphenyltetrazolium bromide (MTT) and 5,5',6,6'-tetrachloro-1,1',3,3'-tetraethyl-imidacarbocyanine iodide (JC-1), DMBA, TPA (Sigma, St. Louis, MO, USA); anti-cytochrome c antibody, phycoerythrin (PE)-conjugated annexin V, and 7-amino-actinomycin D (7-AAD) (BD Pharmingen, Franklin Lake, NJ, USA); antibodies against cleaved poly (ADP-ribose) polymerase (PARP), Bcl-xL, and cleaved caspase-3, -7, -8, and -9 (Cell Signaling Technology, Beverly, MA, USA); antibodies against Bcl-2, Bax, Bad and heat shock protein (HSP) 60 (Santa Cruz Biotechnology, Santa Cruz, CA, USA); licochalcone A (Calbiochem, La Jolla, CA, USA). Dried licorice roots (Glycyrrhiza inflata Bat., Dea Guang Medical, Chuncheon, South Korea).
Preparation of ethanol and aqueous extracts of roasted and un-roast licorice
To compare the anti-carcinogenic effects of roasted and unroasted licorice, we prepared 4 different extracts; EERL, EEUL, aqueous extract of roasted licorice (WERL), and aqueous extract of un-roasted licorice (WEUL). In order to prepare roasted licorice, licorice roots (200 g) were sliced and placed in a glass bottle that was rotated for 100 min at a speed of 30 rpm in a heated oil bath at 150℃. The roasted or un-roasted licorice slices were ground into a fine powder in a blender (Waring, Model 51BL30). To prepare ethanol extracts, the powder (100 g) was refluxed in 900 mL of 95% ethanol at 80℃ for 3 h. This extraction procedure was repeated 3 times. The ethanol extracts were filtered (Whatman #2) and dried in a vacuum at a low temperature (< 40℃). The yields from roasted and un-roasted licorice were 6.5 g/100 g and 5.9 g/100 g, respectively.
In order to prepare the aqueous extracts, 100 g of each powder was refluxed in 2 L of water at 100℃ for 10 h. The extracts were filtered and freeze-dried. The yields were 20.5 g/100 g and 19.2 g /100 g from roasted and un-roasted licorice, respectively.
The fractionation of EERL and identification of the active compound
The EERL powder (6.2 g) was suspended in distilled water and then partitioned with methylene chloride (MC; yield, 3.2 g). The MC-soluble fraction (3.0 g) was subjected to medium pressure liquid chromatograph (MPLC) in vacuo (VLC) with silica gel (100 g, 70-230 mesh, Merck, Germany), eluted by gradient systems of n-pentane-n-hexane-ethyl acetate (10:0:0-0:5:5). Twenty one fractions were isolated, and the ability of these 21 fractions to inhibit DU145 cell growth was examined by MTT assays. MPLC fraction # 5 (0.8 g) was determined to be most active in inhibiting DU145 cell growth, was further purified by recrystallization with ethyl acetate using thin layer chromatography and recycle high performance liquid chromatography, and compound 1 was obtained as yellow powder (final yield, 257 mg/100 g roasted licorice). The structure of compound 1 was identified by 1H NMR and 13C NMR. 1H NMR and 13C NMR spectra were recorded on a Bruker DPX 400 (400 MHz) spectrometer with tetramethylsilane as an internal standard and was compared with literature values. The detailed data from 1H NMR and 13C NMR are presented as Supplement 1.
Cell culture
DU145 human and MAT-Ly-Lu (MLL) rat prostate cancer cells, HT-29 human colon cancer cells, MCF-7 and MDA-MB-231 human breast cancer cells, B16-F10 mouse melanoma cells, A375 and A2058 human skin cancer cells, IEC-6 rat intestinal crypt cells, and CCD1108SK normal fibroblasts were obtained from the American Type Culture Collection (Manassas, VA, USA). DU145, MLL, HT-29, IEC-6 and CCD1108SK cells were maintained in Dulbeco's Modified Eagle's Medium (DMEM)/F12 containing 100 mL/L fetal bovine serum (FBS). MCF-7 and MDA-MB-231 cells were maintained in DMEM/F12 containing 100 mL/L FBS and 300 mg/L L-glutamine with 10 mg/L (MCF-7) or 2 mg/L (MDA-MB-231) insulin. B16-F10, A375 and A2058 cells were maintained in DMEM containing 100 mL/L FBS. All cells were maintained in the presence of 100,000 U/L penicillin and 100 mg/L streptomycin.
Cell viability assay
In order to assess the effects of licorice extracts on the growth of various cells, cells were plated in multi-well plates with their respective maintenance media. The cell monolayers were serum-deprived or serum-starved for 24 h with the following media: DMEM/F-12 supplemented with 10 mL/L charcoal-stripped FBS (DU145, MLL); DMEM/F-12 supplemented with 5 mg/L transferrin, 1 g/L BSA, and 5 µg/L selenium (HT-29, IEC-6, CCD 1108SK); DMEM/F-12 supplemented with 5 mg/L transferrin, 1 g/L BSA, 5 µg/L selenium, and 10 mL/L L-glutamine (MCF-7 and MDA-MB-231); and DMEM (B16-F10, A375 and A2058). After serum-deprivation or starvation, we replaced the medium with fresh serum-deprivation or starvation medium, with or without various concentrations of licorice extracts, the MC fraction, MPLC fractions or licochalcone A. Viable cell numbers were estimated via MTT assays. We utilized the serum-deprivation or -starvation media to minimize the possible effects of various phytochemicals present in the FBS. Serum deprivation medium (SDM) contained 1% FBS while serum starvation medium (SFM) contained 0% FBS, 5 mg/L transferrin, 1 g/L BSA, and 5 µg/L selenium. Both SFM and SDM supported cell growth as judged by MTT assays at least for a few days. Cell viability of these cancer cells was not differentially affected under the current experimental conditions. As shown in Table 1 and Fig. 1, the numbers of cells were increased during the experimental period. EERL, EEUL, the MC fraction, MPLC fractions and licochalcone A were dissolved in dimethylsulfoxide (DMSO), and the aqueous extracts of licorice were dissolved in distilled water. Control cells were treated with DMSO or H2O at a final concentration of 1 mL/L.
Table 1.
The effect of the ethanol extract of roasted licorice (EERL) on viable cell numbers of several cancer cells, normal intestinal epithelial cells, and normal skin fibroblasts.
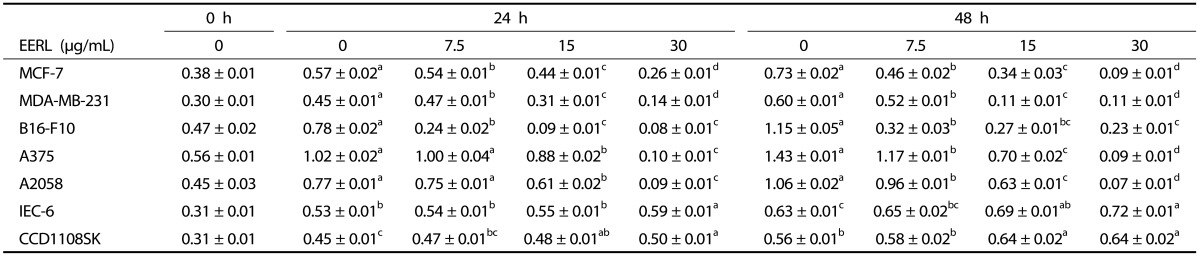
Cells were incubated for 24 or 48 h in serum-free medium or serum-deprivation medium containing 0-30 mg/mL of the EERL. Cell numbers were estimated by MTT assay. Absorbance data (at 570 nm) represent the mean ± SEM (n = 6). Means with different letters differ significantly, P < 0.05.
Fig. 1.

Ethanol extract of roasted licorice (EERL) is more potent and efficacious in inhibiting cancer cell growth than ethanol extract of un-roasted licorice (EEUL). Tumor cells were serum-deprived or starved with their respective media and were incubated with the indicated extract as described in the Methods section. The viable cell numbers were estimated by MTT assay. Each bar represents the mean ± SEM. Means with different letters differ significantly, P < 0.05.
Quantification of apoptotic cells and measurement of the mitochondrial membrane potential (Δψm)
To quantify apoptotic cell numbers, serum-deprived DU145 cells were treated with various concentrations of EERL or licochalcone A for 24 h. The number of early apoptotic cells was assessed by PE-annexin V and 7-AAD staining as previously described. [16]
To measure the mitochondrial membrane potential, serum-deprived DU145 cells were treated for 18 h with various concentrations of EERL and the dual-emission potential-sensitive probe JC-1 was utilized as previously described. [16]
Cell cycle analysis by flow cytometry
Cells were plated in 24-well plates at a density of 50,000 cells/well and subsequently serum-deprived and treated with EERL or licochalcone A for 24 h. After treatment, the cells were collected, fixed, and stained with 0.5 g/L propidium iodide. DNA content data were acquired using FACScan (Becton Dickinson, Franklin Lakes, NJ) and analyzed using Modfit version 1.2 software.
Western blot analysis
Cells were lysed as previously described. [17] Cytosolic and mitochondrial proteins were separated as described by Eguchi et al. [18] The protein contents of total cell lysates, cytoplasmic and mitochondrial fractions were determined using a BCA protein assay kit (Pierce, Rockford, IL, USA). Western blot analyses were conducted as previously described. [17]
Two-stage skin carcinogenesis in mice
Female ICR mice (6 weeks of age) were divided randomly into four groups, each consisting of 20 animals. The dorsal regions of the mice were shaved with an electric clipper. The shaved dorsal skin was then treated topically with a single dose of 0.2 µmol dimethylbenzanthracene (DMBA) dissolved in 0.2 mL of vehicle [acetone:DMSO (85:15, v/v)]. One week after the application of DMBA, the mice were treated topically with 10 nmol of TPA in 0.2 ml of vehicle twice weekly for 20 weeks. DMBA and TPA were used as a tumor initiator and tumor promoter, respectively. To assess the antitumor-promoting activity of EERL, 1 or 10 mg of EERL dissolved in 0.2 ml of vehicle was applied topically 30 min prior to each TPA treatment. Control mice were treated topically with the same volumes of vehicle alone. The incidence and number of tumors that were at least 1 mm in diameter were monitored and counted on a weekly basis. The results were expressed as the percentage of tumor-bearing mice (tumor incidence) and the average number of tumors per mouse (tumor multiplicity). All animal experimental protocols were approved by the Animal Care and Use Committee of Hallym University (Protocol approval #: Hallym2013-23).
Statistical analysis
The data were expressed as the means ± SEM and analyzed via analysis of variance. Differences between the treatment groups were assessed by Duncan's multiple range test or Student's t-test. Differences were considered significant at P < 0.05. All statistical analyses were conducted using the SAS system for Windows V 9.1. (SAS Institute, Cary, NC, USA).
RESULTS
EERL is more potent than EEUL in inhibiting the growth of MLL, DU145, and HT-29 cancer cells.
When DU145 cells were cultured with various concentrations of EERL, the viable cell numbers were decreased in a dose-dependent manner, with a 72 h IC50 = 6 ± 0.1 µg/mL. EEUL also dose-dependently decreased the viable DU145 cell numbers, but the potency of EEUL was approximately half that of EERL, with a 72 h IC50 = 12.3 ± 0.1 µg/mL (Fig. 1A and 1B). However, WERL and WEUL slightly inhibited the growth of DU145 cells only after 72 h (data not shown). Similar results were obtained with MLL prostate cancer cells and HT-29 colon cancer cells (Fig. 1C, 1D, 1E, 1F). The 72 h IC50 values of EERL and EEUL on MLL cell growth were 4.3 ± 0.1 µg/mL and 12 ± 0.1 µg/mL, respectively, and on HT-29 cell growth, were 12.2 ± 0.1 µg/mL and 31 ± 0.4 µg/mL, respectively.
EERL inhibits the growth of several cancer cells but slightly increases the growth of normal cells
We next determined whether EERL reduces the growth of MCF-7 and MDA-MB-231 breast cancer cells, B16-F10 melanoma cells, and A375 and A2058 skin cancer cells. EERL induced a marked reduction in viable cell numbers in a dose-dependent manner, with 88% (MCF-7), 82% (MDA-MB-231), 80% (B16-F10), 94% (A375) and 93% (A2058) decreases 48 h after the addition of 30 µg/mL of EERL (Table 1). However, EERL slightly (~14%) increased the growth of ICE-6 intestinal epithelial cells and CCD1108SK human fibroblasts (Table 1).
EERL induces apoptosis of DU145 prostate cancer cells
To test whether the decrease in cell viability observed after treatment with EERL was attributable to the induction of apoptosis, we estimated apoptotic cell numbers using annexin V staining followed by flow cytometry. Apoptotic DU145 cell numbers were increased in an EERL dose-dependent manner (Fig. 2A). Additionally, to determine whether EERL regulates cell cycle progression, DU145 cells were treated with various concentrations of EERL, and DNA was stained with propidium iodide. EERL treatment did not alter cell cycle progression in DU145 cells (Fig. 2B). Treatment of DU145 cells with various concentrations of EERL markedly increased the levels of cleaved caspase-8, -9, -7 and -3. Additionally, EERL significantly increased the levels of cleaved PARP in a dose-dependent manner (Fig. 2C). EERL treatment increased the levels of cytochrome c in the cytoplasm, and caused a concomitant reduction of this protein in the mitochondria of DU145 cells (Fig. 2D). Consistent with these results the permeability of mitochondrial membranes was significantly increased in EERL-treated cells. Results from JC-1 staining revealed that EERL treatment reduced the proportion of cells with intact mitochondrial membranes (red-positive and green-negative) and increased the proportion of cells with depolarized mitochondrial membranes (green-positive and red-negative) (Fig. 2E). EERL at 15 µg/mL significantly increased the protein levels of pro-apoptotic Bax, but did not affect those of pro-apoptotic Bad or the anti-apoptotic Bcl-2 and Bcl-xL (Fig. 2E).
Fig. 2.

EERL induces apoptosis in DU145 cells. Serum-deprived cells were treated with 0, 5, 10 or 15 µg/mL EERL for 24 h. (A) Cells were trypsinized, loaded with 7-aminoactinomycin D and Annexin V, and then analyzed by flow cytometry. The number of living cells and apoptotic cells is expressed as a percentage of total cell number. (B) Cells were stained with propidium iodide, and the cell cycle was analyzed via flow cytometry. Each bar represents the mean ± SEM (n = 6). (C,D,F) Total cell lysates (C,F) and cytosolic and mitochondrial fractions (D) were prepared and analyzed by Western blotting with the indicated antibodies. Photographs of the chemiluminescent detection of the blots, which were representative of three independent experiments, are shown. The relative abundance of each band to their own b-actin, α-tubulin, or heat shock protein (HSP) 60 was quantified, and the control levels were set at 1. The adjusted mean ± SEM (n = 3) of each band is shown above each blot. (E) Cells were loaded with JC-1 and then analyzed by flow cytometry. The number of cells with normally polarized mitochondrial membranes (red) and cells with depolarized mitochondrial membranes (green) was expressed as a percentage of total cell number. Each bar represents the mean ± SEM (n = 6). Means without a common letter differ, P < 0.05.
The active compound of EERL is licochalcone A.
Hwang et al. [15] previously reported that the levels of non-polar components were significantly increased in EERL, as compared to those in EEUL. Thus, EERL was dissolved in MC, and the MC-soluble fraction was separated from residues (Fig. 3A). The ability of the MC fraction to inhibit DU145 cell growth was examined by MTT assay. The MC fraction decreased the viable DU145 cell numbers in a dose-dependent manner at concentrations between 1 and 3 µg/mL (Fig. 3B). In order to identify the active component(s), the MC fraction was applied to a SiO2 column, and 21 fractions were collected using the MPLC. Among the 21 fractions, fraction #5 was determined to be the most effective in reducing the numbers of viable DU145 cells (Fig. 3C). Compound 1 was obtained from further purification of fraction #5 via recrystallization, and the 1H NMR and 13C NMR spectra of compound 1 were completely matched to the literature value of licochalcone A (LCA) [19]. The structure is shown in Fig. 4A.
Fig. 3.

Non-polar components of EERL inhibit the growth of DU145 cells. (A) Fractionation plan for the identification of the active compound in EERL. (B) Cells were treated with 0, 1, 2 or 3 mg/mL of the MC fraction. The viable cell numbers were determined by MTT assay and represent means ± SEM (n = 6). Means without a common letter differ, P < 0.05. (C) Cells were treated with 3 mg/mL of the individual MPLC fractions for 48 h. Cell numbers were estimated by the MTT assay. Each bar represents the mean ± SEM (n = 6). *Significant difference from the MC fraction, P < 0.01.
Fig. 4.

The active compound of EERL licochalcone A induces apoptosis in DU145 cells. (A) Structure of Licochalcone A. (B) Serum-deprived cells were treated with 0-8 mmol/L licochalcone A, and cell numbers were estimated by MTT assay. Each bar represents the mean ± SEM (n = 6). (C) Serum-deprived cells were treated with licochalcone A for 24 h. Cells were stained with 7-aminoactinomycin D and Annexin V, and were then analyzed by flow cytometry. The number of living cells and apoptotic cells is expressed as a percentage of total cell number. Each bar represents the mean ± SEM (n = 6). (D) Cell lysates were analyzed by Western blotting with the indicated antibodies. Photographs of the chemiluminescent detection of the blots, which were representative of three independent experiments, are shown. The relative abundance of each band to their own β-actin was quantified and the control levels were set at 1. The adjusted mean ± SEM (n = 3) of each band is shown above each blot. (E) Cells were treated with or without 8 µmol/L licochalcone A for 24 h and stained with propidium iodide, and the cell cycle was analyzed via flow cytometry. Each bar represents the mean ± SEM (n = 6). Means with different letters differ significantly, P < 0.05.
The active compound licochalcone A induces apoptosis of DU145 cells
Licochalcone A induced dose-dependent reductions in the number of viable DU145 cells at concentrations of 2-8 µmol/L (Fig. 4B), and induced apoptotic cell death (Fig. 4C). Additionally, licochalcone A significantly increased the levels of cleaved caspase-8, -9, -7 and -3 as well as those of cleaved PARP (Fig. 4D). However, licochalcone A did not alter cell cycle progression of DU145 cells (Fig. 4E).
EERL suppresses DMBA-initiated, TPA-promoted skin tumorigenesis in mice
To determine whether EERL inhibits tumor promotion in vivo, the DMBA-initiated, TPA-promoted skin carcinogenesis mouse model was employed. When EERL (1 or 10 mg) was topically applied to mouse skin prior to treatment of the tumor promoter TPA, tumor incidence (Fig. 5B) and tumor multiplicity (Fig. 5A, C) were drastically decreased, as compared with the vehicle-treated group. EERL treatment with 1 and 10 mg/mice decreased tumor multiplicity by 22.5 and 88%, respectively.
Fig. 5.

EERL inhibits skin papilloma formation. Tumor formation in mice was initiated with the topical application of DMBA (0.2 µmol/mouse) and promoted with TPA (10 nmol/mouse) twice weekly, starting one week after the initiation. The control group received vehicle only. EERL was applied topically 30 min prior to each TPA treatment. (A) Photographs of mice skin. (B) Percentage of mice with papillomas. (C) Average number of papillomas/mouse. Means without a common letter differ, P < 0.05.
DISCUSSION
Extensive use of licorice as a cancer chemopreventive agent is not considered to be desirable despite its excellent antitumor activity [20] because glycyrrhizin, present in licorice, causes hypokalemia by inhibiting the renal enzyme 11-beta-hydorxysteroid dehydrogenase, which is responsible for renal conversion of cortisol to locally inactive cortisone. [21,22] In order to modify the composition of the extracts and to exclude toxic fractions, roasting as well as the use of different solvents has been employed. [13,23] The ethanol extracts of roasted licorice have been reported to exert potent anti-inflammatory properties, [24] neuro-protective effects against ischemic damage, [13] and protective effects against mesangial fibrosis and glomerulosclerosis in diabetes nephropathy. [25] The present results clearly demonstrated that EERL is more potent and efficacious than EEUL in inhibiting the growth of prostate and colon cancer cells (Fig. 1). The aqueous extract of either roasted or un-roasted licorice did not inhibit cancer cell growth. Additionally, EERL effectively inhibits the growth of human skin cancer cells and mouse melanoma cells, as well as human breast cancer cells. However, EERL stimulated the growth of normal rat intestinal epithelial cells and normal fibroblasts (Table 1). Furthermore, the topical application of EERL effectively suppressed chemically induced skin carcinogenesis in mice (Fig. 5). These results indicate that roasted rather than un-roasted licorice can be used to develop an additional supplementary and single agent for the prevention and/or treatment of cancer.
Kim et al. reported that the oral administration of 2000 mg/kg licochalcone A had no toxic effect. [26] However, we did not determine the bioavailability of orally administered EERL and licochalcone A in mice. Our results showed that licochalcone A inhibits the growth and induces apoptosis at the concentration range of 2-8 µmol/L in DU145 cells. It is not known whether the concentrations of licochalcone A used in cell culture studies can be achievable in the blood of animals. 1-10 mg of EERL (from 15.4-153.8 mg roasted licorice) which was used in this study contained 39.5-395.3 µg of licochalcone A. Kim et al. also showed that oral administration of 15-30 mg/kg licochalcone A (5.8-11.7 g/kg roasted licorice) exerted antitumor effect in a colitis-associated colon cancer model. [28] However, orally administered 10 mg/kg EERL was effective in inhibiting the progression and severity of collagen-induced arthritis [15] and orally administered 2 mg/kg EERL was effective in inhibiting the breast cancer cell-induced bone destruction. [42] These previous results suggest that EERL contains other biologically active compounds or licochalcone A in EERL is biologically more active. Future studies are needed to compare the relative potency and efficacy of EERL and licochalcone A for anti-carcinogenic effects in both animals and humans. Together, these results suggest that the consumption of roasted licorice (0.15-11.7 g/kg) can be beneficial to prevent inflammation and cancer.
Hwang et al. previously reported that the levels of non-polar components were significantly increased in EERL as compared to EEUL. [15] In the present study, we noted that the aqueous extract of either roasted or un-roasted licorice did not substantially inhibit cancer cell growth. In an attempt to prepare a fraction highly enriched with non-polar components, EERL was extracted with MC. Because the MC fraction was approximately 5 times more effective than EERL in inhibiting DU145 cell growth (Fig. 1A, Fig. 3B), the MC fraction was further separated into 21 fractions by MPLC. MPLC fraction #5 was the most effective in inhibiting DU145 cell growth, and the principal component of the MPLC #5 was licochalcone A. The final yield of licochalcone A was 257 mg/100 g of dried licorice. These results are consistent with our previous results that licochalcone A inhibits solid tumor growth, [26,27] angiogenesis, [26] and metastasis [28] in mouse tumor models. Future studies are needed to identify other non-polar compounds in roasted licorice that can enhance the anti-carcinogenic effects of licochalcone A.
Several reports showed that licorice and its active compounds inhibit cancer cell growth through the induction of apoptosis, [29,30] cell cycle arrest [29,31,32] and autophagy. [20] Licochalcone A was reported to induce apoptosis in androgen-sensitive LNCaP prostate cancer cells [33] and to induce cell cycle arrests in androgen-insensitive PC-3 prostate cancer cells. [34] In the present study EERL and licochalcone A were observed to induce apoptosis in androgen-insensitive DU145 cells (Fig. 2A, 2B, Fig. 4C and 4E) indicating that the induction of apoptosis is one of the mechanisms by which EERL and licochalcone A exert anti-carcinogenic effects. Our results showed that EERL and licochalcone A did not induce cell cycle arrest of DU145 cells. This was probably due to using the different concentrations of licochalcone A and/or different cell lines.
The two major pathways that trigger apoptosis are the extrinsic pathway, initiated by death receptors on the plasma membrane, and the intrinsic pathway, which occurs through the mitochondria. The mitochondrial pathway is associated with changes in the permeability of the outer mitochondrial membrane, the collapse of membrane potential, and the release of cytochrome c from the mitochondrial intermembrane space into the cytoplasm. [35] Cytochrome c in the cytoplasm activates caspase-9, which in turn activates executioner caspases, such as caspase-3 and caspase-7 [36,37] which plays pivotal roles in the execution of apoptosis by cleaving a wide variety of cellular proteins. [38] The best characterized caspase substrate is PARP, a nuclear protein associated with DNA repair. [4,39] In DU145 cells, EERL treatment induced a marked increase in mitochondrial membrane permeability (Fig. 2E), the release of cytochrome c from mitochondria (Fig. 2D), and the increase in the levels of activated caspase-9, -7, and -3, as well as cleaved PARP (Fig. 2C). Licochalcone A also induced increases in the levels of these cleaved caspases and cleaved PARP (Fig. 4D). Members of the Bcl-2 family include both pro- as well as anti-apoptotic molecules and are the key regulators of mitochondrial membrane permeability. [40,41] EERL treatment also increased the levels of Bax (Fig. 2F), a pro-apoptotic protein, whereas EERL did not change the protein levels of other Bcl-2 family members. These results indicate that EERL and licochalcone A induce the activation of the mitochondrial apoptotic pathway. However, the levels of cleaved capase-8 were also increased in DU145 cells by the treatment of EERL (Fig. 2C) and licochalcone A (Fig. 4D). The extrinsic (death-receptor mediated) pathway is initiated by binding of the ligand with its death receptor, which subsequently activates casapse-8 and -3 (reviewed in [4]). Together, these results indicate that a death receptor pathway was also activated by EERL and licochalcone A.
In summary, we have shown that EERL is more efficacious and potent than EEUL in inhibiting the growth of several tumor cells. EERL also inhibits DMBA-initiated, TPA-promoted skin carcinogenesis in mice. Licochalcone A has been identified as one of the active compounds of EERL. Both EERL and licochalcone A induces the apoptosis of androgen-insensitive DU145 human prostate cancer cells. The present results provide the molecular basis for using EERL and licochalcone A to develop an additional supplementary and single agent for the prevention and/or treatment of cancer.
Footnotes
This work was supported by National Research Foundation of Korea (NRF) grants, as funded by the Korean government (MSIP) (2013R1A2A2A05004533, 2008-0062618, 2011-0028637, 2012R1A1A3010664).
Supplementary Material
The structure of compound 1
References
- 1.Ouédraogo M, Charles C, Ouédraogo M, Guissou IP, Stévigny C, Duez P. An overview of cancer chemopreventive potential and safety of proanthocyanidins. Nutr Cancer. 2011;63:1163–1173. doi: 10.1080/01635581.2011.607549. [DOI] [PubMed] [Google Scholar]
- 2.Kinghorn AD, Chai HB, Sung CK, Keller WJ. The classical drug discovery approach to defining bioactive constituents of botanicals. Fitoterapia. 2011;82:71–79. doi: 10.1016/j.fitote.2010.08.015. [DOI] [PubMed] [Google Scholar]
- 3.Tan AC, Konczak I, Sze DM, Ramzan I. Molecular pathways for cancer chemoprevention by dietary phytochemicals. Nutr Cancer. 2011;63:495–505. doi: 10.1080/01635581.2011.538953. [DOI] [PubMed] [Google Scholar]
- 4.Vermeulen K, Van Bockstaele DR, Berneman ZN. Apoptosis: mechanisms and relevance in cancer. Ann Hematol. 2005;84:627–639. doi: 10.1007/s00277-005-1065-x. [DOI] [PubMed] [Google Scholar]
- 5.Wang ZY, Nixon DW. Licorice and cancer. Nutr Cancer. 2001;39:1–11. doi: 10.1207/S15327914nc391_1. [DOI] [PubMed] [Google Scholar]
- 6.Asl MN, Hosseinzadeh H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother Res. 2008;22:709–724. doi: 10.1002/ptr.2362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yasue H, Itoh T, Mizuno Y, Harada E. Severe hypokalemia, rhabdomyolysis, muscle paralysis, and respiratory impairment in a hypertensive patient taking herbal medicines containing licorice. Intern Med. 2007;46:575–578. doi: 10.2169/internalmedicine.46.6316. [DOI] [PubMed] [Google Scholar]
- 8.van Uum SH. Liquorice and hypertension. Neth J Med. 2005;63:119–120. [PubMed] [Google Scholar]
- 9.Leitolf H, Dixit KC, Higham CE, Brabant G. Licorice - or more? Exp Clin Endocrinol Diabetes. 2010;118:250–253. doi: 10.1055/s-0029-1224123. [DOI] [PubMed] [Google Scholar]
- 10.Omar HR, Komarova I, El-Ghonemi M, Fathy A, Rashad R, Abdelmalak HD, Yerramadha MR, Ali Y, Helal E, Camporesi EM. Licorice abuse: time to send a warning message. Ther Adv Endocrinol Metab. 2012;3:125–138. doi: 10.1177/2042018812454322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Murphy SC, Agger S, Rainey PM. Too much of a good thing: a woman with hypertension and hypokalemia. Clin Chem. 2009;55:2093–2096. doi: 10.1373/clinchem.2009.127506. [DOI] [PubMed] [Google Scholar]
- 12.Sung MW, Li PC. Chemical analysis of raw, dry-roasted, and honey-roasted licorice by capillary electrophoresis. Electrophoresis. 2004;25:3434–3440. doi: 10.1002/elps.200305988. [DOI] [PubMed] [Google Scholar]
- 13.Hwang IK, Lim SS, Choi KH, Yoo KY, Shin HK, Kim EJ, Yoon-Park JH, Kang TC, Kim YS, Kwon DY, Kim DW, Moon WK, Won MH. Neuroprotective effects of roasted licorice, not raw form, on neuronal injury in gerbil hippocampus after transient forebrain ischemia. Acta Pharmacol Sin. 2006;27:959–965. doi: 10.1111/j.1745-7254.2006.00346.x. [DOI] [PubMed] [Google Scholar]
- 14.Majima T, Yamada T, Tega E, Sakurai H, Saiki I, Tani T. Pharmaceutical evaluation of liquorice before and after roasting in mice. J Pharm Pharmacol. 2004;56:589–595. doi: 10.1211/0022357023286. [DOI] [PubMed] [Google Scholar]
- 15.Kim KR, Jeong CK, Park KK, Choi JH, Park JH, Lim SS, Chung WY. Anti-inflammatory effects of licorice and roasted licorice extracts on TPA-induced acute inflammation and collagen-induced arthritis in mice. J Biomed Biotechnol. 2010;2010:709378. doi: 10.1155/2010/709378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Jung JI, Lim SS, Choi HJ, Cho HJ, Shin HK, Kim EJ, Chung WY, Park KK, Park JH. Isoliquiritigenin induces apoptosis by depolarizing mitochondrial membranes in prostate cancer cells. J Nutr Biochem. 2006;17:689–696. doi: 10.1016/j.jnutbio.2005.11.006. [DOI] [PubMed] [Google Scholar]
- 17.Cho HJ, Kim WK, Kim EJ, Jung KC, Park S, Lee HS, Tyner AL, Park JH. Conjugated linoleic acid inhibits cell proliferation and ErbB3 signaling in HT-29 human colon cell line. Am J Physiol Gastrointest Liver Physiol. 2003;284:G996–G1005. doi: 10.1152/ajpgi.00347.2002. [DOI] [PubMed] [Google Scholar]
- 18.Eguchi Y, Srinivasan A, Tomaselli KJ, Shimizu S, Tsujimoto Y. ATP-dependent steps in apoptotic signal transduction. Cancer Res. 1999;59:2174–2181. [PubMed] [Google Scholar]
- 19.Wang B, Zou K, Yang XB, He WY, Zhao YY, Zhang RY. Two new flavanone glycosides from Glycyrrhizia inflata. Yao Xue Xue Bao. 1997;32:199–202. [PubMed] [Google Scholar]
- 20.Yo YT, Shieh GS, Hsu KF, Wu CL, Shiau AL. Licorice and licochalcone-a induce autophagy in LNCaP prostate cancer cells by suppression of Bcl-2 expression and the mTOR pathway. J Agric Food Chem. 2009;57:8266–8273. doi: 10.1021/jf901054c. [DOI] [PubMed] [Google Scholar]
- 21.Mumoli N, Cei M. Licorice-induced hypokalemia. Int J Cardiol. 2008;124:e42–e44. doi: 10.1016/j.ijcard.2006.11.190. [DOI] [PubMed] [Google Scholar]
- 22.Sontia B, Mooney J, Gaudet L, Touyz RM. Pseudohyperaldosteronism, liquorice, and hypertension. J Clin Hypertens (Greenwich) 2008;10:153–157. doi: 10.1111/j.1751-7176.2008.07470.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Choi HJ, Seon MR, Lim SS, Kim JS, Chun HS, Park JH. Hexane/ethanol extract of Glycyrrhiza uralensis licorice suppresses doxorubicin-induced apoptosis in H9c2 rat cardiac myoblasts. Exp Biol Med (Maywood) 2008;233:1554–1560. doi: 10.3181/0807-RM-221. [DOI] [PubMed] [Google Scholar]
- 24.Kim JK, Oh SM, Kwon HS, Oh YS, Lim SS, Shin HK. Anti-inflammatory effect of roasted licorice extracts on lipopolysaccharide-induced inflammatory responses in murine macrophages. Biochem Biophys Res Commun. 2006;345:1215–1223. doi: 10.1016/j.bbrc.2006.05.035. [DOI] [PubMed] [Google Scholar]
- 25.Li J, Lee YS, Choi JS, Sung HY, Kim JK, Lim SS, Kang YH. Roasted licorice extracts dampen high glucose-induced mesangial hyperplasia and matrix deposition through blocking Akt activation and TGF-beta signaling. Phytomedicine. 2010;17:800–810. doi: 10.1016/j.phymed.2010.02.004. [DOI] [PubMed] [Google Scholar]
- 26.Kim YH, Shin EK, Kim DH, Lee HH, Park JH, Kim JK. Antiangiogenic effect of licochalcone A. Biochem Pharmacol. 2010;80:1152–1159. doi: 10.1016/j.bcp.2010.07.006. [DOI] [PubMed] [Google Scholar]
- 27.Lee CK, Son SH, Park KK, Park JH, Lim SS, Kim SH, Chung WY. Licochalcone A inhibits the growth of colon carcinoma and attenuates cisplatin-induced toxicity without a loss of chemotherapeutic efficacy in mice. Basic Clin Pharmacol Toxicol. 2008;103:48–54. doi: 10.1111/j.1742-7843.2008.00238.x. [DOI] [PubMed] [Google Scholar]
- 28.Kim JK, Shin EK, Park JH, Kim YH, Park JH. Antitumor and antimetastatic effects of licochalcone A in mouse models. J Mol Med (Berl) 2010;88:829–838. doi: 10.1007/s00109-010-0625-2. [DOI] [PubMed] [Google Scholar]
- 29.Jo EH, Kim SH, Ra JC, Kim SR, Cho SD, Jung JW, Yang SR, Park JS, Hwang JW, Aruoma OI, Kim TY, Lee YS, Kang KS. Chemopreventive properties of the ethanol extract of chinese licorice (Glycyrrhiza uralensis) root: induction of apoptosis and G1 cell cycle arrest in MCF-7 human breast cancer cells. Cancer Lett. 2005;230:239–247. doi: 10.1016/j.canlet.2004.12.038. [DOI] [PubMed] [Google Scholar]
- 30.Jo EH, Hong HD, Ahn NC, Jung JW, Yang SR, Park JS, Kim SH, Lee YS, Kang KS. Modulations of the Bcl-2/Bax family were involved in the chemopreventive effects of licorice root (Glycyrrhiza uralensis Fisch) in MCF-7 human breast cancer cell. J Agric Food Chem. 2004;52:1715–1719. doi: 10.1021/jf035012t. [DOI] [PubMed] [Google Scholar]
- 31.Seon MR, Park SY, Kwon SJ, Lim SS, Choi HJ, Park H, Lim do Y, Kim JS, Lee CH, Kim J, Park JH. Hexane/ethanol extract of Glycyrrhiza uralensis and its active compound isoangustone A induce G1 cycle arrest in DU145 human prostate and 4T1 murine mammary cancer cells. J Nutr Biochem. 2012;23:85–92. doi: 10.1016/j.jnutbio.2010.11.010. [DOI] [PubMed] [Google Scholar]
- 32.Rafi MM, Vastano BC, Zhu N, Ho CT, Ghai G, Rosen RT, Gallo MA, DiPaola RS. Novel polyphenol molecule isolated from licorice root (Glycrrhiza glabra) induces apoptosis, G2/M cell cycle arrest, and Bcl-2 phosphorylation in tumor cell lines. J Agric Food Chem. 2002;50:677–684. doi: 10.1021/jf010774e. [DOI] [PubMed] [Google Scholar]
- 33.Szliszka E, Czuba ZP, Mazur B, Sedek L, Paradysz A, Krol W. Chalcones enhance TRAIL-induced apoptosis in prostate cancer cells. Int J Mol Sci. 2009;11:1–13. doi: 10.3390/ijms11010001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fu Y, Hsieh TC, Guo J, Kunicki J, Lee MY, Darzynkiewicz Z, Wu JM. Licochalcone-A, a novel flavonoid isolated from licorice root (Glycyrrhiza glabra), causes G2 and late-G1 arrests in androgen-independent PC-3 prostate cancer cells. Biochem Biophys Res Commun. 2004;322:263–270. doi: 10.1016/j.bbrc.2004.07.094. [DOI] [PubMed] [Google Scholar]
- 35.Martinou JC, Green DR. Breaking the mitochondrial barrier. Nat Rev Mol Cell Biol. 2001;2:63–67. doi: 10.1038/35048069. [DOI] [PubMed] [Google Scholar]
- 36.Green DR, Reed JC. Mitochondria and apoptosis. Science. 1998;281:1309–1312. doi: 10.1126/science.281.5381.1309. [DOI] [PubMed] [Google Scholar]
- 37.Budihardjo I, Oliver H, Lutter M, Luo X, Wang X. Biochemical pathways of caspase activation during apoptosis. Annu Rev Cell Dev Biol. 1999;15:269–290. doi: 10.1146/annurev.cellbio.15.1.269. [DOI] [PubMed] [Google Scholar]
- 38.Plati J, Bucur O, Khosravi-Far R. Apoptotic cell signaling in cancer progression and therapy. Integr Biol (Camb) 2011;3:279–296. doi: 10.1039/c0ib00144a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ho PK, Hawkins CJ. Mammalian initiator apoptotic caspases. FEBS J. 2005;272:5436–5453. doi: 10.1111/j.1742-4658.2005.04966.x. [DOI] [PubMed] [Google Scholar]
- 40.Gross A, McDonnell JM, Korsmeyer SJ. BCL-2 family members and the mitochondria in apoptosis. Genes Dev. 1999;13:1899–1911. doi: 10.1101/gad.13.15.1899. [DOI] [PubMed] [Google Scholar]
- 41.Wong RS. Apoptosis in cancer: from pathogenesis to treatment. J Exp Clin Cancer Res. 2011;30:87. doi: 10.1186/1756-9966-30-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lee SK, Park KK, Park JH, Lim SS, Chung WY. The inhibitory effect of roasted licorice extract on human metastatic breast cancer cell-induced bone destruction. Phytother Res. 2013;27:1776–1783. doi: 10.1002/ptr.4930. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The structure of compound 1