

Abstract
McCarty and colleagues (1999) developed the elevated spoon task to measure motor planning in human infants. In this task, a spoon containing food was placed on an elevated apparatus that supported both ends of the spoon. The handle was oriented to the left or right on different trials. We presented naïve adult rhesus monkeys (Macaca mulatta) with the elevated spoon problem, and observed how monkeys learned the affordances of spoons over sessions. Strikingly, monkeys developed two different strategies for efficient spoon transport in just 12 to 36 trials. In subsequent testing with a novel double bowl spoon approximately 1 year later, monkeys demonstrated that they were attending to the baited spoon bowl and continued to select efficient grips for transporting the spoon. Monkey data were contrasted with previous studies in human infants using a perception-action perspective in an effort to understand the fundamentals of tool use and motor planning that may be common in the development of these abilities across species and their origins in human behavior.
INTRODUCTION
Motor planning can be studied by examining goal-directed actions. In order to effectively solve a motor problem, an individual must first perceive the goal of the task and reason about possible solutions. After selecting an action, the individual must then determine whether the goal was achieved, or if another action should be tried. Thus, motor planning requires cognitive ability in selecting actions and acknowledging errors as well as motor ability in executing actions, even though it is often studied in a purely motor context (Rosenbaum, 2005).
Recently, attention has been focused on characterizing the origins of motor planning by examining the ontogeny of planning ability in human infants (McCarty, Clifton & Collard; 1999; McCarty & Clifton, 2001; Claxton, Keen & McCarty, 2003; McCarty & Keen, 2005; Claxton, McCarty & Keen, 2009). In a task designed by McCarty et al. (1999), a spoon baited with food was placed on elevated bookends and presented to infants of various ages. The orientation of the spoon varied by trial creating two distinct trial conditions: easy trials where the handle of the spoon matched the infant’s preferred hand, and difficult trials where the handle of the spoon was matched to the infant’s non-preferred hand. On every trial, the goal of the task was to obtain the food on the spoon. Because poor planning had a negative consequence (e.g., food could slide off the spoon), this task was a good indicator of planning ability.
Infants’ performance on the spoon task varied as a function of age. Nine-month-old infants persisted in using their preferred hand regardless of the spoon’s orientation and frequently put the handle end of the spoon into the mouth on difficult trials. After making this error, infants often corrected their mistake and put the bowl of the spoon in the mouth. Fourteen-month-old infants also largely used their preferred hand, but adjusted their movements on difficult trials so that the bowl of the spoon was put into the mouth first. By 19 months-of-age, infants alternated hands based on the orientation of the spoon and did not need to correct their movements (McCarty et al., 1999).
The ability to select efficient movements on the spoon task may in part be related to the amount of spoon experience infants typically have at the ages examined by McCarty et al. (1999). Parents usually begin feeding their infant from a spoon between 6 and 12 months-of-age. Through this process, infants learn the relationship between the spoon and food, but cannot yet use the spoon effectively. Sometime early in the second year of life, infants begin trying to feed themselves with the spoon, and spoon-using skills develop over the next several months (Connolly & Dalgleish, 1989). By 18 months, infants have consistent grasping preferences for using spoons (Connolly & Dalgleish, 1993). Efficient planning on the elevated spoon task appears to develop between 14 and 19 months, corresponding to the time period when infants establish grip preferences for spoons.
The improvement in the ability to use a spoon as a tool for transporting food observed over the course of infant development suggests that the solution to the elevated spoon problem is not based on insight, but rather increasing knowledge of the affordances of the spoon and the relationships between the components of the spoon, the food, and the infant. Lockman (2000) advocated for a perception-action based model of tool use development that focuses on examining the processes that occur during learning to use tools, rather than a pass or fail outcome on a particular task. The ability to use tools may therefore reflect a gradual, dynamic learning process that is often discounted in tool-using studies, particularly those conducted with nonhuman primates that have examined the evolutionary origins of tool-using abilities.
It is this perception-action process that may share a similar underlying mechanism in both human infants and nonhuman primates, even if the adult patterns of tool use and planning differ in sophistication across species. Visalberghi and Fragaszy (2006) adopted this perspective in describing tool use in capuchin monkeys (Cebus apella) and noted that very little is actually known about how nonhuman primates learn how objects and object parts are related. Examining the development of movement strategies in both human and nonhuman subjects will provide insight into the way perception and action are used to explore the affordances of tools that leads to their effective use as well as the origins of tool use and motor planning in human behavior. Nonhuman primate models offer an opportunity to observe the fundamental components of this developmental process with greater control over the general amount of interaction that subjects have with objects in a laboratory setting as compared to human infants.
The underlying assumption in observing tool use development in nonhuman primates is that subjects are motivated by efficiency to refine their movements and plan appropriate actions. Recent work in both captivity and the wild has suggested that nonhuman primates are indeed motivated by efficiency and select grips based on utility for future actions (Chapman, Weiss & Rosenbaum, 2010; Gumert, Kluck & Malaivijitnond, 2009; Weiss, Wark & Rosenbaum, 2007). Weiss et al. (2007) demonstrated that captive cotton-top tamarins (Saguinus oedipus) choose how to grip a cup stem according to the cup’s orientation and its future possible movement. When the cup was an upright position, monkeys used a thumb-up grip because the cup did not need to be rotated to retrieve a treat that had been placed inside the cup bowl. When the cup was in an inverted position however, monkeys used a thumb-down grip, indicating that they had anticipated the rotation of the cup and chose a grip that would facilitate turning the cup over. Although not as robust, the same gripping pattern was found in five species of captive lemurs (collared lemur, Eulemur collaris; mongoose lemur, Eulemur mongoz; eastern lesser bamboo lemur, Hapalemur griseus; ringtailed lemur, Lemur catta; red ruffed lemur, Varecia rubra) using a similar cup paradigm (Chapman et al., 2010).
In another recent report by Gumert et al. (2009), wild long-tailed macaques (Macaca fascicularis aurea) were observed to select different types of stone tools depending on the food item to be opened. Monkeys used a larger stone called a pounding hammer to open nuts and loose shellfish and a smaller stone called an axe hammer to open oysters attached to rocks. Importantly, monkeys used specific grips with each tool type. A power grip was often used with the pounding hammer to bring the stone down onto the food item from above. In comparison, a precision grip was associated with the axe hammer, which allowed for rapid pounding and greater hand control. If monkeys gripped the pounding hammer poorly, an easy correction was to set the tool down on the pounding surface and adjust the hand(s). With the axe hammer however monkeys were pounding in a vertical space and poor grip selection could cause the monkey to miss the target or even drop the stone. Thus grip choice impacts foraging efficacy, and poor grip selection can have negative consequences. Taken together, these studies suggest that motor planning is an ecologically relevant problem for nonhuman primates, and that nonhuman primates are motivated by efficiency in selecting actions.
In the current study, we presented adult rhesus monkeys (Macaca mulatta) that had no prior spoon experience with the elevated spoon problem and observed the monkeys’ behavior over various sessions (Experiment 1). We were interested in both the initial action the monkeys chose and any mistakes that were made in transporting the spoon. Furthermore, we asked whether mistakes would be corrected and if so, at what point in the movement sequence. We hypothesized that rhesus monkeys would select movements based on efficiency, and predicted that monkeys would alternate hands according to the orientation of the spoon like older infants. We acknowledge that as adults, monkeys have fully developed motor skills compared to human infants. The monkeys may also have an additional cognitive advantage over infants in that these particular subjects have experience participating in problem solving tasks. However, we did not expect the monkeys to instantly solve the task. Rather, we expected to observe an unfolding of ability over time as the monkeys discovered the properties of the spoon, similar to what has been observed in human infants. Therefore, we also predicted that reaches to the spoon (measured as movement time) would decrease over sessions as monkeys became more efficient in transporting spoons. Based on the results of Experiment 1, the same subjects were tested with a modified spoon in Experiment 2 to further examine how monkeys perceived the relationships between spoon components.
EXPERIMENT 1: SINGLE BOWL CONDITION
METHOD
Subjects
Seven adult rhesus monkeys (two males; five females) housed at the University of Massachusetts participated in this study. Monkeys ranged in age from 7 to 23 years old (mean age = 11). All monkeys were born in captivity. Six monkeys were surrogate peer-reared according to the protocol described by Shannon, Champoux, and Suomi (1998). The rearing history of the remaining monkey was unknown. Four monkeys were individually housed in close proximity to other monkeys, and three monkeys were socially housed with one or two other monkeys (Table 1). All monkeys had prior experience participating in cognitive studies but no experience manipulating spoons. Four monkeys had previously participated in tool-using studies, and could proficiently use a rake to retrieve a treat placed out of reach outside of the rake head (Metevier, 2006). The remaining three monkeys had no previous tool-using experience (Table 1). Monkeys had access to food and water throughout the experiment. The study was conducted in accordance with the Guide for the Care and Use of Laboratory Animals and complied with the Animal Welfare Act. The University of Massachusetts Amherst Institutional Animal Care and Use Committee (IACUC) approved the research. Monkeys were tested individually in their home cages on the following tasks.
Table 1.
Number of trials to reach criterion (mastery of efficient grip) on the elevated spoon task for the single bowl condition by subject including the strategy used at criterion, hand preference, sex, age, and previous experience on a rake tool-using task prior to spoon testing
| Subject | Criterion | Strategy | Hand Preference* | Sex | Age | Rake Experience** |
|---|---|---|---|---|---|---|
| Thelmaa | 12 trials | Alternate hands | −1.00 (0.00) | F | 23 | Not tested |
| Ivana | 18 trials | Alternate hands | 0.82 (0.11) | M | 10 | Proficient with rake |
| Zoeyb | 24 trials | Use preferred hand | 0.96 (0.08) | F | 7 | Not tested |
| Kaylab | 24 trials | Use preferred hand | 0.32 (0.58) | F | 7 | Not tested |
| Taza | 30 trials | Use preferred hand | 0.88 (0.17) | F | 10 | Proficient with rake |
| Violetc | 36 trials | Alternate hands | −0.12 (0.32) | F | 10 | Proficient with rake |
| Cobyc | NA | NA | −0.78 (0.18) | M | 10 | Proficient with rake |
Denotes animals housed in the same room.
Calculated by the formula HI=(R−L)/(R+L), where HI = Handedness Index, R=Right hand reaches, L=Left hand reaches. Values range from −1.00 (exclusive left hand use) to 1.00 (exclusive right hand use). Standard deviations given in parentheses.
Data from Metevier (2006).
Reaching Task
Baseline reaching data were collected for each monkey to determine individual hand preferences prior to spoon testing. Monkeys reached from a seated position for a small food item (Gerber® Graduates® Fruit Puffs) that was presented at midline outside their home cages. Monkeys were required to locomote to a new location between trials to prevent rote reaching. Monkeys were given 10 trials per day over 10 days for a total of 100 trials.
Elevated Spoon Task
In the elevated spoon task, monkeys were required to lift a metal spoon measuring 19 cm in length with a bowl diameter of 4 cm from a wooden platform measuring 22.9 cm by 22.9 cm at the base and 29.2 cm high. The spoon rested on bookends that were spaced 11.4 cm apart, allowing the bowl and handle ends of the spoon to be supported while leaving the middle portion accessible (Figure 1A). The spoon was elevated 20.3 cm from the base of the platform. A handle on the back of the platform allowed the apparatus to be secured to the inside of the monkey’s cage. Spoons were baited with either applesauce or yogurt.
Figure 1.
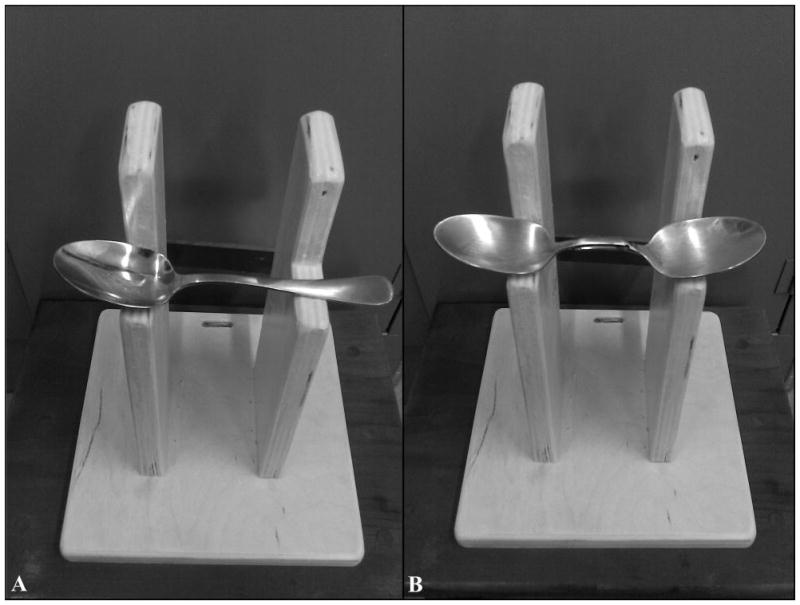
Elevated spoon task. Left panel (A) shows the single bowl condition (Experiment 1). The orientation of the spoon is “easy” for a right-preferent monkey and “difficult” for a left-preferent monkey. Right panel (B) shows the double bowl condition (Experiment 2).
The testing procedure for the spoon task required that the monkey be moved to a separate holding quadrant of their home cage during parts of each testing session. The monkey was first moved to the holding location to allow the spoon apparatus to be attached to the inside of their enclosure. An experimenter then baited a spoon and placed the spoon on the holder with the spoon handle held between the thumb and index finger. Monkeys had an obstructed view of the experimenter during spoon placement due to the dividing panels separating the testing area from the holding location. The orientation of the spoon handle was randomized left or right on different trials. Trials where the spoon handle was oriented to the monkey’s preferred hand were considered easy trials whereas trials where the spoon handle was oriented to the monkey’s non-preferred hand were considered difficult trials. The first trial of any session was randomized easy or difficult based on individual hand preferences as determined from the reaching task previously described, and an equal number of easy and difficult trials were given each session.
A trial began when the monkey was released into the testing quadrant and ended when the monkey finished eating the food, or subsequently spilled the food. Once the trial ended, the monkey was moved to the holding location and the spoon was reset. After the last trial, the monkey was moved to the holding quadrant and the spoon apparatus was removed from the enclosure. Monkeys received 6 trials per session and were tested until mastery of efficient grip use (criterion of 10 or more efficient grips over two consecutive test days) or until they had participated in a maximum of 6 sessions. All spoon trials were videotaped for later analysis.
Analysis
A Handedness Index (HI) was used to characterize hand preferences from the reaching task. Mean HI scores were computed by subtracting the total number of left hand reaches from the total number of right hand reaches and then dividing by the total number of reaches summed across testing days, HI = (R − L)/(R + L). HI scores were also computed for each testing day to determine the variability in hand use preference. HI scores range from −1.00 (exclusively left) to 1.00 (exclusively right) with this index. Negative values indicated a left hand bias and positive values indicated a right hand bias.
Spoon trials were analyzed from video in two ways. In the preliminary analysis, the primary observer noted the orientation of the spoon, whether the trial was easy or difficult for the individual tested, and the monkey’s initial grip on the spoon. Spoon grips were scored as radial (hand on spoon handle with palm facing down and thumb towards the spoon bowl), underhand (hand on spoon handle with palm facing up), ulnar (hand on spoon handle with palm facing down and thumb towards handle end), bowl (any portion of hand gripping spoon bowl), or no grip (monkeys attempted to consume the food by licking or finger dipping but did not manipulate the spoon with either hand). The radial grip and the underhand grip were considered efficient motor strategies because they allowed the spoon to be transported to the mouth in a single movement, and were therefore considered to be examples of good planning. The ulnar and bowl grips were considered inefficient strategies because they required a correction to correctly maneuver the spoon bowl to the mouth, and often resulted in spilling the food. Licking or finger dipping without using a spoon grip was also inefficient because the spoon typically fell off the platform before all the food could be eaten, and was thus considered to be poor planning.
To demonstrate mastery of spoon transport, individual monkeys were required to make 10 or more efficient grips over two consecutive test days to meet a statistically significant criterion (two-tailed binomial probability test, P < 0.05). The primary observer reviewed each session for the subset of monkeys that met this criterion in a frame-by-frame analysis (30 frames per second) using the program MPEG Streamclip (Squared 5 srl). The observer scored reach onset and spoon contact according to the following criteria. Reach onset was the first frame of arm movement towards the spoon apparatus. Spoon contact was the first frame of hand contact with the spoon. Movement time was calculated by subtracting reach onset from spoon contact. A second observer who was blind to condition independently scored approximately 20% of the sessions for inter-rater reliability using percent agreement. For timing measures, a difference was counted if observers differed by more than 5 frames. Reliability was 100% for hand choice, 96% for grip choice, 96% for reach onset, and 88% for spoon contact. Linear regression was used to examine changes in movement time across sessions for each monkey using the statistical computing program R (R Development Core Team, 2009). Data were plotted using R graphics packages (Lemon et al., 2009; Sakar, 2009).
RESULTS
Mean Handedness Index (HI) scores on the reaching task ranged from −1.00 indicating only left hand use to 0.96 indicating mostly right hand use (Table 1). Three monkeys were classified as left-preferent with HI scores of −0.12 (SD=0.32), −0.78 (SD=0.18), and −1.00 (SD=0.00). The remaining four monkeys were classified as right-preferent with mean HI scores of 0.32 (SD=0.58), 0.82 (SD=0.11), 0.88 (SD=0.17), and 0.96 (SD=0.08).
On the elevated spoon task, the majority of the monkeys gripped the spoon, brought the spoon to the mouth, and ate from the bowl. Overall six of the seven monkeys met the criterion of 10 or more efficient grips over 2 consecutive test sessions (Table 1). Individual monkeys met criterion at different points varying from 12 to 36 trials. The number of trials to criterion did not vary as a function of previous tool-using experience. Monkeys that were experienced in using a rake reached criterion in 28 trials (SD=9) on average, whereas monkeys that had no previous experience using a rake averaged 20 trials (SD=7). One monkey reached for the spoon, but did not consistently use a manual grip by the end of the 6 sessions, despite previous successful rake experience (Coby). This monkey’s scores were excluded from further analysis.
Monkeys picked up the spoon on 95% of the total trials (137/144), and use of an efficient grip varied by trial type and session (Table 2). The radial and the underhand grips were considered efficient, whereas the ulnar, bowl, and no grip were considered inefficient. Efficient grips on easy trials increased from 61% in the first session to 100% at criterion for the group (Figure 2). The radial grip was the predominant grip used on easy trials, although two monkeys (Taz and Kayla) used an underhand grip at least once. For the radial grip, the spoon was held horizontally as monkeys ate. In contrast, the spoon was tilted up vertically when an underhand grip was used to facilitate feeding. Efficient grips on difficult trials increased from 28% to 94% for the group (Figure 2). As predicted, some monkeys alternated hands based on the spoon’s orientation, as did older infants (McCarty et al., 1999). These monkeys will be referred to as the alternators. Alternators achieved mastery of efficient grip use in 22 trials (SD=12) on average. Unlike infants however, other monkeys adjusted the position of their body in relationship to the spoon platform and used their preferred hand on every trial (Table 1). This group of monkeys will be referred to as the maintainers. Maintainers reached criterion in 26 trials on average (SD=3). Only radial grips were used on difficult trials regardless of the strategy that monkeys developed (Figure 3).
Table 2.
Grip type by condition and trial type for monkeys as a group
| Easy Trials | Difficult Trials | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Radial | Under | Ulnar | Bowl | No Grip | Radial | Under | Ulnar | Bowl | No Grip | |
| Condition | ||||||||||
| Single Bowla | 11 | 0 | 0 | 3 | 4 | 5 | 0 | 3 | 7 | 3 |
| Single Bowlb | 15 | 3 | 0 | 0 | 0 | 17 | 0 | 0 | 1 | 0 |
| Double Bowl | 10 | 5 | 1 | 2 | 0 | 12 | 0 | 2 | 4 | 0 |
First Session,
Criterion.
Figure 2.
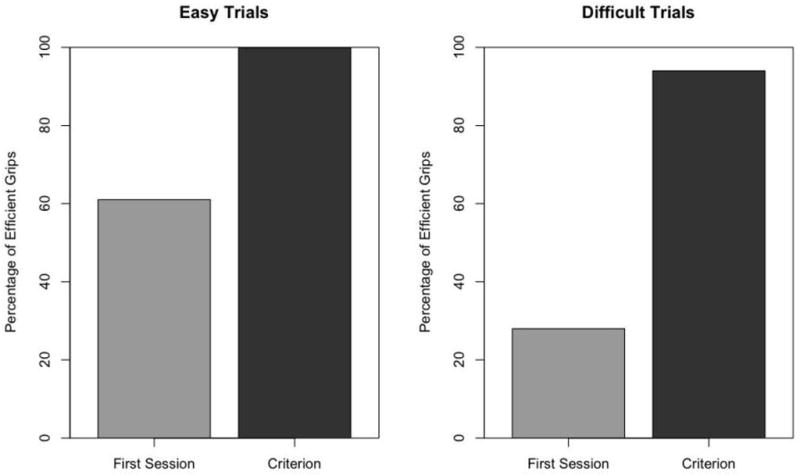
Percentage of efficient grips for single bowl condition at first session compared to criterion session. Efficient grips on easy trials increased from 61% to 100% for the group (left panel). Efficient grips on difficult trials increased from 28% to 94% for the group (right panel).
Figure 3.
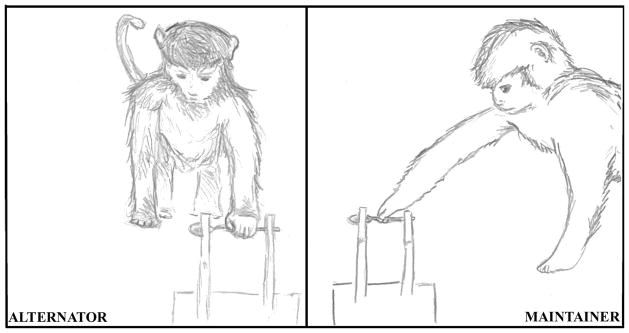
Spoon transport strategies illustrated. The orientation of the spoon indicates a difficult trial for a right-preferent monkey in both panels. Left: Alternators use a radial grip with the non-preferred hand. Right: Maintainers use a radial grip with the preferred hand. Note the differences in body orientation and arm angle.
When gripping errors were made, monkeys adjusted their movements before bringing the spoon to the mouth. When all of the sessions were examined, the majority of errors were bowl grips. Following this error, monkeys added the opposite hand to the spoon handle and then ate the food. Ulnar grips were uncommon, but when such a mistake was made, monkeys attempted to adjust their grip by rotating the spoon. The food was often spilled during this process. Unlike human infants, monkeys did not put the handle of the spoon into the mouth after an ulnar grip.
The effect of session on movement time was explored with linear regression analyses that examined all of the sessions in which individual monkeys participated, varying from 2 to 6 based on time to criterion (Figure 4). Outliers were identified from boxplots as values 2 times the interquartile range and were removed (movement time values > 700 ms). A significant decrease in movement time over sessions was noted for the three monkeys that developed the alternating hand strategy (Thelma: F(1,9)=12.100, P<0.01, R2=0.573; Ivan: F(1,14)=7.539, P < 0.05, R2=0.350; Violet: F(1,29)=8.714, P<0.01, R2=0.231) but not for the three monkeys who continued to use their preferred hand by changing their body position (Kayla: F(1,20)=3.221, P>0.05, R2=0.139; Zoey: F(1,21)=0.856, P>0.05, R2=0.039; Taz: F(1,25)=0.006, P>0.05, R2<0.001). The decreasing pattern observed in alternators was explored in greater detail by examining the performance of monkeys’ preferred and non-preferred hands separately. In two of the three monkeys, the decrease in movement time was found to be a result of the non-preferred hand becoming faster, while the preferred hand remained constant (Thelma and Violet). In the third monkey (Ivan), neither the preferred hand nor the non-preferred hand showed significant change over sessions.
Figure 4.
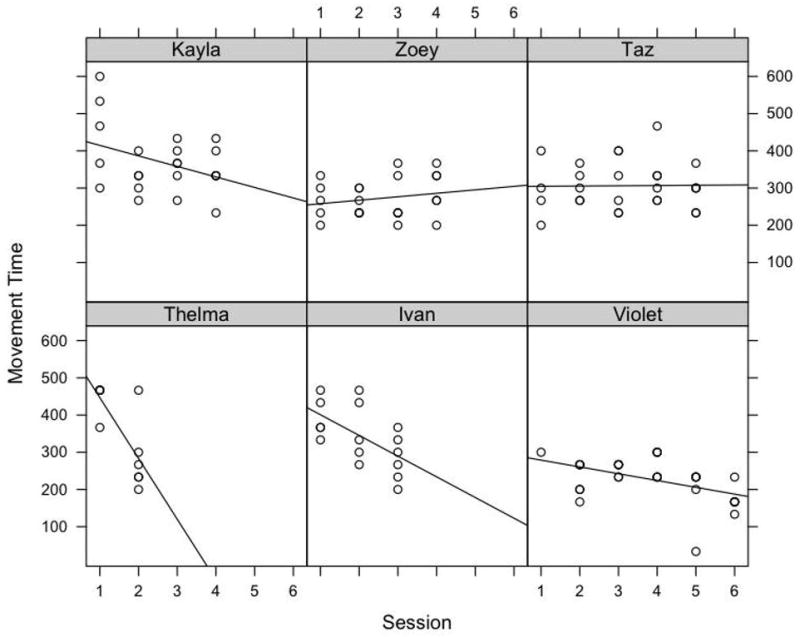
Movement time by session for individual monkeys for the single bowl condition (Experiment 1). Linear regression analyses revealed movement time significantly decreased for the three subjects that developed the alternating hand strategy (Thelma, Ivan, and Violet; P < 0.05), but remained constant for the three subjects who adjusted their body position and continued to use their preferred hand (Kayla, Zoey, and Taz; P > 0.05).
DISCUSSION
The majority of monkeys selected efficient grips to transport the spoon to the mouth and obtain a food reward. Interestingly, monkeys developed two different transport strategies (Table 1). As expected, some monkeys alternated hands to pick up the spoon (n = 3). Similar to the 19-month-old infants studied by McCarty et al. (1999), alternators used the orientation of the spoon to guide their hand choice. The preferred hand was used on easy trials, and the non-preferred hand was used on difficult trials. In every case, monkeys faced the platform directly, and the handle of the spoon was in line with the width of the palm. Other monkeys unexpectedly executed an efficient grip with their preferred hand on both easy and difficult trials (n = 3). To achieve a radial grip with the preferred hand on a difficult trial, the monkeys changed how their body was oriented to the platform as well as how their hand was positioned with respect to the spoon. Approaching from an oblique angle enabled monkeys to grasp the spoon with the handle oriented lengthwise to the underside of the hand (Figure 3). In contrast, 9-month-old and 14-month-old infants that used the preferred hand on difficult trials employed bowl or ulnar grips that required a correction to successfully obtain the food (McCarty et al., 1999). However, infants were constrained to a high chair or booster seat during testing and therefore did not have the option of reorienting their body to the apparatus.
The grip strategy monkeys used on the spoon task affected latency to reach to the spoon. Movement time did not vary over sessions in the maintainer group. In contrast, all three monkeys in the alternator group showed significant decreases in movement time over sessions. Additional analyses revealed that the observed decrease in movement time was a result of the non-preferred hand improving in the majority of the alternator monkeys. Movement time for trials where the preferred hand was used did not change over sessions in any of the monkeys regardless of their spoon transport strategy. These data suggest that monkeys may have been performing at floor levels from the start when the preferred hand was used to reach to the spoon.
It is striking that monkeys learned efficient use of the single bowl spoon in just 12 to 36 trials. Similar to the observations by Kellogg and Kellogg (1933) of a chimpanzee infant learning to use a spoon proficiently months before her human counterpart, rhesus monkeys appear to gain knowledge of some spoon properties faster than human infants. With no prior experience, the majority of monkeys developed an efficient grip strategy very quickly with minimal exposure to spoons. In comparison, infants are slowly introduced to spoons and accumulate months of spoon experience before their performance on the elevated spoon task is at the level of the spoon-naïve monkeys (McCarty et al., 1999).
The question remains as to how the monkeys learned to grip the spoon efficiently. Previous tool use experience of maneuvering a rake to retrieve an object placed out of reach did not appear to influence monkeys’ performance on the elevated spoon task. Three monkeys that had never been tested on the rake task transported spoons efficiently, and one monkey that was a rake user did not reliably manipulate the spoons. Imitation of the experimenter is also an unlikely explanation, as monkeys had an obstructed view of the experimenter baiting the apparatus between trials. In addition, no monkey ever used the thumb to index finger precision grip that the experimenter used when placing the spoon on the testing platform. Rather, we propose that monkeys progressed through various stages in learning to plan efficient movements just as a human infant would. McCarty et al. (1999) described a developmental model of planning based on the elevated spoon task. Initially a feedback-based strategy is used in young infants. Gripping errors are not corrected until after the spoon has been transported to the mouth. As planning develops in infants, a partially planned strategy is used in which errors are perceived and corrected during the transport action. Finally, actions are fully planned before the spoon is gripped, and errors are avoided.
For monkeys, the consequence of losing the food coupled with other feedback may have shaped the rapid progression from a feedback-based to a fully planned strategy. During the first session, every monkey put his or her hand in the food, either by grabbing the bowl of the spoon or finger dipping. Through feedback, monkeys learned something about the affordances of the spoon. Monkeys showed partial planning on trials where a gripping error was made. For example, monkeys rotated the spoon after initially selecting an inefficient ulnar grip. Although the food was often lost in this maneuvering, the monkey correctly prevented the handle of the spoon from going into to the mouth. At criterion, monkeys’ consistent use of an efficient grip demonstrated full planning. Overall, monkeys resembled older infants that at first adjusted mistakes before eating the food and ultimately were able to efficiently transport the spoon in a single movement when it was placed in different orientations by either alternating hands or maintaining hand preference and adjusting their body.
EXPERIMENT 2: DOUBLE BOWL CONDITION
In Experiment 1, monkeys demonstrated that they could solve the elevated spoon problem at first by using feedback and then by implementing fully planned strategies. The strategies varied on difficult trials, with some monkeys switching to their non-preferred hand and other monkeys reorienting their body to the apparatus to facilitate continued use of the preferred hand. On easy trials, all monkeys used their preferred hand. Experiment 2 was designed to explore what spoon feature monkeys were attending to that signaled how the spoon should be transported efficiently. We created a novel spoon that resembled a barbell with a handle connecting two bowl ends of equal size, and only baited one of the bowls per trial (Figure 1B). If monkeys had learned to distinguish between the bowl and the handle of the regular spoon and were using the direction of the handle to guide grip selection, we predicted monkeys would plan poorly with the double bowl spoon when the ends of the spoon were identical. However if monkeys were using the direction of the spoon bowl for movement choice and had represented the location of the food, we predicted that monkeys would show good planning (i.e., select efficient grips) for transporting the double bowl spoon.
METHOD
Subjects
The subjects that demonstrated mastery of efficient spoon transport in Experiment 1 were tested in Experiment 2 approximately 1 year later (N = 6). Monkeys continued to participate in other cognitive and motor studies during the time between experiments; however, they were not exposed to additional tool-using tasks during this year interval.
Modified Elevated Spoon Task
In this version of the elevated spoon task, monkeys were required to lift a modified metal spoon constructed with two bowl ends from the wooden platform described in Experiment 1 (Figure 1B). The double bowl spoon was equivalent in length to the single bowl spoon, and the dimensions of each spoon bowl matched that of the spoons used in Experiment 1. The portion of the spoon connecting the two bowls was approximately 5 cm in length. Only one bowl of the spoon was baited with either applesauce or yogurt on any given trial, and the testing procedure was identical to that of previous testing with the single bowl spoon. As in Experiment 1, there were both easy and difficult trials. An easy trial consisted of baiting the spoon bowl contralateral to the monkey’s preferred hand and a difficult trial consisted of baiting the spoon bowl ipsilateral to the monkey’s preferred hand. The location of the food (left or right spoon bowl) was randomized across trials, and each bowl was baited the same number of times, creating an equal number of easy and difficult trials. Monkeys received a single session of 6 trials and all trials were videotaped for later analysis.
Analysis
Spoon trials from Experiment 2 were analyzed from videotape in a similar manner to data collected in Experiment 1. The primary observer noted the location of the food, whether the trial was easy or difficult for the individual tested, and the monkey’s initial grip on the spoon. Spoon grips were scored as radial (hand on spoon handle or unbaited spoon bowl with palm facing down and thumb towards the baited spoon bowl), underhand (hand on spoon handle with palm facing up), ulnar (hand on spoon handle or unbaited spoon bowl with palm facing down and thumb towards unbaited spoon bowl), bowl (any portion of hand gripping baited spoon bowl), or no grip, meaning monkeys attempted to consume the food by licking or finger dipping. To clarify, a bowl grip indicated that the monkey put some portion of its hand in the food. A grip to the non-baited bowl was not considered an error unless the thumb pointed away from the food (i.e., ulnar grip). Because monkeys were given a single session, movement time was not scored. Paired samples t-tests were used to compare the percentage of efficient grip use (radial or underhand) between the single bowl condition at criterion (Experiment 1) and the double bowl condition (Experiment 2) for easy and difficult trials. Alpha was 0.05 for all tests.
RESULTS AND DISCUSSION
Like the single bowl spoon, monkeys raised the double bowl spoon to the mouth and ate from the baited bowl. Monkeys picked up the double bowl spoon on 100% of the total trials (36/36) and use of an efficient grip varied by trial type (Table 2). A grip was considered efficient if it allowed the spoon to be transported to the mouth in a single movement without any adjustment or corrections. On easy trials, monkeys used an efficient grip 83% of the time. Two-thirds of the efficient grips were radial grips, while one-third were underhand grips. On difficult trials, monkeys used an efficient grip 67% of the time. All of the efficient grips seen on difficult trials were radial grips. Paired samples t-tests revealed that monkeys did not differ in their percentage of efficient grip use between the double bowl spoon and the single bowl spoon at criterion (Experiment 1) on easy trials, t(5)=1.460, P>0.05 (SEM=11.418), or on difficult trials, t(5)=1.740, P>0.05 (SEM=15.901).
Monkeys did not simply generalize their previously developed strategy for gripping and transporting the spoon to the double bowl condition. Monkeys in the alternator group continued to use both the preferred and non-preferred hands on different trials. However, two of the three monkeys in this group (Thelma and Violet) made at least one efficient underhand grip on easy trials in the double bowl condition. These monkeys had not used an underhand grip during Experiment 1. For monkeys in the maintainer group, a mix of preferred hand and non-preferred hand use was observed on difficult trials. Two monkeys (Kayla and Zoey) used their non-preferred hand once each on difficult trials with the double bowl spoon, resembling monkeys in the alternator group. These results indicate that grip choice may have varied as a function of the different properties of the single and double bowl spoons.
Although alternators and maintainers largely continued to transport the spoon efficiently, the types of errors each group made differed (Table 3). Maintainers made errors on easy and difficult trials in the double bowl condition. Moreover, these errors included both the inefficient ulnar and bowl grips. In contrast, monkeys that had learned to alternate hands based on the spoon’s orientation did not make any mistakes on easy trials in the double bowl condition. On difficult trials, these monkeys only made bowl grip errors. One possibility is that maintainers had been attending to both the bowl and the handle end of the spoon, and were more disrupted than alternators by the loss of the information that the handle had provided. These differences in error patterns deserve consideration in future studies to advance our understanding of the significance of different types of mistakes in the development of planning as well as the variability seen in motor strategies.
Table 3.
Error type by trial type for each transport strategy in the double bowl condition
| Alternators | Maintainers | |||
|---|---|---|---|---|
| Bowl | Ulnar | Bowl | Ulnar | |
| Trial Type | ||||
| Easy | 0 | 0 | 2 | 1 |
| Difficult | 4 | 0 | 0 | 2 |
GENERAL DISCUSSION
Rhesus monkeys engage in a great deal of oral object manipulation and could have easily chosen to lick from the spoon rather than picking it up with the hands. Notably however, monkeys chose to use the spoon as a tool and quickly developed strategies for efficiently transporting the spoon to the mouth in a single movement in Experiment 1. These strategies varied from using the preferred hand on easy trials and the non-preferred hand on difficult trials (alternator group) to using the preferred hand on all trials and varying body position (maintainer group). The degree of hand preference lateralization did not appear to influence the type of strategy monkeys adopted. The monkey with the strongest hand preference overall (Thelma, HI = −1.00) was the first to demonstrate mastery of efficient spoon transport, and did so by breaking hand preference and using her non-preferred hand on difficult trials. Similarly, a monkey with a weaker hand preference compared to the others (Kayla, HI = 0.32) consistently used her preferred hand on both easy and difficult trials. The direction of hand preference lateralization may have been related to grip strategy. The subset of monkeys in the maintainer group consisted only of right-preferent individuals, whereas monkeys in the alternator group were both left- and right-preferent. These results suggest that right-preferent monkeys may be less likely to use their non-preferred hand compared to left-preferent monkeys, a pattern that has also been reported for reaching studies in human adults (e.g., Bryden, Pryde & Roy, 2000).
The monkeys in the maintainer group may have been following the higher order motor planning constraint of end-state comfort that has been documented in adult human studies (Rosenbaum, Marchak, Barnes, Vaughan, Slotta & Jorgensen, 1990; Rosenbaum & Jorgensen, 1992). Briefly, adults were asked to pick up an elevated horizontal bar and move it to various vertical positions using their preferred hand. Participants gripped the bar differently depending on the rotational movement they had been asked to make, alternating between an overhand grip and an underhand grip. The underhand grip was initially awkward, but allowed the hand to be in a comfortable position at the end of the required movement. By selecting for end-state comfort, adults revealed they had planned the hand’s trajectory before contacting the bar. In a related set of studies with cotton-top tamarins and various lemur species, subjects selected an underhand grip for rotating an inverted cup, suggesting that end-state comfort plays a role in planning in both human and nonhuman primates (Chapman et al., 2010; Weiss et al., 2007).
For the rhesus monkeys in the present study, the combination of the radial grip and the preferred hand on difficult trials appeared to result in a hand position that was initially awkward, but allowed the hand to rest comfortably when the spoon was raised to the mouth. Monkeys that consistently used the preferred hand may have anticipated their own hand’s trajectory much like the adults on the bar task and the nonhuman primates on the cup task and compensated for awkwardness by angling their body rather than approaching the task head-on. By showing end-state comfort, individuals are selecting an initial grip that anticipates future utility. Although monkeys that were guided by end-state comfort may have perceived the spoon task differently than monkeys that used the strategy of alternating hands, both groups developed efficient grip solutions.
An outstanding question is whether monkeys in the maintainer group were more sophisticated planners than monkeys in the alternator group. Monkeys that used their preferred hand on difficult trials selected a grip that was not only efficient, but also appeared to take into account the ending comfort of the movement. However, an alternative interpretation is that these monkeys were cognitively less flexible than the other monkeys and showed an over-reliance on their preferred hand. In Experiment 2, these monkeys showed a different error pattern when mistakes were made transporting the double bowl spoon compared with monkeys in the alternator group. Maintainers made mistakes on easy trials and difficult trials, whereas alternators only made mistakes on difficult trials. Furthermore, maintainers made ulnar grip errors but alternators did not. Nevertheless, both groups made bowl grip errors, indicating a propensity to reach directly to the goal. Bowl grip errors were also common in early sessions with the single bowl spoon. Inhibitory control may therefore play an important role in the elevated spoon task in suppressing the natural tendency to reach directly to the food, or to use the preferred hand.
Another paradigm that has been used to measure inhibitory control in rhesus monkeys and human infants is the object retrieval task (Diamond, 1990). In this task, an object is placed in a clear box. The front of the box is closed such that the individual cannot reach forward to the object, but rather must reach around to an opening on one of its sides. Rhesus monkeys perform well on this task at 3 to 4 months-of-age, whereas human infants aren’t successful until 11 to 12 months-of-age (Diamond, 1990). Perhaps 9-month-old infants do poorly on the elevated spoon task because they cannot yet inhibit reaching directly to a goal. It would be valuable to test infant rhesus monkeys on the elevated spoon task and compare their errors and strategies to that of human infants as well as to the adult monkey data presented here.
The elevated spoon paradigm offered an opportunity to observe the development of motor planning in a nonhuman primate species using adult rhesus monkeys that were unfamiliar with spoons. The monkeys’ knowledge of the affordances of spoons was further examined with a novel double bowl spoon. Our findings indicate that monkeys attended to the baited bowl of the spoon and did not simply generalize a learned strategy from their previous experiences with the single bowl spoon. We acknowledge that the double bowl spoon task was more difficult than the original spoon task, and a distinct spoon handle may have helped monkeys perform more accurately, particularly those in the maintainer group who were slightly disrupted on both easy and difficult trials. Many questions still remain regarding what monkeys understand about the affordances of each spoon and which elements of the task are the most challenging, particularly with regards to the development of different efficient transport strategies. Future work should also be directed at examining how monkeys perceive affordances of other tools in contexts that require planning, especially during infancy, and how different objects and object relations may impact movement selection.
The dynamic nature of transporting a spoon loaded with food required monkeys to continually monitor their movements and to select efficient grips in advance to avoid spilling. Other tool using tasks such as using a rake to retrieve a treat out of reach are also dynamic in nature, but previous experience on a rake task did not affect performance on the elevated spoon task, suggesting that monkeys learn something about the affordances of each tool, rather than possess a general tool-using ability. The spoon paradigm could be extended in monkeys to understanding how efficient spoon loading is sequenced with efficient spoon transport, where monkeys must first monitor getting the food onto the spoon and then monitor getting the food to the mouth. The development of efficient nutcracking, also a dynamic planning sequence, has previously been explored in capuchin monkeys using this perception-action framework (de Resende, Ottoni & Fragaszy, 2008).
An essential take-home message from this work is that the ability to use tools and plan movements is a process, not a dichotomy, in both human infants and nonhuman primates. This underlying process is deeply rooted in perception and action relationships, and a comparative approach can be a valuable tool for uncovering the fundamental elements of these abilities, despite species differences in the level of ultimate sophistication. By employing a perception-action framework from multiple perspectives including ontogeny within different species and phylogeny across species, we will further our knowledge of the origins of motor planning and tool-using behavior.
Acknowledgments
We would like to thank Samantha Babcock, Maurine Braun, Amanda Hamel, and Catherine Martin for their assistance in collecting or scoring data, and Joseph Bergman, Gary Cormier, and Tony Scarnici for their creativity and effort in constructing the testing materials. We are also very grateful to Gina Fortunato for the drawings used in Figure 3 and to Dan Weiss and an anonymous reviewer for helpful comments on previous versions of the manuscript.
References
- Bryden PJ, Pryde KM, Roy EA. A performance measure of the degree of hand preference. Brain and Cognition. 2000;44:402–414. doi: 10.1006/brcg.1999.1201. [DOI] [PubMed] [Google Scholar]
- Chapman KM, Weiss DJ, Rosenbaum DA. Evolutionary roots of motor planning: The end-state comfort effect in lemurs. Journal of Comparative Psychology. 2010;124:229–232. doi: 10.1037/a0018025. [DOI] [PubMed] [Google Scholar]
- Claxton LJ, Keen R, McCarty ME. Evidence of motor planning in infant reaching behavior. Psychological Science. 2003;14:354–356. doi: 10.1111/1467-9280.24421. [DOI] [PubMed] [Google Scholar]
- Claxton LJ, McCarty ME, Keen R. Self-directed action affects planning in tool-use tasks with toddlers. Infant Behavior and Development. 2009;32:230–233. doi: 10.1016/j.infbeh.2008.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Connolly K, Dalgleish M. The emergence of a tool-using skill in infancy. Developmental Psychology. 1989;25:894–912. [Google Scholar]
- Connolly K, Dalgleish M. Individual patterns of tool use by infants. In: Kalverboer AF, Hopkins B, Geuze R, editors. Motor development in early and later childhood: Longitudinal approaches. Cambridge: Cambridge University Press; 1993. pp. 174–204. [Google Scholar]
- de Resende BD, Ottoni EB, Fragaszy DM. Ontogeny of manipulative behavior and nut-cracking in young tufted capuchin monkeys (Cebus apella): A Perception-action perspective. Developmental Science. 2008;11:828–840. doi: 10.1111/j.1467-7687.2008.00731.x. [DOI] [PubMed] [Google Scholar]
- Diamond A. Developmental time course in human infants and infant monkeys, and the neural basis of inhibitory control in reaching. Annals of the New York Academy of Sciences. 1990;608:637–736. doi: 10.1111/j.1749-6632.1990.tb48913.x. [DOI] [PubMed] [Google Scholar]
- Gumert MD, Kluck M, Malaivijitnond S. The physical characteristics and usage patterns of stone axe and pounding hammers used by long-tailed macaques in the Andaman Sea region of Thailand. American Journal of Primatology. 2009;71:594–608. doi: 10.1002/ajp.20694. [DOI] [PubMed] [Google Scholar]
- Kellogg WN, Kellogg LA. The ape and the child: A study of environmental influence on early behavior. New York: McGraw-Hill; 1933. [Google Scholar]
- Lemon J, Bolker B, Oom S, Klein E, Rowlingson B, Wickham H, Tyagi A, Eterradossi O, Grothendieck G, Toews M, Kane J, Cheetham M, Turner R, Witthoft C, Stander J, Petzoldt T, Duursma R, Biancotto E, Levy O. plotrix: Various plotting functions. R package version 2.6-4. 2009 http://CRAN.R-project.org/package=plotrix.
- Lockman JL. A perception-action perspective on tool use development. Child Development. 2000;71:137–144. doi: 10.1111/1467-8624.00127. [DOI] [PubMed] [Google Scholar]
- McCarty ME, Clifton RK, Collard RR. Problem solving in infancy: The emergence of an action plan. Developmental Psychology. 1999;35:1091–1101. doi: 10.1037//0012-1649.35.4.1091. [DOI] [PubMed] [Google Scholar]
- McCarty ME, Clifton RK, Collard RR. The beginnings of tool use by infants and toddlers. Infancy. 2001;2:233–256. [Google Scholar]
- McCarty ME, Keen R. Facilitating problem-solving performance among 9-and 12-month-old infants. Journal of Cognition and Development. 2005;6:209–228. [Google Scholar]
- Metevier CM. Dissertation Abstracts International. 4. B67. University of Massachusetts Amherst; 2006. Tool-using in rhesus monkeys and 36-month-old children: Acquisition, comprehension, and individual differences; p. 2263. [Google Scholar]
- R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing; Vienna, Austria: 2009. URL http://www.R-project.org. [Google Scholar]
- Rosenbaum DA. The Cinderella of psychology: The neglect of motor control in the science of mental life and behavior. American Psychologist. 2005;60:308–317. doi: 10.1037/0003-066X.60.4.308. [DOI] [PubMed] [Google Scholar]
- Rosenbaum DA, Jorgensen MJ. Planning macroscopic aspects of manual control. Human Movement Science. 1992;11:61–69. [Google Scholar]
- Rosenbaum DA, Marchak F, Barnes HJ, Vaughan J, Slotta JD, Jorgensen MJ. Constraints for action selection: Overhand versus underhand grips. In: Jeannerod M, editor. Attention and performance XIII: Motor representation and control. Hillsdale, NJ: Erlbaum; 1990. pp. 321–342. [Google Scholar]
- Sarkar D. lattice: Lattice Graphics. R package version 0.17-25. 2009 http://CRAN.R-project.org/package=lattice.
- Shannon C, Champoux M, Suomi SJ. Rearing condition and plasma cortisol in rhesus monkey infants. American Journal of Primatology. 1998;46:311–321. doi: 10.1002/(SICI)1098-2345(1998)46:4<311::AID-AJP3>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- Visalberghi E, Fragaszy D. What is challenging about tool use? The capuchin’s perspective. In: Wasserman EA, Zentall TR, editors. Comparative Cognition: Experimental Explorations of Animal Intelligence. New York: Oxford University Press; 2006. pp. 529–552. [Google Scholar]
- Weiss DJ, Wark JD, Rosenbaum DA. Monkey see, monkey plan, monkey do. Psychological Science. 2007;18:1063–1068. doi: 10.1111/j.1467-9280.2007.02026.x. [DOI] [PubMed] [Google Scholar]